IMMUNREGULÁCIÓ KÍSÉRLETES ENCEPHALOMYELITISBEN: AZ NKT SEJTEK SZEREPE
|
|
|
- László Balázs
- 6 évvel ezelőtt
- Látták:
Átírás
1 IMMUNREGULÁCIÓ KÍSÉRLETES ENCEPHALOMYELITISBEN: AZ NKT SEJTEK SZEREPE Egyetemi doktori (Ph.D.) értekezés DR. PÁL ENDRE PROGRAM-, ALPROGRAM- és TÉMAVEZETŐ: DR. SZEKERES-BARTHÓ JÚLIA Egyetemi tanár ORVOSI MIKROBIOLÓGIAI ÉS IMMUNITÁSTANI INTÉZET, PTE, PÉCS 2001
2 Tartalomjegyzék Rövidítések 3 1. BEVEZETÉS 4 2. MÓDSZEREK EREDMÉNYEK MEGBESZÉLÉS ÖSSZEFOGLALÁS IRODALOM ÁBRÁK 56 Köszönetnyilvánítás 69 Függelék 70 Az értekezéshez kapcsolódó közlemények Cikkek és előadások jegyzéke
3 Rövidítések APC: Antigen presenting cell APL: Altered peptide ligand CP: krónikus progresszív CTLA-4: Cytotoxic T lymphocyte antigen 4 DN: Double negative (kettős negatív sejtek) EAE: Kísérletes encephalomyelitis (experimental autoimmune encephalomyelitis) EDSS: Expanded disability status scale GC: α-galactosylceramide GVHD: Graft versus host disease IFN-γ: Interferon gamma IL: Interleukin KO: Knockout MHC: Major hisztokompatibilitási komplex NKT: Natural Killer T sejt OGP: Oligoclonalis gammopathia RR: relapszus-remisszió SM: Sclerosis multiplex TCR: T sejt receptor TGFβ: Transforming growth factor beta Th: T helper sejt TME(V): Theiler murin encephalomyelitis (vírus)
4 1. Bevezetés 1.1. A sclerosis multiplex kóreredete A sclerosis multiplex (SM) a központi idegrendszer egyik leggyakoribb szerzett megbetegedése (1,2,3,4). Klinikailag 2 fő típus különíthető el. 1. A betegek 80%-át érintő, relapszusokkal-remissziókkal (RR) jellemzett forma, mely általában a 3. évtizedben kezdődik, nőkben 2-3x gyakoribb (1). A betegség korai szakaszában az átmeneti tüneteket (relapszus) teljes remisszió követheti, azonban a későbbiekben egyre több maradandó tünet jelenik meg és a betegség egy szekunder progresszív fázisba megy át. 2. Kevésbé gyakori a kezdettől fogva folyamatos progressziót mutató forma (CP), melyben remissziók nem ismerhetők fel (5). A betegség a központi idegrendszer fehérállományának többgócú demyelinizációjával jellemezhető. Többféle skálát használnak a klinikai tünetek leírására. A legelterjedtebb, és elfogadottabb a Kurtzke által leírt EDSS (expanded disability status scale), mely a különböző pályarendszerek érintettségét egy pozitív skálán osztályozza (6). Az SM biztos diagnózisa patológiai, azonban klinikai szinten elkülönítenek lehetséges, valószínű és határozott formát a klinikai és paraklinikai gócok száma és a tünetek dinamikája alapján (7). Az SM diagnosztikájának alapvető lépései: a klinikai tünetek megállapítása, követése, elektrofiziológiai vizsgálatok (kiváltott válaszok) képalkotó vizsgálatok (MRI), liquorvizsgálatok (intratekális immunglobulin szintézis kimutatása).
5 Az SM pontos patomechanizmusa nem ismert. A jelenleg elfogadott elmélet szerint a betegséget az élet korai szakaszában elszenvedett (vírus) infekció indítja el. Ezt követően alakul ki elsősorban celluláris, másodsorban humorális immunválasz autoimmun reakció a központi idegrendszer myelin antigénjeivel szemben (8). A környezeti hatások mellett genetikai tényezők, elsősorban bizonyos MHC-II allélek asszociációja is ismertek (9,10). A patológiai folyamat alapvető jellemzője a myelinhüvely destrukciója. Az axonok a betegség kezdetén rendszerint megkíméltek, a tünetek előrehaladásával azonban egyre inkább érintetteké válnak (11). A legújabb közlemények szerint az axonális károsodás nemcsak a betegség késői szakaszára jellemző, de változó mértékben már a korai stádiumban is kimutatható (12). Viszonylag kevés ismerettel rendelkezünk a myelin újraképződéséről, de bizonyították, hogy a kifejlődött idegrendszer is rendelkezik regenerációs (remyelinizációs) kapacitással (13). 1.2 A sclerosis multiplex állatmodelljei A spontán kialakuló SM-hez hasonló, állatokban előforduló betegséget nem ismerünk. Kísérleti állatokban alapvetően kétféle immun-mediált központi idegrendszeri demyelinizációs betegség idézhető elő: A, Theiler murine encephalomyelitis (TME), B, Kísérletes autoimmun encephalomyelitis (EAE) (14). A TME neuro-patogén vírusnak direkt a központi idegrendszerbe juttatásával idézhető elő. A vírusinfekció következtében a myelinhüvely károsodik, belőle myelin antigének válnak szabaddá, amelyek másodlagos (auto) immunreakciót eredményeznek (15).
6 Az EAE alapja a különböző egér-, patkány, vagy tengerimalac, stb. törzsek immunizációja myelin antigénekkel (aktív EAE), vagy encephalitogén T sejtek átvitele fogékony törzsekbe (passzív EAE) (16,17) A kísérletes encephalomyelitis 1930-ban sikerült elsőként Rivers és mtsainak kísérletes encephalomyelitist előidézni majmokon (18). Kezdetben agy és/vagy gerincvelő homogenizátummal történt többszörös immunizáció. Később azonosították a myelin komponenseit, tisztították, majd szintetikusan előállították a fehérjéket. A központi idegrendszer myelinjének fő alkotóelemei a myelin bázikus protein (MBP), proteolipid protein (PLP). A kis mennyiségben előforduló myelin oligodendrocita glikoprotein (MOG) és myelin asszociált glikoprotein (MAG) azonban szintén alkalmas betegség kiváltására (19,20). A megfelelő immunválasz eléréséhez a fenti proteineket, illetve azok egy rövid fragmentumát Freund adjuvánssal együtt juttatjuk a szervezetbe (21,22). A különböző törzsek eltérő fogékonyságot mutatnak az EAE iránt. A kísérletek tanúsága szerint ennek magyarázata az eltérő genetikai háttér, elsősorban az MHC-II molekulák meghatározó szerepe emelhető ki (22). Azonosították az egyes proteinek azon fragmentumait (immunogén régió), mely az MHC és TCR molekulákkal lép kölcsönhatásba, valamint meghatározták, hogy egyes törzsekben mely peptiddel (encephalitogén peptid) lehet betegséget kiváltani (22,23,24,25,26).
7 Az EAE kritikája A klinikai tünetek hasonlatossága ellenére az EAE-t, mint az SM állatmodelljét kritikával illetik. Különböző immunizációs protokollok alkalmazásával egy törzsön belül is előidézhető eltérő klinikai kép, azaz monofázisos, remissziókkal-relapszusokkal zajló, valamint krónikus progresszív forma is (25,26). Az EAE nem spontán kezdetű, immunizáció, vagy encephalitogén sejtek átvitele szükséges hozzá (kivételt képeznek egyes MBP-specifikus TCR transzgenikus egerek, amelyekben a betegség spontán alakul ki). A patológiai folyamat során dominálhat myelin pusztulás (gerincvelő homogenizátummal történt immunizáció után), míg más esetekben perivaszkuláris kereksejtes infiltráció, a parenchima és a meninxek gyulladásos beszűrődése a vezető patológiai elváltozás (myelin peptidekkel történt immunizáláskor) (11,27,28) Az SM és EAE immunpatológiája A patomechanizmus alapvető lépése a myelin-peptid-reaktív T sejtek klonális szelekciója, expanziója, a központi idegrendszer inváziója és a lokális myelin károsodás ( bystander demyelinisatio ) (28,29). Habár a patogenezis fenti lépései több-kevesebb bizonyítást nyertek, nem ismert, hogy az autoreaktiv sejtek aktivációja hol következik be (8). Fontos kérdés, hogy milyen mechanizmusok állhatnak autoimmun reakciók hátterében. A lehetséges magyarázat: 1, molecular mimicry (=vírus epitóp és myelin epitóp keresztreagáló T-sejt klónok), 2, az autoreaktiv sejtek nem specifikus aktivációja vírus kódolt szuperantigének által, 3, a myelin destrukciója során szabaddá váló auto-epitópok okozta Tsejt aktiváció ( epitope spreading ), ez utóbbi lehet felelős az immunreakció fennmaradásáért, az ismételt relapszusok előidézéséért (8,30).
8 Immunreguláció EAE-ben A betegség kiváltásában alapvető szerepet játszanak Th1 fenotípusú CD4+ T sejtek: a klinikai tünetegyüttes előidézhető encephalitogén sejtek passzív átvitelével (31,32). A humorális immunreakció jelentősége valószínűleg másodlagos: a beteg állatok/emberek széruma önmagában nem alkalmas a betegség átvitelére (33), azonban (MOG ellenes) autoantitestek fontosak a lokális immunreakcióban és szövetkárosodásban, amint azok kimutathatók a demyelinizációs plakkokban, valamint a betegek szérumában is (34). A fogékony állattörzsekben csak bizonyos aminosav-szekvencia (domináns epitóp) képes immunreakciót kiváltani. Ennek molekuláris biológiai alapja az, hogy a peptid antigén meghatározott aminosav-csoportjai képesek az MHC- hez, és mások csak a TCR-hez kapcsolódni. A kapcsolat specifitásából következik, hogy 1. módosított peptidek (APL) ellenkező irányú, protektiv immunválaszt képesek előidézni (35), 2. homogén MHC-vel rendelkező beltenyésztett állattörzsekben a TCR-használat korlátozott (36). Az autoimmunitás kutatása az utóbbi években új fordulatot vett. Az autoreaktiv sejtek tanulmányozása mellett egyéb, regulációs funkcióval rendelkező sejtpopulációk kerültek a vizsgálatok középpontjába, mint T sejtek, NK és NKT sejtek (26,37,38,39,40,41,42,43,44). A jelen munka is a regulációs sejtek szerepével, működésével foglalkozik. A betegség során 2 meghatározó pont van: 1: az immunválasz és betegség indukciója, 2: a klinikai tünetek remissziója. Ad 1: Az immunizációt követően az antigén-specifikus T sejtek Th1 irányú differenciálódása indul el. Ebben a szakaszban meghatározó jellegű a citokin-környezet: IL-12, IFN-γ jelenléte
9 a Th1 típusú, míg IL-4, IL-10 jelenléte Th2-irányú differenciálódásnak kedvez (39,45,46). Aktivált Th2 típusú sejtek átvitelével az EAE kialakulása gátolható (47,48). Az NK sejtek IFN-γ termelése révén a Th1 típusú differenciálódást segítik elő (49,50). Az NKT sejtek képesek mind Th1 (IFN-γ), Th2 (IL-4) és Th3 (TGFβ) citokinek termelésére, ezáltal szerepet játszhatnak az immunológiai folyamatok szabályozásában (50,51,52,53). Az NKT sejteknek EAE-ben játszott szerepe ezideig nem ismert. Nem tudjuk, hogy antigén stimulációt követően mikor, hogyan aktiválódnak és mely irányba befolyásolják az autoimmun folyamatot. Nem ismert szerepük a helyi (agyi) szövetkárosodásban sem. Ad 2. A klinikai tünetek csúcspontján többféle immunológiai változás történik, amely a remisszió elindításáért felelős. Az autoreaktiv T sejtek száma csökken apoptosis révén (54), valamint a sejtek fenotípusa is megváltozik, Th1 Th2 irányba (55,56). Az apoptosis előidézésében fontos szerepet játszik az autonóm idegrendszer (glukokortikoidok és noradrenalin apoptosist indukáló hatása révén) (57,58) és a regulátoros sejtek aktivitása (NK sejtek, valamint más, szuppresszor (CD8+, vagy CD25+, γδ) T sejtek okozta T sejt inaktiváció, pusztulás (38,40,41,59). Jelen munka célja az NKT sejtek szerepének tanulmányozása volt az EAE során.
10 2. Anyagok és Módszerek 2.1. Egerek Vad típusú C57BL/6 (B6) egereket a CLEA (Tokyo, Japan) laboratórium biztosította. Interleukin (IL)-4 -/- és interferon (IFN)-γ -/- egereket a Jackson Laboratory (Bar Harbor, ME, USA) szállította. Vα14-Jα281 (NKT) -/-, valamint Vα14-Jα281 (NKT) transzgenikus (tg) egereket Dr. Taniguchi és mtsai hozták létre és tenyésztették (60). Valamennyi állatot speciális, pathogen-free körülmények között tartottuk, csak 6-12 hetes nőstény egyedeket használtunk fel a kísérletekhez Reagensek A Pharmingen (San Diego, CA, USA)-től vásároltuk a következő monoklonális antitesteket: In vitro blokkolási kísérletekhez tisztított és azid-mentes anti- CD1d (1B1), CD3e (145-2C11), CD11a (LFA1, M17/4), CD28 (37.51), CD40 (HM40-3), CD54 (ICAM-1 (3E2) CD80 (B7.1, 1G10), CD86 (B7.2, GL1), CD152 (CTLA-4, 9H10); flow cytometriához tisztított CD16/32 (2.4G2), FITC jelölt anti-cd3e (145-2C11), CD40 (HM40-3), CD80 (B7.1, 1G10), CD86 (B7.2, GL1), mouse I-Ab (AF ), PE jelölt anti-cd1d (1B1), CD3e (1452C11), CD11c (HL3), CD19 (1D3), NK1.1 (PK136), valamint ELISA céljára tisztított és biotin jelölt anti-ifn-γ (XMG1.2), IL-4 (BVD6 és 24G2), IL-10 (SXC-1), IL-12 (p40/p70, C17.8), és rekombináns egér (rm) citokineket: IL-2, IL-4, IL-12, IFN-γ. A szérum citokineket Quantikine M kittel (R&D System) határoztuk meg. In vivo NK sejt delécióhoz anti asialo-gm1 (Wako, Richmond, VA), míg az NK és NKT sejtek együttes eltávolításához anti NK1.1 (PK-136, ATCC hibridoma) antitestet használtunk.
11 RPMI 1640 médiumot, Iscove s Modified Dulbeco s médiumot, dimethyl-sulfoxidot (DMSO), pertussis toxin (PT)-t, trypsint és DNA-se-t, L-glutamint, fetal calf serum-ot (FCS), PHK-26 lipofil membránfestéket és diethyl-aether-t a Sigma (St. Louis, MO, USA)-tól, 2merkaptoethanolt, penicillin-streptomycin oldatot, Freund adjuvánst és hővel inaktivált Mycobacterium tuberculosist (H37Ra) a Difco (Detroit, MI, USA) laboratóriumtól vásároltuk. Myelin oligodendrocyta glycoprotein (MOG) peptidet (MEVGWYRSPFSRVVHLYRNGK) a CHIRON Technologies (Clayton, Victoria, Australia) szintetizálta. A kísérletekhez 2 mg/ml törzsoldatot használtunk, ehhez a peptidet PBS-ben oldottuk fel. Αz α-galactosylceramide (GC) szintézise Morita és Motoki szerint történt (61,62). A törzsoldat 100 µg/ml-es volt, 10%-os dimethyl sulfoxide (DMSO)-ban feloldva. In vivo kísérleteinkhez az oldatot tovább hígítottuk PBS-sel és 100 µg/kg adagban alkalmaztuk intraperitonealisan. In vitro kultúrákhoz RPMI médiummal hígítottuk és 100 ng/ml koncentrációban használtuk Aktív EAE indukció (Zhang és mtsai szerint [26]) Az immunizációhoz a MOG peptid és Freund adjuváns 1:1 arányú emulzióját használtuk. Az emulziót valamennyi esetben frissen készítettünk és egerenként 200 µl-t injektáltunk mindkét oldalon a talppárnába, szubkután. Egerenként 100 µg peptidet adtunk be. A fenti beavatkozás után röviddel valamennyi egér 500 ng pertussis toxint (PT)-t kapott, amit 48 óra múlva megismételtük. A 7. napon ismételt immunizáció történt MOG és PT-adásával, a 9. napon ismét PT-t adtunk önállóan. Az ismertetett protokollal jól reprodukálható, enyhe lefolyású, monofázisos EAE volt előidézhető, mely az immunizációt követő napon kezdődött, néhány napos plató fázist követően spontán remissziót mutatott a napon. Az egerek 100%-ban megbetegedtek. Az egereket naponta vizsgáltuk a klinikai tünetek
12 súlyosságának megítélése végett. A klinikai tüneteket egy 6 pontos skálán osztályoztuk: 0: egészséges, 1: farok tónusának megszűnése, 2: járási nehezítettség, 3: hátsó lábak gyengesége, 4: hátsó lábak teljes bénulása, 5: hátsó és elülső lábak súlyos gyengesége, 6: halál az EAE következtében. IFN-γ és IL-4 knockout (KO) egerek esetében a fenti kezelés súlyos, általában halálos kimenetelű betegséget eredményezett, emiatt mind az antigén, mind a PT adagját felére csökkentettük. NKT KO egerekben a vad típusnál ismertetett protokollt alkalmaztuk Passzív EAE indukció MOG immunizációt követően 10 nappal a regionális nyirokcsomókat eltávolítottuk, és sejt szuszpenziót készítettünk. A nyirokcsomó sejteket 6 well lemezen (1.5x107/well) restimuláltuk RPMI médiumban, 25 µg/ml peptiddel, 37 oc-on, 5% CO2 jelenlétében 72 óráig. Többszörös mosást követően az élő sejteket megszámoltuk tripánkék festés után, majd egerenként 1x107 aktivált nyirokcsomó-sejtet juttatunk a recipiensekbe intravénásan. A sejt transzfert megelőzően a befogadó egereket röntgen készülékkel besugaraztuk (350 rad), utána minden esetben 500 ng PT-t adtunk. A speciális beavatkozások (pl. GC beadás) a sejt transzfert követően 1 órán belül történtek.
13 2.5. In vitro kultúra A sejtek szeparálása Az egereket éterrel elaltattuk, majd a lépet (egyes esetekben más nyirokszerveket, mint inguinalis és poplitealis nyirokcsomókat, thymust, májat, néhány esetben agyat) eltávolítottuk és a szerveket homogenizáltuk. Máj, vagy agyi mononukleáris sejtek gyűjtéséhez a sejt-homogenizátumot percoll-grádienssel választottuk szét (a lymphoid sejteket a 40/80%-os fázisok felszínéről gyűjtöttük össze). Csontvelői sejtek szeparálásához a végtagcsontokról a bőrt és izmokat eltávolítottuk, a csontokat kettévágtuk és a velőűr sejteket PBS öblítéssel távolítottuk el. A lépsejteket 96-well steril lemezeken tenyésztettük, 5x105/200 µl koncentrációban, RPMI 1640 médiumban, amelyet 5x10-5 M 2-mercaptoethanollal, 2mM L-glutaminnal, 100 U/µg/ml penicillin-streptomycin oldattal és 5% FCS-sel egészítettünk ki. A kultúrákat a kísérlettől függő ideig (általában óráig) inkubáltuk 5% CO2 tartalmú, párásított levegőjű inkubátorban 37 oc-on. Stimuláció esetén GC-t adtunk a kultúrához 100 ng/ml koncentrációban, a kontroll esetekben csak vivőanyag (PBS-DMSO) került a sejtekhez. Blokkolási kísérleteknél tisztított, monoklonális antitesteket adtunk a wellekhez a GC hozzáadásával egyidőben, 10 µg/ml koncentrációban Másodlagos sejtkultúrák, NKT sejtvonalak Egyes kísérleteknél izolált T (NKT) sejteket (5x104/100 µl/well) stimuláltunk GC-vel (100 ng/ml) antigén prezentáló sejtek (APC, 5x105/ 100 µl/well) jelenlétében. A T/NKT sejtek szeparálása a következő módon történt. A frissen preparált lépsejt szuszpenziót Petri
14 csészékben inkubáltuk 1 óráig, ennek során a monociták nagy része kitapadt. Ezt követően a mintát paramágneses bead-del jelölt I-Ab (B6 egértörzsre jellemző MHC II) antitesttel inkubáltuk (B sejtek jelölésére), majd MACS készülék (Miltényi Biotec) segítségével különítettük el a T/NKT sejteket negatív szeparálással. A minta tisztaságát valamennyi esetben FACS mérésekkel ellenőriztük, eszerint a minta kontaminációja < 10-15% volt egyéb (pl. NK) sejtek tekintetében. Az APC-hez friss lépsejt szuszpenziót készítettünk, majd a sejteket besugárzásnak tettük ki (4000 Rad). GC-specifikus sejtvonalakat állítottunk elő a limiting dilution technika alkalmazásával. Ennek során a primer lépsejt kultúrákat az előzőekben leirt módon készítettük el. A T/NKT sejtek életképességének fenntartása érdekében a médiumot 3-4 naponta cseréltük és IL-2 tartalmát pótoltuk (Con-A szupernatánssal: ezt patkány lépsejtek Con-A stimulációjával állítottuk elő és a sejtmentes szupernatánst fagyasztva tároltuk). Hosszútávú sejtvonalak esetében a médium FCS tartalma 5% volt. A sejtek antigén specificitásának megőrzése érdekében 10 naponként az IL-2 kimosása után GC-vel (100ng/ml) és APC-vel (irradiált lépsejtek, 5x105/100 µl/well) újra stimuláltuk az NKT sejteket (5x104/100 µl/well). Azokat a well-eket használtuk fel a továbbiakban, amelyekben a sejtek antigén-függő proliferációs képessége a legjobb volt. A említett módon legalább 1 hónapig fenntartott sejtek antigén specificitását, fenotípusát és funkcionális sajátosságait vizsgáltuk (citokin profil, receptor-blokkolási kísérletek, stb.) APC-transzfer kísérletek B6 egerekből készített lépsejteket besugaraztuk (4000 Rad), majd GC-vel (100 ng/ml) és/vagy a megfelelő monoklonális antitesttel (10 µg/ml) inkubáltuk standard médiumban, termosztátban 4 óráig. Alapos mosás után a sejteket 4%-os paraformaldehydel fixáltuk 5 percig. Intenzív mosást követően a sejteket (1x107/egér) az 1 nappal korábban immunizált
15 egerekbe injektáltuk intravénásan. Az immunizációs protokoll és az egerek megfigyelése a korábbiakban leírtaknak megfelelően történt APC preparálás NKT sejtek stimulációjához, különböző szervekből Lép és májsejteket a fentiekben leirt módon szeparáltuk. Agysejtek preparálásához kezeletlen, felnőtt B6 egereket használtunk. A steril körülmények között eltávolított agyat az erek és meninxek eltávolítása után - ollóval apró darabokra vágtuk, majd trypsin (0.125%) és DNA-se (1 mg/ml) oldatban inkubáltuk 37 oc-on, percig. A sejtszuszpenziót szűrés, mosás után Iscove s médium felhasználásával, szövettenyésztő edényekben inkubáltuk. A mediumot naponta cseréltük, a kitapadt sejteket minimum 7 napos inkubáció után használtuk fel. A sejtek fenotípusát flow cytométerrel ellenőriztük. Dendritikus sejtek (DC) előállításához csontvelői, vagy lépsejteket használtunk. A sejtek izolálása után 2 óráig inkubáció következett Petri csészékben. A továbbiakban csak a kitapadt sejteket tenyésztettük. A dendritikus sejtek feldúsításához a mediumot rgm-csf-el (4ng/ml) és ril-4-gyel (20 ng/ml) egészítettük ki nap elteltével a sejtek fenotípusának flow cytometriás ellenőrzése után történtek a kísérletek Proliferációs teszt A mérésekhez 96-well, lapos (flat-bottomed) lemezeket használtunk. A sejtek proliferációs aktivitását 72 óra után határoztuk meg antigén jelenlétében és anélkül. Az utolsó 16 órára 1 µci {3H} thymidint adtunk a wellekhez, majd a sejteket harvester-rel szűrőpapírra szárítottuk. Scintillációs folyadék hozzáadását követően az aktivitást beta-1205 counterrel mértük (Pharmacia, Uppsala, Sweden). A stimulációs indexet az antigén stimulált és kontroll wellek hányadosaként számítottuk ki.
16 2.7. ELISA In vitro kísérleteinkben a citokinek meghatározásához a sejteket U-alakú well-ekben inkubáltuk, a mérésekhez a kultúrák felülúszóit használtuk. A stimuláció kezdetétől számított 48 óra múlva gyűjtöttük össze a felúszókat, a sejteket centrifugálással távolítottuk el. A sejtmentes szupernatánst azonnal felhasználtuk, vagy későbbi méréshez 80 oc-on tároltuk. Az ELISA a standard Sandwich elv alapján történt a Pharmingen ajánlása szerint és reagenseinek felhasználásával. In vivo kísérleteink során a retroorbitalis vénák punkciójával gyűjtöttünk vért az állatoktól, majd a szérumot centrifugálás után fagyasztva tároltuk. A szérum citokinek mérése ugyancsak Sandwich ELISA-val történt, R&D System ELISA kittek felhasználásával (Quantikine M). Néhány esetben az EAE különböző stádiumában a szérumból MOG specifikus IgG izotipusok meghatározása történt. Ehhez ELISA lemezeket (Maxisorp, Nunc, Rochester, NY) coatoltunk MOG-gal (10 µg/ml, Na2HPO4 pufferben). BSA (1%) blokkolást követően a vizsgált szérumok x-es hígításait (vagy PBS-t) a lemezeken inkubáltuk, majd biotinnal jelölt nyúl anti-egér IgM, IgG1 és IgG2a antitestekkel kezeltük 1 óráig (Southern Biotechnology, Birmingham, AL). Streptavidin-biotinnal történt inkubáció után a lemezeket o-phenylene-diaminnal (Sigma) hívtuk elő és a wellek abszorpcióját fotométerrel 470 nm-en mértük. Valamennyi esetben legalább 3 párhuzamos well-t használtunk mintánként. Az antitest titereket a dilúció/absorpció görbék alapján számítottuk ki Flow cytometria A mérésekhez a sejteket a perifériás nyirokszervekből preparáltuk, néhány kísérletben az agyból és májból különítettük el a fent leírtak szerint. Sejtkultúrából is végeztünk méréseket, ehhez a kultúrák sejtjeit többször alaposan mostuk. A fent említett fluorescens
17 markerekkel történt jelölés jégen, 20 perces inkubációval (az antitestek koncentrációja 0.5 µg/ 106 sejt/0.1 ml volt). A jelölés előtt a sejteket tisztított anti-cd16/32 antitesttel (1.0 µg/106 sejt/0.1 ml) kezeltük a nem-specifikus kötődés megelőzésére. Az inkubációhoz PBS-BSA (1%)-NaN3 (0.1%) puffert használtunk valamennyi esetben. Többszörös mosást követően a mérés propidium-jodid (10 µg/ml) hozzáadása után történt FACSort (Becton Dickinson) flow cytométerrel. Valamennyi esetben sejtet elemeztünk, az elpusztult sejteket (propidium-jodid felvétel) az analízisből kizártuk, a további értékeléshez a lymphocyta populációt használtuk fel. Az értékelés a Cellquest szoftver felhasználásával történt Citotoxikus aktivitás mérése flow cytometria segítségével (63) A kísérleti állatokból lépsejteket preparáltunk a fentiekben ismertetett módon, majd RPMI médiumban GC, Concanavalin-A, vagy vivőanyag (kontroll) hozzáadása után 72 óráig tenyésztettük. Ezt követően a sejteket többször mostuk és az élő sejtek számát meghatároztuk. A mérésekhez YAC-1 T limfóma sejteket használtunk (ATCC) targetként, melyeket folyamatos kulturában tartottunk a kísérletek idején. A mérés előtt többszöri mosás után a YAC-1 sejteket PHK-26 lipofil festékkel jelöltük, amely lehetővé tette a sejtek azonosítását a flow cytometria során (FL-2 detektor). A citotoxicitás teszteket 5 ml-esl Ualakú műanyagcsőben végeztük. Valamennyi csőben azonos számú YAC-1 target sejtet tettünk 250 µl volumenben. A csövekhez a hígítási értékeknek megfelelő számú effektor sejtet (aktivált és kontroll lépsejtek) adtunk szintén 250 µl-ben. A megfelelő sejtkölcsönhatások kialakulását 1 perces centrifugálással értük el, majd a sejt-keveréket 4 óráig inkubáltuk 37 oc-os, 5% CO2 atmoszférában. Az inkubáció után a sejteket propidium jodiddal jelöltük meg, az elpusztult alakok azonosíthatósága végett. Valamennyi mintánál legalább 2-3 párhuzamos csövet mértünk le és átlagoltunk. A háttér (YAC-1 sejtek spontán
18 pusztulása és a lépsejtek spontán pusztulásának) megítélése végett negatív kontrollként a lépsejteket és a YAC-1 sejteket külön-külön is inkubáltuk. Az elpusztult sejtek számának meghatározásakor a mért értékekből mindig levontuk a háttérértékeket. A YAC-1 sejtek és lépsejtek elkülönítését az FL-2 fluoreszcencia-beli eltérés tette lehetővé (lép negativ, YAC-1 poziziv). További méréstechnikai részleteket tekintve ugyanazon elveket alkalmaztuk, mint a hagyományos méréseknél. 2.9 Statisztika Az EAE klinikai paramétereinek összehasonlítását (kumulatív score, maximum score, kezdet, tartam, stb.) Mann-Whitney teszttel végeztük. A proliferációs, citokin és flow citometriás mérések eredményeinek összehasonlításánál kétmintás T próbát alkalmaztunk.
19 3. Eredmények 3.1 Aktív és passzív EAE indukció normális B6 egerekben A bevezető kísérletekben egy korábban már publikált EAE-modellt reprodukáltunk (26). Az immunizációt követő 3. héten, a napon kezdődtek a klinikai tünetek. Néhány napos progresszió után elérték a maximális, a kontrollok esetében 1-2-es score-t, majd ismét néhány napos plató-fázist fokozatos regresszió követte a teljes gyógyulásig, kb napig. A megfigyelési periódus (ált. 50 nap) során nem észleltünk relapszust (1. ábra). Ezzel szemben a passzív EAE indukciója általában sikertelen volt 1x107 sejtszám ellenére. Nagyobb sejtszámokat rutinszerűen nem alkalmaztunk az eredmények nagy variabilitása miatt ( x107 sejtszámnál az egerek kb. 25%-ánál alakult ki EAE, míg ennél magasabb sejtszám súlyos, néhány nap alatt letális betegséget idézett elő). 3.2 NKT sejtek számának és aktivitásának változása az EAE során Klinikai vizsgálataink bizonyították, hogy az NK-sejt aktivitás változó SM betegekben, azonban határozottan csökken a relapszusok során /I. közlemény/. Ugyancsak ismert az NKT sejtek (klónok) számbeli csökkenése (57). Ezen előzmények alapján vizsgáltuk a fenti sejtpopulációk szerepét EAE-ben. Az immunizációt követően különböző időpontokban (0, 5, 15, 30, és 45.-napon) az egereket feláldoztuk és a lépsejtek flow cytometriás mérését (NK 1.1/CD3 kettős jelöléssel) végeztük el, valamennyi alkalommal ugyanazon körülmények között. A lép teljes sejtszámának ismeretében az NK és NKT sejtek abszolút számát határoztuk meg az értékelő program %-os eredményeinek segítségével. Amint a 2. ábra mutatja, a remisszió során észleltük az NK és NKT sejtek számának mérsékelt emelkedését. A citotoxicitást szintén flow cytométerrel, az
20 ismertetett protokoll szerint mértük. Kifejezett individuális különbségek mellett nem észleltünk szignifikáns eltérést a különböző időpontokban. 3.3 EAE az NK és NKT sejtek eltávolítása után A következő lépésben indirekt módszert alkalmaztunk az NK és NKT sejtek funkciójának megítélésére. Az NK sejteket asialo-gm1 monoklonális antitest injekciójával, más esetekben az NK és NKT sejteket együtt távolítottuk el PK 136 (NK 1.1) antitest beadásával, 1 nappal az immunizáció előtt. (A kezelés hatékonyságát flow cytometriával ellenőriztük, a vér NK 1.1+ sejtek számának meghatározásával.) Az NKT sejtek szerepét knockout egereken vizsgáltuk (Invariábilis Vα14 lánc az NKT sejtek többségében /de nem minden NKT sejtben/ a TCR jellemzője. Az NKT knockout (KO), más néven Vα14-/- egerekben az invariáns lánc hiánya miatt az NKT sejtek többsége hiányzik, az NKT sejtszám lényegesen alacsonyabb (<a vad típus 50%-a). A 3. ábrán látható, hogy az NK sejtek eltávolítása súlyos klinikai tünetekkel járó, progresszív betegséget eredményezett (maximum score, vad típus: 1.6±0.3, NK deléció után: 4.3±0.6, p<0.01). A betegség kezdete változatlan volt, ugyanakkor krónikus tünetek alakultak ki (az egerek egy részénél a betegség letális volt). Az eredmények arra utaltak, hogy az NK sejtek a remisszióban játszhatnak fontos szerepet. Ezzel ellentétben az NKT KO egerekben a klinikai tünetek súlyossága nem különbözött a kontrollokétól, és a remisszió is normálisan zajlott. A legfontosabb különbség, hogy a betegség kezdete korábbra tolódott (vad típusban 17.3±0.3 nap, NKT KO egérben 6.8±1.1 nap, p<0.01). Az eredmények arra utalhatnak, hogy az NKT sejtek az immunválasz kezdeti stádiumában, az indukciós szakaszban játszhatnak szerepet (4. ábra). Hipotézisünket
21 megerősíti, hogy vad típusú, vagy NKT tg lépsejtek beadása NKT KO egerekbe (az immunizációval egyidejűleg) megszüntette a korai kezdetet (5. ábra). Ezen indirekt adatok alapján arra következtethetünk, hogy mind az NK, mind az NKT sejtek szuppresszív hatást fejtenek ki az autoimmun folyamatra, azonban a hatás ideje és mechanizmusa eltérő. 3.4 Galactosylceramide, egy specifikus NKT-sejt ligand jellemzői /2. Közlemény/ A további kísérletekben a regulátor sejtek (elsősorban NKT sejtek) aktív szuppresszív hatását szerettük volna bizonyítani. Ehhez a közelmúlt egy felfedezése volt segítségünkre. Japán kutatóknak tumor-gátló hatású vegyületet sikerült előállítaniuk a tengeri szivacsból. Jelenleg a vegyület teljes szerkezete ismert és szintetikusan is előállítható (61,62). Az αgalactosylceramid (GC) egy glikolipid. Kawano és mtsai kísérletei szerint a vegyület annak ellenére, hogy az emlősök szervezetében nem fordul elő és α helyzetű cukormolekulát tartalmaz, szelektíven képes aktiválni mind egér, mind humán NKT sejteket (64). Az általunk használt GC hatékonyságának bizonyítására in vitro és in vivo rendszerben vizsgáltuk az aktivált NKT sejteket In vitro hatás Normális lépsejtek GC stimulálása sejtkultúrában a lépsejtek egy részének proliferációját és citokinek termelését idézi elő. A kísérletek során valamennyi alkalommal nagy mennyiségű IFN-γ és IL-4 volt mérhető 48 órával a stimuláció kezdete után, míg IL-10 és IL-12 valamennyi esetben a mérési határ alatti volt.(6/a. ábra). (IL-10 és IL.12 termelését csak NKT sejtvonalak stimulációjával sikerült kimutatni, nem közölt adatok). Jól ismert, és az anti-
22 tumor hatásban fontos tulajdonsága a GC-nek, hogy az NKT sejtek cytotoxicitását a concanavalin-a-val csaknem azonos mértékben fokozza (6/b ábra). Megjegyzendő, hogy a GC hatás szempontjából fontos az NKT sejtek forrása: lép és vér NKT sejtek a fenti módon aktiválhatók, ezzel szemben thymus és máj NKT sejtek nem reagálnak GC stimulálásra (7. ábra). Ennek hátterében állhat az NKT sejtek eltérő fenotípusa, valamint az antigén prezentálás helyi sajátosságai. Utóbbit alátámasztandó lép NKT sejteket különböző szervekből preparált APC-vel és GC-vel aktiváltuk. A kísérlet tanúsága szerint lép és máj APC esetén, szingén dendritikus sejtek alkalmazásával szignifikáns proliferációt és citokin termelést tudtunk kimutatni. Thymus sejtek nem váltottak ki aktivációt. Ugyancsak nem találtunk aktivációt friss, vagy kultúrában tartott agysejtek alkalmazásával (mononukleáris sejtek normális és EAE agyból, vagy glia + neuron kultúrából) (8. ábra). Az ábrán mutatott proliferáció mellett valamennyi kísérletben mérhető mennyiségű IFN-γ és IL-4 termelést is megfigyeltünk. A citokinek termelése szoros korrelációt mutatott a proliferációval. A kísérlet előtt (és egyes esetekben után) az APC fenotípusát flow cytometriás mérésekkel ellenőriztük. Az antigén prezentáció szempontjából fontos megjegyezni, hogy a lép-, máj- thymus sejtek, dendritikus sejtek többsége CD1+ volt, míg agyszövet-kultúrákban sosem találtunk CD1 expessziót. A későbbiekben említendő B7.1 (CD80) és B7.2 (CD86) kostimulációs molekula szintén kimutatható volt az agysejtek kivételével In vivo hatás Intraperitoneális GC injekciót követően különböző időpontokban gyűjtöttünk vért (szérumot) a kezelt és kontroll egerekből. Amint a 9. ábra mutatja egyszeri injekciót követően a citokinek szekvenciális termelése mutatható ki. A legkorábban az IL-4 (max. 2h) termelődik,
23 majd IL-12 (max. 6h) és végül a legnagyobb mennyiségben és legtartósabban IFN-γ mutatható ki (2-48h, max. 12 h). Fontos megjegyezni, hogy NKT KO egerekben sem in vitro, sem in vivo citokin termelést (IFNγ <0.1 ng/ml, IL-4 <0.01 ng/ml), vagy proliferációt (stimulációs index <2) nem tapasztaltunk, ami bizonyítja, hogy valamennyi hatásért a Vα14 invariábilis NKT sejtek tehetők felelőssé Galactosylceramide hatása aktív EAE-re; normális és citokin knockout egerek Az előző kísérletek bizonyítják, hogy a GC sikeresen és szelektíven aktiválja B6 egér lép NKT sejtjeit. Ennek alapján megkíséreltük az NKT sejteket aktiválni az EAE során. Ehhez az immunizációt követően egyszeri, 100 µg/kg GC-t adtunk az egereknek ip. Amint a 10/A ábra mutatja, az említett kezelés kismértékben, nem szignifikánsan fokozta a maximális score-t, de a betegség kezdete és tartama nem változott. Különböző dózisokat ( µg/kg) alkalmazva nem találtunk eltérő klinikai hatást. Ugyancsak hatástalan volt a GC kezelés a 15. napon, a klinikai tünetek kezdete előtt. A következőkben többszöri, ismételt GC kezeléssel próbálkoztunk (0, 3, 6, 12. napon). A kezelt egerek klinikailag hasonló súlyosságú tüneteket mutattak, mint a kontrollok, azonban a betegség tartama számottevően rövidebb volt. Tekintettel arra, hogy az ismételt kezelés humorális immunválaszt is elindít, ami az eredmények értékelését nehezíti, további therápiás alternatívákat kerestünk. Az előző kísérletek arra utaltak, hogy az NKT sejtek szerepe elsősorban az immunválasz kezdetén döntő. Feltételeztük, hogy a normális egerekben azért nem tapasztalunk változást egyszeri kezelést követően, mert az NKT sejtek szimultán termelték az IFN-γ-t, és az IL-4-et, így a Th1/Th2 egyensúly nem változott meg (a GC indukált IFN-γ/IL-4 aránya normálisan, a vad típusú egerekben közötti volt az in
24 vitro stimulációs kísérletekben). Hipotézisünk igazolására olyan lépsejteket és egereket vizsgáltunk, amelyek nem voltak képesek IFN-γ (IFN-γ KO), vagy IL-4 termelésére (IL-4 KO). Amint a 11. ábra mutatja, az IFN-γ KO sejtek csak IL-4-et, az IL-4 KO sejtek csak IFN-γ t termelnek, míg az NKT KO sejtek egyáltalán nem mutatnak aktivitást a GC hatására. Várakozásunknak megfelelően az EAE IL-4 KO erekben súlyosabb lefolyású volt, míg az IFN-γ KO egerekben enyhébb tünetekkel járt (10/B-C ábra). (NKT KO egerekben GC nem befolyásolta a betegség lefolyását, az adatok nem kerültek bemutatásra). A kísérlet adatainak összegzését az 1. táblázat tartalmazza. A fentiek alátámasztják, hogy a GC indukálta IFN-γ potencirozza, az IL-4 gátolja az EAE-t. Kumulatív score Kezelés Vad típusú B6 Kontroll 17.6 ± ± ± 4.2 Vad típusú B6 GC 17.8 ± ± ± 1.7* IL-4 KO Kontroll 18.0 ± ± ± 5.1 IL-4 KO GC 18.2 ± ± ± 10.4** IFN-γ KO Kontroll 20.0 ± ± 0.29 IFN-γ KO GC Kezdet Maximum Score Egértörzs 20.3 ± ± ± 13.3 *** 57.7 ± 17.2*** 1. Táblázat GC kezelés hatása aktív EAE-re EAE B6 vad tipusú, IL-4 KO and IFN-γ KO egerekben MOG-immunizálást követően. A kezelt egerek 100 µg/kg GC adásában részesültek az immunizáció napján. A táblázat több kísérlet eredményének összefoglalása, csoportonként 12 egér átlagát mutatjuk. A szignifikáns eltéréseket az alábbiak szerint jelöltük: * (p<0.05), **(p<0.03), ***(p<0.01).
25 3.6. A GC-indukált NKT sejt-aktiváció megváltoztatása a kostimuláció befolyásolása révén Eredményeink bizonyították, hogy az NKT sejtek effektív immunregulációs képességgel rendelkeznek, mivel mind Th1 és Th2 típusú citokinek termelésére képesek. Feltettük a kérdést, milyen módon lehetne elérni az NKT sejtek polarizált aktiválódását, amellyel csak IFN-γ, vagy csak IL-4 termelését idéznénk elő. A hagyományos T sejtek irodalmát áttekintve az egyik legjobban ismert és tanulmányozott módszer a kostimulációs szignálok befolyásolása (65). A konvencionális T sejtek analógiájára kipróbáltunk számos blokkoló antitestet, milyen hatást gyakorolnak a GC-kiváltotta NKT aktiválódásra. A 2. táblázat mutatja néhány blokkoló antitest hatását a proliferációra és a citokin termelésre. Az eredmények alapján 2 jól ismert, a T sejtek kostimulációjában fontos szerepet játszó antitestet (anti-b7.1/b7.2 és anti CD40) választottunk ki a további kísérletekhez, mivel ezeknél tapasztaltunk a Th0 fenotípus Th1, illetve Th2 irányú eltolódását (az IFN-γ/IL-4 arány megváltozását). Megfigyelhető, hogy a B7.1 blokkolás nem befolyásolta a GC hatását és nem változtatta meg az NKT sejtek fenotípusát. Ezzel szemben a B7.2 blokkolás Th2 irányú eltolódást eredményezett, elsősorban az IFN-γ termelés gátlása révén. Meglepő módon a CD40-CD40L interakció blokkolása ellenkező eredményt hozott: az IL-4 termelés csökkenését és a fenotípus Th1 irányú eltolódását. Várakozásainknak megfelelően a CD1-TCR interakció gátlása jelentős mértékben gátolta mind az IFN-γ, mind IL-4 termelést. Hasonló eredményre vezetett az NK1.1 molekulák gátlása. Ez utóbbi szerepe nem ismert a GC kiváltotta NKT-sejt aktiválódásban. Eredményeink alapján az NK1.1 receptor és ligandjának kölcsönhatása az NKT sejtek stimulációját segíti elő. A 12. ábra mutatja be a kiválasztott antitestekkel végzett mérések eredményeit grafikus formában.
26 Blokkoló antitest Proliferáció (SI) IFN-γ (ng/ml) IL-4 (ng/ml) Nincs (háttér) - 0,2±0,05 0,2±0, Nincs (GC) 3.9±0.9 6,7±1.1 1,27±0, Th0 0.5±0.17 (-60.6%) Th2 0.32±0.08 (-95.2%) 0.42±0.14 (-66.9%) Th2 CD1 3.24±0.59 (-16.9%) 0.35±0.07 (-94.7%) IFN-γ/IL-4 Fenotípus arány NK ±0.65 (+12.8%) B ±0.83 (+15.8%) 5.8±1.1 (-13.4%) 0.95±0,17 (-25.2%) Th0 B ±0.66 (-58.9%) 1,7±0,7 (-74.6%) 0,57±0,17 (-55%) Th2 CD ±0.18 (-58.2%) 6.6±0.63 (-1.5%) 0.05±0,005 (-96%) Th1 2. táblázat Monoklonális antitestek hatása a GC-kiváltotta proliferációra és citokin termelésre Valmennyi fent jelzett antitestet a kultúra indításakor, 10 µg/ml koncentrációban alkalmaztunk. A proliferáció és citokinek mérése a módszerek részben leírtak szerint történt. Az értékek 3 mérés átlagát±sd mutatják, zárójelben tüntettük fel a az antitestek nélküli kontrollhoz (GC) viszonyított eltérést %-át) Az EAE befolyásolása ex vivo kezelt APC segítségével A következőkben arra kerestük a választ, hogyan alkalmazhatjuk fenti eredményeinket az EAE szabályozásában. Egyik lehetőség a fenti blokkoló antitestek beadása az immunizáció idején. Ezt a lehetőséget elvetettük, ugyanis ez nemcsak az NKT sejtek aktiválódását, de a konvencionális, a betegség előidézéséért felelős encephalitogén T sejtek aktiválódását, polarizálódását is befolyásolná (ellentétes irányba: pl. anti-b7.2 a konvencionális T sejteket Th1, az NKT sejteket Th2 irányba; [65]). Ezen korlátok miatt egy másik NKT aktiválási módszert dolgoztunk ki. Irradiált (és/vagy) fixált normális lép sejteket in vitro GC-vel és
27 blokkoló antitestekkel inkubáltuk együtt 4 óráig, majd többszörös mosást követően az így előkezelt sejteket, mint antigén prezentáló sejtek (APC) jutattuk a szervezetbe. Feltételezéseink szerint ez a sejt-preparátum csak az NKT sejteket aktiválja a beadás után (mivel csak azok képesek a GC-t felismerni). Eredményeinket a 13. ábra mutatja. Kontrollként az APC-t kezelés nélkül, csak antitest-kezelés, csak GC kezelés után, illetve GC+B7.1 kezelés után juttattuk immunizált egerekbe. Jól látható, hogy valamennyi említett a kontroll csoport esetében az EAE lefolyása, súlyossága nem változott. Ezzel szemben a GC+B7.2 kezelt csoportban csak az állatok fele betegedett meg, és a betegség rövidebb lefolyású volt. Mindezeket együttvéve az átlagos score (A) és a kumulatív score (B) is szignifikánsan csökkent az említett csoportban (3. táblázat). Ezzel ellentétben anti-cd40 kezelt APC beadása az EAE klinikai tüneteinek súlyosbodását idézte elő. Mindemellett a betegség kezdete 10 nappal eltolódott és nem volt remisszió a kezelt csoportban (14. ábra). Nem tudtunk hatást kimutatni NKT KO egerek APC és/vagy antitest kezelésével, mely indirekt módon bizonyítja az NKT sejtek kulcsszerepét (15. ábra, 3. táblázat). A Th egyensúly megváltozását az IgG izotípus titerének megváltozásával igyekeztünk alátámasztani. Az EAE 30 napján, a klinikai tünetek csúcspontján az egerek szérumából meghatároztuk a MOG-specifikus IgG1-et és IgG2a-t. Az eredmények azt mutatták, hogy a kontroll, kezeletlen egerekben igen alacsony titer volt mérhető. Anti-B7.2-APC kezelés hatására az IgG1 jelentős emelkedést mutatott, ami a humorális immunválasz Th2 irányú eltolódását jelezte. Anti-CD40 kezelés nem eredményezett szignifikáns eltérést az izotipustiterekben (16. ábra).
28 Egértörzs Kezelés B6 vad típus APC Kezdet (nap) Maximum Score Kumulatív score 17.0± ± ±3.1 APC+GC 18.4± ± ±3.34* APC+B ±0 1.3± ±3.4 APC+GC+B ± ±0.15 APC+GC+B ± ± ±1.15* APC 17.0± ± ±2.8 APC+CD ± ± ±7 APC+GC+CD ± ± ±8.5* NKT KO APC+GC 14.5± ± ±6 III. APC+B ± ± ±2.4 APC+GC+B ±0 2.0± ±1.4 I. II. 3.0±3.9*** 3. táblázat APC transzfer hatása aktív EAE klinikai tüneteire vad típusú és NKT KO egerekben. A táblázatban jelzett módon, a módszerek részben leírtak szerint előkezelt APC-t (106 sejt/egér) az immunizálást követő napon juttattuk a recipiensekbe (N>=5 az I kísérletnél, N=3 a II. és III. és valamennyi csoportban). A kumulatív score esetén tapasztalt szignifikáns eltéréseket az alábbiak szerint jelöltük: * (p<0.05), ** (p<0.01), *** (p<0.001). 3.8 A B7 kostimuláció funkcionális vizsgálata /III. Közlemény/ A fenti eredmények arra utaltak, hogy a B7.2 kostimuláció fontos szerepet játszhat az NKT sejtek aktiválódásában. A következőkben a B7.2 szint változását vizsgáltuk az EAE során, in vivo, majd in vitro kultúrákban különböző sejteken. A normális lépsejtek igen kis arányban expresszálják a B7 receptorokat (~1 %). Az EAE során nem mutatható ki számottevő változás (nem közölt adatok).
29 Jellegzetes kinetikát figyeltünk meg a GC in vivo beadása után. Már 6 óra után jól mérhető a B7.2-t expresszáló sejtek számának növekedése. Ez a tendencia követhető 72 órán keresztül is. Ezzel szemben nem mutatható ki változás a B7.1 esetében, illetve ugyancsak nem volt szignifikáns változás látható az NKT KO egerek GC stimulációjával (17 ábra). Ugyanezen jelenség figyelhető meg lépsejtek in vitro GC stimulációjakor (18. ábra). Fontos megjegyezni, hogy a B7.1 expresszió nem változott, mindössze 72 óra után volt kimutatható kis mértékü emelkedés. A B7.2 szelektív up-regulációja elsősorban a B sejtek, mint fő antigén prezentáló sejtek (APC) felszínén figyelhető meg (de más potenciális APC, mint T sejtek, dendritikus sejtek, mikroglia esetében is nő az expresszió; utóbbiak nem kerültek bemutatásra). Blokkoló antitestek segítségével kerestük a választ, hogy mely sejtközötti interakciók felelősek a B7.2 up-regulációért. Amint vártuk, a CD1-TCR kapcsolat meghatározó (anti-cd1d antitest szignifikánsan blokkolta az antigén-indukálta B7.2 expressziót), emellett citokinek jelenléte szükséges (pl. az IL-4 blokkolásával kivédhető a B7.2 változása és a jelenség nem alakul ki IL-4 KO lépsejtek GC stimulációjakor). Rekombináns egér citokineket alkalmazva in vitro kultúrákban az IL-4 meghatározó szerepét találtuk. IL-4 mind a B7.1 és B7.2 up-regulációját elősegítette, de IFN-γ egyedül nem változtatta meg a B7 expressziót. IFN-γ + IL-4 együttes adása sikeresen reprodukálta a GC hatását, azaz a B7.2 szelektív emelkedését a B7.1 változása nélkül (19. ábra). Eredményeink szerint az NKT sejtek szimultán IFN-γ és IL-4 termelése szükséges az optimális B7.2 expresszióhoz az APC felszínén GC-specifikus NKT sejtvonalak előállítása és funkcionális jellemzése /IV. Közlemény/ A módszerek részben leirtak szerint GC-specifikus sejtvonalakat állítottunk elő.
30 Az NKT sejtek életképességének fenntartása érdekében a médiumot IL-2-vel (Con-A szupernatans) egészítettük ki, A sejtek antigén specifitásának megőrzése érdekében naponként az IL-2 kimosása után GC-vel és APC-vel újra stimuláltuk az NKT sejteket. Az ismertetett módon legalább 1 hónapig fenntartott sejtek antigén specificitását, fenotípusát és funkcionális sajátosságait vizsgáltuk. 10 GC specifikus sejtvonalat sikerült előállítanunk, amelyek proliferációval és citokin termeléssel reagáltak. 6 sejtvonal részletes vizsgálatát sikerült elvégeznünk, ezeket az alábbiakban részletesen ismertetjük. A fenotípus tekintve valamennyi sejtvonal CD3+NK1.1+-nek mutatkozott flow cytometriás mérésekkel. Az NKT sejtek-hez hasonlóan mindkét sejfelszini receptor expressziója alacsonyabb volt, mint a konvencionális T sejtek CD3, illetve NK sejtek NK1.1 expressziója. 3 sejtvonal mutatott CD4-CD8- kettős negativitást (DN), másik három CD4-CD8+ volt. TCR expessziót tekintve a Vβ8 dominanciáját találtuk (20. ábra). GC stimuláció hatására valamennyi esetben határozott proliferáció és IFN-γ termelés volt kimutatható, de mindössze 1 sejtvonal termelt kimutatható mennyiségben IL-4-et. Valamennyi esetben a CD1 receptor blokkolása szignifikánsan csökkentette mind a proliferációt, mind a citokin termelést, alátámasztva a CD1-en keresztül történő GC prezentálást. A kostimulációs szignálok tekintetében a lépsejtekhez hasonló viselkedést észleltünk: B7.2 blokkolás szignifikánsan gátolta az IFN-γ termelést. Az eredményeket a 4. táblázatban foglaltuk össze. Két jellemző sejtvonalat szemléltet a 20. ábra.
31 Sejtvonal SI IFN-γ IL-4 α-cd1 α Β7.2 Fenotípus TCRVβ Gci CD8+ 8 GC CD4-CD8-8 GC CD8+ 8 GC CD4-CD8- GC /- CD4-CD8- GC CD táblázat A GC-specifikus NKT sejtvonalak jellemzői (SI: stimulációs index, α-cd1, α-b7.2: a citokin termelés/proliferáció >50%-os gátlása CD1 és B7.2blokkkoló antitestek hatására. Az 1. oszlop a sejtvonalak jelzéseit tartalmazza) Az in vitro kísérleteket követően megkíséreltük a sejtvonalak in vivo tanulmányozását. Az in vitro aktivált NKT sejtvonalakat immunizált egerekbe juttatuk az immunizációt követő napon. A várakozásokkal ellentétében egy kivétellel nem tapasztaltunk változást az EAE lefolyásában. Mindössze 1 sejtvonal (GC24) fokozta az EAE klinikai tüneteit. Megjegyzendő, hogy ennél találtuk a legmagasabb IFN-γ termelést in vitro körülmények között, ami magyarázhatja a hatást (21. ábra) GC hatása passzív EAE klinikai lefolyására EAE GC kezelt recipiensekben In vitro re-stimulált MOG specifikus nyirokcsomó sejteket (107/egér) juttatunk a megelőzőleg teljes test besugárzáson átesett recipiensekbe intravénásan. A kontroll, kezeletlen egerekben nem sikerült betegséget előidézni a fenti sejtszámmal. Az állatok egy csoportját
32 GC-vel kezeltünk ip. (100 µg/kg) a sejttranszferrel egyidejűleg. Ezen egerekben súlyos klinikai tünetekkel járó, monofázisos betegséget figyeltünk meg (22. ábra) EAE indukciója MOG+GC aktivált T sejtekkel Az egerek egy másik csoportját olyan nyirokcsomó sejtekkel kezeltük, amelyeket MOG-immunizált egerekből preparáltunk, majd a sejttranszfer előtt in vitro MOG35-55 (25 µg/ ml) és GC-vel (100 ng/ml) egyidejűleg re-stimuláltuk. Amint a 23. ábrán megfigyelhető, a fenti kezelés szintén súlyos, progressziv tüneteket eredményezett. In vitro kísérletek szerint utóbbi jelenség magyarázata, hogy GC MOG35-55 peptiddel együtt szignifikánsan fokozza a T sejtek proliferációját és IFN-γ termelését, ezáltal encephalitogén potenciálját (a csak MOG35 tel re-stimulált sejtekkel összehasonlítva, nem közölt adatok). 55
33 4. Megbeszélés 4.1. NKT sejtek szerepe EAE-ben, immunreguláció Az EAE Th1 típusú CD4+ T sejtek által előidézett autoimmun betegség (31,32). Az immunológiai folyamat és a klinikai tünetek lefolyása változatos, függ attól melyik törzsben, melyik immunogén peptiddel (22) és milyen körülmények között történik az immunizáció, illetve az encephalitogén sejtek beadása. Eddigi tudásunk szerint számos regulációs mechanizmus befolyásolja az autoimmun betegségek kialakulását és lefolyását, mint például az autoimmun sejtek fenotípusának változása (55,56) neuro-endokrin kölcsönhatások (57,58) és regulációs funkcióval rendelkező sejtpopulációk jelenléte. Utóbbiak közé tartoznak CD25+ T sejtek, CD8+ citotoxikus sejtek, anti-idiotipus hálózat sejtjei, γδ T sejtek, NK-sejtek, melyek szerepét már bizonyították EAE-ben (26,37,38,39,40,41,59). Az NKT sejtek sajátos sejtpopuláció, amelynek tagjai NK- és T sejt receptorokkal is rendelkeznek. Állatkísérletek adatai alapján mind az NK 1.1 receptor keresztkötésével, mind a CD3-TCR komplexen keresztül aktiválhatók. Az aktiváció citokin termelést és fokozott citotoxikus aktivitást eredményez (50,51,52,53,66). Ezen tulajdonságok alapján merült fel a lehetőség, hogy ezek a sejtek immunregulációs szerepet játszanak. A hipotézist alátámasztja, hogy a Vα14+ NK 1.1+ T sejtek szelektív pusztulása mutatható ki autoimmun állatmodellekben ( autoimmune prone mouse, non-obese diabetic mouse [NOD]; 38,66,67,68), illetve az EAE humán megfelelőjében, sclerosis multiplexben (69). Az NKT sejtek hiánya szerepet játszhat egyes egértörzsek fokozott EAE-iránti fogékonyságában (70). Ezzel szemben az NKT sejtek jelenléte szükséges a Concanavalin-A indukált hepatitis kialakulásához (71). Az NKT sejtek a tumor ellenes immunitásban (52,66,72,73,74), bizonyos kórokozók elleni védelemben (66,75,76), toleranciában (77) és a graft versus host betegségben (78) is szerepet játszanak.
34 Ezen adatok alapján vizsgáltuk az NKT sejtek szerepét C57BL/6 egértörzsben kiváltott EAE során. Az NK és NKT sejtek száma a remisszió során mutatott emelkedést, ami utalhat szerepükre a remisszió előidézésében, azonban a citotoxikus aktivitásban nem találtunk szignifikáns különbséget a betegség különböző stádiumaiban (1. és 6. ábra). A 3. ábrán bemutatott kísérlet megerősíti a korábban már közölt eredményeket: az NK sejtek eltávolítása után a klinikai tünetek súlyosbodnak, nem ritkán halállal végződő betegség alakul ki, és a túlélő egyedekben sem mutatható ki remisszió. Az NKT sejtek eltávolítása nem befolyásolta a betegség súlyosságát, ugyanakkor a betegség kezdete korábbra tolódott (4. ábra). Ezek az eredmények megerősítik, hogy az NKT sejtek protektiv szerepet játszanak EAE-ben, elsősorban az immunválasz indukciójában. A NOD modellben a Vα14 gén bejuttatása (NKT transgenikus egér) megakadályozta a diabetes kialakulását (79,80). A diabetestől eltérően az EAE NKT sejtek beadásával nem előzhető meg, azonban az NKT KO egerekben tapasztalt korai kezdet NKT sejtek beadásával megszűnik (5. ábra). Kísérleteink alapján az NK és NKT sejtek protektiv mechanizmusa eltérő, mivel előbbiek az effektor, utóbbiak elsősorban az indukciós fázist befolyásolják. Az NKT sejtek szerepének további tanulmányozásához olyan ligandot kerestünk, ami az NKT sejteket szelektíven aktiválja. A Kawano és Taniguchi munkacsoportja által a tengeri szivacsból izolált, majd mesterségesen előállított α-galactosyl-ceramide (GC) in vitro aktiválni képes mind a humán (81,82,83), mind egér NKT sejteket (64,83,84,85). A kölcsönhatás lényege, hogy a GC prezentálása a CD1 molekulán keresztül történik (82,83,84,85,86,87). A GC-aktivált NKT sejtek immunregulációs funkcióját bizonyították a közelmúltban (60,85,88), azt vártuk, hogy a módszer felhasználásával az EAE eredményesen befolyásolható lesz. Fontos megjegyezni, hogy az NKT sejtekkel kapcsolatos megfigyelések nem egybehangzóak, a különböző kísérleti rendszerekben egymásnak ellentmondó eredményekre
35 jutottak (Cui és mtsai szerint GC aktiválás gátolja, míg Singh és mtsai, valamint Burdin és mtsai szerint GC hatás elősegíti a hagyományos T sejtek Th2 irányú differenciálódását (60,85,88). A GC in vivo adása jelentős IL-12, IL-4 és IFN-γ termelést indított el (9. ábra), de csak kis mértékű változást (súlyosbodást) eredményezett az EAE lefolyásában. Ezzel szemben, IFN-γ és IL-4 KO egerekben a GC szignifikáns hatást váltott ki, amit az in vitro kísérletek tanúsága szerint az IFN-γ/IL-4 egyensúly eltolódásával tudtunk kapcsolatba hozni. A kísérletek bizonyítják, hogy az aktivált NKT sejtek immunregulációs kapacitása az IFN-γ és IL-4 termelés arányától függ, és ez nem változik meg a GC kezelt vad típusú egérben, ami megmagyarázza, hogy a klinikai tünetek csak kevéssé súlyosbodtak ( ábra). Kísérleteink nem foglalkoztak az NKT sejt működés valamennyi aspektusával, így nem vizsgáltuk pl. a TGFβ termelést, vagy a citotoxikus aktivitás változását, ami további kísérletek célja lehet és szintén szerepet játszhat a regulációs mechanizmusban. A továbbiakban megkíséreltünk olyan módszert kidolgozni, amellyel a vad típusú egerek NKT sejtjeinek polarizált citokin termelését idézi elő. A hagyományos T sejteken kapott eredmények szerint módosított ligand (APL), vagy a kostimulációs kapcsolatok blokkolásával a T sejtek aktivációja szub-optimális lesz, amelynek eredménye a Th1-Th2-Th3 irányú differenciálódás megváltozása (35,65). Mivel nem állt rendelkezésünkre módosított ligand, az utóbbi utat választottuk. Az in vitro GC + blokkoló antitestekkel előkezelt APC sikeresnek mutatkozott: a B7.2 receptor gátlásával az NKT sejtek citokin termelése IL-4 irányába, a T sejtek differenciálódása Th2 irányba tolódott el, míg a CD40-CD40L kapcsolat gátlása fokozott IFN-γ termeléshez és Th1 irányú differenciálódáshoz vezetett (13. ábra). Anti-B7.2 kezelt APC ennek megfelelően gátolta, anti-cd40 kezelt APC súlyosbította az EAE klinikai tüneteit. Az NKT sejtek kulcsszerepét bizonyítja, hogy NKT KO egerekben nem tudtunk hasonló hatást kimutatni ( ábra). Hipotézisünket alátámasztotta a kezelt
36 egerekben mért szérum citokinek szintjének változása (az előbbieknél IL-4, utóbbinál IFN-γ dominancia; nem közölt adatok), valamint a MOG specifikus IgG altípusok megfelelő irányú eltolódása is (kezeletlen egérben IgG2a, GC+B7.2 kezeltben IgG1 dominancia, 16. ábra). A fentiek alapján valószínű, hogy az NKT sejtek citokin fenotípusa határozza meg azok regulációs képességeit és az eredmények megmagyarázzák, hogy a különböző rendszerekben az NKT sejtek viselkedése eltérő lehet. Eredményeink bizonyítják, hogy az NKT sejtek citokin termelésének megváltoztatása racionális stratégia lehet az autoimmun betegségek befolyásolására. Passzív EAE-ben a klinikai tünetek szignifikáns súlyosbodását sikerült a recipiensek GC kezelésével előidézni. Ennek mechanizmusa nem ismert. Hátterében állhat az encephalitogén sejtek további aktivációja, a központi idegrendszerbe való könnyebb bejutása, vagy kiterjedtebb lokális szöveti destrukció. Más esetben a MOG35-55 specifikus nyirokcsomó sejteket MOG+GC együttes in vitro restimulációját követően juttattuk a recipiensekbe. Eredményeink szerint a GC dózisfüggő módon fokozta az antigén-specifikus proliferációt és IFN-γ termelést (nem közölt adatok). A várakozásnak megfelelően súlyosabb klinikai tüneteket észleltünk ( ábra). Az eredmények arra utalnak, hogy a GC-aktivált NKT sejtek szerepet játszanak az EAE effektor fázisában is, bár ennek jelentősége kisebb, mint az indukciós fázisban, hiszen az aktiv EAE-ben csak a betegség kezdetét befolyásolta az NKT sejtek eltávolítása, sem a klinikai tüntek súlyossága, sem a remisszió nem változott meg.
37 4.2. Az NKT sejt aktiválódás mechanizmusa In vitro eredményeink megerősítették az NKT sejtek GC-specifikus proliferációját, citokin termelését és citotoxikus aktivitását (6. ábra). Megjegyzendő, hogy a GC aktivált NKT sejtek IFN-γ-t termelése 5-10x magasabb, mint az IL-4 szekréció (6. és 11. ábra). Ugyanakkor az in vitro és in vivo citokin termelés kinetikája változó. IL12, IL-4 és IFN-γ termelés követi egymást (9. ábra). A pillanatnyi citokin arány tehát függ attól, mely időpontban történik a meghatározás (85,89,90). Ugyan a kísérletek többségénél nem szeparált lépsejteket használtunk, de a korábbi közlemények egyértelműen igazolták, hogy kizárólag NKT sejtek aktiválódnak GC hatására (84,91). Munkánk rávilágít az NKT aktiválódás néhány eddig ismeretlen aspektusára. GC más hagyományos stimulációval (CD3, NK1.1, IL-12) összehasonlítva heterogén válaszreakciót eredményezett: lép és perifériás vér NKT sejtek aktiválódtak, míg thymus és máj sejtek nem (7. ábra). Ennek hátterében számos tényező állhat. 1. A különböző szervek NKT sejtjeinek eltérő fenotípusa, 2. az antigén-prezentáció helyi sajátosságai. Számos laboratórium bizonyította, hogy az NKT sejtek aránya a limfocita populáción belül a különböző szervekben eltérő (lép: 1-5%, máj, csontvelő 20-40%, vér, thymus: 1-2%), azonban az abszolút sejtszám hasonló x106 (66,92). Az NKT sejtek a felszíni markerek alapján 2 csoportra oszthatók: Vα14- és Vα14+ populációra, az utóbbiak játszanak szerepet az immunregulációban (66,92,93). A sejtek fenotípusa eltérő: a lép, thymus döntően DN és CD4+ sejtekből áll, addig a csontvelő, máj CD4+, DN és CD8+ sejteket is tartalmaz. (94,95,96). CD1d KO egerek analízise alapján a thymus és máj döntően CD1d-dependens NKT sejtekből áll, míg a lép és csontvelő heterogénebb, CD1d dependens és independens NKT sejteket
38 tartalmaz. Előbbiek DN és CD4+-ak, utóbbiak CD8+ sejtek (94,97). A kísérletek tanúsága szerint a CD8+ NKT sejtek extrathymikusan fejlődnek, szemben a többi populációval, amelyek fejlődéséhez a thymus nélkülözhetetlen (66,98). Az antigén prezentáció sajátosságait vizsgáltuk a 8. ábrán bemutatott kísérletben. Azt tapasztaltuk, hogy a thymus sejtek képtelenek voltak a GC prezentálására, ezáltal a lépből izolált NKT sejtek aktiválására. Hasonló megfigyeléseket tettek NKT sejt hibridómák esetében is (99). Ezzel szemben lép, máj és DC sejtek esetén szignifikáns proliferációt és citokin termelést találtunk. Ennek egyik magyarázata lehet a CD1d, vagy a kostimulációs molekulák expressziójának a hiánya. Az előbbi nem valószínű, mivel valamennyi említett esetben magas (>90%) CD1d expressziót találtunk (nem mutatott adatok). A ko-stimulációs molekulák esetében nagy különbségek mutathatók ki. A DC sejtek többsége B7.1 és B7.2+, a többi szerv esetében igen alacsony (1-2%) B7.1 és B7.2 expresszió mutatható ki, melynek sorrendje lép>csontvelő>máj>thymus. GC kezelés hatására a B7.2 expresszió növekedését tapasztaltuk lép, csontvelő sejteken, máj és thymus esetén a B7.2 nem változott. Megjegyzendő, hogy a lépben elsősorban B sejtek prezentálják a GC-t, amik a thymusban nem találhatók meg. Az előzőeken túl vizsgáltuk az agyszövet lehetséges szerepét az immunregulációban. Nem sikerült aktivációt kimutatnunk lépből izolált NKT sejtekkel sem friss, sem kultúrában tartott agyszövet, mint APC felhasználásával. Ugyancsak nem találtunk B7 expressziót sem. Utóbbi magyarázhatja a negatív eredményeket, de nem zárja ki az NKT sejtek in vivo szerepét a lokális szöveti reakcióban. Bizonyított ugyanis, hogy központi idegrendszeri sejtek képesek peptid antigén prezentálására hagyományos T sejteknek (100,101). Mindezeken túl feltételezhetjük, egy szöveti faktor/antigén jelenlétét, amely szükséges az effektív NKT-APC felismeréshez és magyarázza, hogy a thymus sejtek miért képtelenek a GC-t lép NKT sejteknek prezentálni.
39 4.3. NKT-APC interakció A konvencionális T sejtek stimulációja során a TCR-CD3-Antigén- komplex és az MHC molekulák interakcióján túl a többféle kostimulációs molekula kapcsolódása játszódik le, előidézve a T sejtek és az APC kölcsönös aktiválódását. A legismertebb kostimulációs kapcsolatok a CD40-CD40L (L=ligand) és a CTLA-4/CD28-B7.1/B7.2 kölcsönhatás (102,103,104). Egyes megfigyelések szerint a CD28-B7 kötés a TH2, míg a CD40-CD40L interakció TH1 irányú differenciálódásnak kedvez (103,105,106). Jól ismert, hogy az NKT sejtek teljes aktiválásához is szükséges a CD28/B7 (60) és a CD40/CD40L kölcsönhatás (64,91). Jelen munka az első, amely bizonyítja, hogy ezen kölcsönhatások befolyásolása megváltoztatja az NKT sejtek fenotípusát. Eredményeink a B7.1 és B7.2 a hagyományos T sejtekétől eltérő szerepére utalnak. Kuchroo és Ranger szerint a B7.1 kostimuláció TH1 irányú differenciálódásnak kedvez, míg B7.2 Th2 irányú polarizációt idéz elő a hagyományos T sejtek esetében (65,107). Ezzel szemben úgy találtuk, hogy a B7.1 kostimuláció nem játszik szerepet az NKT sejtek aktiválásában, míg a B7.2 jelenléte esszenciális, elsősorban az IFN-γ termelés vonatkozásában (12. ábra). Mindkét kostimulációs mechanizmus alapvető szerepét bizonyították korábban EAE-ben (108,109,110,111,112,113), és feltételezik SM-ben (114,115,116). A B7 expresszió kinetikai vizsgálata tovább erősíti a hipotézist, mivel csak a B7.2 expresszió emelkedését figyeltük meg az APC (elsősorban B sejtek) felszínén ( ábra). Ennek előidézésében fontos szerepet játszik a primer TCR-CD1 interakció, azonban egyes citokinek, (IFN-γ + IL-4) jelenléte szükséges (19. ábra, 117,118,119,120). Jelen és korábbi munkák is alátámasztják, hogy GC kezelés során az NKT sejteken keresztül a lépsejtek (B sejtek, monociták, NK sejtek) poliklonális aktivációja mutatható ki (121,122). A
40 dendritikus sejtek esetében mutatható ki a legmagasabb B7 expresszió, igy nem meglepő, hogy effektiven tudják aktiválni az NKT sejteket (123). Végül az NKT és APC sejtek kölcsönhatásának hipotézisét, az aktiváció lehetséges lépéseit mutatjuk be. NKT sejt APC Vα14Jα281 - TCR CD1- GC CD40 L expresszió CD28 expresszió IL12 R expresszió CD 40 expresszió B7.2 expresszió IL-12 termelés IFN-γ /IL-4 termelés B7.2 expresszió 4.4 NKT sejtvonalak GC-specifikus (NKT) sejtvonalak fenotípusának és citokin profiljának elemzése sajátos eredményeket mutatott. A perifériás NKT sejtektől eltérően a sejtvonalak fele CD8+ volt (a másik fele DN). Feltételezzük, hogy ezen fenotípus a GC stimuláció sajátos következménye lehet, mivel korábbi NKT hibridómák és sejtvonalak kizárólag DN fenotípust mutattak (99). Igazolták továbbá, hogy a B7 kostimuláció is szerepet játszhat a CD4/CD8 fenotípus alakításában (124). A sejtek megőrizték GC-, CD1- és B7.2 -specifitásukat, ugyanakkor citokin termelésük egy kivételével Th1 irányú deviációt mutatott. EAE kísérletekben csak 1 sejtvonal esetében találtunk szignifikáns (a klinikai tüneteket
41 fokozó) hatást. A stimuláció körülményeit változtatva elméletileg lehetséges olyan sejtvonalakat előállítani, melyekkel az EAE különböző irányokba befolyásolható Perspektívák: NKT sejtek felhasználása az immunregulációban Az NKT sejtek aktiválásának számos lehetősége van, ezeket az 5. Táblázatban tüntettük fel. A megfelelő utat és módszert választva az NKT sejtek Th1, vagy Th2 irányú differenciálódását segíthetjük elő. A Th1 Th2 shift lehetősége az autoimmun betegségek (SLE, NOD/Diabetes mellitus, EAE/SM) gyógyításában használható fel (67,68,69,125). A Th2 Th1 shift allergiás betegségek befolyásolására ad lehetőséget (126,127). Biztató eredményeket értek el in vivo aktivált, vagy ex vivo kezelt NKT sejtek felhasználásával kísérletes tumorokban (72,73,74,128,129) és NOD egérben (130). Eredményeink biztonságos kezelési alternatívát kínálnak eddig gyógyíthatatlan kórképekben. A napjainkban széles körben alkalmazott dexamethason kezelés protektiv hatásmechanizmusában is szerepet tulajdonitanak az NKT sejteknek (131). Ugyankakkor fontos megjegyezni, hogy számos más lehetőség is kínálkozik az NKT sejtek polarizált/inkomplett stimulálására (5. táblázat), igy az NKT sejteken keresztül a betegségek sora válhat befolyásolhatóvá (132,133,134,135).
42 IFN-γ termelés IL-4 termelés Citotoxicitás NK Arase, 1996 (50) IL Leite de Moraes, 1998 (132) Leite de Moraes, 1999 (133) (1.,3.,9. napon) +++ +? Cui, 1999 (60) GC+α-CD ? Pál, közlés alatt Kawano, 1997 (64); Nieda, 1999 (81) Chen, 1997 (51) + ++? Gombert,1996 (134), Hameg,1999 (135) GC+α-B ? Pál, közlés alatt GC immunizáció + ++? Burdin, 1999 (88); Singh, 1999 (85) Aktiváció Th fenotípus IL-18 Th1 GC repetitiv GC Th0 α-cd3 IL-7 Th2 Referencia 5. táblázat Egér NKT sejtek stimulációjának lehetőségei Az NKT sejtek különböző stimulációja a citokin profil megváltozását eredményezi, ennek következtében az effektor T sejtek differenciálódására hatást gyakorolhatnak. +, ++, +++ a funkció stimulációjának relativ erőssége.
43 5. Összefoglalás Kísérleteink bizonyítják, hogy az NKT sejtek hasonlóan más autoimmun betegségekhez szerepet játszanak az EAE immunológiai folyamatainak szabályozásában. 1. Az NKT sejtek eltávolítása a klinikai tünetek korai megjelenéséhez vezetett, ugyanakkor a tünetek súlyossága és a gyógyulási hajlam nem változott. 2. A GC-aktivált NKT sejtek többféle citokint termelnek és citotoxikus hatást fejtenek ki. Az NKT sejtek szabályozó funkciója a termelt citokinek (IFN-γ/IL-4) arányától függ. 3. A termelt citokinek aránya a ko-stimuláción keresztül befolyásolható: CD 40 blokkolás az IFN-γ/IL-4 arány növekedéséhez, Th1 irányú deviációhoz és a klinikai tünetek súlyosbodásához vezet, mig B7.2 blokkolás az IFN-γ/IL-4 arány csökkenését, Th2 irányú deviációt és az EAE enyhébb lefolyását idézi elő. 4. A passzív EAE kísérletek alapján az NKT sejtek szerepet játszhatnak az effektor fázisban is, ennek pontos mechanizmusa nem világos, további vizsgálatokat igényel. 5. A GC kezelés szelektíven aktiválja az NKT sejteket, azonban (az NKT sejteken keresztül) más sejtféleségek aktivált állapota is kialakul: B sejtek (monociták, dendritikus sejtek) felszínén B7.2 (és más aktivációs markerek) expressziója és IL-12 termelés mutatható ki. 6. A fenti eredmények felhasználásával kidolgoztunk egy módszert, amelynek során in vitro GC aktivált és blokkoló antitestekkel kezelt APC sejtek beadásával az NKT sejtek citokin termelésének modulációján keresztül az EAE hatásosan befolyásolható. A módszer biztonságos alternatívát jelenthet a humán autoimmun, allergiás és daganatos betegségek kezelésében.
44 6. Irodalom 1. Kurtzke, J.F. MS epidemiology world wide One view of current status. Acta Neurol. Scand. 1995, 91, S23-S Pálffy, G. MS in Hungary including the Gipsy population. In Multiple sclerosis East and West. Kurland L.T., Kuroiwa Y. eds. Kyushu University Press, Fukuoka, 1982, pp Guseo, A., Jófejü, E., Kocsis, A. Epidemiology of multiple sclerosis in western Hungary In: Firnhaber W., Lauer K. eds. Multiple sclerosis in Europe: an epidemiological update. Darmstadt, Leuchtturm Verlag/LTV Press, 1994, Bencsik, K., Rajda, C., Klivényi, P., Járdánházy, T., Vécsei, L. The prevalence of multiple sclerosis in the Hungarian city of Szeged. Acta Neurol. Scand. 1998, 97, McDonnel, G.V., Hawkins, S.A. Primary progressive multiple sclerosis: a distinct syndrome? Multiple sclerosis, 1996, 2, Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: an expanded disability status scale (EDSS). Neurology, 1983, 43, Poser, C.M., Paty, D.W., Scheinbeerg, L. et al. New diagnostic criteria for multiple sclerosis: guidelines for research protocols. Ann. Neurol. 1983, 13, Gran, B., Hemmer, B., Vergelli, M., McFarland, H.F., Martin, R. Molecular mimicry and multiple sclerosis: degenerate T-cell recognition and the induction of autoimmunity. Ann. Neurol. 1999, 45, Marcadet, A., Massart, C., Semana G. et al. Association of Class II HLA-DQ beta chain DNA restriction fragments with multiple sclerosis. Immunogenetics 1985, 22, Takacs K., Kalman, B., Gyodi E. et al. Association between the lack of HLA-DQw6 and the low incidence of multiple sclerosis in Hungarian gypsies. Immunogenetics, 1990, 31, Raine, C.S, Traugott, U. Immunopathology of the lesion in multiple sclerosis and chronic relapsing experimental allergic encephalomyelitis. In: Immunoregulatory process in experimental allergic encephalomyelitis and multiple sclerosis. VandenBark, A.A., Raus, J.C.M. eds. Elsevier, pp
45 12. Trapp, B.D., Peterson, J., Ransohoff, R.M. et al. Axonal transection in the lesions of multiple sclerosis. N. Engl. J. Med. 1998, 338, Prineas, J.W., Barnaard, R.O., Kwon, E.E. et al. Multiple sclerosis: remyelination of nascent lesions. Ann. Neurol. 1993, 33, Tsunoda, I., Fujinami, R.S. Two models for multiple sclerosis: experimental autoimmune encephalomyelitis and Theiler s murine encephalomyelitis virus. J. Neuropathol. Exp. Neurol. 1996, 55, Miller, S.D., Gerety, S.J. Immunologic aspects of Theiler s murine encephalomyelitis virus (TMEV)-induced demyelinating disease. Seminol. Virol. 1990, 1, Weller, R.O. Animal models of demyelinating disease. Curr. Op. Neurol. Neurosurg. 1991, 4, Brown, A.M., McFarlin, D.E. Relapsing experimental allergic encephalomyelitis in the SJL/J mouse. Lab. Invest. 1981, 45, Rivers, T.M., Sprunt, D.H., Berry, G.P. Observation on attempts to produce acute disseminated encephalomyelitis in monkeys. J. Exp. Med. 1933, 58, Rosbo, N.K., Milo, R., Lees, M.B., Burger, D., Bernard, C.C.A., Ben-Nun, A. Reactivity to myelin antigens in multiple sclerosis. Peripheral blood lymphocytes respond predominantly to myelin oligodenrocyte glycoprotein. J. Clin. Invest. 1993, 92, Storch, M.K., Stefferi, A., Brehm, U., Weissert, R., Wallström, E., Kerschensteiner, M., Olsson, T., Linington, C., Lassmann, H. Autoimmunity to myelin oligodendrocyte glycoprotein in rats mimics the spectrum of multiple sclerosis pathology. Brain Pathol. 1998, 8, Freund, J., McDermott, K. Sensitization to horse serum by means of adjuvants. Proc. Soc. Exp. Biol. Med. 1942, 49, Fazekas, G., Tabira, T. What transgenic and knockout mouse models teach us about experimental autoimmune encephalomyelitis. Rev. Immunogenet. 2000, 2, Encinas, J.A., Lees, M.B., Sobel, R.A. et al. Genetic analysis of susceptibility to experimental autoimmune encephalomyelitis in a cross between SJL/J and B10.S mice. J. Immunol. 1996, 157, Kuchroo, V.K., Sobel, R.A., Laning, J.C. et al. Experimetal allergic encephalomyelitis mediated by cloned T cells specific for a synthetic peptide of myelin proteolipid protein: fine specificity and T cell receptor Vβ usage. J. Immunol. 1992, 148,
46 25. Mendel, I., Kerlero de Rosbo, N., Ben.Nun, A. A myelin oligodendrocyte glycoprotein peptide induces typical chronic experimental autoimmune encephalomyelitis in H-2b mice: fine specificity and T cell receptor V beta expression of encephalitogenic T cells. Eur. J. Immunol. 1995, 25, Zhang, B-N, Yamamura T., Kondo, T., Fujiwara, M., Tabira T. Regulation of experimental autoimmune encephalomyelitis by natural killer (NK) cells. J. Exp. Med, 1997, 186, Lassmann, H., Wisniewski, H.M. Chronic relapsing experimental allergic encephalomyelitis. Clinicopathological comparison with multiple sclerosis. Arch. Neurol. 1979, 36, Kálmán, B., Lublin, F.D. Immunopathogenic mechanisms in experimental allergic encephalomyelitis. Curr. Op. Neurol. Neurosurg. 1993, 6, Traugott, U., McFarlin, D.E., Raine, C.S. Immunpathology of the lesion in chronic relapsing experimental autoimmune encephalomyelitis in the mouse. Cell. Immunol. 1986, 99, Vanderlugt, C.L., Neville, K.L., Nikcevich, K.M., Eagar, T.N., Bluestone, J.A., Miller, S.D. Pathologic role and temporal appearance of newly emerging autoepitopes in relapsing experimental autoimmune encephalomyelitis. J. Immunol. 2000, 164, Alvord, E.C, Kies, M.W., Suckling, A.J. Experimental allergic encephalomyelitis: a useful model for multiple sclerosis. Alan R. Liss, 1984, New York. 32. Steinman, L. Assessment of animal models for MS and demyelinating disease in design of rational therapy. Neuron, 1999, 24, Tabira, T., Webster, H., Wray, S.H. Multiple sclerosis cerebrospinal fluid produces myelin lesions in tadpode optic nerves. N.E.J. M. 1976, Genain, C.P., Cannella, B., Hauser, S.L., Raine, C.S. Identification of autoantibodies associated with myelin damage in multiple sclerosis. Nature Med. 1999, 5, Kuchroo, V.K., Greer, J.M., Kaul, D., Ishioka, G., Frano, A., Sette, A., Sober, R.A., Lees, M.B. A single TCR antagonist peptide inhibits experimental allergic encephalomyelitis mediated by a diverse T cell repertoire. J. Immunol. 1994, 153, Hafler, D.A., Saadeh, M.G., Kuchroo, V.K., Milford, E., Steinman, L. TCR usage in human and experimental demyelinating disease. Immunol. Today 1996, 17,
47 37. Kuchroo, V.K., Chen, Y.C., Inobe, J-I., Hafler, D.A., Weiner, H.L. Regulatory T cell clones induced by oral tolerance: suppression of autoimmune encephalomyelitis. Science, 1994, 265, Sakaguchi, S., Sakaguchi, N., Asano. M., Itoh, M., Toda, M. Immunologic selftolearnace maintaned by activated T cells expressing IL-2 receptor α-chains (CD25). J. Immunol. 1995, 155, Falcone, M., Yeung, B., Tucker, L., Rodriguez, E., Sarvetnick, N. A defect in interleukin 12-induced activation and interferon-γ secretion of peripheral natural killer T cells in nonobese diabetic mice suggests new pathogenic mechanisms for insulindependent diabetes mellitus. J. Exp. Med. 1999, 190, Stochkman, S.A., Pei, L., Cua, D.J., Li, Z., Hinton, D.R. Activation of regulatory cells suppresses experimental allergic encephalomyelitis via secretion of IL-10. J. Immunol. 1999, 163, Lohse, A.W., Mor, F., Kalin, N., Cohen, IR. Control of experimental autoimmune encephalomyelitis by T cells responding to activated T cells. Science, 1989, 244, Rajan, A.J., Gao, Y.L, Raine, C.S., Brosnan, C.I. A pathogenic role for gamma delta T cells in relapsing-remitting experimental autoimmune encephalomyelitis in the SJL mouse. J. Immunol. 1996, 157, Matsumoto, Y., Kohyama, K., Aikawa, Y., Shin, T., Kawazoe, Y., Suzuki, Y., Tamema, N. Role of natural killer cell and TCRγδ T cells in acute autoimmune encephalomyelitis. Eur. J. Immunol. 1998, 28, Spahn, T.W., Issazadah, S., Salvin, A., Weiner, H.L. Decreased severity of myelin oligodendrocyte glycoprotein peptide induced experimental autoimmune encephalomyelitis in mice with a disrupted TCR δ chain gene. Eur. J. Immunol. 1999, 29, Seder, R.A., Paul, W.E. Acquisition of lymphokine-producing phenotype by CD4+ T cells. Annu. Rev. Immunol. 1994, 12, Assenmacker, M., Löhning, M., Scheffold, A., Richter, A., Miltényi, S., Schmitz, J., Radbuch, A. Commitment of individual Th1-like lymphocytes to expression of IFN-γ versus IL-4 and IL-10: selective induction of IL-10 by sequential stimulation of naive Th cells with IL-12 and IL 4. J. Immunol. 1998, 161,
48 47. Van der Veen, R., Stohlman, S.A. Encephalitogenic Th1 cells are inhibited by Th2 cells with related peptide specificity: relative roles of interleukin (IL)-4 and IL-10. J. Neuroimmunol. 1993, 48, Ramirez, F., Mason, D. Induction of resistance to active experimental allergic encephalomyelitis by myelin basic protein-specific Th2 cell lines generated in the presence of glucocorticoids and IL-4. Eur. J. Immunol, 2000, 30, Fernandez, N.C., Lozier, A., Flament, C. et al. Dendritic cells directly trigger NK cell functions: cross-talk relevant in innate anti-tumor immune responses in vivo. Nature Med. 1999, 5, Arase, H., Arase, N., Saito, T. Interferon γ production by natural killer (NK) cells and NK1.1+ T cells upon NKR-P1 cross-linking. J. Exp. Med. 1996, 183, Chen, H., Paul, W.E. Cultured NK1.1+CD4+ T cells produce large amounts of IL-4 and IFN-γ upon activation by anti-cd3 or CD1. J. Immunol. 1997, 159, Tamada, K., Harada, M., Abe, K., Li, T., Tada, H., Onoe, Y., Nomoto, K. Immunosuppressive activity of cloned natural killer (NK1.1+) T cells established from murine tumor-infiltrating lymphocytes. J. Immunol. 1997, 158, Asea, A., Stein-Streilein, J. Signalling through NK 1.1 triggers NK cells to die but induces NK T cells to produce interleukin-4. Immunology, 1998, 93, Schmied, M., Breitschopf, H., Gold, R., Zischer H., Rothe, G., Wekerle, H., Lassmann, H. Apoptosis of T lymphocytes in experimetal autoimmune encephalomyelitis: evidence for programmed cell death as a mechanism to control inflammation in the brain. Am. J. Pathol. 1993, 143, Kennedy, M.K., Torrance, D.S., Picha, K.S., Mohler, K. M. Analysis of cytokine mrna expression in the central nervous system of mice with experimental autoimmune encephalomyelitis reveals that IL-10 mrna expression correlates with recovery. J. Immunol. 1992, 149, Begolka, W.S., Miller, S.D. Cytokines as intrinsic and exogenous regulators of pathogenesis in experimental autoimmune encephalomyelitis. Res. Immunol. 1998, 149, Nguyen, K.B., McCombe, P.A., Pender, M.P. Increased apoptosis of T lymphocytes and macrophages in the central and peripheral nervous system of Lewis rat with experimental autoimmune encephalomyelitis treated with dexamethasone. J. Neuropath. Exp. Neurol. 1997, 56,
49 58. Josefsson, E., Bergquist, J., Ekman, R., Tarkowski, A. Catecholamines are synthesized by mouse lymphocytes and regulate function of these cells by induction of apoptosis. Immunology, 1996, 88, Smeltz, R.B., Wolf, N.A., Swanborg, R.H. Inhibition of autoimmune T cell responses in the DA rat by bone marrow-derived NK cells in vitro: implications for autoimmunity. J. Immunol 1999, 163, Cui, J., Watanabe, N., Kawano, T. et al. Inhibition of T helper cell type 2 cell differentiation and immunoglobulin E response by ligand-activated Vα14 natural killer T cells. J. Exp. Med. 1999, 190, Morita, M., Natori, T., Akimoto, K., Osawa, T., Fukushima, H., Koezuka, Y. Synthesis of α-, β-monoglycosylceramides and four diastereomers of an αgalactosylceramide. Bioorganic and Medicin. Chem. Lett. 1995, 5, Motoki, K., Morita, M., Kobayashi, E., Uchida, T., Akimoto, K., Fukushima, H., Koezuka, Y. Immunostimulatory and antitumor activities of monoglycosyl-ceramides having various sugar moieties, Biol. Pharm. Bull. 1995, 18, Slezak, SE., Horan, PK. Cell-mediated cytotoxicity. A highly sensitive and informative flow cytometric assay. J. Immunol. Methods 1989, 117, Kawano, T., Cui, J., Koezuka, Z. et al. CD1-restricted and TRC-mediated activation of Vα14 NKT cells by glycosylceramides. Science 1997, 278, Kuchroo, V.K., Das, M.P., Brown, J.A., Ranger, A.M., Zamvil, S.S., Sober, R.A., Weiner, H.L., Nabavi, N., Glimcher, L.H. B7-1 and B7-2 costimulatory molecules activate differentially the Th1/Th2 developmental pathways: application to autoimmune disease therapy. Cell, 1995, 80, Bendelac, A., Rivera, M.N., Park, S-H., Roark, J.H. Mouse CD1-specific NK1 T cells: development, specificity, and function. Annu. Rev. Immunol. 1997, 15, Mieza, M.A., Itoh, T., Cui, J.Q. et al. Selective reduction of Vα14+ NK T cells associated with disease development in autoimmune-prone mice. J. Immunol. 1996, 156, Gombert, J.M., Herbelin, A., Tancrede-Bohin, E, Carnaud, Dy, M., Carnaud, C., Bach, J.F. Early quantitative and functional deficiency of NK1.1+-like thymocytes in the NOD mouse. Eur. J. Immunol. 1996, 26, Illés, Zs., Kondo, T., Newcombe, J., Oka, N., Tabira, T., Yamamura, T. Differential expression of NK T cell Vα24JαQ invariant TCR chain in the lesions of multiple
50 sclerosis and chronic inflammatory demyelinating polyneuropathy. J. Immunol. 2000, 164, Yoshimoto, T., Bendelac, A., Hu-Li, J., Paul, W.E. Defective IgE production by SJL mice is linked to the absence of CD4+, NK1.1+ T cells that promptly produce interleukin-4. Proc. Natl. Acad. Sci. USA 1995, 92, Kaneko, Y., Harada, M., Kawano, T., Yamashita, M., Shibata, Y., Gejyo, F., Nakayama, T., Taniguchi, M. Augmentation of Vα14 NKT cell-mediated cytotoxicity by interleukin 4 in an autocrine mechanism resulting in the development of concanavalin A-induced hepatitits. J. Exp. Med. 2000, 191, Nicol, A., Nieda, M., Koezuka, Y., Porcelli, S., Suzuki, K., Tadokoro, K., Durrant, S., Juji, T. Human invariant Vα24+ natural killer T cells activated by αgalactosylceramide (KRN7000) have cytotoxic anti-tumor activity through mechanisms distinct from T cells and natural killer cells. Immunology, 2000, 99, Smyth, M.J., Thia, K.Y.T., Street, S.E.A. et al. Differential tumor surveillance by natural killer (NK) and NKT cells. J. Exp. Med. 2000, 191, Cui, J., Shin, T., Kawano, T. et al. Requirement for Vα14 NKT cells in IL-12mediated rejection of tumors. Science, 1997, 278, Nilsson, N., Bremell, T., Tarkowski, A., Carlsten, H. Protective role of NK1.1+ cells in experimental Staphylococcus aureus arthritis. Clin. Exp. Immunol. 1999, 117, Naiki, Y., Nishimura, H., Kawano, T., Tanaka, Y., Itohara, S., Taniguchi, M., Yoshikai, Y. Regulatory role of peritoneal NK1.1+αβ T cells in IL-12 production during Salmonella infection. J. Immunol. 1999, 163, Sonoda, K-H., Exley, M., Snapper, S., Balk, S.P., Stein-Streilein, J. CD1-reactive natural killer T cells are required for development of systemic tolerance through an immune-privileged site. J. Exp. Med. 1999, 190, Zeng, D., Lewis, D., Dejbakhsh-Jones, S., Lan, F., Garcia-Ojeda, M., Sibley, R., Strober, S. Bone marrow NK1.1- and NK1.1+ T cells reciprocally regulate acute graft versus host disease. J. Exp. Med. 1999, 189, Lehuen, A., Lanty, O., Beaudion, L., Laloux, V., Carnaud, C., Bendelac, A., Bach, J.F., Monteiro, R.C. Overexpression of natural killer T cells protects Vα14-Jα281
51 transgenic nonobese diabetic mice against diabetes. J. Exp. Med. 1998, 188, Zeng, D., Dick, M., Cheng, L., Amano, M., Dejbakhsh-Jones, S., Huie, P, Sibley, R., Strober, S. Subsets of transgenic T cells that recognize CD1 induce or prevent murine lupus: role of cytokines. J. Exp. Med. 1998, 187, Nieda, M., Nicol, A., Koezuka, Y. et al. Activation of human Vα24NKT cells by αglycosylceramide in a CD1d-restricted and Vα24TCR-mediated manner. Hum. Immunol. 1999, 60, Spada, F., Koezuka, Y., Porcelli, S.A. CD1-restricted recognition of synthetic glycolipid antigens by human natural kiler T cells. J. Exp. Med. 1998, 188, Brossay, L., Chioda, M., Burdin, N., Koezuka, Y., Casorati, G., Dellabona, P., Kronenberg, M. CD1d-mediated recognition of an α-galactosylceramide by natural killer cells is highly conserved through mammalian evolution. J. Exp. Med. 1998, 188, Burdin, N., Brossay, L., Koezuka, Y., Smiley, S.T., Grusby, M.J., Gui, M., Taniguchi, M., Hayakawa, K., Kronenberg, M. Selective ability of mouse CD1 to present glycolipids: α-galactosylceramide specifically stimulates Vα14+ NK T lymphocytes. J. Immunol. 1998, 161, Singh, N., Hong, S., Scherer, D.C., Serizawa, I., Burdin, N., Kronenberg, M., Koezuka, Y., Kaer, L.V. Activation of NK T cells by CD1d and α-galactosylceramide directs conventional T cells to the acquisition of a Th2 phenotype. J. Immunol. 1999, 163, Burdin, N., Kronenberg, M. CD1-mediated immune responses to glycolipids. Curr. Op. Immunol. 1999, 11, Kronenberg, M., Brossay, L., Kurepa, Z., Forman, J. Conserved lipid and peptide presentation functions of nonclassical class I molecules. Immunol. Today 1999, 20, Burdin, N., Brossay, L., Kronenberg, M. Immunization with α-galactosylceramide polarizes CD1-reaktive NK T cells towards Th2 cytokine synthesis. Eur. J. Immunol. 1999, 29, Kitamura, H., Ohta, A., Sekimoto, M. et. al. α-galactosylceramide induces early Bcell activation through IL-4 production by NKT cells. Cell. Immunol. 2000, 199, 3742.
52 90. Tomura, M., Yu, W-G., Ahn, H-J. et al. A novel function of Vα14+CD4+NKT cells: stimulation of IL-12 production by antigen-presenting cells in the innate immune system. J. Immunol. 1999, 163, Kitamura, H., Iwakabe, K., Zahata, T. et al. The natural killer T (NKT) cell ligand αgalactosylceramide demonstrates its immunopotentiating effect by inducing interleukin (IL)-12 production by dendritic cells and IL-12 receptor expression on NKT cells. J. Exp. Med. 1999, 189, Makino, Y., Kanno, B., Ito, T., Higashino, K., Taniguchi, M. Predominant expression of invariant Vα14+ TCRα chain in NK1.1+ T cell populations. Int. Immunol. 1995, 7, Masuda, K., Makino, Y., Cui, J. et al. Phenotypes and invariant αβ TCR expression of peripheral Vα14+ NK T cells. J. Immunol. 1997, 158, Eberl, G., Lees, R., Smiley, S.T., Taniguchi, M., Grusby, M.J., MacDonald, H.R. Tissue-specific segregation of CD1d-dependent and CD1d-independent NK T cells. J. Immunol. 1999, 162, Hammond, K.J.L., Pelikan, S.B., Crowe, N.Y. et al. NKT cells are phenotypically and functionally diverse. Eur. J. Immunol. 1999, 29, Zeng, D., Gazit, G., Dejbakhsh-Jones, S., Balk, S.P., Snapper, S., Taniguchi, M., Strober, S. Heterogeneity of NK1.1+ T cells in the bone marrow: divergence from the thymus. J. Immunol. 1999, 163, Coles, M.C., Raulet, D.H. Class I dependence of the development of CD4+CD8NK1.1+ thymocytes. J. Exp. Med. 1994, 180, Hammond, K., Cain, W., van Driel, I., Godfrey, D. Three day neonatal thymectomy selectively depletes NK1.1+ T cells. Int. Immunol. 1998, 10, Park, S-H., Roark, J., Bendelac, A. Tissue-specific recognition of mouse CD1 molecules. J. Immunol. 1998, 160, Xiao, B-G., Link, H. Immune regulation within the central nervous system. J. Neurol. Sci. 1998, 157, Katz-Levy, Y., Neville, K.L., Girvin, A.M., Vanderlugt, C.L., Pope, J.G., Tau, L.J, Miller, S.D. Endogenous presentation of self myelin epitopes by CNS-resident APCs in Theilers virus infected mice. J. Clin. Invest, 1999, 104, Greenfield, E.A., Nguyen, K.A., Kuchroo, V.K. CD28/B7 costimulation: a review. Crit. Rev. Immunol. 1998, 18,
53 103. Howland, K.C., Ausubel, L.J., London, C.A., Abbas, A.K. The roles of CD28 and CD40 ligand in T cell activation and tolerance. J. Immunol. 2000, 164, Durie, F.H., Foy, T.M., Masters, S.R., Laman, J.D., Noelle, R.J. The role of CD40 in the regulation of humoral and cell-mediated immunity. Immunol. Today, 1994, 15, Rulifson, I.C., Sperling, A.I., Fields, P.E., Fitch, F.W., Bluestone, J.A. CD28 costimulation promotes the production of Th2 cytokines. J. Immunol. 1997, 158, Stout, R., Stuttles, J. The many roles of CD40:CD40L interactions in cell-mediated inflammatory responses. Immunol. Today, 1996, 17, Ranger, A.M., Das, M.P., Kuchroo, V.J., Glimcher, L.H. B7-2 (CD86) is essential for the development of IL-4 producing T cells. Int. Immunol. 1996, 8, Chang, T.T., Jabs, C., Sobel, R.A., Kuchroo, V.J., Sharpe, A.H. Studies in B7deficient mice reveal a critical role for B7 costimulation in both induction and effector phases of experimental autoimmune encephalomyelitis. J. Exp. Med. 1999, 190, Schaub, M., Issazadeh, S., Stadlbauer, T.H.W., Peach, R., Sayegh, M.H., Khoury, S.J. Costimulatory signal blockade in murine relapsing experimental autoimmune encephalomyelitis. J. Neuroimmunol. 1999, 96, Miller, S.D., Vanderlugt, C.L., Lenschow, D.J., Pope, J.G., Karandikar, N.J., Dal Canto, M.C., Bluestone, J.A. Blockade of CD28/B7-1 interaction prevents epitope spreading and clinical relapses of murine EAE. Immunity, 1995, 3, Grewal, I.S., Foellmer, H.G., Grewal, K.D., Xu, J., Hardardottir, F., Baron, J.L., Janeway, C.A., Flavell, R.A. Requirement for CD40 ligand in costimulation induction, T cell activation, and experimental allergic encephalomyelitis. Science, 1996, 273, Karandikar, N.J., Vanderlugt, C.L., Bluestone, J.A., Miller, S.D. Targeting the B7/CD28:CTLA-4 costimulatory system in CNS autoimmune disease. J. Neuroimmunol. 1998, 89, Karandikar, N.J., Vanderlugt, C.L., Eagar, T., Tan, L., Bluestone, J.A., Miller, S.D. Tissue-specific up-regulation of B7-1 expression and function during the course of murine relapsing experimental autoimmune encephalomyelitis. J. Immunol. 1998, 161,
54 114. Correale, J. McMillan, M., Li, S., McCarthy, K., Le, T. Weiner, L.P. Antigen presentation by autoreactive proteolipid protein peptide-specific T cell clones from chronic progressive multiple sclerosis: roles of co-stimulatory B7 molecules and IL12. J. Neuroimmunol. 1997, 72, Monteyne, P., Guillauma, B., Sindic, C.J.M. B7-1 (CD80), B7-2 (CD86), interleukin12 and transforming growth factor-β mrna expression in CSF and peripheral blood mononuclear cells from multiple sclerosis patients. J. Neuroimmunol. 1998, 91, Windhagen, A., Maniak, S., Gebert, A., Ferger, I., Heidenreich, F. Costimulatory molecules B7-1 and B7-2 on CSF cells in multiple sclerosis and optic neuritis. J. Neuroimmunol. 1999, 96, Constant, S.L. B lymphocytes as antigen-presenting cells for CD4+ T cell priming in vivo. J. Immunol. 1999, 162, Roth, R., Nakamura, T., Mamula, M.J. B7 costimulation and autoantigen specificity enable B cells to activate autoreactive T cells. J. Immunol. 1996, 157, Hathcock, K.S., Laszlo G., Pucillo, C., Linsley, P., Hodes, R.J. Comparative analysis of B7-1 and B7-2 costimulatory ligands: expression and function. J. Exp. Med. 1994, 180, Lenschow, D.J., Walunas, T.L., Bluestone, J.A. CD28/B7 system of T cell costimulation. Annu. Rew. Immunol. 1996, 14, Carnaud, C., Lee, D., Donnars, O., Park, S-H., Beavis, A., Koezuka, Y., Bendelac, A. Cross-talk between cells of innate immune system: NKT cells rapidly activate NK cells. J. Immunol. 1999, 163, Eberl, G., MacDonald, R. Selective induction of NK cell proliferation and cytotoxicity by activated NKT cells. Eur. J. Immunol. 2000, 30, Inaba, K., Witmer-Pack, M., Inaba, M., et al. The tissue distribution of the B7-2 costimulation in mice: abundant expression on dendritic cells in situ and during maturation in vitro. J. Exp. Med. 1994, 180, Yu, X., Fournier, S., Allison, J.P., Sharpe, A.H., Hodes, R.J. The role of b7 costimulation in CD4/CD8 T cell homeostasis. J. Immunol. 2000, 164, Wilson, S.B., Kent, S., Patton, K.T. et al. Extreme Th1 bias of invariant Vα24JαQ T cells in type 1 diabetes. Nature 1998, 391,
55 126. Hofer, M.F., Jirapongsababuruk, O., Trumble, A.E., Leung, D.Y.M. Upregulation of B7.2 but not B7.1 on B cells from patients with allergic asthma. J. Allergy Clin. Immunol. 1998, 101, Jirapongsananuruk, O., Hofer, M.F., Trumble, A.E., Norris, D.A., Leung, D.Y.M. Enhanced expression of B7.2 (CD86) in patients with atopic dermatitis: a potential role in the modulation of IgE synthesis. J. Immunol. 1998, 160, Toura, I., Kawano, T., Akutsu, Y., Nakayama, T., Ochiai, T., Taniguchi M. Inhibition of experimental tumor metastasis by dendritic cells pulsed with α-galactosylceramide. J. Immunol. 1999, 163, Kawano, T., Cui, J., Koezuka, Y., et al. Natural killer-like nonspecific tumor cell lysis mediated by specific ligand-activated Vα14 NKT cells. Proc. Natl. Acad. Sci. USA. 1998, 95, Hammond, K.J.L., Poulton, L.D., Palmisano, L.J., Silveira, P.A., Godfrey, D.I., Baxter, A.G. α/β-t cell receptor (TCR)+CD4-CD8- (NKT) thymocytes prevent insulin-dependent diabetes mellitus in nonobese diabetic (NOD)/Lt mice by the influence of interleukin (IL)-4 and/or IL-10. J. Exp. Med. 1998, 187, Milner, J.D., Kent, S.C., Ashley, T.A., Wilson, S.B., Strominger, J.L., Hafler, D.A. Differential responses of invariant Vα24JαQ T cells and MHC class II-restricted CD4+ T cells to dexamethasone. J. Immunol. 1999, 163, Leite-de-Moraes, M.C., Moreau, G., Arnould, A., Machavoine, F., Garcia, C., Papiernik, M., Dy, M. IL-4-producing NK T cells are biased towards IFN-γ production by IL-12. Influence of the microenvironment on the functional capacities of NK T cells. Eur. J. Immunol. 1998, 28, Leite-de-Moraes, M.C., Hameg, A., Arnould, A., Machavoine, F., Koezuka, Y., Schneider, E., Herbelin, A., Dy, M. A distinct IL-18-induced pathway to fully activate NK T lymphocytes independently from TCR engagement. J. Immunol. 1999, 163, Gombert, J-M., Tancrede-Bohin, E., Hameg, A., Leite.de.Moraes, M., Vicari, A., Bach, J.F., Herbelin, A. IL-7 reverses NK1+ T cell-defective IL-4 production in the non-obese diabetic mouse. Int. Immunol. 1996, 8, Hameg, A., Gouarin, C., Gombert, J-M., Hong, S., Kaer, L.V., Bach, J-F., Herbelin, A. IL-7 up-regulates IL-4 production by splenic NK1.1+ and NK1.1- MHC class Ilike/CD1-dependent CD4+ T cells. J. Immunol. 1999, 162,
56 7.Ábrák
57
58
59
60
61
62
63
64
65

66
67
68
69 Köszönetnyilvánítás Hálával tartozom családomnak a vég nélküli türelemért, amivel munkámat lehetővé tették. Köszönet illeti Dr. Czopf József professzort, amiért támogatta e tudományos munka elvégzését. Köszönetemet fejezem ki Dr. Takeshi Tabirának, akinek nemzetközileg is elismert intézetében dolgozhattam. Dr. Takashi Yamamura nemcsak szakmai de baráti tanácsaival is támogatta munkámat. Köszönettel tartozom Dr. Szekeres Júliának, értékes segítségéért és tanácsaiért, amivel hozzájárult munkám elkészítéséhez.
70 Függelék Az értekezéshez kapcsolódó közlemények

71
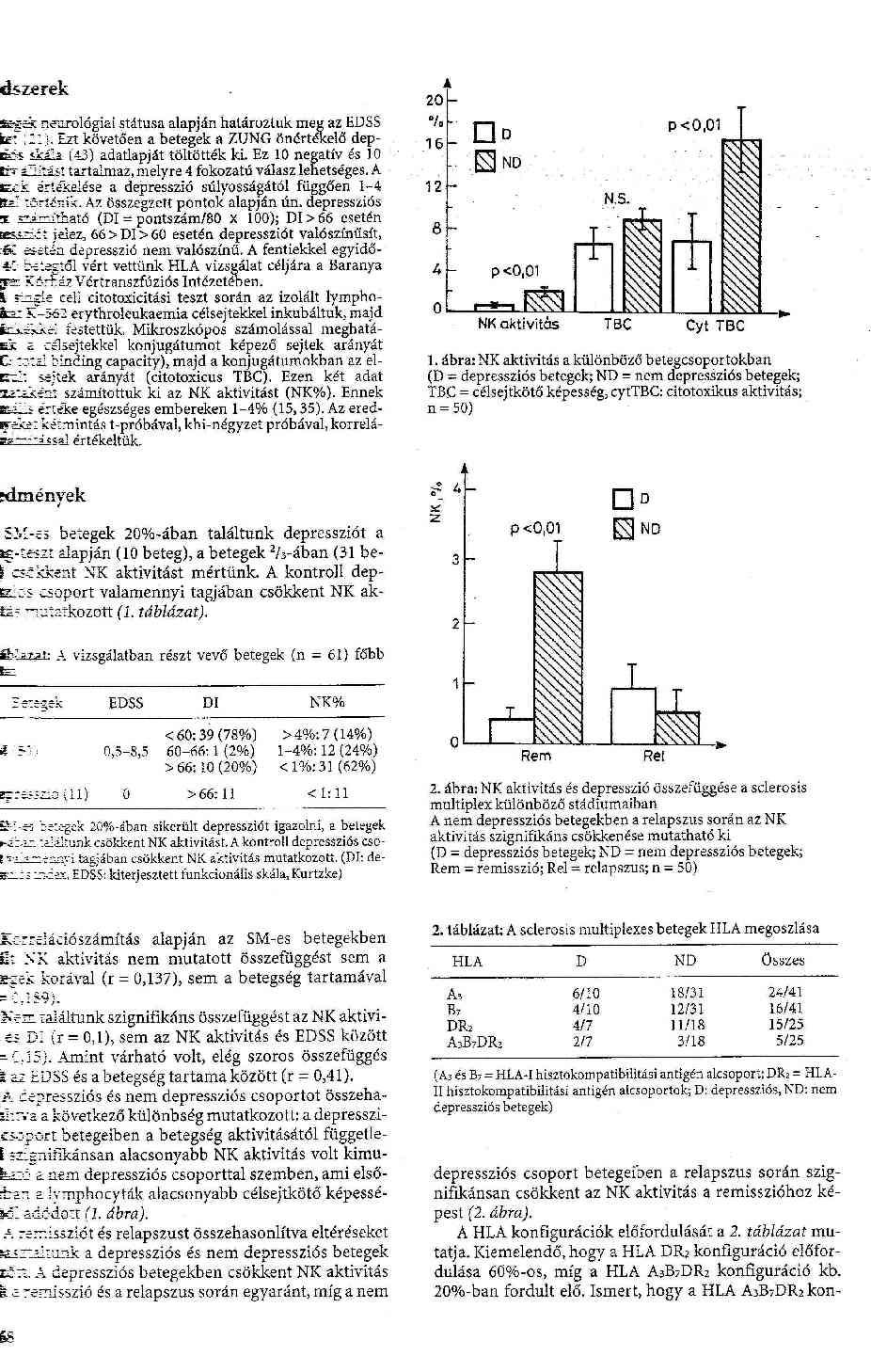
72

73

74

75

76
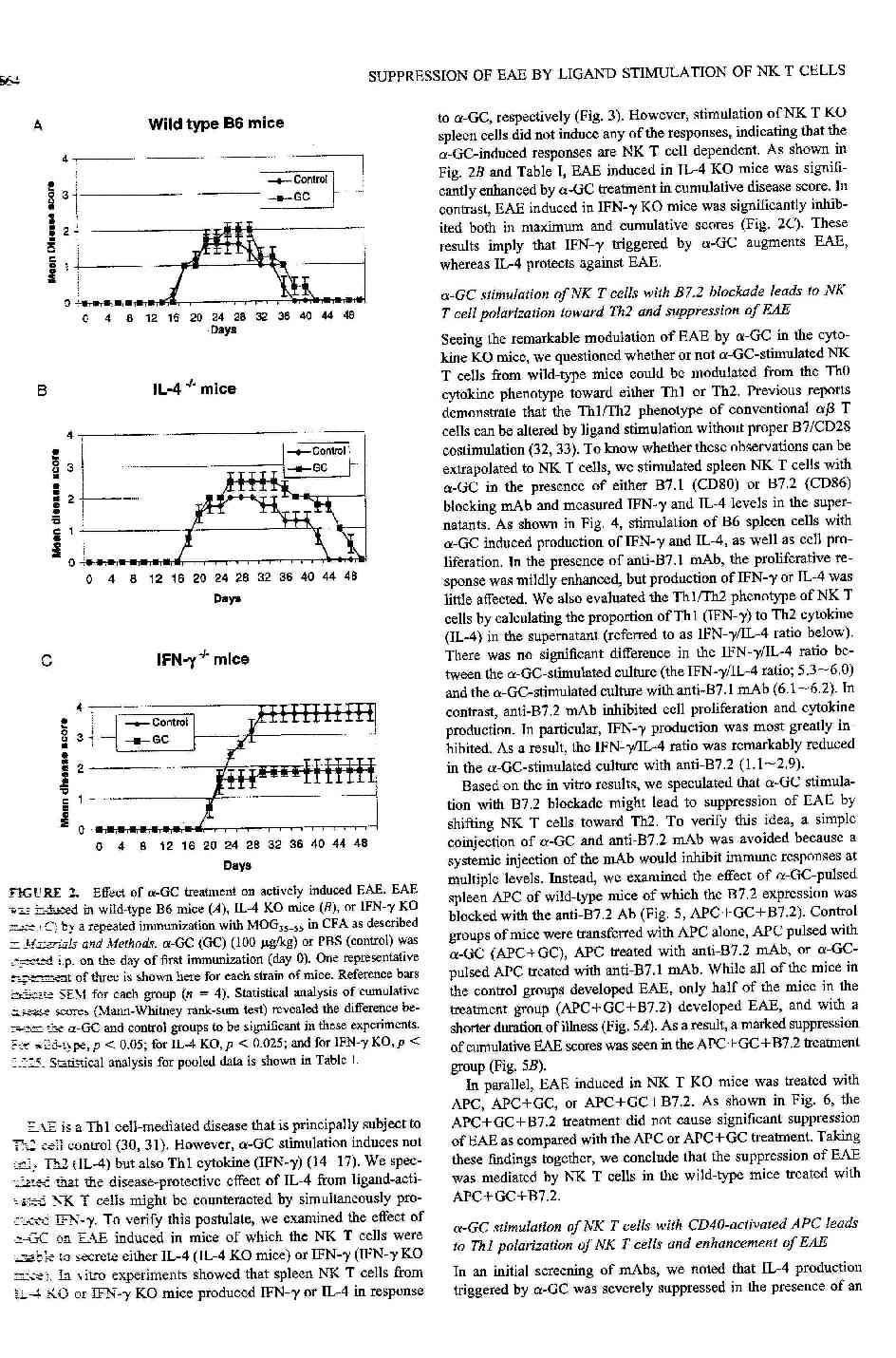
77
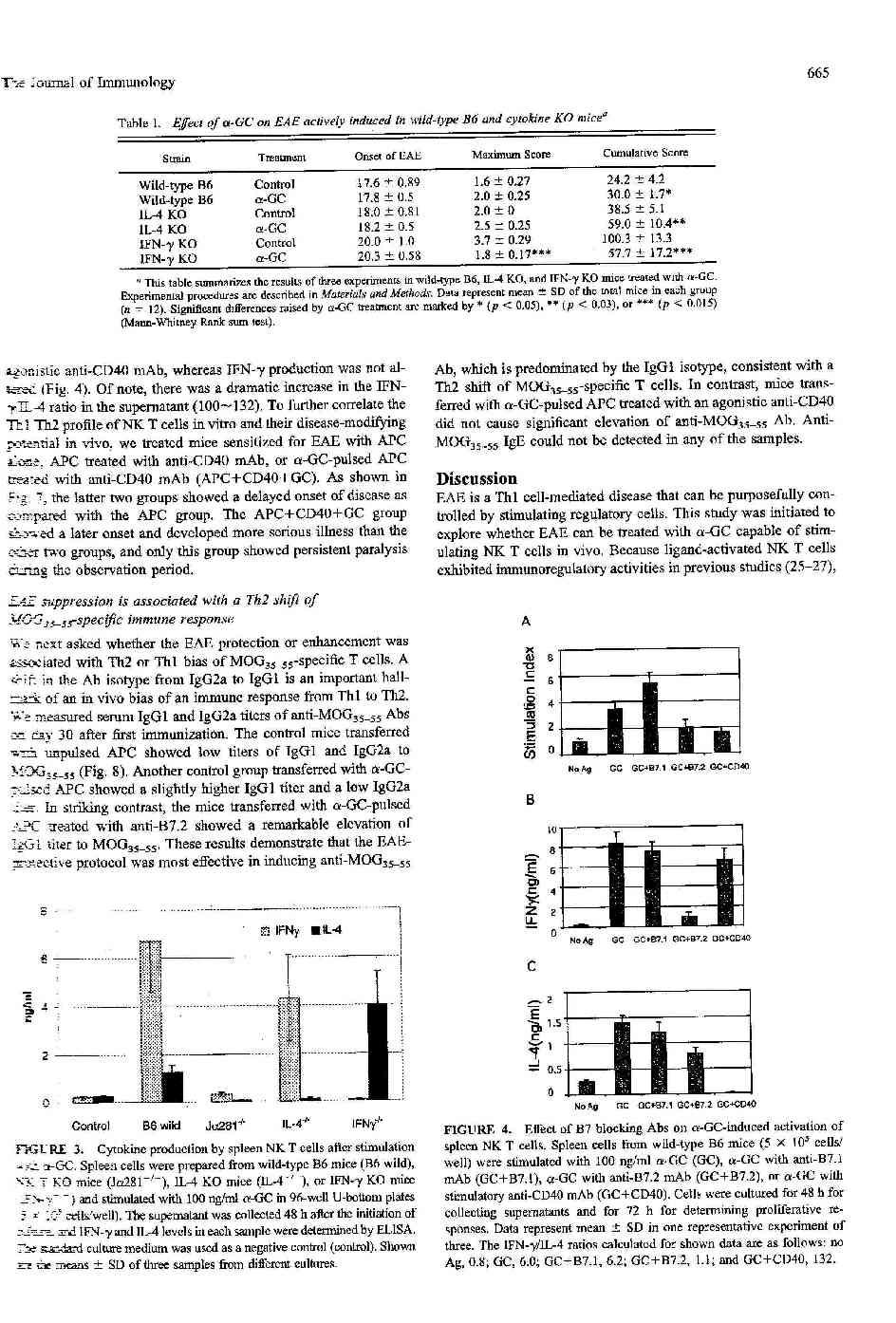
78
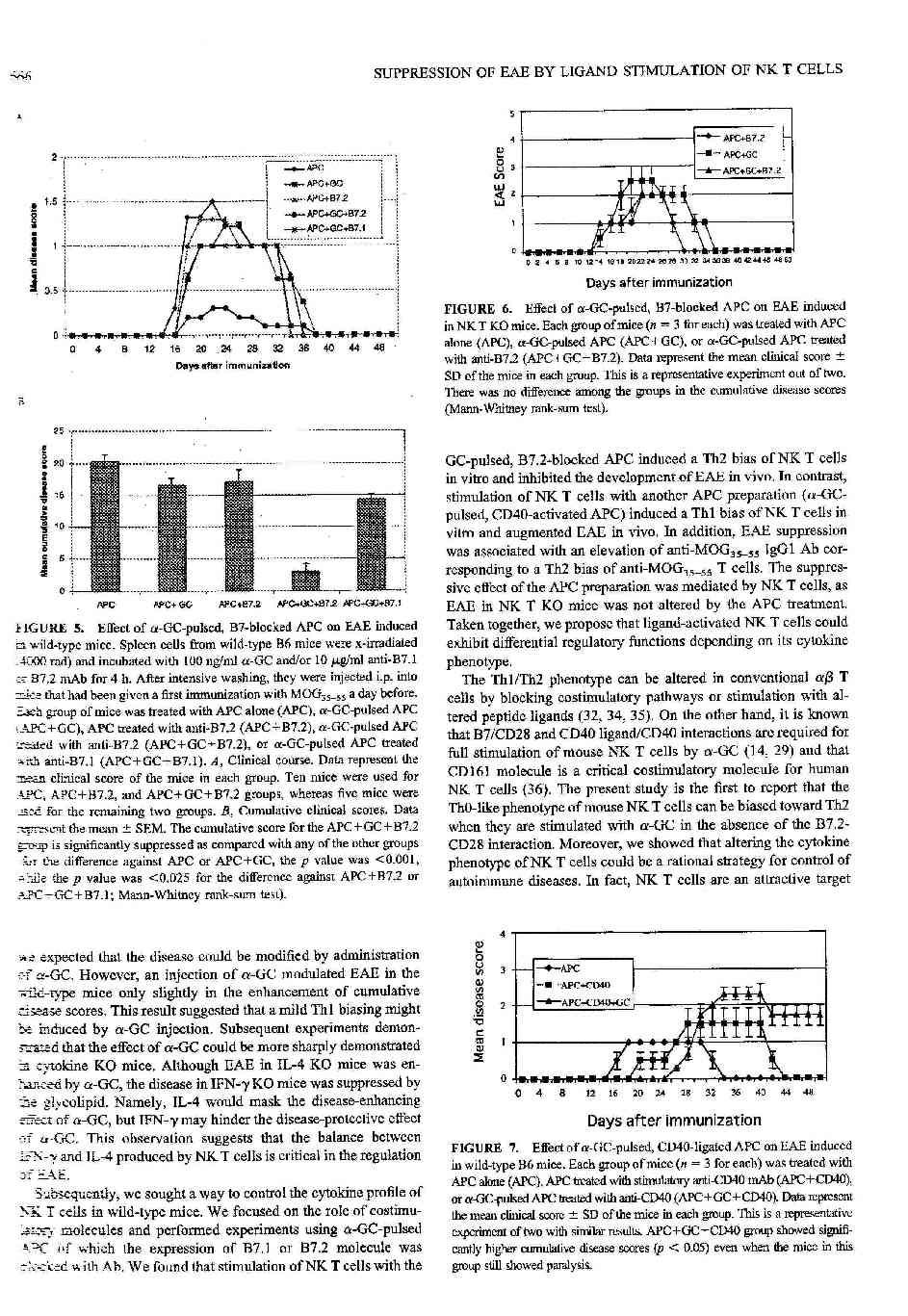
79

80

81
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
IMMUNREGULÁCIÓ KÍSÉRLETES ENCEPHALOMYELITISBEN: AZ NKT SEJTEK SZEREPE
 IMMUNREGULÁCIÓ KÍSÉRLETES ENCEPHALOMYELITISBEN: AZ NKT SEJTEK SZEREPE Ph.D. tézisek DR. PÁL ENDRE PÉCSI TUDOMÁNYEGYETEM Általános Orvostudományi Kar NEUROLÓGIAI KLINIKA 2001 2 Bevezetés A sclerosis multiplex
IMMUNREGULÁCIÓ KÍSÉRLETES ENCEPHALOMYELITISBEN: AZ NKT SEJTEK SZEREPE Ph.D. tézisek DR. PÁL ENDRE PÉCSI TUDOMÁNYEGYETEM Általános Orvostudományi Kar NEUROLÓGIAI KLINIKA 2001 2 Bevezetés A sclerosis multiplex
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
INTRACELLULÁRIS PATOGÉNEK
 INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
Áramlási citometria, sejtszeparációs technikák. Immunológiai és Biotechnológiai Intézet ÁOK, PTE
 Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Doktori értekezés tézisei. A komplementrendszer szerepe EAE-ben (Experimental Autoimmune Encephalomyelitis), a sclerosis multiplex egérmodelljében
, a sclerosis multiplex egérmodelljében") Doktori értekezés tézisei A komplementrendszer szerepe EAE-ben (Experimental Autoimmune Encephalomyelitis), a sclerosis multiplex egérmodelljében Készítette: Terényi Nóra Témavezető: Prof. Erdei Anna Biológia
Doktori értekezés tézisei A komplementrendszer szerepe EAE-ben (Experimental Autoimmune Encephalomyelitis), a sclerosis multiplex egérmodelljében Készítette: Terényi Nóra Témavezető: Prof. Erdei Anna Biológia
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
A preventív vakcináció lényege :
 Vakcináció Célja: antigénspecifkus immunválasz kiváltása a szervezetben A vakcina egy olyan készítmény, amely fokozza az immunitást egy adott betegséggel szemben (aktiválja az immunrendszert). A preventív
Vakcináció Célja: antigénspecifkus immunválasz kiváltása a szervezetben A vakcina egy olyan készítmény, amely fokozza az immunitást egy adott betegséggel szemben (aktiválja az immunrendszert). A preventív
A fiziológiás terhesség hátterében álló immunológiai történések
 A fiziológiás terhesség hátterében álló immunológiai történések APAI Ag ANYAI Ag FERTŐZÉS AUTOIMMUNITÁS MAGZATI ANTIGEN ALACSONY P SZINT INFERTILITAS BEÁGYAZÓDÁS ANYAI IMMUNREGULÁCIÓ TROPHOBLAST INVÁZIÓ
A fiziológiás terhesség hátterében álló immunológiai történések APAI Ag ANYAI Ag FERTŐZÉS AUTOIMMUNITÁS MAGZATI ANTIGEN ALACSONY P SZINT INFERTILITAS BEÁGYAZÓDÁS ANYAI IMMUNREGULÁCIÓ TROPHOBLAST INVÁZIÓ
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
B-sejtek szerepe az RA patológiás folyamataiban
 B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE
 Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Immunológia I. 4. előadás. Kacskovics Imre
 Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia alapjai előadás. A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és
 Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai előadás. Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás.
 Immunológia alapjai 24-25. előadás Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás. Tolerált bőr graftok MHC (H2) azonos egereken TOLERANCIA & AUTOIMMUNITÁS Toleranciáról beszélünk, ha
Immunológia alapjai 24-25. előadás Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás. Tolerált bőr graftok MHC (H2) azonos egereken TOLERANCIA & AUTOIMMUNITÁS Toleranciáról beszélünk, ha
Kutatási beszámoló ( )
") Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 7. előadás Immunizálás. Poliklonális és monoklonális ellenanyag előállítása, tisztítása, alkalmazása Az antigén (haptén + hordozó) sokféle specificitású ellenanyag
Immunológiai módszerek a klinikai kutatásban 7. előadás Immunizálás. Poliklonális és monoklonális ellenanyag előállítása, tisztítása, alkalmazása Az antigén (haptén + hordozó) sokféle specificitású ellenanyag
Újabb ismeretek a Graves-ophthalmopathia kórisméjében
 Újabb ismeretek a Graves-ophthalmopathia kórisméjében Dr. Molnár Ildikó 2004 Tézisek Az utóbbi 11 évben végzett tudományos munkám új eredményei 1. Graves-kórhoz társult infiltratív ophthalmopathia kialakulásában
Újabb ismeretek a Graves-ophthalmopathia kórisméjében Dr. Molnár Ildikó 2004 Tézisek Az utóbbi 11 évben végzett tudományos munkám új eredményei 1. Graves-kórhoz társult infiltratív ophthalmopathia kialakulásában
Az adaptív immunválasz kialakulása. Erdei Anna Immunológiai Tanszék ELTE
 Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Egy, a Curcuma longa-ból izolált poliszacharid kivonat immunserkentı tevékenységei.
 http://www.ncbi.nlm.nih.gov/pubmed/20609432 Egy, a Curcuma longa-ból izolált poliszacharid kivonat immunserkentı tevékenységei. Yue GG 1, Chan BC, Hon PM, Kennelly EJ, Yeung SK, Cassileth BR, Fung KP,
http://www.ncbi.nlm.nih.gov/pubmed/20609432 Egy, a Curcuma longa-ból izolált poliszacharid kivonat immunserkentı tevékenységei. Yue GG 1, Chan BC, Hon PM, Kennelly EJ, Yeung SK, Cassileth BR, Fung KP,
T sejtek II Vizler Csaba 2010
 T sejtek II Vizler Csaba 2010 DENDRITIKUS SEJT Tc CD8+ Th CD4+ B SEJT CTL DTH Ab T sejtek és B sejtek - az elnevezés eredete A T sejt receptor (TcR) kialakulása T sejt érés és szelekció a tímuszban A T
T sejtek II Vizler Csaba 2010 DENDRITIKUS SEJT Tc CD8+ Th CD4+ B SEJT CTL DTH Ab T sejtek és B sejtek - az elnevezés eredete A T sejt receptor (TcR) kialakulása T sejt érés és szelekció a tímuszban A T
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása. Uher Ferenc, PhD, DSc
 A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
Áramlási citometria 2011. / 4. Immunológiai és Biotechnológiai Intézet PTE KK
 Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
Immunológia alapjai előadás MHC. szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
Ha nem akarsz mellé-nyúl-ni, használj Nobivac Myxo-RHD-t! MSDay-MOM park, 2013.02.21. dr. Schweickhardt Eszter
 Ha nem akarsz mellé-nyúl-ni, használj Nobivac Myxo-RHD-t! MSDay-MOM park, 2013.02.21. dr. Schweickhardt Eszter Áttekintés Miért éppen a nyuszik? Védekezés módja Vakcina jellemzői Vakcina működése Ráfertőzési
Ha nem akarsz mellé-nyúl-ni, használj Nobivac Myxo-RHD-t! MSDay-MOM park, 2013.02.21. dr. Schweickhardt Eszter Áttekintés Miért éppen a nyuszik? Védekezés módja Vakcina jellemzői Vakcina működése Ráfertőzési
Sejtfeldolgozás Felhasználás
 Sejtterápia Sejtfeldolgozás Felhasználás Fagyasztva tárolás Sejtmosás Alap sejtszelekció Komplex sejtszelekció Ex vivo sejtszaporítás Sejtaktiválás Immunizálás Génmodifikálás Köldökzsinórvér bank Limfocita
Sejtterápia Sejtfeldolgozás Felhasználás Fagyasztva tárolás Sejtmosás Alap sejtszelekció Komplex sejtszelekció Ex vivo sejtszaporítás Sejtaktiválás Immunizálás Génmodifikálás Köldökzsinórvér bank Limfocita
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői.
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban között. Dr. Hortobágyi Judit
 A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban 2007-2011 között Dr. Hortobágyi Judit Pikkelysömörre gyakorolt hatása 2007-től 2009-ig 1. Lokális hatása 2. Szisztémás hatása 3. Állatkísérlet
A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban 2007-2011 között Dr. Hortobágyi Judit Pikkelysömörre gyakorolt hatása 2007-től 2009-ig 1. Lokális hatása 2. Szisztémás hatása 3. Állatkísérlet
Antigén, Antigén prezentáció
 Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai
 Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
A szervspecifikus autoimmun betegségek pathomechanizmusa. Dr. Bakó Gyula DE OEC III. Belklinika Ph.D. Kurzus, Debrecen, 2011.
 A szervspecifikus autoimmun betegségek pathomechanizmusa Dr. Bakó Gyula DE OEC III. Belklinika Ph.D. Kurzus, Debrecen, 2011. Centrális és perifériás tolerancia Az adaptív immunitás esetén a tolerantia
A szervspecifikus autoimmun betegségek pathomechanizmusa Dr. Bakó Gyula DE OEC III. Belklinika Ph.D. Kurzus, Debrecen, 2011. Centrális és perifériás tolerancia Az adaptív immunitás esetén a tolerantia
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL
 TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai
 Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Immunológia alapjai előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok:
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
A Flowcytometriás. en. Sinkovichné Bak Erzsébet,
 A Flowcytometriás keresztpróba jelentısége élıdonoros veseátültet ltetést megelızıen. en. Sinkovichné Bak Erzsébet, Schmidt Lászlóné Mikor végezhetv gezhetı el a veseátültet ltetés? Veseátültetés s akkor
A Flowcytometriás keresztpróba jelentısége élıdonoros veseátültet ltetést megelızıen. en. Sinkovichné Bak Erzsébet, Schmidt Lászlóné Mikor végezhetv gezhetı el a veseátültet ltetés? Veseátültetés s akkor
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Az ellenanyagok szerkezete és funkciója. Erdei Anna Immunológiai Tanszék ELTE
 Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Zárójelentés. A) A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.
 A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.") Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
http://www.rimm.dote.hu Tumor immunológia
 http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
ANTICANCER RESEARCH [Rákkutatás]
![ANTICANCER RESEARCH [Rákkutatás]](/thumbs/39/19250750.jpg "ANTICANCER RESEARCH [Rákkutatás]") ANTICANCER RESEARCH [Rákkutatás] Nemzetközi rákkutatási és rákgyógyászati folyóirat Az MGN-3/Biobran rizskorpából kivont módosított arabinoxilán in vitro növeli a metasztatikus emlőráksejtek érzékenységét
ANTICANCER RESEARCH [Rákkutatás] Nemzetközi rákkutatási és rákgyógyászati folyóirat Az MGN-3/Biobran rizskorpából kivont módosított arabinoxilán in vitro növeli a metasztatikus emlőráksejtek érzékenységét
Az ophthalmopathia autoimmun kórfolyamatára utaló tényezôk Bizonyított: A celluláris és humorális autoimmun folyamatok szerepe.
 Az ophthalmopathia autoimmun kórfolyamatára utaló tényezôk Bizonyított: A celluláris és humorális autoimmun folyamatok szerepe. szemizom, retrobulbaris kötôszövet, könnymirigy elleni autoantitestek exophthalmogen
Az ophthalmopathia autoimmun kórfolyamatára utaló tényezôk Bizonyított: A celluláris és humorális autoimmun folyamatok szerepe. szemizom, retrobulbaris kötôszövet, könnymirigy elleni autoantitestek exophthalmogen
Immunológia I. 2. előadás. Kacskovics Imre (imre.kacskovics@ttk.elte.hu)
") Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Autoimmun vizsgálatok (ML AU)
") mennyisége Autoimmun vizsgálatok (ML AU) ML AU 1.100 ANA ELISA(QuantaLite) ELISA - Szérum 200 l < 20 U/ml ML AU 1.200 Anti-dsDNA ELISA Szérum 200 l < 20 IU/ml ML AU 1.210 Anti-dsDNA IgG (Alegria) ELISA
mennyisége Autoimmun vizsgálatok (ML AU) ML AU 1.100 ANA ELISA(QuantaLite) ELISA - Szérum 200 l < 20 U/ml ML AU 1.200 Anti-dsDNA ELISA Szérum 200 l < 20 IU/ml ML AU 1.210 Anti-dsDNA IgG (Alegria) ELISA
2.6.16. VIZSGÁLATOK IDEGEN KÓROKOZÓKRA HUMÁN ÉLŐVÍRUS-VAKCINÁKBAN
 2.6.16. Vizsgálatok idegen kórokozókra Ph.Hg.VIII. - Ph.Eur.7.0 1 2.6.16. VIZSGÁLATOK IDEGEN KÓROKOZÓKRA HUMÁN ÉLŐVÍRUS-VAKCINÁKBAN 01/2011:20616 Azokhoz a vizsgálatokhoz, amelyekhez a vírust előzőleg
2.6.16. Vizsgálatok idegen kórokozókra Ph.Hg.VIII. - Ph.Eur.7.0 1 2.6.16. VIZSGÁLATOK IDEGEN KÓROKOZÓKRA HUMÁN ÉLŐVÍRUS-VAKCINÁKBAN 01/2011:20616 Azokhoz a vizsgálatokhoz, amelyekhez a vírust előzőleg
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a PD-L1 és PD-1 fehérjék túlélésre gyakorolt hatása
 Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Az Ames teszt (Salmonella/S9) a nemzetközi hatóságok által a kémiai anyagok minősítéséhez előírt vizsgálat, amellyel az esetleges genotoxikus hatás
 a nemzetközi hatóságok által a kémiai anyagok minősítéséhez előírt vizsgálat, amellyel az esetleges genotoxikus hatás") Az Ames teszt (Salmonella/S9) a nemzetközi hatóságok által a kémiai anyagok minősítéséhez előírt vizsgálat, amellyel az esetleges genotoxikus hatás kockázatát mérik fel. Annak érdekében, hogy az anyavegyületével
Az Ames teszt (Salmonella/S9) a nemzetközi hatóságok által a kémiai anyagok minősítéséhez előírt vizsgálat, amellyel az esetleges genotoxikus hatás kockázatát mérik fel. Annak érdekében, hogy az anyavegyületével
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Intézeti Beszámoló. Dr. Kovács Árpád Ferenc
 Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok.
 Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
T helper és T citotoxikus limfociták szerepének vizsgálata allergológiai és autoimmun bőrgyógyászati kórképekben
 T helper és T citotoxikus limfociták szerepének vizsgálata allergológiai és autoimmun bőrgyógyászati kórképekben A most befejezett OTKA pályázat célkitűzése volt immunmediált bőrgyógyászati ill. bőrgyógyászati
T helper és T citotoxikus limfociták szerepének vizsgálata allergológiai és autoimmun bőrgyógyászati kórképekben A most befejezett OTKA pályázat célkitűzése volt immunmediált bőrgyógyászati ill. bőrgyógyászati
Immunológia 4. A BCR diverzitás kialakulása
 Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Rövid és hosszútávú sejtkultúrák, funkcionális tesztek. Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem
 Rövid és hosszútávú sejtkultúrák, funkcionális tesztek Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem Bevezetés Definíció: az élő szervezeten kívül (in( vitro) ) tartunk élő sejteket Cél:
Rövid és hosszútávú sejtkultúrák, funkcionális tesztek Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem Bevezetés Definíció: az élő szervezeten kívül (in( vitro) ) tartunk élő sejteket Cél:
Az áramlási citometria gyakorlati alkalmazása az ondó rutin analízisben. Hajnal Ágnes, Dr Mikus Endre, Dr Venekeiné Losonczi Olga
 Az áramlási citometria gyakorlati alkalmazása az ondó rutin analízisben Hajnal Ágnes, Dr Mikus Endre, Dr Venekeiné Losonczi Olga Labmagister Kft aktivitásai HumanCell SynLab Diagnosztika MensMentis LabMagister
Az áramlási citometria gyakorlati alkalmazása az ondó rutin analízisben Hajnal Ágnes, Dr Mikus Endre, Dr Venekeiné Losonczi Olga Labmagister Kft aktivitásai HumanCell SynLab Diagnosztika MensMentis LabMagister
Immunpatológia kurzus, - tavaszi szemeszter
 Immunpatológia kurzus, - tavaszi szemeszter Prof. Sármay Gabriella, Dr. Bajtay Zsuzsa, Dr. Józsi Mihály, Prof. Kacskovics Imre Prof. Erdei Anna Szerdánként, 10.00-12.00-ig, 5-202-es terem 1 2016. 02. 17.
Immunpatológia kurzus, - tavaszi szemeszter Prof. Sármay Gabriella, Dr. Bajtay Zsuzsa, Dr. Józsi Mihály, Prof. Kacskovics Imre Prof. Erdei Anna Szerdánként, 10.00-12.00-ig, 5-202-es terem 1 2016. 02. 17.
Mikrobiális antigének
 Mikrobiális antigének Dr. Pusztai Rozália SZTE, ÁOK, Orvosi Mikrobiológiai és Immunbiológiai Intézet 2008. november 17. Antigének Konvencionális antigének Superantigének Antigén - az érett immunrendszer
Mikrobiális antigének Dr. Pusztai Rozália SZTE, ÁOK, Orvosi Mikrobiológiai és Immunbiológiai Intézet 2008. november 17. Antigének Konvencionális antigének Superantigének Antigén - az érett immunrendszer
Dr. Nemes Nagy Zsuzsa Szakképzés Karl Landsteiner Karl Landsteiner:
 Az AB0 vércsoport rendszer Dr. Nemes Nagy Zsuzsa Szakképzés 2011 Az AB0 rendszer felfedezése 1901. Karl Landsteiner Landsteiner szabály 1901 Karl Landsteiner: Munkatársai vérmintáit vizsgálva fedezte fel
Az AB0 vércsoport rendszer Dr. Nemes Nagy Zsuzsa Szakképzés 2011 Az AB0 rendszer felfedezése 1901. Karl Landsteiner Landsteiner szabály 1901 Karl Landsteiner: Munkatársai vérmintáit vizsgálva fedezte fel
Részletes szakmai beszámoló Az erbb proteinek asszociációjának kvantitatív jellemzése című OTKA pályázatról (F049025)
") Részletes szakmai beszámoló Az erbb proteinek asszociációjának kvantitatív jellemzése című OTKA pályázatról (F049025) A pályázat megvalósítása során célunk volt egyrészt a molekulák asszociációjának tanulmányozására
Részletes szakmai beszámoló Az erbb proteinek asszociációjának kvantitatív jellemzése című OTKA pályázatról (F049025) A pályázat megvalósítása során célunk volt egyrészt a molekulák asszociációjának tanulmányozására
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek
 A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
4. A humorális immunválasz október 12.
 4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
Immunológia alapjai előadás. Sej-sejt kommunikációk az immunválaszban.
 Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
A plazminogén metilglioxál módosítása csökkenti a fibrinolízis hatékonyságát. Léránt István, Kolev Kraszimir, Gombás Judit és Machovich Raymund
 A plazminogén metilglioxál módosítása csökkenti a fibrinolízis hatékonyságát Léránt István, Kolev Kraszimir, Gombás Judit és Machovich Raymund Semmelweis Egyetem Orvosi Biokémia Intézet, Budapest Fehérjék
A plazminogén metilglioxál módosítása csökkenti a fibrinolízis hatékonyságát Léránt István, Kolev Kraszimir, Gombás Judit és Machovich Raymund Semmelweis Egyetem Orvosi Biokémia Intézet, Budapest Fehérjék
A krónikus hepatitis C vírus infekcióhoz társuló elégtelen celluláris immunválasz pathogenezise
 Szakmai zárójelentés dr Pár Gabriella A krónikus hepatitis C vírus infekcióhoz társuló elégtelen celluláris immunválasz pathogenezise A hepatitis C virus (HCV) nagy közegészségügyi probléma. A világon
Szakmai zárójelentés dr Pár Gabriella A krónikus hepatitis C vírus infekcióhoz társuló elégtelen celluláris immunválasz pathogenezise A hepatitis C virus (HCV) nagy közegészségügyi probléma. A világon
Mindennapi pajzsmirigy diagnosztika: antitestek, hormonszintek
 Mindennapi pajzsmirigy diagnosztika: antitestek, hormonszintek Nagy V. Endre Endokrinológia Tanszék Debreceni Egyetem Orvos- és Egészségtudományi Centrum Belgyógyászati Intézet, I. Belklinika A pajzsmirigybetegségek
Mindennapi pajzsmirigy diagnosztika: antitestek, hormonszintek Nagy V. Endre Endokrinológia Tanszék Debreceni Egyetem Orvos- és Egészségtudományi Centrum Belgyógyászati Intézet, I. Belklinika A pajzsmirigybetegségek
Bevezetés a vércsoport-szerológiai vizsgálatokba
 Bevezetés a vércsoport-szerológiai vizsgálatokba dr. Fődi Éva OVSZ Szegedi Regionális Vérellátó Központ 2018. január Mivel foglalkozik a vércsoportszerológia? Vércsoport szerológiai kompatibilitás: Kompatibilitás:
Bevezetés a vércsoport-szerológiai vizsgálatokba dr. Fődi Éva OVSZ Szegedi Regionális Vérellátó Központ 2018. január Mivel foglalkozik a vércsoportszerológia? Vércsoport szerológiai kompatibilitás: Kompatibilitás:
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-8/1/A-29-11 Az orvosi biotechnológiai
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-8/1/A-29-11 Az orvosi biotechnológiai
AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA. Balicza-Himer Leonóra. Doktori értekezés tézisei
 AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA Doktori értekezés tézisei Balicza-Himer Leonóra Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola / Immunológia
AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA Doktori értekezés tézisei Balicza-Himer Leonóra Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola / Immunológia
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata
 sejtek mikro RNS (mir) profiljának vizsgálata") Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Rh VÉRCSOPORT RENDSZER GENETIKÁJA. Rh ANTIGÉNEK ÉS ANTITESTEK. EGYÉB VÉRCSOPORTRENDSZEREK
 Rh VÉRCSOPORT RENDSZER GENETIKÁJA. Rh ANTIGÉNEK ÉS ANTITESTEK. EGYÉB VÉRCSOPORTRENDSZEREK HISTORY Antitestet találtak egy koraszülött gyermek anyjának szérumában; ez lenne felelős a gyermek haláláért?
Rh VÉRCSOPORT RENDSZER GENETIKÁJA. Rh ANTIGÉNEK ÉS ANTITESTEK. EGYÉB VÉRCSOPORTRENDSZEREK HISTORY Antitestet találtak egy koraszülött gyermek anyjának szérumában; ez lenne felelős a gyermek haláláért?
Immunológia alapjai. 23-24. előadás. Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás.
 Immunológia alapjai 23-24. előadás Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás. Tolerált bőr graftok MHC (H2) azonos egereken TOLERANCIA & AUTOIMMUNITÁS Toleranciáról beszélünk, ha
Immunológia alapjai 23-24. előadás Immunológiai tolerancia. Fiziológiás és patológiás autoimmunitás. Tolerált bőr graftok MHC (H2) azonos egereken TOLERANCIA & AUTOIMMUNITÁS Toleranciáról beszélünk, ha
Mit tud a genetika. Génterápiás lehetőségek MPS-ben. Dr. Varga Norbert
 Mit tud a genetika Génterápiás lehetőségek MPS-ben Dr. Varga Norbert Oki terápia Terápiás lehetőségek MPS-ben A kiváltó okot gyógyítja meg ERT Enzimpótló kezelés Őssejt transzplantáció Genetikai beavatkozások
Mit tud a genetika Génterápiás lehetőségek MPS-ben Dr. Varga Norbert Oki terápia Terápiás lehetőségek MPS-ben A kiváltó okot gyógyítja meg ERT Enzimpótló kezelés Őssejt transzplantáció Genetikai beavatkozások
A PET szerepe a gyógyszerfejlesztésben. Berecz Roland DE KK Pszichiátriai Tanszék
 A PET szerepe a gyógyszerfejlesztésben Berecz Roland DE KK Pszichiátriai Tanszék Gyógyszerfejlesztés Felfedezés gyógyszertár : 10-15 év Kb. 1 millárd USD/gyógyszer (beleszámolva a sikertelen fejlesztéseket)
A PET szerepe a gyógyszerfejlesztésben Berecz Roland DE KK Pszichiátriai Tanszék Gyógyszerfejlesztés Felfedezés gyógyszertár : 10-15 év Kb. 1 millárd USD/gyógyszer (beleszámolva a sikertelen fejlesztéseket)
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
A CYTOKIN AKTIVÁCIÓ ÉS GÉN-POLIMORFIZMUSOK VIZSGÁLATA HEL1COBACTER PYLORI FERTŐZÉSBEN ÉS CROHN BETEGSÉGBEN
 % A CYTOKIN AKTIVÁCIÓ ÉS GÉN-POLIMORFIZMUSOK VIZSGÁLATA HEL1COBACTER PYLORI FERTŐZÉSBEN ÉS CROHN BETEGSÉGBEN BEVEZETÉS Mind a Helicobacter pylori fertőzésre, mind pedig a gyulladásos bélbetegségekre, mint
% A CYTOKIN AKTIVÁCIÓ ÉS GÉN-POLIMORFIZMUSOK VIZSGÁLATA HEL1COBACTER PYLORI FERTŐZÉSBEN ÉS CROHN BETEGSÉGBEN BEVEZETÉS Mind a Helicobacter pylori fertőzésre, mind pedig a gyulladásos bélbetegségekre, mint
1. Bevezetés. Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban
 Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban Doktori tézisek Dr. Jakus Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori
Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban Doktori tézisek Dr. Jakus Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori
Túlérzékenységi reakciók Gell és Coombs felosztása szerint.
 Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI. - autokrin. -neurokrin. - parakrin. -térátvitel. - endokrin
 A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
KLINIKAI IMMUNOLÓGIA I.
 Kórházhigienikus képzés, DE OEC KLINIKAI IMMUNOLÓGIA I. AZ IMMUNRENDSZER MŰKÖDÉSE Dr. Sipka Sándor DE OEC III. sz. Belgyógyászati Klinika Regionális Immunológiai Laboratórium A főbb ábrák és táblázatok
Kórházhigienikus képzés, DE OEC KLINIKAI IMMUNOLÓGIA I. AZ IMMUNRENDSZER MŰKÖDÉSE Dr. Sipka Sándor DE OEC III. sz. Belgyógyászati Klinika Regionális Immunológiai Laboratórium A főbb ábrák és táblázatok
FUSARIUM TOXINOK IDEGRENDSZERI HATÁSÁNAK ELEMZÉSE
 FUSARIUM TOXINOK IDEGRENDSZERI HATÁSÁNAK ELEMZÉSE Világi Ildikó, Varró Petra, Bódi Vera, Schlett Katalin, Szűcs Attila, Rátkai Erika Anikó, Szentgyörgyi Viktória, Détári László, Tóth Attila, Hajnik Tünde,
FUSARIUM TOXINOK IDEGRENDSZERI HATÁSÁNAK ELEMZÉSE Világi Ildikó, Varró Petra, Bódi Vera, Schlett Katalin, Szűcs Attila, Rátkai Erika Anikó, Szentgyörgyi Viktória, Détári László, Tóth Attila, Hajnik Tünde,
CANDIDA FERTŐZÉSSEL SZEMBENI IMMUNITÁS AZ I-ES TÍPUSÚ AUTOIMMUN POLIENDOKRIN SZINDRÓMÁS BETEGEKBEN
 Egyetemi doktori (PhD) értekezés tézisei CANDIDA FERTŐZÉSSEL SZEMBENI IMMUNITÁS AZ I-ES TÍPUSÚ AUTOIMMUN POLIENDOKRIN SZINDRÓMÁS BETEGEKBEN Dr. Sarkadi Adrien Katalin Témavezető: Dr. Erdős Melinda DEBRECENI
Egyetemi doktori (PhD) értekezés tézisei CANDIDA FERTŐZÉSSEL SZEMBENI IMMUNITÁS AZ I-ES TÍPUSÚ AUTOIMMUN POLIENDOKRIN SZINDRÓMÁS BETEGEKBEN Dr. Sarkadi Adrien Katalin Témavezető: Dr. Erdős Melinda DEBRECENI
Terhesség és immunitás 2012.03.02. Immunhematológiai terhesgondozás ÚHB Újszülöttek transzfúziója
 Egészségügyi szakdolgozók transzfúziológiai továbbképzése 2012. március 5-9. Immunhematológiai terhesgondozás ÚHB Újszülöttek transzfúziója Dr. Nemes Nagy Zsuzsa Országos Vérellátó Szolgálat Terhesség
Egészségügyi szakdolgozók transzfúziológiai továbbképzése 2012. március 5-9. Immunhematológiai terhesgondozás ÚHB Újszülöttek transzfúziója Dr. Nemes Nagy Zsuzsa Országos Vérellátó Szolgálat Terhesség
Monoklonális antitestek előállítása, jellemzői
 Monoklonális antitestek előállítása, jellemzői 2011. / 3 Immunológiai és Biotechnológiai Intézet PTE KK Antitestek szerkezete IgG Monoklonális ellenanyag előállítás: Hybridoma technika Lehetővé teszi kezdetben
Monoklonális antitestek előállítása, jellemzői 2011. / 3 Immunológiai és Biotechnológiai Intézet PTE KK Antitestek szerkezete IgG Monoklonális ellenanyag előállítás: Hybridoma technika Lehetővé teszi kezdetben
Immunológia II GY. 1. Bevezetés febr. 16. Bajtay Zsuzsa
 Immunológia II GY 1. Bevezetés 2018. febr. 16. Bajtay Zsuzsa Tankönyv: Immunológiai módszerek szerkesztette: Erdei Anna (Medicina, 2006) Laboratóriumi gyakorlatok MSc MIM: - halim1mb17lm Haladó immunológiai
Immunológia II GY 1. Bevezetés 2018. febr. 16. Bajtay Zsuzsa Tankönyv: Immunológiai módszerek szerkesztette: Erdei Anna (Medicina, 2006) Laboratóriumi gyakorlatok MSc MIM: - halim1mb17lm Haladó immunológiai
LIQUORVIZSGÁLAT. A lumbálpunkció helye a klinikai neurodiagnosztikában. Tantermi előadás V.évf. 2014. szeptember 24. Bors László Neurológiai Klinika
 LIQUORVIZSGÁLAT A lumbálpunkció helye a klinikai neurodiagnosztikában Tantermi előadás V.évf. 2014. szeptember 24. Bors László Neurológiai Klinika Összefoglalás 1. A liquor termelődése, keringése, felszívódása
LIQUORVIZSGÁLAT A lumbálpunkció helye a klinikai neurodiagnosztikában Tantermi előadás V.évf. 2014. szeptember 24. Bors László Neurológiai Klinika Összefoglalás 1. A liquor termelődése, keringése, felszívódása
Rovarméreg (méh, darázs) - allergia
 - allergia") Rovarméreg (méh, darázs) - allergia Herjavecz Irén Országos Korányi TBC és Pulmonológiai Intézet Epidemiológia I. Prevalencia: - nagy helyi reakció: felnőtt 10-15 % - szisztémás reakció: gyerek 0.4-0.8
Rovarméreg (méh, darázs) - allergia Herjavecz Irén Országos Korányi TBC és Pulmonológiai Intézet Epidemiológia I. Prevalencia: - nagy helyi reakció: felnőtt 10-15 % - szisztémás reakció: gyerek 0.4-0.8
Új könnyűlánc diagnosztika. Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006
 Új könnyűlánc diagnosztika Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006 1845 Bence Jones Protein vizelet fehérje 1922 BJP I-II típus 1956 BJP
Új könnyűlánc diagnosztika Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006 1845 Bence Jones Protein vizelet fehérje 1922 BJP I-II típus 1956 BJP
Immunitás és evolúció
 Immunitás és evolúció (r)evolúció az immunrendszerben Az immunrendszer evolúciója Müller Viktor ELTE Növényrendszertani, Ökológiai és Elméleti Biológiai Tanszék http://ramet.elte.hu/~viktor Az immunitás
Immunitás és evolúció (r)evolúció az immunrendszerben Az immunrendszer evolúciója Müller Viktor ELTE Növényrendszertani, Ökológiai és Elméleti Biológiai Tanszék http://ramet.elte.hu/~viktor Az immunitás
Esettanulmányok. Dr. Toldi József OVSZ Szegedi Regionális Vérellátó Központ
 Esettanulmányok Dr. Toldi József OVSZ Szegedi Regionális Vérellátó Központ Esetismertetések: ABO vércsoport meghatározással kapcsolatos nehézségek és megoldások RhD bizonytalanság kivizsgálása Antitest-azonosítással
Esettanulmányok Dr. Toldi József OVSZ Szegedi Regionális Vérellátó Központ Esetismertetések: ABO vércsoport meghatározással kapcsolatos nehézségek és megoldások RhD bizonytalanság kivizsgálása Antitest-azonosítással
eljárásokkal Doktori tézisek Szatmári Tünde Semmelweis Egyetem Klinikai Orvostudományok Doktori Iskola Sugárterápia Program
 Agydaganatok sugárterápia iránti érzékenységének növelése génterápiás eljárásokkal Doktori tézisek Szatmári Tünde Semmelweis Egyetem Klinikai Orvostudományok Doktori Iskola Sugárterápia Program Doktori
Agydaganatok sugárterápia iránti érzékenységének növelése génterápiás eljárásokkal Doktori tézisek Szatmári Tünde Semmelweis Egyetem Klinikai Orvostudományok Doktori Iskola Sugárterápia Program Doktori
Kompatibilitás. Type & Screen
 Kompatibilitás Type & Screen Dr. Toldi József OVSZ Szegedi Regionális Vérellátó Központ A kompatibilitási vizsgálatok célja nil nocere elv ( a transzfúziós szövődmény megelőzése) a donor vörösvérsejtek
Kompatibilitás Type & Screen Dr. Toldi József OVSZ Szegedi Regionális Vérellátó Központ A kompatibilitási vizsgálatok célja nil nocere elv ( a transzfúziós szövődmény megelőzése) a donor vörösvérsejtek
Immunológia alapjai
 Immunológia alapjai 2011.11.03. A sejt-mediálta immunválasz effektor mechanizmusai (CMI): 1. Citotoxicitás 2. T H sejt mediálta makrofág aktiváció (Késői típusú hyperszenzitivitás = DTH.) Az adaptív immunválasz
Immunológia alapjai 2011.11.03. A sejt-mediálta immunválasz effektor mechanizmusai (CMI): 1. Citotoxicitás 2. T H sejt mediálta makrofág aktiváció (Késői típusú hyperszenzitivitás = DTH.) Az adaptív immunválasz
Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése.
 F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban
F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban