Doktori (PhD) értekezés
|
|
|
- Csongor Bakos
- 6 évvel ezelőtt
- Látták:
Átírás

1 Cigarettafüst-indukálta gyulladásos folyamatok COPD-ben szisztémás gyulladás kialakulása a Wnt5a és a gyulladásos citokin tartalmú extracelluláris vezikulák által Doktori (PhD) értekezés FELLER DIÁNA Pécsi Tudományegyetem Gyógyszerésztudományi Kar, Általános Orvostudományi Kar Gyógyszerészi Biotechnológia Intézet, Farmakológiai és Farmakoterápiai Intézet Elméleti Orvostudományok Doktori Iskola Témavezetők: Prof. Dr. Pongrácz Judit Erzsébet, egyetemi tanár Prof. Dr. Helyes Zsuzsanna, egyetemi tanár Programvezető: Prof. Dr. Berki Tímea, egyetemi tanár Doktori Iskola vezetője: Prof. Dr. Szekeres-Barthó Júlia, egyetemi tanár Pécsi Tudományegyetem Általános Orvostudományi Kar Pécs, 2019
2 Rövidítések jegyzéke AC: adenocarcinoma (adenokarcinóma) ANKRD1: ankyrin repeat domain 1 (ismétlődő ankirin domén 1) AP1: activator protein 1 (aktivátor protein 1) APC: adenomatous polyposis coli (adenomatózus polipózis kóli) ARDS: acute respiratory distress syndrome (akut respirációs distressz szindróma) AQP5: aquaporin-5 (akvaporin-5) ATI cells/ati-es sejtek: alveolar type I epithelial cells (I. típusú alveoláris epitél sejtek) ATII cells/atii-es sejtek: alveolar type II epithelial cells (II. típusú alveoláris epitél sejtek) BAL: bronchoalveolar lavage (bronchoalveoláris mosás) Bim: Bcl-2-like protein 11 (Bcl-2 kölcsönhatást közvetítő fehérje/bcl-2 típusú fehérje 11) CAMKII: Ca2+/calmodulin-dependent protein kinase II (Ca 2+ /Kalmodulin függő protein kináz II) CBA: cytometric bead array (citometriás gyöngy array) CS/CF: cigarette smoke (cigarettafüst) CSE/CFE: cigarette smoke extract (cigarettafüst extraktum) COPD: chronic obstructive pulmonary disease (krónikus obstruktív tüdőbetegség) CRE: camp response element (camp-ra reagáló elem) CREB: camp response element-binding protein (CRE-kötő fehérje) CRP: C-reactive protein (C-reaktív fehérje) CYP4B1: cytochrome P450 family 4 subfamily B member 1 (citokróm P450 család 4, alcsalád B, polipeptid 1) DKK-1: Dickkopf -1 (Dickkopf -1) DVL: Dishevelled (Dishevelled) EGM-2: endothelial growth medium-2 (endotél növekedési médium) EMP2: epithelial membrane protein 2 (epiteliális membrán fehérje 2) EpCAM: epithelial cell adhesion molecule (epiteliális sejtadhéziós molekula) EMT: epithelial mesenchymal transition (epiteliális mezenchimális átalakulás) EV: extracellular vesicles (extracelluláris vezikulák) FACS: fluorescence-activated cell sorting (fluoreszcencia aktivált sejtválogatás) FBS: fetal bovine serum (borjú szérum) FEV1: forced expiratory volume-one second (első másodpercre eső erőltetett kilégzési térfogat) 2
3 FGM-2: fibroblast growth medium-2 (fibroblaszt növekedési médium) FITC: fluorescein isothiocyanate (fluoreszcens izotiocianát) FVC: forced vital capacity (erőltetett kilégzési kapacitás) FZD: Frizzled (Frizzled) GM-CSF: granulocyte-macrophage colony-stimulating factor (granulocita-makrofág kolónia stimuláló faktor) GOLD: nemzetközi COPD ajánlás (Global Initiative for COPD) GSK-3β: glycogen synthase kinase 3 beta (glikogén szintetáz kináz 3 béta) HDAC: histone deacetylase (hiszton deacetiláz enzim) HMVEC-L: human lung microvascular endothelial cells (humán mikrovaszkuláris tüdő endotélsejt) ITGAV: integrin subunit alpha V (integrin alfa V) IKK: I kappa B kinase (I kappa B kináz) LRP5/6: low-density lipoprotein receptor-related protein 5/6 MACS: magnetic-activated cell sorting (mágneses aktivált sejtválogatás) MAPK: mitogen activated protein kinase (mitogén aktivált protein kináz) mirna: microrna (mikrorns) MMP: matrix metalloproteinase (mátrix metalloproteináz) MV: microvesicle (mikrovezikulum) MVB: multivesicular body (multivezikuláris test) NFAT: nuclear factor of activated T-cells (aktivált T-sejtek nukleáris faktora) NFκB: nuclear factor kappa-light-chain-enhancer of activated B cells (nukleáris faktor kappa B) NHLF: normal human lung fibroblast (normál humán tüdő fibroblaszt) PBMC: peripheral blood mononuclear cell (perifériás vér mononukleáris sejtek) PBS: phosphate-buffered saline (foszfát puffer) PCP: planar cell polarity (poláris sejt polaritás) PE: phycoerythrin (fikoeritrin) PPAR-γ: peroxisome proliferator-activated receptor γ (peroxiszóma proliferátor aktivált receptor γ) RAGE: receptor for advanced glycation endproducts (vég-glikációs termék receptor) RSG: rosiglitazone (roziglitazon) SFP: surfactant protein (szörfaktáns fehérje) SFTPC: surfactant protein-c (szörfaktáns protein-c) 3
4 SCC: squamous cell carcinoma (laphámkarcinóma) TGM2: transglutaminase 2 (transzglutamináz-2) THBS1: trombospondin 1 (trombospondin 1) TD: transdifferentiation (transzdifferenciáció) WHO: World Health Organization (Egészségügyi Világszervezet) 4
5 Tartalom 1. Bevezetés A tüdő gyulladásos megbetegedései és a dohányzás kapcsolata COPD COPD epidemiológiája COPD tünetei és azok hátterében álló molekuláris mechanizmusok COPD diagnosztikája COPD terápiája Wnt jelátvitel Wnt jelátvitel jellemzői és szerepe Kanonikus jelátvitel Nem-kanonikus jelátvitel Wnt5a és PPAR-γ szerepe a gyulladásos folyamatokban Wnt5a gyulladást indukáló szerepe Peroxiszóma proliferátor-aktivált receptor γ (PPAR-γ) gyulladáscsökkentő szerepe Extracelluláris vezikulák szerepe a krónikus gyulladásban Extracelluláris vezikulák definíciója és szerepe Az exoszómák definíciója és szerepe Tüdő regenerációs folyamatai és a Wnt jelátvitel kapcsolata ATI és ATII típusú sejtek szerepe a tüdőben és a transzdifferenciációban Wnt jelátvitel szerepe a tüdő regenerációs folyamataiban Célkitűzés Anyagok és módszerek Etikai engedély Anyagok Humán vérminták gyűjtése Kísérleti állatok In vivo cigarettafüst (CF) expozíció Egértüdő sejtek izolálása Sejtszortolás Cigarettafüst extraktum (CFE) elkészítése Sejtvonalak és primer sejtkultúrák Primer humán monociták izolálása
6 dimenziós (3D) humán tüdőmodell RNS izolálás, cdns szintézis, valós idejű kvantitatív PCR (qpcr) TaqMan Array Immunhisztokémiai és immunfluoreszcens festések Áramlási citometriás gyöngy array (CBA) Extracelluláris vezikulák izolálása humán szérum mintákból Extracelluláris vezikulák (EV) izolálása A549 és Wnt5a-A549 sejtvonalakból Mikrovezikula/Exoszóma homing Exoszómák lokalizációjának fluoreszcens analízise Mikrovezikulák (exoszómák) elektronmikroszkópos detektálása Enzim kapcsolt szendvics immunoszorbens vizsgálat (ELISA) Tüdő epiteliális sejtek izolálása primer humán tüdőszövetből Primer tüdő epiteliális sejtek in vitro transzdifferenciációja (TD) Mikroarray analízis Protein ANalysis THrough Evolutionary Relationships (PANTHER) Mesterséges neuronális hálózat analízis (ANN - artificial neural network) Statisztikai analízisek Eredmények A dohányzás kiváltotta gyulladásos folyamatok és a Wnt5a kapcsolata in vivo egérmodellben A dohányoztatás kiváltotta gyulladás és a Wnt5a kapcsolata 3D humán tüdőmodellben A makrofágok polarizációjának és a Wnt jelátviteli útvonalnak a kapcsolata in vitro dohányoztatás hatására Az extracelluláris vezikulák és a gyulladásos citokinek szerepe a szisztémás gyulladás kialakulásában Wnt jelátvitel aktiválódása az ATII-ATI transzdifferenciáció során in vitro Az ATII-ATI TD-ban szerepet játszó Wnt jelátviteli molekulák hierarchiájának definiálása a mikroarray adatok ANN analízisével Megbeszélés Új eredmények, összefoglalás Hivatkozások Köszönetnyilvánítás
7 1. Bevezetés 1.1. A tüdő gyulladásos megbetegedései és a dohányzás kapcsolata Az 1960-as években írták le először, hogy a dohányzás által kiváltott betegségek gyakoribb előfordulását részben a cigarettafüstnek az immunrendszerre gyakorolt hatása okozza (1). A dohányfüst krónikus inhalációjának következménye az immunológiai funkciók változása, beleértve a veleszületett és a szerzett immunitást egyaránt. A dohányzás az immunrendszer szabályozásában kettős szerepet játszik, egyrészt a kóros immunválaszok súlyosbodását okozza, másrészt az immunrendszer csökkent működését eredményezi. A dohányfüst az immunitásban szerepet játszó immunsejtek fejlődésében és azok termelődésében indukál változásokat. Ezen változások a veleszületett immunrendszer esetében a dendritikus sejteket, makrofágokat, NK-sejteket, míg a szerzett immunrendszer során pedig a citotoxikus CD8 + T- sejteket, helper CD4 + Th-sejteket, regulatorikus T-sejteket és a B-sejteket érintik (2) (3) (4). A dohányzás okozta leggyakoribb megbetegedések közül első helyen állnak a kardiovaszkuláris rendellenességek. Ezt követik a daganatos megbetegedések, melyekből első helyen a tüdődaganatok állnak, majd további légzőszervi rendellenességek, úgymint a krónikus obstruktív tüdőbetegség (COPD - Chronic Obstructive Pulmonary Disease) (5) (1. ábra). Kutatások bizonyítják, hogy a tüdő gyulladásos megbetegedéseiért úgy az aktív, mint a passzív dohányzás felelős, melynek hátterében egyrészt az immunrendszer kóros aktiválása, másrészt védekező mechanizmusok szupressziója állhat (6). A cigarettában található dohány elégetésével több, mint 8000 olyan komponens szabadul fel, melyek immunmoduláló toxinok révén gyulladást váltanak ki. Az égéstermékek főbb összetevői közé tartozik a szén-monoxid, az akrolein, a nikotin és a reaktív oxigén származékok (ROS). Emellett további 73 olyan összetevőt is tartalmaz, melyek karcinogén tulajdonsággal rendelkeznek, mint pl. a kadmium, az arzén és a formaldehid (7). A tüdő gyulladásos megbetegedéseinek időbeli lefolyását tekintve megkülönböztetjük az akut és a krónikus gyulladásos elváltozásokat. Az akut gyulladásos folyamatok a tüdőgyulladásban és az akut légzési elégtelenségben (ARDS - Acute Respiratory Distress Syndrome) jellemzőek, ezzel szemben a krónikus gyulladás asztmában és COPD-ben jelenik meg (8). A dohányzáson kívül a tüdőben kialakuló gyulladásos folyamatokért különféle patogének, toxinok, ipari szennyezőanyagok, allergének is felelősek lehetnek. 7
(10).")
8 COPD-ben és tüdődaganatban szenvedő betegek egyaránt csökkent tüdőfunkcióval és krónikus gyulladás kialakulásával jellemezhetőek. A tüdőrákos esetek 50-70%-ban COPD-vel társulnak, mely utalhat a két kórkép közötti szoros összefüggésre (9)(10). Legújabb tanulmányok szerint a krónikus gyulladás központi szerepet játszik ezen kórképek patogenezisében. A krónikus gyulladás mellett megfigyelhetőek immunológiai diszfunkciók, mint az IL-1β és TNF-α termelődésének hatására a nukleáris faktor kappa B (NFκB - Nuclear Factor Kappa-Light- Chain-Enhancer of Activated B cells) transzkripciós faktor rendellenes aktivációja, epiteliálismezenchimális átalakulás (EMT - Epithelial Mesenchymal Transition), megváltozott adhéziós jelátviteli útvonalak, vagy az extracelluláris mátrix lebomlása is (11). Felső légúti és emésztőszervi daganatok 4% Tüdőrák 21% Egyéb 6% Szív és érrendszeri betegségek 41% Egyéb daganatos betegségek 9% COPD 13% Egyéb légzőszervi betegségek 6% 1. ábra: Dohányzás okozta megbetegedések és azok előfordulási aránya. 8
 becslése szerint 2030-ra a harmadik vezető halálok lesz a világon (12). Kialakulásáért a tüdőt érő szövetkárosító gázok és részecskék lehetnek felelősek, mint pl.")
9 1.2. COPD COPD epidemiológiája Napjainkban a COPD az ötödik vezető morbiditású és mortalitású betegségek közé tartozik világszerte. Előfordulása jelenleg is folyamatosan növekszik. A WHO (World Health Organization) becslése szerint 2030-ra a harmadik vezető halálok lesz a világon (12). Kialakulásáért a tüdőt érő szövetkárosító gázok és részecskék lehetnek felelősek, mint pl. a légköri szennyezés, foglalkozási inhalációs károsító anyagok, fertőzések, genetikai prediszpozíció és az öregedés is. A leggyakoribb kiváltó oknak azonban a dohányzást tartják, amelyet az az adat is alátámaszt, hogy a COPD-ben szenvedő betegek 90-95%-a dohányzik, a dohányzók 15%-nál pedig tüdőrák alakul ki. A betegség kialakulását tekintve a legveszélyeztetettebb csoport a dohányzó, 40 éves kor feletti korosztály, de a COPD súlyosságának szempontjából prognosztikai jelentőséggel bír a dohányzás kezdete (életkor) és intenzitása (cigaretta doboz/év), amelyek a mortalitást is meghatározzák (13) (2. ábra). 2. ábra: A COPD előfordulásának gyakorisága összefüggésben az életkorral és a dohányzás mértékével (14). 9
10 COPD tünetei és azok hátterében álló molekuláris mechanizmusok A COPD a tüdőben zajló, szöveti destrukcióval és légúti obstrukcióval járó betegség, melyet a légúti áramlás ellenállás-fokozódása jellemez. Morfológiailag a COPD-beteg tüdejében rendszerint az emfizéma (alveoláris destrukció) és a krónikus bronchitis (bronchiolusok falvastagságának átmérője kevesebb, mint 2 mm) tüdőbetegségek kombinációja ismerhető fel (15). A betegség tünetei közé tartozik a krónikus, hosszan fennálló produktív köhögés, fokozott nyáktermelés, köpetürítés, nehézlégzés és a fizikai terhelésre jelentkező légszomj (16). A tünetek hátterében álló egyik jellegzetes folyamat a tüdő krónikus gyulladásos folyamata, mely progresszív, visszafordíthatatlan légúti obstrukciót eredményez. Ismert, hogy a COPD progressziójában szerepet játszó dohányfüst mediálta gyulladásos válasz egy rendkívül komplex hálózaton keresztül, különböző immunsejtek, molekuláris mediátorok és a tüdőszövetben található további sejtek aktív részvételével és azok felhalmozódásával indukálódik (17). A dohányzás következtében és COPD során akkumulálódnak makrofágok, neutrofilek, dendritikus sejtek és T-limfociták, míg a gyulladásos mediátorok közül a citokinek, kemokinek és proteázok, mint pl. a mátrix metalloproteázok (MMP) játszanak fontos szerepet. Emellett a tüdőt felépítő epiteliális sejtek is képesek lehetnek az immunfolyamatok befolyásolására. A fent említett résztvevők mindegyike aktiválódhat cigarettafüst hatására is (18). Funkciójuk egyenként jól ismert, de a progresszív, dinamikus és többfázisú gyulladásos COPD kialakulásában résztvevő immunsejtek és mediátorok interakcióját kevésbé ismerjük. Korábbi eredmények alapján elmondhatjuk, hogy a COPD-betegekben az IL-6, IL-8, a C- reaktív protein (CRP) és a fibrinogén szintje szignifikánsan emelkedett összehasonlítva egészséges, nem dohányzó önkéntesekkel (19). Az IL-6 főként pro-inflammatorikus citokinként ismert, de pontos hatásmechanizmusa COPD-ben nem tisztázott. Az IL-6 COPDben megemelkedett szintje miatt felmerült biomarkerként való alkalmazása is (20) COPD diagnosztikája A betegség diagnosztizálása alapvető légzésfunkciós vizsgálattal, (spirometriás méréssel) történik, ahol az első másodpercre eső erőltetett kilégzési térfogat (FEV1 - Forced Expiratory Volume) és az erőltetett kilégzési vitálkapacitás (FVC - Forced Vital Capacity) hányadosának csökkenése és a csökkenés mértéke utal az obstruktív légzészavarra és a légúti szűkület súlyosságára. A betegségnek a FEV1 értéke alapján enyhe, középsúlyos, súlyos és nagyon súlyos stádiumait különböztetjük meg a nemzetközi COPD ajánlás (GOLD - Global Initiative for Chronic Obstructive Lung Diasease) alapján. Bár legtöbbször a COPD-t elsőként 10
11 tüdőbetegségként diagnosztizálják, számos esetben szisztémás manifesztációk és komorbiditások is társulnak ehhez a kórképhez, melyek érintik a szívet (ateroszklerózis, miokardiális infarktus, pulmonális hipertónia, angina, stroke), a hasnyálmirigyet és más szerveket is. Illetve elősegíti a diabétesz, vázizom-diszfunkció vagy oszteoporózis kialakulását is (21) COPD terápiája A COPD gyógyszeres kezelésében meghatározó a tartós hatású hörgőtágítók inhalációban történő alkalmazása, bár a betegség enyhébb szakaszában a gyors hatású muszkarin receptor antagonistákat és a β 2 adrenerg receptor-agonistákat, illetve ezek kombinációit is alkalmazzák hörgőtágítóként. A szisztémás gyulladás csökkentésére a hörgőtágítók adása mellett az inhalációs kortikoszteroidok bevezetése elengedhetetlen, amikor az akut exacerbáció (a beteg légúti tüneteinek, úgymint légszomj, köhögés és köpetürítés súlyosbodásában jelentkező akut esemény) kezelése szükségessé válik. Eddigi vizsgálatok azt mutatják, hogy jelenleg a kezeléseknek limitált a hatékonysága, illetve a nem kívánt mellékhatások is gyakoriak lehetnek. Emellett fontos megemlíteni, hogy a kortikoszteroidok gyulladáscsökkentő hatásukat egyrészről a hiszton deacetiláz-2 (HDAC-2) működésén keresztül fejtik ki, így a COPDbetegek tüdejében és alveoláris makrofágjaiban oxidatív stressz hatására kialakuló HDAC-2 redukciója hozzájárul a kortikoszteroidok ellen fellépő terápiás hatástalansághoz (22) (23). Mindezek alapján a COPD igazi terápiás kihívást jelent ezért a betegség patomechanizmusának pontos megismerésére és új gyógyszercélpontok azonosítására irányuló felfedező kutatások különösen fontosak Wnt jelátvitel Wnt jelátvitel jellemzői és szerepe A Wnt jelátvitelben résztvevő Wnt fehérjék olyan szekretált glikoproteinek, melyek kanonikus és nem-kanonikus Wnt jelátviteli utakon keresztül kifejtve hatásukat jelentős szerepet játszanak az embrionális fejlődésben, a szövetek homeosztázisában, szabályozzák a sejtek osztódását, a sejtek sorsát, migrációját, adhézióját és polarizációját is. Emlősökben 19 különböző Wnt molekulát különböztetünk meg, melyek a sejtek membránjában elhelyezkedő úgynevezett Frizzled (FZD) receptorokhoz kötődnek, így aktiválva a Wnt jelátviteli útvonalat (24). A Wnt molekulákat csoportosíthatjuk kanonikus (Wnt1, Wnt3, Wnt3a, Wnt7a, Wnt7b, Wnt8) és nemkanonikus (Wnt5a, Wnt11) Wnt-okra a jelátvitel útvonalak aktiválása alapján (25). A Wnt 11
12 jelátviteli útvonal a gasztrointesztinális és a keringési rendszer, illetve a csontvelő fejlődésében és az öregedésben is részt vesz, de bizonyítottan jelentős szerepet vállal a karcinogenezisben és a gyulladásos folyamatok kialakulásában Kanonikus jelátvitel A kanonikus útvonal aktiválódása során a Wnt ligand a FZD receptorhoz kötődik, majd az LRP5/6 ko-receptorral együtt alakít ki stabil receptor komplexet. A Wnt-FZD-LRP5/6 komplex létrejötte a Dishevelled (Dvl) citoszolikus jelátvivő molekula 3 doménjének foszforilációját eredményezi. A Dvl aktivációja a GSK-3β (glycogen synthase kinase 3 beta) foszforilációjához és ennek eredményeként a gátlásához vezet. A GSK-3β gátlása a β-katenin stabilizációjához és ennek következtében citoszolikus akkumulációjához vezet. Ezt követően a stabilizálódott β- katenin transzkripciós koaktivátorként a sejtmagba transzlokálódik és a TCF/LEF transzkripciós faktor család egyik tagjához kapcsolódik. A transzkripciós komplex megfelelő kapcsolódása különböző célgének aktivációját okozza, úgy, mint a cyclin-d1, c-myc és c-jun. A Wnt ligand kötődésének hiányában a β-katenin nem képes felhalmozódni a citoplazmában, mivel az APC (Adenomatous Polyposis Coli) -Axin-GSK-3β komplex segítségével foszforilálódik, majd ubiquitinálódik és a citoplazmában proteoszómában proteolízissel lebomlik Nem-kanonikus jelátvitel A nem-kanonikus jelátvitel esetében két útvonalat különböztetünk meg. Az egyik a Jun N- terminális kináz által aktivált planáris sejt polaritás (PCP- Planar Cell Polarity) útvonal, illetve a kalmodulin kináz II-n és protein kináz C-n keresztül aktiválódó Wnt/Ca 2+ jelátviteli útvonal (26). A PCP útvonal esetében a Dvl aktiválódása a JNK és az AP1 transzkripciós faktorok aktivációjához vezet. Az AP1 egy fehérje komplex, mely több kisméretű fehérjéből áll, pl. cjun, JunB, JunD, cfos, FosB, Fra1, Fra2, ATF2 és CREB. A nem-kanonikus Wnt jelátvitel kiváltja az AP1 fehérjék expresszióját, amelyek szabályozzák a ciklin D1-et, mátrix metalloproteinázt, Bim-et és GM-CSF-et (27). 12
, a Ca 2+ -Kalmodulin Kináz II (CaMKII), a kálcium szenzitív foszfatáz és a")
 transzkripciós faktor aktiválódásával immunfolyamatokban")
 (3. ábra).")
13 A nem-kanonikus útvonal másik fő típusának, a Ca 2+ függő jelátvitelnek számos célmolekulája létezik, mint például a protein kináz C (PKC), a Ca 2+ -Kalmodulin Kináz II (CaMKII), a kálcium szenzitív foszfatáz és a kalcineurin. Végül az aktivált T-sejtek nukleáris faktora (NFAT) transzkripciós faktor aktiválódásával immunfolyamatokban fontos szerepet játszó gének átíródása stimulálódik, úgymint az interleukin-2 (IL-2), interleukin-4 (IL-4) és a tumor nekrózis faktor alfa (TNF-alfa) (3. ábra). Kanonikus útvonal PCP útvonal Nem-kanonikus útvonalak Wnt/Ca 2+ útvonal 3. ábra: A Wnt jelátviteli útvonalak és főbb jelátviteli komponenseik összefoglalása. 13
14 1.4. Wnt5a és PPAR-γ szerepe a gyulladásos folyamatokban Wnt5a gyulladást indukáló szerepe A Wnt jelátviteli útvonal minden tagját megvizsgálva, a rendelkezésre álló irodalom alapján a legtöbbet kutatott és a karcinogenezisben, illetve a gyulladásos folyamatokban a legjelentősebb szerepet betöltött molekula a nem-kanonikus, szekretált Wnt fehérje, a Wnt5a. Számos kutatási eredmény bizonyította a Wnt5a emelkedett expresszióját több karcinóma esetében is, pl. emlőkarcinómában, petefészek tumorban, prosztata karcinómában és nem-kissejtes tüdődaganatokban (28)(29). A nem-kissejtes tüdőrákok vizsgálatával további Wnt molekulák fontos szerepét bizonyították, ahol a Wnt1, Wnt2, Wnt3 és Wnt11, illetve a β-katenin, Frizzled- B emelkedett expressziója volt mérhető, ezzel párhuzamosan pedig a jelátvitel negatív szabályozójaként számon tartott Axin, GSK3β, Dkk-1 csökkenését figyelték meg (30). Különböző Wnt molekuláknak a daganatok kialakulásában betöltött szerepét vizsgálva, a Wnt5a hiányát leírták már leukémiában (31), petefészek karcinómában (32) és pajzsmirigyrákban is (33). Az említett malignitásokban a Wnt5a tumor szupresszorként viselkedik, a karcinogenezisben szerepet játszó Wnt/β-katenin útvonal blokkolójaként. A Wnt5a alapvetően a nem-kanonikus jelátvitel molekulájaként a kálcium függő útvonalat aktiválja (34), azonban legutóbbi tudományos eredmények bizonyítják, hogy a β-katenin függő, kanonikus Wnt jelátviteli útvonalon is képes hatását kifejteni (35). Ellentétben tumor szupresszáló hatásával, más humán karcinómákban, mint pl. melanomában és hasnyálmirigy daganatban a Wnt5a fokozza a tumorsejtek invázióját és migrációját az aktin átrendeződésén, sejt adhéziós változásokon és az EMT folyamatán keresztül (36). Ezek alapján elmondhatjuk, hogy a Wnt5a képes gátolni a β-katenin által szabályozott karcinogenezist, de ezzel ellentétesen elősegíthet tumor progressziót is β-katenintól független mechanizmuson keresztül. Ezek alapján feltételezhetjük, hogy a Wnt5a emberi tumorokban vállalt pontos szerepe jelentősen függ a tumor mikrokörnyezetétől is. Kutatócsoportunk eredményei alapján a Wnt5a aktivációját nem-kissejtes tüdődaganatokon belül a laphámkarcinómában (SCC) figyeltük meg a Wnt11 molekula emelkedésével párhuzamosan (37). Összefoglalva elmondhatjuk, hogy a Wnt5a molekulát pro-inflammatorikus molekulaként tartjuk számon, mely dohányfüst-indukálta gyulladásos citokinek aktivátoraként játszik szerepet (38) (39) (40). A Wnt5a a gyulladásos folyamatokat a mitogén aktiválta protein kináz (MAPK) és NFκB kaszkádokon keresztül, TNFα és IL-1β inflammatorikus mediátorok termelődésének hatására az IκB kinázok (IKK) aktiválódásával és az IκB inhibitorok foszforiláció függő degradációjával indukálja (34) (41) (42). 14
15 Ismert, hogy a dohányfüst a gyulladás indukálásával egy időben a makrofágok aktivációját, polarizációját eredményezi a klasszikus M1 és az alternatív M2 útvonalak irányába (43). A makrofágok jelenléte elengedhetetlen a szervezet általános homeosztázisának fenntartásában, a gyulladásos folyamatokban, a különböző immunreakciókban és az immunitás kialakításában (28). A Wnt5a expresszióját mérték már perifériás vér mononukleáris sejtekből (PBMC) (44) származó makrofágokban, csontvelőből származtatott makrofágokban (BMDM) (45), alveoláris makrofágokban, peritoneális makrofágokban és mikrogliákban is (46). Emellett a Wnt5a receptorát, a FZD5 expresszióját is kimutatták makrofágokban. Kutatások bizonyítják, hogy a makrofágokat pro-inflammatorikus citokinek képesek indukálni a Wnt5a szint emelkedésének hatására, ahol a makrofág eredetű Wnt5a molekulák nem csak szolubilis fehérjeként szekretálódnak, hanem mikrovezikulákban is megjelenhetnek. A Wnt5a a makrofág eredetű exoszómák, mikrovezikulák segítségével is szállítódhat egészen a recipiens sejtekig (47) Peroxiszóma proliferátor-aktivált receptor γ (PPAR-γ) gyulladáscsökkentő szerepe A gyulladás komplex természetét tekintve elmondhatjuk, hogy rendkívül sok tényező befolyásolja, így természetesen a Wnt5a mellett más faktorok is szerepet jászhatnak a folyamat szabályozásában. Az utóbbi években leírták, hogy a PPAR-γ szintén egy központi szereplője a komplex gyulladásos folyamatoknak (48) (49). A PPAR-γ lipidek által aktivált transzkripciós faktor, mely a ligand aktivált nukleáris hormon receptor szupercsalád tagja. Szerepét tekintve a lipid metabolizmussal, adipogenezissel, gyulladással és a metabolikus homeosztázissal kapcsolatos gének átíródásában vesz részt. A PPAR-γ összességében egy kiemelkedően fontos anti-inflammatorikus és anti-oxidatív molekuláris célpont (50) (51) (48) (52). A PPAR-γ agonisták számos gyulladásos mediátor termelődésének gátlásával képesek antiinflammatorikus, anti-aterogenikus (érfalban lerakódást gátló) és anti-oxidáns hatásokat kifejteni (53) (54)(55) (56) (57) (58) (59) (60). Korábbi kutatások leírták, hogy a PPAR-γ regulációja részben Wnt jelátvitel függő (61), ahol legtöbbször a kanonikus vagy β-katenin függő jelátvitel aktiválódása mellett a PPAR-γ inaktiválódása figyelhető meg (62) (63) (27). Emellett, a PPAR-γ a nem-kanonikus Wnt jelátviteli útvonalon keresztül is szabályozza az erek növekedését egy Wnt5a függő jelátviteli mechanizmuson keresztül (38). 15
16 Az eddigi megfigyelések alapján a PPAR-γ anti-inflammatorikus aktivitását összefüggésbe hozhatjuk gyulladásos tüdőbetegségekkel, mely erősíti azt a hipotézist, hogy a PPAR-γ agonisták képesek csökkenteni a gyulladást (64) (65), illetve visszafordítani a cigarettafüstindukálta emfizémát is (66). A PPAR-γ gyulladásban betöltött szerepét bizonyítja, hogy PPARγ deficienciával rendelkező makrofágok hozzájárulnak spontán gyulladás kialakulásához a tüdőben (67), illetve a PPAR-γ agonisták az immunszupresszív útvonalat képviselő M2 makrofágok differenciációját segítik elő (68). Ennek következtében valószínű, hogy a multifaktoriális folyamatként számon tartott makrofág polarizáció COPD betegségben összefüggésbe hozható a PPAR-γ működésével. Korábbi publikációk és a saját kutatócsoportunk által végzett megfigyelések alapján is feltételezzük, hogy bár mind a kanonikus, mind pedig a nem-kanonikus Wnt jelátvitel szabályozza a PPAR-gammát, a nemkanonikus jelátviteli útvonal az, amely dohányfüst expozíció hatására aktiválódik. Összefoglalva, a PPAR-γ molekuláris szabályozása egy rendkívül összetett folyamat, melynek aktiválódása, vagy éppen gátlása multifaktoriális szabályozás alatt áll. Ezt a komplexitást korábbi kutatásaink során is bizonyítottuk, ahol eredményeink azt mutatták, hogy tumoros környezetben a PPAR-γ gátlásának hátterében álló egyik mechanizmus a Wnt5a-nak a mir-27b aktiválódására kifejtett hatásán keresztül szabályozódik, amely végül a PPAR-γ csökkenéséhez vezet (38). Ebből arra következtethetünk, hogy amennyiben a PPAR-γ aktiválódásának és gátlásának szerepét akarjuk jobban megismerni, nem hagyatkozhatunk egyetlen faktor vizsgálatára, hanem a funkció mögött álló komplex hálózat feltérképezésére van szükség. Ezen bonyolult szabályozási mechanizmusok mellett a szisztémás gyulladás kialakulásának hátterében feltételezzük, hogy a dohányfüst expozíció-indukálta lokális gyulladás az expozíció helyétől különböző extracelluláris vezikulákba csomagolt úgynevezett gyulladásos üzenetek elszállítódásával válhat az egész szervezetet érintő, szisztémás gyulladássá. 16


17 1.5. Extracelluláris vezikulák szerepe a krónikus gyulladásban Extracelluláris vezikulák definíciója és szerepe Legújabb tudományos eredmények bizonyítják, hogy a sejtek képesek membránba zárt partikulumok felszabadulásán keresztül is a sejt-sejt közötti kommunikációra. Ez a fajta információcsere evolúciósan konzervált folyamat, ahol a partikulumok az extracelluláris vezikulák (EV), melyek kettős foszfolipid membránba csomagolva bármilyen típusú sejtből képesek az extracelluláris térbe felszabadulni. Jelenleg az extracelluláris vezikulák populációit méretük, sűrűségük, szubcelluláris eredetük és funkciójuk alapján különíthetjük el egymástól. Megkülönböztetünk exoszómákat ( nm), mikrovezikulákat (MV) és a nemrég felfedezett tumor eredetű EV populációját, az úgynevezett apoptotikus testeket vagy nagy onkoszómákat (4. ábra). 4. ábra: Extracelluláris vezikulák típusainak és azok eredetének összefoglalása (69) Az exoszómák definíciója és szerepe Az exoszómák olyan sejtekből származó kisméretű mikrovezikulák csoportja, melyek mérete nm között változik. Szerkezetüket tekintve kettős rétegű lipidmembrán határolja tartalmukat, melyek anyagcseréből származó fehérjéket és a ribonukleinsavak több változatát tartalmazzák, főként mikrorns-t, de tartalmaznak mrns-eket, fehérjéket és lipideket is (70) (71). Az exoszómák extracellulárisan szabadulnak fel multivezikuláris endoszómák 17
18 sejtmembránnal történő fúziója következtében. Termelődésük helyétől az egész szervezetbe képesek eljutni, majd a célsejtekben internalizálódni endocitózis és membránfúzió segítségével. A fent említett tulajdonságaik alapján tekintik őket a sejt-sejt közötti kommunikáció mediátorainak, mely megvalósulhat egymás mellett lévő sejtek és egymástól távoli sejtek esetében egyaránt (72) (73) (74). Az exoszóma által mediált sejtek közötti kommunikáció jelentősen befolyásolhatja a betegségek megjelenési formáit. Szerepük a különböző betegségekben gyakran kontextusfüggő, melyet nagyban meghatároz az őket termelő sejt típusa (73) (75). Az exoszómák legtöbbször olyan sejtekből szabadulhatnak fel, melyek fontos szerepet töltenek be az immunreakciók kialakításában, mint pl. a B-sejtekből, (76), dendritikus sejtekből (77), T-sejtekből (78), hízósejtekből (79) és epitél sejtekből (80). Előfordulásukat tekintve megtalálhatóak fiziológiás folyadékokban, mint pl. a bronchoalveoláris folyadék (BAL) (81), a szérum (82), a vizelet (83) és az anyatej (84). Az exoszómák lehetséges szerepét a COPD szisztémás gyulladásos folyamatainak kialakításában már korábban feltételezték, COPD-betegek plazmájában mért emelkedett exoszómák mennyiségéből és ezzel korrelláló szisztémás gyulladásos biomarkerek, C-reaktív protein (CRP) és IL-6 szintjének emelkedésére alapozva (85) Mikrovezikulák és apoptotikus testek (onkoszómák) definíciója és szerepe A mikrovezikulák az exoszómákhoz viszonyítva nagyobb, nm-es mérettartományba eső, a plazmamembránból lefűződéssel keletkező extracelluláris vezikulák, melyek foszfatidilszerinben gazdagok és egyéb membránkomponenseket is tartalmaznak. A legnagyobb extracelluláris vezikulák az apoptotikus testek, másik nevükön az onkoszómák, melyek nm-es mérettartományba tartoznak, a sejtek apoptózisa során a plazmamembránból való felszabadulással keletkeznek. Az onkoszómák a plazmamembránból és a citoszólból származó fehérjéket, DNS fragmentumokat tartalmazhatnak (86). Hasonlóan az exoszómákhoz, a nagyobb méretű extracelluláris vezikulák esetében is több kísérletben bizonyították már szerepüket az immunreakciók promotálásában. Korábbi kutatásokban cigarettafüst extraktummal (CFE) kezelt makrofágokból izolált EV-ban pro-inflammatorikus mediátorként alarmint mértek. A CFE indukált makrofágokból és epitél sejtekből, illetve dohányos humán BAL mintákból izolált EV-ban az IL-6 és IL-8 gyulladásos citokinek emelkedett szintjét találták (87) (88). Ezek alapján következtethetünk, hogy az extracelluláris vezikulák nagymértékben hozzájárulhatnak a gyulladás kialakulásához és szerepük lehet az immunsejtek toborzásában is (89). 18
19 1.6. Tüdő regenerációs folyamatai és a Wnt jelátvitel kapcsolata ATI és ATII típusú sejtek szerepe a tüdőben és a transzdifferenciációban A tüdő szerkezetét felépítő alveolusok nagy és magasan vaszkularizált felülettel rendelkeznek, melyek a hatékony légcseréért felelősek. Az alveoláris felület állandó kapcsolatban van a környezettel, esetekben a dohányfüsttel is, mely képes károsítani a gázcsere felületét (90)(91). Ahogy korábban már említettük, a COPD során a légzőfelület jelentős csökkenése figyelhető meg, mely a gázcserében szerepet játszó alveolusok állapotát is nagymértékben érinti. Ennek eredményeképpen a tüdő regenerációs képességének és a légzőfelület differenciálódásának mélyebb megismerése kiemelkedően fontos a terápiás lehetőségeket tekintve. Az alveolusokat két különböző epitél sejt építi fel, az alveoláris epitél I. (ATI) és az alveoláris epitél II. (ATII) sejtek. Az ATI-es típusú sejteken keresztül megy végbe a gázcsere, míg az ATII-es típusú sejtek funckiója komplexebb, többek között feladatuk a szöveti regeneráció és a felületi feszültség csökkentése. Az ATI-es sejtek laposak, vékonyak és az alveoláris felszín 95%-át teszik ki. Az ATII-es típusú sejtekhez képest érzékenyebbek a tüdőt érő sérülésekre, amelyekre apoptózissal reagálnak. Ezzel szemben az ATII-es sejtek rendelkeznek őssejt funkcióval, azaz fakultatív progenitor sejtek a tüdő alveoláris régiójában, melyek megújítják saját magukat és sérülés esetén klonális proliferációra és migrációra képesek. Az ATII-es alveoláris sejtek termelik a szörfaktáns fehérjéket (SFP), melyek a felületi feszültséget csökkentő lipoproteinek az alveolusokban. Különféle kísérleti rendszerekben bizonyították, hogy az ATII-es típusú sejtek transzdifferenciációval (TD) ATI-es sejtekké alakulhatnak (92) (93) Wnt jelátvitel szerepe a tüdő regenerációs folyamataiban Korábbi, a szervek regenerációjával foglalkozó kísérletek bizonyították, hogy a regenerációs folyamatok hátterében a korábbi fejlődési mechanizmusok újra aktiválódása megy végbe (94). Felnőttkorban is a BMP, FGF, Notch és Wnt jelátviteli útvonalak vesznek részt a szöveti regeneráció folyamataiban (95). A Wnt családhoz tartozó ligandok a tüdő fejlődése során szabályozzák a sejtproliferációt, differenciációt, polaritást, adhéziót és migrációt (27), amíg a Wnt β-katenin szignalizáció az alveoláris morfogenezishez szükséges (96). További Wnt ligandok, mint a Wnt5a (97), a Wnt7b (98) és a legtöbb FZD receptor központi szerepet töltenek be a tüdő fejlődésében (99). Bár a Wnt jelátvitel szerepe a tüdő regenerációjában ismert, a TD folyamatában betöltött pontos szerepe és mechanizmusa nem feltérképezett. 19
20 Mindezek alapján feltételezhetjük, hogy a Wnt jelátvitel szükséges lehet az ATII-ATI TD indukciójában és szabályozásában, amely akár lehetőséget biztosíthat a cigarettafüst által károsított tüdőszövet irányított regenerálásában. 20
21 2. Célkitűzés Kutatócsoportunk korábbi eredményei igazolták, hogy a tüdő daganatos elváltozásaiban, differenciáltan expresszálódnak a Wnt ligandok. Míg a tüdő adenokarcinómára (AC) a kanonikus Wnt molekulák jellemzőek, addig a tüdő laphámkarcinómában (SCC) a nemkanonikus Wnt5a molekula emelkedése mérhető (37) (38). A tüdőt érintő súlyos betegségek, úgymint a tüdőkarcinóma és a COPD szorosan összefüggő komorbiditások lehetnek, melyeknek fő kiváltó oka a dohányzás. Ismert, hogy a dohányzás szisztémás gyulladásos folyamatokat indít be az emberi szervezetben, mely később karcinogenezishez vezethet (100) (101). Mindezek alapján PhD kutatásaim során a következő központi kérdésekre kerestük a választ: 1. Kiváltja-e a cigarettafüst a nem-kanonikus Wnt5a molekula szintjének emelkedését és a Wnt5a-val kapcsolatosan COPD-re jellemző gyulladásos környezet kialakulását? 2. Kimutatható-e a Wnt5a emelkedett szintje COPD-betegek szérumában, amely magyarázhatja az emberre, de a kísérleti állatokra nem jellemző szisztémás gyulladás kialakulását? 3. Szabályozza-e, és ha igen, milyen módon a Wnt jelátviteli útvonal a pulmonális regenerációt és az epitéliális transzdifferenciációs folyamatokat? 21
22 3. Anyagok és módszerek 3.1. Etikai engedély In vivo állatkísérleteinket a Pécsi Tudományegyetem Állatkísérletes Etikai Bizottsága által az Állatkísérletek Etikai Kódexe alapján jóváhagyott engedéllyel végeztük (BA02/2000-5/2011). Kísérleteink során követett protokollokat a Pécsi Tudományegyetem Állatkísérletes Etikai Bizottsága jováhagyta. További kísérleteink során felhasznált humán vérmintákat a Pécsi Tudományegyetem Klinikai Központ I.sz. Belgyógyászati Klinika, Pulmonológia Osztályáról kaptuk. A mintákat a Pécsi Tudományegyetem Klinikai Központ Humán Klinikai Vizsgálatok Regisztrációs Központjának jóváhagyásával gyűjtöttük (ETT 6444/2016). A szérumok a betegek írásos beleegyezését követően, anonim mintaként kerültek laboratóriumi feldolgozásra Anyagok Az in vitro kezelésekhez a cigarettafüst extraktumot sejtkultúra médiumban higítottuk, 0,5%- os koncentrációban. A humán rekombináns Wnt5a, Wnt4 és Wnt7a fehérjéket az R&D Systems-től vásároltuk (Chinese Hamster Ovary Cell Line, CHO-derived Gln38-Lys380, Minneapolis, USA) és kísérleteink során 1 μg/ml-es koncentrációban alkalmaztuk a sejteken. A PPAR-γ agonistát, roziglitazont (RSG) és antagonistát (GW9662) a Sigma-Aldrich-tól vásároltuk (St. Louis, USA) és 10 μm koncentrációban alkalmaztuk a kezelésekhez Humán vérminták gyűjtése A humán vérmintákat az I. számú Belgyógyászati Klinika Pulmonológia Osztályáról (Általános Orvostudományi Kar, Pécsi Tudományegyetem) gyűjtöttük. Összesen 5 COPD-beteg és 5 korban megegyező egészséges kontroll vérmintáját gyűjtöttük pro-koagulációs vérgyűjtő csövekben (BD Vacutainer, SST: BD SST Tubes with Silica Clot Activator and Polymer Gel, Franklin Lakes, NJ, USA). A mintákat 30 percig szobahőmérsékleten állni hagytuk, majd 1500 rpm-n 10 percig centrifugáltuk. További felhasználásig a szérum mintákat -80 C tároltuk Kísérleti állatok In vivo kísérleteinkhez C57BL/6 hím 8 hetes egereket (20 25 g; n=4-5/csoport) használtunk, melyeket a Pécsi Tudományegyetem Általános Orvostudományi Karának Farmakológiai és Farmakoterápiai Intézetének állatházában tartottak. További kísérleteinkben 8 hetes BALB/c hím egereket (20 25 g; n=3/csoport) használtunk, amelyek a PTE ÁOK Immunológiai és 22
23 Biotechnológiai Intézet állatházában voltak fenntartva. Víz és étel korlátlan mennyiségben állt rendelkezésükre In vivo cigarettafüst (CF) expozíció C57BL/6 hím egerek (20 25 g; n=4-5/csoport) teljes test cigarettafüstnek voltak kitéve, ahol a dohányoztatáshoz 3R4F típusú kísérleti cigarettákat használtunk a füstszűrő eltávolítása után (College of Agriculture, University of Kentucky; Lexington, KY). A cigarettafüst expozíció egy manuális rendszerben történt, naponta 2 x 2db cigarettával, alkalmanként 30 percen keresztül (TE2; Teague Enterprises, Woodland, CA). Az összes partikulum átlag koncentrációja a dohányoztató kamrában 70 mg/m 3 (102) Egértüdő sejtek izolálása A kísérleti egerek altatása intraperitoneálisan ketamin-xylazinnal (5 mg/kg) történt, majd nyaki diszlokációt követően az egerek tüdejét eltávolítottuk. A tüdőszöveteket három csoportba osztottuk, így a tüdőszöveteket további molekuláris biológiai kísérletekhez RNA laterben (Qiagen; Hilden, Germany), Western-blot elemzésekhez 1 ml lízis pufferben, immunhisztokémiai vizsgálatokhoz pedig 4%-os formaldehid oldatban (Szkarabeusz Ltd.; Pécs, Hungary) tároltuk további felhasználásig. A tüdőben lévő sejtek izolálásához három C57BL/6 egeret használtunk fel csoportonként. Az egerek abdominális aortájának átvágása után az állatok perfúziója következett, ahol a tüdőben maradó vér eltávolítása érdekében 10 ml PBS-sel a jobb kamrán keresztül végeztük a perfúziót. A tüdőszövet teljes elemésztéséhez 3 ml tripszint használtunk, majd 10 ml PBS-t alkalmaztunk a tripszin kimosására. A légcsövek feltöltéséhez 3 ml kollagenáz-diszpázt (3 mg/ml kollagenáz (Sigma-Aldrich, St. Louis, USA), 1 mg/ml diszpázt (Roche F. Hoffmann-LaRoche Ltd. Basel, Switzerland) és 1u/μl DNáz I-et (Sigma-Aldrich, St. Louis, USA) használtunk. Ezután a tüdőlebenyeket kisebb darabokra vágtuk, majd 10 ml kollagenáz-diszpáz oldatban folyamatos mágneses kevergetés mellett 50 percig emésztettük. Az elemésztett tüdőszövetet 70 μm-es szűrőn átszűrtük (BD Becton, Dickinson and Company, Franklin Lakes, NJ, USA) Sejtszortolás Az egértüdő egysejt szuszpenziót anti-egér CD45 - FITC és anti-egér EpCAM1 (G8.8 anti-rat- PE) (40) antitesttel jelöltük meg, melyet a Pécsi Tudományegyetem Általános Orvostudományi Karának Immunológiai és Biotechnológiai Intézetében termeltek. A sejtszortolást a FACSAria III (BD Biosciences, San Jose, CA, USA) szorterrel végeztük. A szortolás alapján a következő 23
24 populációkat tudtuk elkülöníteni: EpCAM - CD45 -, EpCAM + CD45 -, EpCAM + CD45 + és EpCAM + CD45 -. A szortolás tisztasága 99% feletti volt. A humán tüdőből frissen izolált sejteket (n=12) 0,1% BSA-t (Sigma-Aldrich, St. Louis, Missouri, USA) és 0,1% Na-Azidot (Sigma-Aldrich, St. Louis, Missouri, USA) tartalmazó PBS oldatban mostuk. Ezután a sejteket az antitestekkel 30 percig inkubáltuk. EpCAM-FITC, CD208-PE és Podoplanin-APC konjugált antitesteket használtunk az ATI-es típusú (EpCAM + Podoplanin + CD208 - populáció) és az ATII-es típusú (EpCAM + Podoplanin - CD208 + populáció) epiteliális sejtek differenciáltatására. A sejteket Beckman-Coulter MoFlo XDP high-speed szorter segítségével szortoltuk. A szortolás eredményeként átlagosan 4.43x10 4 ATIes és 1.39x10 5 ATII-es típusú sejteket gyűjtöttünk Cigarettafüst extraktum (CFE) elkészítése A cigarettafüst extraktum oldat elkészítéséhez egy korábban már leírt, de a laboratóriumunkban módosított módszert alkalmaztuk (41). Két, 3R4F típusú kísérleti cigaretta füstjét átbuborékoltattuk 10 ml FBS mentes RPMI médiumban (Lonza, Basel, Switzerland) perisztaltikus pumpa (Harvard Apparatus, P-1500, Holliston, USA) segítségével, 550ml/perc áramlási sebességgel, 3 percen keresztül. Az átbuborékoltatott CFE oldatot 0,22 μm átmérőjű szűrőn (TPP, Trasadingen, Switzerland) átszűrtük és ezt tekintettük 100%-os oldatnak, melyet a további kísérletekhez sejttenyésztő médiummal higítottunk és 0,5%-os koncentrációban alkalmaztunk a sejteken. A CFE oldatok ph értékeit minden alkalommal lemértük, melynek átlaga ph 7,28 volt. Az elkészített CFE oldatok minden esetben 30 percen belül felhasználásra kerültek. A CFE oldatokban beoldódott részecskék optikai denzitását 360 nm-en spektrofotometriásan határoztuk meg és a CF-el nem átbuborékoltatott, kontroll oldatokhoz hasonlítottuk Sejtvonalak és primer sejtkultúrák A humán adenokarcinóma A549 sejtvonalat az ATCC-től (American Type Culture Collection, Rockville, MD) vásároltuk, a transzgenikus Wnt5a overexpresszáló-a549 sejtvonalat pedig lentivirális transzgenezissel állítottuk elő saját laboratóriumunkban (42). A549 sejtvonalat komplett Dulbecco's Modified Eagle Medium (DMEM) sejttenyésztő médiumban növesztettük kiegészítve 10% borjú szérummal (FBS), 200mM L-glutaminnal, illetve penicillinsztreptomicin oldattal. Primer sejtekként normál, primer kislégúti epitélsejtet (SAEC, small airway epithelial cells), normál tüdő fibroblaszt sejtet (NHLF, normal human lung fibroblast) és normál mikrovaszkuláris endotélsejtet (HMVEC-L, human lung microvascular endothelial 24
25 cells) alkalmaztunk a kísérletek során. A sejtek tenyésztése során a gyártó által javasolt médiumokat (SAGM, Small Airway Growth Medium, FGM-2, Fibroblast Growth Medium és EGM-2, Endothelial Growth Medium) használtuk és a sejteket standard sejttenyésztési körülmények között (37 C, 5% CO2) tartottuk Primer humán monociták izolálása A monociták izolálása egészséges donorok véréből történt, melyeket a Pécsi Regionális Vérellátó Központból kaptunk. Heparinizált csövekben 20 ml perifériás vér felhasználásával és Fikoll grádiens centrifugálás segítségével perifériás vér mononukleáris sejteket (PBMC, peripheral blood mononuclear cells) izoláltunk (GE Healthcare, Sigma-Aldrich, St. Louis, USA). A monocitákat mágneses aktivált sejtválogatással (MACS, Miltenyi Biotec, GmbH, Bergisch Gladbach, Germany) anti-humán CD14 antitesttel konjugáltatott mikrogyöngyök segítségével, pozitív szelekció útján nyertük ki. A PBMC sejtszuszpenziót CD14 + mikrogyöngyökkel inkubáltuk és egy mágneses oszlop, a MACS MultiStand, OctoMACS (Miltenyi Biotech GmbH, Bergisch Gladbach, Germany) segítségével a CD14 + sejteket kiszortoltuk. A monocita populáció tisztasága > 85% volt. A makrofágok differenciációját 10 nm PMA (Phorbol 12-myristate 13-acetate) kezeléssel indukáltuk 37 C-on, 5%-os CO 2 tartalom mellett 5 órán keresztül dimenziós (3D) humán tüdőmodell A 3D tüdő aggregátumokat, másik néven szferoidokat, egy korábban született szabadalom (P ) alapján készítettük el (103). Kísérleteink során a 3D tüdőmodellünkhöz az előzőekben leírt humán primer sejteket használtuk fel (kislégúti epitélsejt - SAEC, normál tüdő fibroblaszt - NHLF, normál mikrovaszkuláris endotélsejt - HMVEC-L), illetve a primer sejtek mellett makrofágot tartalmazó 3D tüdőmodelleket is készítettünk. A primer sejteket a 3D modellhez a korábban leírt módon tenyésztettük (103). Miután a sejtek elérték a 80%-os konfluenciát, tripszinnel történt emésztés után a sejteket lecentrifugáltuk, majd a háromféle sejttípusból sejtszuszpenziót készítettünk. A sejteket a következő arányban kevertük össze: makrofágot nem tartalmazó 3D aggregátum esetében: 40% NHLF, 30% SAEC, 30% HMVEC- L és makrofágokat tartalmazó 3D aggregátum esetében: (30% NHLF, 25% SAEC, 25% HMVEC-L és 20% makrofágok). A sejtszuszpenziót egy 96-lyukú, nem kitapadó sejtekhez kialakított, U-aljú sejttenyésztő lemezre (Corning, New York, USA) osztottuk szét, majd a lemezeket 10 percig 600 g-n szobahőmérsékleten centrifugáltuk. A lemezeket ezután standard sejttenyésztési körülmények között inkubátorban hagytuk egy éjszakán keresztül. A 25
26 szferoidokat 2:2:1 arányban összekevert SAGM:EGM:FGM sejttenyésztő médiumokban tenyésztettük. Kísérleteink során a tüdő aggregátumokat 0,5%-os CFE-vel, emellett 1 μg/ml humán Wnt5a rekombináns fehérjével kezeltük 48 órán keresztül RNS izolálás, cdns szintézis, valós idejű kvantitatív PCR (qpcr) A 3D aggregátumokból a totál RNS kivonását az oszlopon történt DNáz emésztéssel együtt NucleoSpin RNAII kittel (Macherey-Nagel, Düren, Germany) végeztük. Az izolált RNS minta koncentrációját és minőségét NanoDrop készülékkel mértük. A cdns készítéséhez egy magas hatékonyságú cdns író kit random hexamer primerjeit alkalmaztuk (Thermo Fisher Scientific, Waltham, MA USA). A cdns szintéziséhez 1 μg totál RNS-t használtunk a gyártó ajánlása szerint. A génexpressziós változásokat valós idejű kvantitatív PCR-al, SensiFAST SYBR Green reagens segítségével detektáltuk (BioLine, London, UK), a génexpressziós analíziseket pedig ABI StepOne és StepOnePlus (Thermo Fisher Scientific, Waltham, MA USA) készülékkel végeztük. A génexpresszióban bekövetkező változások kiszámítására a 2 -ddct módszerét alkalmaztuk az alábbi képlet alapján: Ct (célgén) - Ct (belső kontroll gén). A génexpresszió relatív kvantifikációját pedig a következő formula alapján számítottuk ki: RQ=2 - ddct, ahol a ddct a következő egyenletből származik: dct (kezelt minta) - dct (kontroll minta). Az adatokat StepOne szoftverrel analizáltuk, az expressziós változásokat pedig a β-aktin génhez normalizáltuk. Az izolált ATI és ATII sejteket szortolás után lizáltuk, majd azokból RNS-t izoláltunk RNeasy kit (Qiagen, Hilden, Németország) segítségével és 200 ng total RNSt felhasználva cdns írtunk WT Expressziós Kit (Ambion, Thermo Fisher Scientific, Waltham, USA) felhasználásával TaqMan Array A Wnt jelátviteli útvonalhoz tartozó molekulák mrns szintű vizsgálatához 92 gént és 4 belső kontroll gént tartalmazó Wnt lemezt használtunk (Human WNT Pathway, Fast 96-well TaqMan Array, Thermo Fisher Scientific, Waltham, USA). Humán monocitákból totál RNS kivonása után cdns-t készítettünk. A TaqMan master mix keveréket a monocitákból származó cdns mintáinkkal összekevertük, majd ezt a szuszpenziót osztottuk szét a Wnt géneket tartalmazó 96-lyukú lemezen. A génexpressziókat ABI StepOnePlus rendszerrel analizáltuk (Thermo Fisher Scientific, Waltham, MA USA) Immunhisztokémiai és immunfluoreszcens festések Az C57BL/6 egereket ketamin-xilazinnal altattuk el, majd a tüdejüket eltávolítottuk. A tüdőszövetek formaldehides fixálása után a PTE ÁOK Farmakológiai és Farmakoterápiai 26
27 Intézetében paraffinba ágyazott metszeteket készítettünk mikrotóm segítségével. A festési protokoll elvégzése előtt a metszeteket xilénnel deparaffinizáltuk, majd rehidratáltuk. Citrát pufferrel (10 mm citromsav, 0.05% Tween 20, ph 6.0) antigén feltárást végeztünk. A fluoreszcens festéshez patkány anti-humán Wnt5a monoklonális antitestet (1:100, Clone , R&D Systems, Minneapolis, USA), másodlagos antitestként Alexa Fluor 488 szamárban termeltetett anti-patkány IgG poliklonális antitestet (1:200, Thermo Fisher Scientific, Waltham, USA) használtunk. Az egértüdő metszeteken a sejtmagokat DAPI (1:1000, Serva, Heidelberg, Germany) magfestékkel vizualizáltuk. A fluoreszcens képeket egy Olympus IX81 (Shinjuku, Tokyo, Japan), CCD kamerával és analízis szoftverrel felszerelt fluoreszcens mikroszkóppal készítettük. A fluoreszcens képek szerkesztését és a fluoreszcens intenzitás mérését ImageJ képszerkesztő programmal végeztük. Az in vitro kísérleteink során a CFE kezelt 3-dimenziós aggregátumokat óvatosan eltávolítottuk a 96-lyukú sejttenyésztő lemezről és egy Cryomount (Histolab, Finland) beágyazó médium segítségével beágyazó kazettákba helyeztük őket és az aggregátumokat felhasználásig -80 Con tároltuk. Fluoreszcens festésekhez 8 µm-es metszeteket készítettünk Leica kriosztáttal (CM1950, Leica, Wetzlar, Germany). Egy szárítási lépés után a metszeteket hideg acetonban (Molar Chemicals Kft., Hungary) 10 percig fixáltuk. A fixált metszeteket 20 percig 5% BSA-t (Sigma Aldrich, St. Louis, USA) tartalmazó PBS-ben blokkoltuk. A fluoreszcens festéshez elsődleges ellenanyagként patkány elleni Wnt5a monoklonális antitestet (1:100, Clone , R&D Systems, Minneapolis, USA) használtunk, amelyet 1 óráig inkubáltuk a metszeteken, majd háromszori PBS-ben történt mosás után a másodlagos, Alexa Fluor 488 szamárban termeltetett patkány elleni IgG poliklonális antitestet (1:200, Thermo Fisher Scientific, Waltham, USA) 45 percig inkubáltuk a metszeteken. A másodlagos antitest után a metszeteket ismét háromszor mostuk PBS-ben. Továbbiakban az ATI-es típusú sejtek differenciációs markereként az AQP5 fehérjét anti-aq5 nyúl poliklonális IgG-vel (1 100, sc-28628, Santa Cruz Biotechnology, Dallas, USA), az ITGAV fehérjét anti-cd51 poliklonális kecske antitesttel (1:100, PA , Thermo Fisher Scientific, Waltham, USA) mutattuk ki. Másodlagos antitestként Alexa Fluor 568 kecskében termeltetett anti-nyúl IgG-t (1:2000, ab175471, Abcam Plc, Cambridge, United Kingdom) és Alexa Fluor 488 konjugált anti-egér IgG antitestet (1:200, Thermo Fisher Scientific, Waltham, USA) használtunk. A sejtmagokat DAPI (1:1000, Serva, Heidelberg, Germany) magfestékkel vizualizáltuk. A metszeteket Vectashield oldattal (Vector Laboratories, Burlingame, USA) fedtük le. A képeket konfokális mikroszkóppal készítettük (Zeiss LSM 710) 27
28 és a fluoreszcens képek szerkesztését, a festések intenzitásának mérését Fiji képszerkesztő programmal végeztük Áramlási citometriás gyöngy array (CBA) A 3D tüdő aggregátumok sejtjei által a felülúszóba termelt pro-inflammatorikus IL-6 és IL-8 citokinek termelődését multiplex áramlási citometriás gyöngy array (CBA Cytometric Bead Array, Human Inflammatory Cytokines Kit, BD Biosciences, San Jose, CA, USA) segítségével detektáltuk. A 3D tüdő aggregátumok 48 órás kezelése után a felülúszóikból 50 µl-t gyűjtöttünk össze, majd a kit által javasolt assay pufferrel négyszeresre higítottuk. A higított mintákhoz (50 μl) azonos mennyiségben adtuk hozzá a fluoreszcens citokin gyöngy szuszpenziót (50 μl) és a humán gyulladásos citokin-fikoeritrin detekciós reagenst. A mintákat 3 órán keresztül sötétben, szobahőmérsékleten inkubáltuk. A gyöngyöket a kit által biztosított mosó pufferben 1500 rpmn 10 percig centrifugáltuk, majd a lecentrifugált gyöngyöket 300 µl mosó pufferben felszuszpendáltuk. A jelölt gyöngyöket BD FACS DIVA V6 szoftverrel ellátott FACS Canto II áramlási citométer készülékkel (BD Immunocytometry Systems, Erembodegen, Belgium) mértük le és az adatokat FCS Express V3 szoftverrel értékeltük ki Extracelluláris vezikulák izolálása humán szérum mintákból Az extracelluláris vezikulák izolálásához 5 COPD-ben szenvedő beteg és 5 egészséges donor szérumát használtuk fel. Az izoláláshoz a totál exoszóma izoláló kitben (Invitrogen, Thermo Fisher Scientific, Waltham, USA) a gyártó által javasolt protokollt követve a szérumokból vezikulákat izoláltuk. A fagyasztott szérum mintákat szobahőmérsékleten kiolvasztottuk. A mintákat 300 g-n 30 percig centrifugáltuk a sejtmaradványok eltávolítása érdekében, majd az így kapott felülúszót ismét lecentrifugáltuk 2000 g-n, 4 C-on, 30 percig és a pelletben kapott onkoszómákat 200 µl PBS-ben felszuszpendáltuk, majd felhasználásig -80 C-on tároltuk. A centrifugálás után kapott felülúszót a totál exoszóma izoláló reagenssel vortex segítségével összekevertük, ezután 4 C-on 30 percig inkubáltuk, majd g-n, 10 percig, szobahőmérsékleten centrifugáltuk. Az exoszómákat tartalmazó pelletet hideg extracelluláris vezikula szuszpenziós pufferben szuszpendáltuk és további felhasználásig -80 C-on tároltuk Extracelluláris vezikulák (EV) izolálása A549 és Wnt5a-A549 sejtvonalakból Extracelluláris vezikulákat sejtkultúra felülúszójából izoláltunk a totál exoszóma izoláló kit sejtkultúra médiumból (Invitrogen, Cat. number: ) protokollját követve. A549 és Wnt5a-A549 sejtvonalakat DMEM sejttenyésztő médiumban növesztettük kiegészítve 10%-os 28
29 FBS tartalommal standard sejttenyésztési körülmények között (37 C, 5% CO 2). A felülúszót eltávolítottuk, a sejteket PBS-el mostuk, majd FBS mentes médiumban tenyésztettük tovább őket. Két nap után az FBS mentes médiumot eltávolítottuk és a sejteket 300 g-n, 4 C-on, 10 percig centrifugáltuk a sejttörmelék eltávolítása érdekében. A felülúszót 2000 g-n, 4 C-on, 30 percig centrifugáltuk a megaszómák/onkoszómák összegyűjtése érdekében, majd totál exoszóma izoláló reagensben vettük fel őket. A mintákat 4 C-on egy éjszakán át inkubáltuk, utána 1000 g-n, 4 C-on, 1 órán keresztül centrifugáltuk. Az extracelluláris vezikulákat tartalmazó pelletet hideg PBS-ben gyűjtöttük össze és felhasználásig -80 C-on tároltuk Mikrovezikula/Exoszóma homing Az A549 és Wnt5a-A549 sejtvonalakból izolált mikrovezikulákat/exoszómákat 1 órán keresztül 4 C-on centrifugáltuk. A pelletet 1:10000-ben higított Dil fluoreszcens festéket (Molecular Probes, Thermo Fisher Scientific, Waltham, USA) tartalmazó 200 µl PBS-ben felszuszpendáltuk. A mintákat 30 percig 37 C-on inkubáltuk, majd 1000 g-n, 4 C-on, 1 óráig centrifugáltuk. A felülúszót eltávolítottuk, a pelletet pedig PBS-ben felszuszpendáltuk és BALB/c egerek farokvénájába injektáltuk. A kontroll egerek injektálása jelöletlen exoszómákkal történt. Az injektálást követően 20 órával az egereket feláldoztuk Exoszómák lokalizációjának fluoreszcens analízise BALB/c egereket farokvénájukon keresztül egyenlő mennyiségű, 200 µl PBS-ben felszuszpendált festett és festetlen exoszóma szuszpenziókkal injektáltuk. Az egereket 20 órával az exoszómák injektálása után feláldoztuk. A tüdőt, a májat, a tímuszt és a lépet eltávolítottuk a szervekben felhalmozódott exoszómák fluoreszcens detektálásához, amelyet Living Image szoftverrel felszerelt IVIS Lumina III In Vivo Imaging rendszer segítségével (PerkinElmer, Waltham, MA, USA) végeztünk. Az egerek szerveiben felhalmozódott exoszómák fluoreszcens intenzitását a festetlen, kontroll szervekhez viszonyítottuk Mikrovezikulák (exoszómák) elektronmikroszkópos detektálása A sejtvonalakból izolált mikrovezikulák elektronmikroszkópos detektálásához az exoszómákat PBS-ben szuszpendáltuk, azután 2,5%-os glutáraldehid vizes oldatában fixáltuk. A fixált exoszómákból 5 μl-t az elektronmikroszkópos hálóra mértünk, amelyet előtte vízzel átmostunk és 24 óráig hagytuk száradni. A mintákat 5%-os uranil-acetáttal 5 percig ph 7-es körülmények mellett kezeltük. Az elektronmikroszkópos hálót 80Kv-on MorgagniD transzmissziós elektronmikroszkóppal vizsgáltuk (Orvosi Biológiai Intézet és Központi Elektronmikroszkópos Laboratórium, Általános Orvostudományi Kar, Pécsi Tudományegyetem). A mikroszkóppal 29
30 készített képeket integrált MegaView III digitális kamerával (Olympus Soft Imaging Solutions GmbH; Munster, Germany) készítettük Enzim kapcsolt szendvics immunoszorbens vizsgálat (ELISA) Mega EV-t (onkoszómák) és mikro EV-t (exoszómák) 0,5%-os Tween 20-at tartalmazó PBSben oldottuk és minden mintából 100 µl-t mértünk rá az ELISA lemezre (Aviva Systems Biology WNT5A ELISA Kit, OKEH00723). Az ELISA 96-lyukú lemez Wnt5a specifikus antitesttel volt előkezelve. A fehérjeméréshez a kalibrációs minták is biztosítva voltak. A mintákat 2 órán keresztül, 37 C-on inkubáltuk, majd a lemezt háromszor mosó pufferben mostuk. Az ELISA lemezt biotinilált Wnt5a specifikus detektor antitesttel további 1 óráig, 37 C-on inkubáltuk. Az újabb mosási lépés után a lemezt Avidin-HRP konjugátummal 1 óráig, 37 C-on inkubáltuk. Az újabb mosási lépés után TMB (3,3,5,5 -Tetramethylbenzidine) szubsztrátot adtunk a mintákhoz és 15 percig, 37 C-on, sötétben inkubáltuk. A reakciót leállítottuk és a színreakciót 450 nm-en mértük. Az eredményeket a standard görbéhez viszonyítva számoltuk Tüdő epiteliális sejtek izolálása primer humán tüdőszövetből Humán primer tüdő epiteliális sejteket izoláltunk lobektomián átesett, normál tüdőfunkcióval rendelkező betegekből (n=26) származó tüdő szöveti mintákon. Minden beteg beleegyező nyilatkozattal adta írásba, hogy mintáikat és klinikai adataikat kizárólag kutatási célokra felhasználhatjuk (etikai engedély 07/MRE08/42). A sejtek izolálását egy korábban leírt protokollt követve végeztük (104). A szöveti mintákat sós oldatba merítettük, majd meleg tripszinnel emésztettük, amit további finom emésztés követett DNáz (Sigma-Aldrich, St. Louis, Missouri, USA) és magzati borjúszérum (FCS) (Lonza, Basel, Svájc) jelenlétében. Az így kapott sejtszuszpenziót átszűrtük, majd a későbbiekben az izolált sejteket további fluoreszcencia aktivált sejtválogatásra és analízisre (FACS), vagy hosszútávú kultúrában való fenntartásra használtuk fel Primer tüdő epiteliális sejtek in vitro transzdifferenciációja (TD) Tüdő epiteliális sejteket kollagénnel bevont sejttenyésztő edényekre ültettünk ki és 10% FCS-t tartalmazó DCCM-1 médiumban (Biological Industries Ltd. Kibbutz Beit-Haemek, Izrael) 3-6 napig tartottuk őket kultúrában. A 3. és a 6. napon a sejteket lizáltuk és belőlük total RNS-t izoláltunk, majd a továbbiakban mikroarray és kvantitatív valós idejű PCR analízisekre használtuk fel. 30
31 3.23. Mikroarray analízis A betegekből (n=3) származó, 3 és 6 napig in vitro körülmények között kultúrában fenntartott primer sejtekből RNS-t izoláltunk, majd cdns-t készítettünk és fluoreszcensen jelöltük GeneChip WT terminális jelölő Kit (Affymetrix, Santa Clara, USA) segítségével. A fragmentált cdns-t a Humán Gén 1.0 ST array-hez (Affymetrix, Santa Clara, USA) hibridizáltuk. A microarray-ből a próba sejt intenzitási adatokat (CEL, cell intensity data) analizáltuk Expressziós Console szoftver segítségével Protein ANalysis THrough Evolutionary Relationships (PANTHER) A PANTHER klasszifikációs rendszert a fehérjék (és azok génjeinek) klasszifikálására tervezték a high throughput analízisek elősegítésére. A módszer részletes leírása korábbi publikációkban elérhető. (105) (106). A PANTHER része a Gén Ontológia Filogenetika Annotációs Projektnek Mesterséges neuronális hálózat analízis (ANN - artificial neural network) A Wnt jelátviteli útvonalhoz kapcsolódó ATII-ből ATI irányú TD folyamatban résztvevő molekulák vizsgálatához mesterséges neuronális hálózat (ANN) (Neurosolutions 6, NeuroDimension Inc.) szoftvert használtunk (107) (108). A génexpressziós adatokat poolozott cdns minták felhasználásával, Affymetrix array segítségével határoztuk meg ATII-es sejtekből, melyeket kontrolloknak és ATI-es sejtekből, melyeket a kezelt mintáknak tekintettünk. Egy átlag szenzitivitást határoztunk meg, mint 1.0 és minden más szenzitivitás értéket is eszerint ábrázoltunk hőtérkép formájában Statisztikai analízisek A statisztikai elemzés GraphPad 6 szoftverrel történt, független mintás T-próba és egy-utas ANOVA (Bonferroni post hoc) tesztek segítségével, valamint ezen tesztek nem parametrikus megfelelőivel (Kolmogorov-Szmirnov-próba). Az adatokat minden esetben átlag ± standard hiba (SEM) ábrázoltuk és a p <0.05 értékeket tekintettük szignifikáns eredménynek. 31




32 4. Eredmények 4.1. A dohányzás kiváltotta gyulladásos folyamatok és a Wnt5a kapcsolata in vivo egérmodellben A cigarettafüst szisztémás gyulladást indukáló szerepének bizonyítására két hónapos teljes test dohányfüst expozíciónak kitett egerek tüdejét hematoxilin-eozin festéssel vizsgáltuk, ahol jól láthatók a kitágult alveolusok, valamint peribronchiális és perivaszkuláris gyulladás jelenléte makrofág, neutrofil és limfocita beszűrődésekkel (5.ábra). Kontroll 1000µm 100µm 2 hónap dohányzás 100µm 1000µm 5. ábra: Hematoxilin-eozin festés egerek tüdejéből paraffinba ágyazott egészséges kontroll metszeteken, illetve 2 hónap dohányoztatás után. 10x nagyítás, 20x nagyítás, lépték:100 µm, 1000 µm. A nyilak a limfoid beszűrődésre mutatnak. Továbbiakban az in vivo egér modellrendszereinkben a gyulladásban kulcsszerepet játszó Wnt5a fehérje emelkedését mértük 2 hónapos CF expozíció hatására, melyet Western-blot analízissel és paraffinba ágyazott egértüdő metszeteken végzett immunfluoreszcens festések segítségével detektáltunk (6. ábra a, b). 32
 Wnt5a fehérje expressziójának vizsgálata dohányoztatott egértüdőben Westernblottal kimutatva a β-aktin fehérjéhez")
 Wnt5a (piros) fehérje expressziójának vizsgálata immunfluoreszcens festéssel kontroll és 2 hónapig dohányoztatott")
 vizualizáltuk. Lépték:200µm.")

33 a) Wnt5a 49kDa β-aktin 42kDa Kontroll 1 hónap 2 hónap b) Kontroll 2 hónap dohányzás 200µm 200µm 6. ábra a) Wnt5a fehérje expressziójának vizsgálata dohányoztatott egértüdőben Westernblottal kimutatva a β-aktin fehérjéhez viszonyítva. b) Wnt5a (piros) fehérje expressziójának vizsgálata immunfluoreszcens festéssel kontroll és 2 hónapig dohányoztatott paraffinos egértüdő metszeteken, az epitél sejteket (zöld) EpCAM + antitesttel jelöltük, míg a sejtmagokat DAPI festékkel (kék) vizualizáltuk. Lépték:200µm. Hogy részletesebben megismerjük a tüdőben a dohányzás hatására bekövetkező molekuláris változásokat, további kísérleteinkben a tüdőt nagy számban felépítő sejtjeket külön is vizsgáltuk. Ennek eredményeként az egerek tüdejéből tisztított epitél (EpCAM + ) sejtek számában csökkenő tendenciát mértünk dohányzás hatására (7. ábra a.), míg a leukocita (CD45 + ) populációban a CD45 + sejtek számának szignifikáns emelkedését detektáltuk 2 hónapos dohányoztatás hatására (7. ábra b.). 33
34 a) b) 7. ábra a) C57BL/6 egerekben az EpCAM + sejtek számának változása kontroll és dohányoztatott egerek tüdejében 1 és 2 hónap elteltével. b) C57BL/6 egerekben a CD45 + sejtek számának változása kontroll és dohányoztatott egerek tüdejében 1 és 2 hónapot követően (n=3). Átlag ± SEM, Student féle t-próba, ahol a különbség szignifikáns, ha *p<
35 Az EpCAM + (8. ábra a.) és a CD45 + (8. ábra b.) sejtpopulációban 2 hónapos dohányoztatás következtében a Wnt5a mrns szignifikáns emelkedését mutattuk ki valós idejű, kvantitatív PCR segítségével. a) b) 8. ábra: a) Wnt5a mrns expressziójának változása dohányoztatott C57BL/6 egerek tüdejéből kiszortolt EpCAM + epitél sejtekben a kontroll mintákban mért Wnt5a expresszióhoz viszonyítva. b) CD45 + sejtekben mért Wnt5a relatív génexpressziós változása kontroll, 1 és 2 hónapig dohányoztatott C57BL/6 egerek tüdejében (n=3). Átlag ± SEM, ANOVA teszt, ahol a különbség szignifikáns, ha *p<0.05, **p<0.01, *** p<0.001, ****p<
36 A CD45-nek különböző izoformái léteznek, melyek sokféle leukocita populáción expresszálódnak, többek között a T és B limfocitákon, vagy antigén prezentáló sejteken, de további kísérleteinkben mi csak a makrofágokkal vizsgálódtunk tovább, hiszen a makrofágok a gyulladásban központi szerepet töltenek be az antigén prezentáció, fagocitózis és az immunmoduláció folyamatainak irányításában, amely funkciókat számos citokin és növekedési faktor termelésén keresztül hajtják végre. Ezek alapján elmondhatjuk, hogy a makrofágok kritikus szerepet töltenek be a gyulladás kialakításában és fenntartásában egyaránt, Így a makrofágok gyulladásban betöltött molekuláris mechanizmusának megismerése kulcsfontosságú ezen folyamatok mélyebb megismerése érdekében (109) A dohányoztatás kiváltotta gyulladás és a Wnt5a kapcsolata 3D humán tüdőmodellben A CF Wnt5a indukáló hatásának humán tüdőszöveten való teszteléséhez in vitro humán 3D tüdőmodellt használtunk. A 3D humán tüdő aggregátumok SAEC, NHLF, HMVEC-L primer sejteket tartalmaztak humán vérből izolált monocitából differenciált makrofágok jelenlétében, illetve makrofágok jelenléte nélkül, melyeket CFE-vel 48 óráig kezeltünk. A kezelés hatására szignifikánsan emelkedett Wnt5a mrns expressziót tapasztaltunk (9. ábra), immunfluoreszcens festéssel pedig növekedett Wnt5a fehérjeexpressziót detektáltunk, de kizárólag a makrofágot tartalmazó aggregátumokban (10. ábra a, b, c.). 9. ábra: Wnt5a mrns relatív génexpressziós változása kontroll és 48 óráig cigarettafüst extraktummal kezelt 3-dimenziós makrofágot tartalmazó és makrofágot nem tartalmazó humán tüdő aggregátumokban (n=3). Átlag ± SEM, ANOVA teszt, ahol a különbség szignifikáns, ha **p<0.01, *** p<


 3D +")


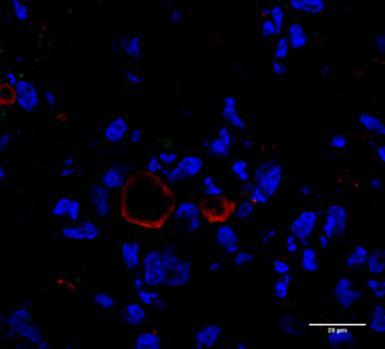
37 a) 3D Kontroll 100µm 20µm 3D + CFE 100µm 20µm b) 3D Kontroll + MO 100µm 20µm 3D + MO + CFE 100µm 20µm 37
38 c) 10. ábra: a) Wnt5a (zöld) és citokeratin 7 (piros) epiteliális marker fehérjeexpressziójának változása 48 órás CFE kezelés hatására 3-dimenziós humán tüdő aggregátumokban, ahol a sejtmagokat DAPI festékkel (kék) vizualizáltuk. b) Wnt5a (zöld) és citokeratin 7 (piros) epiteliális marker fehérjeexpressziójának változása 48 órás CFE kezelés hatására 3- dimenziós humán tüdő aggregátumokban makrofágokkal, ahol a sejtmagokat DAPI festékkel (kék) jelöltük. c) Wnt5a fehérje intenzitásváltozásának ábrázolása kontroll és CFE kezelt humán tüdő aggregátumokban ImageJ képszerkesztővel számolva (n=3). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p<0.05, **p<0.01, **** p< X és 20X nagyítás, lépték: 100 µm, 20 µm. 38
39 In vitro gyulladásos modellrendszerünk alátámasztásaként elmondhatjuk, hogy CFE kezelés után a makrofágot tartalmazó szferoidokban emelkedettebb gyulladásos citokin expressziókat mértünk, mint a kontroll, makrofágot nem tartalmazó szferoidokban (11. ábra). 11. ábra: IL-6 és IL-8 pro-inflammatorikus citokinek génexpressziójának változása 48 órás CFE kezelés hatására primer humán sejtekből álló 3D humán tüdő aggregátumokban (n=3). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p<0.05. Ebből következtethetünk arra, hogy a dohányzás által kiváltott gyulladásos folyamatok összefüggésben állhatnak a makrofágok jelenlétével a tüdőben. A gyulladásos faktorok aktiválódása bizonyos, a szervezetet érő káros ágensek eliminálásának érdekében történik, és sérülés vagy fertőzés esetében segít az immunrendszer funkciójának javításában és a homeosztázis helyreállításában. A makrofágok különböző szinteken vesznek részt a gyulladásos folyamatok szabályozásában, melyre példa, hogy a szövetspecifikus makrofágok és az aktivált monociták különböző aktivációs állapotot képesek felvenni. Bár a folyamat még nem teljesen ismert, az M1 és M2 polarizációs útvonalak adhatnak magyarázatot a makrofágok aktivációjának változására a gyulladásos folyamatok során (110). Ahhoz, hogy a dohányzás immunmoduláló hatásának teszteléséhez megintcsak 3D humán tüdő aggregátum kultúrákat alkalmaztunk, melyeket 48 órán keresztül CFE kezelésnek tettünk ki, majd a gyulladásos citokin és a nem-kanonikus Wnt5a génexpresszió változásokat monitoroztuk. CFE kezelés hatására szignifikáns IL-6 emelkedést tapasztaltunk az aggregátum kultúrákban a kontroll mintákhoz képest, melyet a makrofág jelenlétében és makrofágok hiányában is detektáltunk (11. ábra). Az aggregátum kultúrákon 48 órás humán rekombináns Wnt5a kezelést (1 μg/ml) is 39
40 végeztünk, melynek hatására szignifikáns IL-6 és IL-8 emelkedést detektáltunk, kizárólag a makrofágot tartalmazó aggregátumokban, mely a Wnt5a-nak a pro-inflammatorikus folyamatok indukálásában játszott szerepére utal nem csak makrofágokban, hanem akár epitéliális és endoteliális sejtek jelenlétében is (12. ábra). Relatív mrns expresszió 12. ábra: IL-6 és IL-8 gyulladásos citokinek génexpressziós szintjei 48 órás humán rekombináns Wnt5a fehérje kezelés hatására primer humán sejtekből álló 3D humán tüdő aggregátumokban (n=3). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p< A makrofágok polarizációjának és a Wnt jelátviteli útvonalnak a kapcsolata in vitro dohányoztatás hatására Az in vivo és in vitro dohányoztatás kiváltotta dinamikusan változó molekuláris folyamatok részletezésére 2-dimenziós kultúrában tenyésztett makrofágokat CFE-vel kezeltünk, majd Wnt szignalizációs TaqMan Array segítségével a Wnt gének expressziós mintázatát detektáltuk. A kezeletlen kontrollokhoz képest a dohányoztatott makrofágokban a nem-kanonikus Wnt jelátviteli molekulák, Wnt5a, Wnt5b, Wnt11, és a kanonikus Wnt jelátvitel gátlását végző Dickkopf 2 (DKK-2) inhibitor emelkedését mértük (13. ábra a, b, c.). 40
 Wnt mrns expressziós változások ábrázolása logaritmikus skálán 3 óra CFE kezelés hatására előre összeállított TaqMan lemezen (n=5).")
41 a) b) c) 13. ábra: a) Wnt jelátvitel génjeinek mrns expressziós változásainak ábrázolása hőtérképen humán makrofágokban 3 óra CFE kezelés hatására előre összeállított TaqMan lemezen (n=5). b) Wnt mrns expressziós változások ábrázolása logaritmikus skálán 3 óra CFE kezelés hatására előre összeállított TaqMan lemezen (n=5). c) A nem-kanonikus Wnt5a, Wnt5b és Wnt10a, illetve a kanonikus Wnt10b gének relatív génexpresszióinak változása látható 3 óra CFE kezelés hatására a kontroll mintákhoz képest. Belső kontrollként β-aktint alkalmaztunk (n=5). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p<
42 A specifikus Wnt jelátviteli molekulák mellett a cink-ujj domén fehérjecsaládba tartozó CXXC4 transzkripciójának aktiválódását is mértük. Az expressziós mintázat alapján a CXXC4, mint a kanonikus Wnt jelátviteli útvonal antagonistája működhet. Emellett a CSNKD1, a kazein kináz I (CKI) gén család tagja, a Transducin Like Enhancer of Split 4 (TLE4) és a Wnt10 mrns expresszióinak növekedését is mértük. Amíg a CSNKD1 alapvetően szerin-treonin protein kináz, melynek szerepe van a sejtnövekedésben és a sejt túlélésben, addig a TLE4 egy transzkripciós korepresszorként működik. A Wnt10a expressziójának növekedése jelentős szerepet játszhat a szignalizációban, hiszen a Wnt10a a Wnt-β katenin-tcf jelátviteli útvonal aktiválódásán keresztül fejti ki hatását a karcinogenezisben. Emellett az Axin2 és a kanonikus Wnt1 expressziójának csökkenését párhuzamosan detektáltuk. Feltételezhetjük, hogy az Axin2 csökkent transzkripciójának szerepe lehet a β-katenin stabilitásának szabályozásában. Továbbá a Transducin Like Enhancer of Split 2 (TLE2) és a szekretált Frizzled kapcsolt fehérje 5 (Secreted Frizzled Related Protein 5, SFRP5) csökkent expressziója is alátámasztja a makrofágokban a Wnt szignalizációban végbemenő nagymértékű változásokat, amelyeket CSE kezelés hatására tapasztaltunk. Mind a kanonikus, mind a nem-kanonikus Wnt jelátviteli útvonalak szabályozásában zajló változások a PPAR-γ expressziójában és aktivitásában is módosulást eredményeznek (31). Megvizsgáltuk, hogy a Wnt szignalizációban végbemenő változások milyen mértékben hatnak a makrofágok dinamikus polarizációjára. A pro-inflammatorikus citokinek jellemzően az M1 fenotípushoz tartoznak, míg az anti-inflammatorikus, de tumorindukáló hatással is rendelkező M2 fenotípus (47) a PPAR-γ expressziójának és aktivitásának vonatkozásában már ismert. A PPAR-γ cigarettafüst által indukált Wnt5a emelkedésben és az általa kiváltott proinflammatorikus citokinek termelésében játszott szerepének vizsgálatához humán tüdő aggregátum kultúrákon Wnt5a kezelést és CF jelenlétében, illetve hiányában adott PPAR-γ agonista, roziglitazon (RSG) és antagonista, GW9662 kezelést végeztünk (14. ábra a, b.). 42
43 a) b) 14. ábra: a) IL-23 és IL-10 mrns expressziók változása 48 órás humán rekombináns Wnt5a kezelést követően 3D tüdő aggregátumokban. b) IL-23 mrns és IL-10 expressziós szintje 48 órás CFE, illetve kombinált CFE és PPAR-γ specifikus agonista (RSG) és GW9662 antagonista kezelést követően (n=3). A relatív génexpressziók a kezeletlen kontrollhoz viszonyítva láthatóak. Átlag ± SEM, Student féle t-próba, ahol a különbség szignifikáns, ha *p<0.05, **p<
44 Az M1 fenotípusra jellemző pro-inflammatorikus citokinek az IL-6, IL-8, IL-23, IL-1β, míg az M2 fenotípusra jellemzőek az anti-inflammatorikus citokinek, mint az IL-10 és TGF-β (34). Az anti-inflammatorikus PPAR-γ gátlása az antagonista, GW9662-vel, csökkentette az IL-10 expresszióját, amely az anti-inflammatorikus M2 fenotípus markere (14. ábra b.). A citokin termelés egy dinamikus folyamat, melynek monitorozásához humán vérből izolált monocitából származó makrofágokat CFE-vel kezeltünk 3, illetve 48 órán keresztül, majd a PPAR-γ mennyiségének változásait analizáltuk. A 3 órás kezelés után a PPAR-γ expressziója szignifikánsan emelkedett, ezzel szemben 48 óra után a PPAR-γ szintje a kezeletlen minták szintjére csökkent, mely arra utal, hogy az anti-inflammatorikus PPAR-γ expressziója önmagában nem tartaná fent az elnyújtott gyulladásos stimulust (15. ábra). 2.5 *** Relatív PPAR- mrns expresszió h Kontroll 3h CFE 48h CFE 15. ábra: PPAR-γ relatív mrns expressziós változásai 3 órás és 48 órás CFE kezelés hatására humán makrofágokban (n=3). A relatív génexpressziós változások a kezeletlen kontrollhoz viszonyítva láthatóak. Átlag ± SEM, Student féle t-próbát alkalmaztunk statisztikai próbaként, ahol a különbség szignifikáns, ha *** p<
45 4.4. Az extracelluláris vezikulák és a gyulladásos citokinek szerepe a szisztémás gyulladás kialakulásában A krónikus CF expozíció növeli a Wnt5a fehérje mennyiségét a COPD-s betegek szérumából izolált extracelluláris vezikulákban. Mivel a lipo-glikoprotein Wnt ligandok erősen hidrofób jelleggel rendelkeznek (48), így transzportjuk a test vizes kompartmentjeiben extracelluláris vezikulákon belül, illetve azok membránjába illeszkedve történhet (49). Az EV-ok (megaszómák, más néven onkoszómák vagy mikroszómák, más néven exoszómák) a sejtek közötti kommunikációnak egy újonnan felfedezett módja, mely befolyásolhatja a Wnt jelátviteli útvonalra kifejtett hosszútávú hatásokat (50) (51). A fentieken alapulva feltételezhetjük, hogy a Wnt5a és a pro-inflammatorikus citokinek alacsony szintje mérhető a humán szérumból izolált extracelluláris vezikulákban (52)(53). Ha az extracelluláris vezikulák képesek egy adott szervből egy másik szervbe, vagy az emberi szervezetet felépítő bármilyen sejtbe juttatni a bennük található tartalmakat, ez a transzport folyamat magyarázatot adhat a COPD-t érintő súlyos szisztémás gyulladásos környezet kialakulására. Hogy bizonyítsuk a hipotézisünket nagyobb megavezikulákat/onkoszómák és kisebb, mikrovezikulákat/exoszómákat izoláltunk COPD-betegek és egészséges, korban megegyező emberek szérumából. Pozitív kontrolloknak nem kissejtes karcinóma sejtvonalat (A549) és transzgenikus, Wnt5a overexpresszáló A549 sejtvonalat (Wnt5a-A549) használtunk. A Wnt5a fehérje mennyiségének meghatározásához ELISA kitet, a pro-inflammatorikus citokinek mérésére CBA array-t alkalmaztunk és a gyulladásos markerek mennyiségét áramlási citometria segítégével detektáltuk. Amíg a Wnt5a expressziója sem a COPD-betegek, sem az egészséges kontrolljaik szérumában nem volt mérhető, a COPD-betegek szérumából izolált apoptotikus testekben/onkoszómákban a Wnt5a overexpresszáló A549 sejtvonalban mért Wnt5a mennyiségével megegyező Wnt5a expressziós szintet detektáltunk (16. ábra). 45
46 16. ábra: Wnt5a fehérje mennyiségének változása A549 és Wnt5a túltermelő A549 tüdőkarcinóma sejtvonal felülúszójából izolált onkoszómákban és exoszómákban poolozott mintákon, illetve egészséges és COPD-betegek poolozott szérumában Wnt5a specifikus ELISA kit segítségével mérve (n=5). Hasonlóan a Wnt5a változásához, a pro-inflammatorikus citokinek szintje sem volt detektálható a szérum mintákban (az adatok nincsenek ábrázolva), ezzel ellentétben az onkoszómákban és az exoszómákban is kiemelkedően magas gyulladásos citokin szinteket mértünk, legfőképpen az IL-8 citokin esetében (17. ábra a, b.). 46
47 a) b) 17. ábra: a) Gyulladásos citokinek fehérjemennyiségének változásai poolozott humán egészséges és COPD-s betegek szérumából izolált onkoszómákban CBA módszerrel mérve (n=5). b) Gyulladásos citokinek fehérjemennyiségének változásai poolozott humán egészséges és COPD-s betegek szérumából izolált exoszómákban CBA módszerrel detektálva (n=5). 47
48 A fenti eredmények tükrében a következő kísérlet során arra kerestük a választ, hogy ezek a sejtszintű üzenetek valóban képesek-e elérni más szerveket? Ehhez EV-t izoláltunk Wnt5a overexpresszáló A549 sejtek által termelt felülúszóból és az exoszómák vizualizálására egy fluoreszcens, DiI nevezetű festéket alkalmaztunk, majd a megfestett exoszómákat BALB/c egerek farokvénájába injkektáltuk. Az injektálást követő 20. órában az egereket feláldoztuk, a szerveiket pedig eltávolítottuk, hogy az ott esetlegesen felhalmozódott fluoreszcensen jelölt exoszómák helyét a szervekben nyomon követhessük. Az injektálást követően a jelölt exoszómákat a legnagyobb mennyiségben a májban, majd a tímuszban és a tüdőben figyeltük meg, illetve kisebb mennyiségben a lépben is detektálhatóak voltak. Összefoglalva eredményeinket feltételezhetjük, hogy a gyulladásos citokineket tartalmazó mikrovezikulák/exoszómák képesek a szervezet bármely szervébe eljutni és ott egy lokális dúsulást kialakítani, ahogy azt alább láthatjuk az egerek szerveiről készített felvételen (18. ábra a, b.). 48
49 a) b) b) Átlagos sugárzási hatékonyság (p/s/cm 2 /sr)/ W/cm 2 ) Tímusz Tüdõ Máj Lép 18. ábra: a) BALB/c egerek injektálását követően 20 órával a Wnt5a túltermelő A549 sejtvonal felülúszójából izolált, fluoreszcensen jelölt exoszómák vizualizálása az egerek szerveiben (n=3). b) A fluoreszcensen jelölt Wnt5a túltermelő A549 tüdőkarcinóma sejtvonal felülúszójából izolált BALB/c egerekbe injektált exoszómák átlagos sugárzási hatékonysága az egerek szerveiben (n=3). 49

50 4.5. Wnt jelátvitel aktiválódása az ATII-ATI transzdifferenciáció során in vitro A fenti kísérletekből is látszik, hogy a dohányzás a tüdőben nagymértékű molekuláris változást indukál gyulladásos folyamatok aktiválódásával, melynek eredménye a tüdő szövetét érintő, hosszútávú, súlyos szöveti destrukció, mely sejtszinten strukturális változásokkal jár. Ennek okán a tüdő szöveti regenerációban szerepet kapó transzdifferenciációs folyamatok pontosabb megismerése kulcsfontosságú a károsodott tüdő helyreállítása, terápiája érdekében. Ezért további kísérleteink során a Wnt jelátvitel szerepét vizsgáltuk az ATII-ATI transzdifferenciáció folyamatában. Elsőként Affymetrix array segítségével 1527 gént analizáltunk (donor=3) és a PANTHER (Protein ANalysis THrough Evolutionary Relationships) program segítségével megállapítottuk, hogy azok közül a gének közül, melyek szignifikáns változást mutattak, 73 gén a Wnt jelátviteli úthoz tartozik (19. ábra). A program által számolt arányokat kördiagrammon ábrázoltuk, ahol az expressziós változást reprezentáló gének mennyiségét számokkal ábrázoltuk. A mérés során a 6 napos kultúrákban mért expressziós változásokat a 3 napos kultúrákhoz viszonyítottuk. 19. ábra: Affymetrix mikroarray PANTHER analízisének összefoglalása kördiagrammon. 50
.")
51 A Wnt jelátvitelben a Wnt4, Wnt5a, Wnt7a ligandok és a Fzd 1, Fzd2 és Fzd7 receptorok mrns szintjében szignifikáns változásokat detektáltunk az Affymetrix array segítségével. Az Affymetrix eredményeket kvantitatív valós idejű PCR-al is validáltuk (21. ábra b.). a) b) 5 c) * * ATII-es típusú sejtek ATI-es típusú sejtek Relatív mrns expresszió * 0 Wnt4 Wnt5a Wnt7a Fzd1 Fzd2 Fzd7 20. ábra: a) Az ATII-ATI TD során szignifikáns változást mutató Wnt jelátvitel asszociált gének hőtérképen való ábrázolása (n=3). b) In vitro transzdifferenciációt követően a Wnt ligandok, Wnt4, Wnt5a és Wnt7a, illetve a Wnt receptorok közül a Fzd1, Fzd2 és Fzd7 génexpresszióinak meghatározása mikroarray segítségével (n=3). c) A mikroarray adatok megerősítéseként a korábban eltérést mutató Wnt molekulák és receptoraik expressziós mintázatának vizsgálatát végeztük in vitro ATII-ATI transzdifferenciált sejtekben (n=11). Statisztikai próbaként független mintás t-próbát alkalmaztunk, ahol * p<0.05, ** p<
52 Az in vitro kísérletek során detektált génexpressziós változásokat izolált primer humán ATI és ATII-es sejtek mrns szintjéhez viszonyítottuk. Az alveoláris típusú sejtek karakterizálásához az ATI-es sejtekhez AQP5 és RAGE markereket használtuk, az ATII-es típusú sejtek esetében differenciációs markerként az SFTPC-t alkalmaztuk (21. ábra a.). Bár az SFTPC szintjében nem detektáltunk szignifikáns változást, az ATI-es AQP5 és RAGE markerek szignifikáns emelkedést mutattak az ATI populációban az ATII-es populációhoz képest (21. ábra a.). A Wnt receptorok mrns szintjeiben nem láttunk különbséget, míg a Wnt4, Wnt5a és Wnt7a ligandok expresszója szignifikánsan magasabb volt az ATI-es szortolt sejtekben, mint az ATII-es sejtpopulációban (21. ábra b.). a) Relatív mrns expresszió b) 21. ábra: a) ATII-es és ATI-es típusú sejtek differenciációs markereinek expressziós változásai frissen izolált human tüdő epiteliális sejtekben, n=12. b) Wnt ligandok és receptoraik expressziós szintjeinek változása frissen izolált tüdő epiteliális sejtekben (n=12). Statisztikai próbaként független mintás T-próbát alkalmaztunk, * p<0.05, ** p<
53 A Wnt molekulák ATII-ATI TD-ban játszott szerepét további in vitro 3-dimenziós aggregátum kokultúrákban teszteltük, melyek SAEC, illetve NHLF primer humán sejtekből állnak. Korábbi kísérleteink bizonyították, hogy ezekben a kokultúrákban az epiteliális sejtek ATII-es típusú differenciáción mennek keresztül és SFTPC-t expresszálnak. Ebben a kísérleti rendszerben a rekombináns humán Wnt4 és Wnt5a kezelések csökkentették az SFTPC mrns és fehérje expresszióját egyaránt (103). Ezt a jelenséget tovább vizsgálva jelenlegi kutatásunkban rhwnt4, rhwnt5a és rhwnt7a kezelést alkalmaztunk aggregátum kultúrákban, hogy az SFTPC mrns szintjének csökkenését indukáljuk. Eredményeink alapján azonban, kizárólag a rhwnt7a kezelés emelte az ATI marker, AQP5 mrns (23. ábra a.) és fehérjeszintjét (23. ábra b, c.). a) 53
 ATII-es és ATI-es típusú sejtek differenciációs markereinek expressziós változása 3D humán tüdő aggregátum kokultúrákban Wnt ligandokkal történt")
 3D humán tüdő aggregátumok szöveti metszeteken immunfluoreszcens festés segítségével anti- AQP5 antitesttel (piros) az ATI-es típusú sejteket és DAPI")
54 b) Kontroll rhwnt7a c) Integrált denzitás (%) 22. ábra: a) ATII-es és ATI-es típusú sejtek differenciációs markereinek expressziós változása 3D humán tüdő aggregátum kokultúrákban Wnt ligandokkal történt kezelés hatására. b) 3D humán tüdő aggregátumok szöveti metszeteken immunfluoreszcens festés segítségével anti- AQP5 antitesttel (piros) az ATI-es típusú sejteket és DAPI festékkel a sejtmagokat vizualizáltuk, skála: 50 µm, három biológiai replikátum reprezentatív festése, n=3. c) AQP5 immunfluoreszcens festésének denzitometriai elemzése, (n=3). Statisztikai próbaként független mintás t-próbát használtunk, * p<0.05, ** p<




55 Az előzőekben kapott eredmények alapján a Wnt7a indukálta intracelluláris jelátviteli folyamatokat vizsgáltuk tovább humán tüdő aggregátum kultúrákból készített metszeteken, melyeken β-katenin és foszfo-β-katenin immunfluoreszcens festést végeztünk (23. ábra a, b). A β-katenin szint emelkedett Wnt7a hatására, a β-katenin foszforilációja pedig szignifikánsan csökkent, ezzel bizonyítva a TD folyamatban résztvevő intracelluláris folyamatok komplexitását (23. ábra a, b). a) Kontroll rhwnt7a 55
56 b) 23. ábra: a) 3D tüdő aggregátumokat rekombináns humán Wnt7a fehérje kezelése anti-βkatenin (zöld) és anti-foszfo β-katenin (piros) antitesttel, a sejtmagokat DAPI festékkel (kék) vizualizáltuk. Skála: 50 µm. Három biológiai replikátum reprezentatív festése, (n=3). b) β- katenin és foszfo β-katenin immunfluoreszcens festésének denzitometriai elemzése, (n=3). Statisztikai próbaként független mintás t-próbát használtunk, * p<

57 4.6. Az ATII-ATI TD-ban szerepet játszó Wnt jelátviteli molekulák hierarchiájának definiálása a mikroarray adatok ANN analízisével A megváltozott Wnt mikrokörnyezetben résztvevő molekuláris célpontok meghatározására ANN analízist végeztünk, melynek segítségével az ATII-ATI TD folyamat részeként a nemlineáris kapcsolatokat definiálhatjuk. Az analízis során több molekulát is találtunk, a trombospondin 1 (THBS1), transzglutamináz-2 (TGM2), integrin alfa V (ITGAV), epiteliális membrán fehérje 2 (EMP2), citokróm P450 család 4, alcsalád B, polipeptid 1 (CYP4B1) és ankyrin repeat domain 1 (ANKRD1) faktorokat, melyek a módosult Wnt jelátviteli útvonal célpontjai az ATII-ATI TD során (24. ábra). 24. ábra: Az ATII-ATI es transzdifferenciációban szerepet játszó Wnt jelátviteli útvonal génjeinek és azok közötti kapcsolatoknak a feltérképezése ANN analízis segítségével. Az Affymetrix elemzés alapján a THBS1, EMP2, ITGAV, CYP4B1 és ANKRD1 mrns szintjei szignifikánsan emelkedtek, míg a TGM2 szignifikánsan csökkent a TD során a kontrollhoz képest. A fent említett faktorok közül a legnagyobb mértékben a THBS1 és EMP2 voltak érintettek a Wnt jelátviteli molekula rendszerek változásának hatására (25. ábra). 57
58 25. ábra: A célgének expressziós mintázatának ábrázolása Affymetrix analízis segítségével (n=3). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p<0.05, ** p<0.01. Annak érdekében, hogy az ANN analízis során definiált gének működését és azok funkcióját specifikus Wnt ligandokhoz kapcsolhassuk, 3D aggregátumokat 48 órán keresztül rekombináns humán Wnt fehérjékkel kezeltünk (rhwnt4, rhwnt5a and rhwnt7a) in vitro körülmények között, majd a kokultúrákból izolált cdns mintákon qrt-pcr mérést végeztünk. Az mrns expresszió szintjeinek változásait detektálva azt találtuk, hogy csak az ITGAV mrns mennyisége csökkent kizárólag rhwnt5a kezelés hatására, de a rhwnt4, illetve a rhwnt7a kezelésnek nem volt hatása. Ebből azt a következtetést vonhatjuk le, hogy a TD során egyes Wnt ligandok különböző célpontokkal rendelkeznek (26. ábra). 58
59 26. ábra: A Wnt jelátviteli útvonal target gének expressziós változásai 3D tüdő aggregátum kultúrákban Wnt ligand kezelések hatására (n=3). Átlag ± SEM, ANOVA teszt, ahol a kontroll mintákhoz viszonyított különbség szignifikáns, ha *p<0.05. A rekombináns humán Wnt5a kezelt, 3D sejt aggregátumokból készített metszeteken sejttípus specifikus ITGAV fehérje expresszió emelkedést tapasztaltunk (27. ábra a, b.), melyet az aggregátumokban centrálisan elhelyezkedő mezenhimális fibroblasztokban detektáltunk, míg az aggregátumok külső rétegében található epiteliális sejtekben az ITGAV fehérje expressziója szignifikánsan kevesebb volt (27. ábra c.). 59



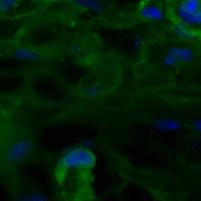
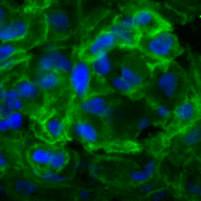
60 a) Kontroll rhwnt4 rhwnt5a rhwnt7a 60
61 b) c) 27. ábra: a) Rekombináns Wnt4, Wnt5a, Wnt7a fehérjével kezelt 3D tüdő szöveti aggregátumok immunfluoreszcens festése, ahol az ITGAV fehérjét anti-itgav antitesttel (zöld) festettük meg, a sejtmagokat pedig DAPI festékkel. Skála: 50 µm. Három biológiai replikátum reprezentatív festése (n=3). b) ITGAV festés denzitometriás analízise (n=3). Statisztikai próbaként független mintás t-próbát alkalmaztunk, * p<0.05. c) ITGAV immunfluoreszcens festésének denzitometriás elemzése (n=3). c) A rekombináns Wnt5a kezelt 3D tüdő szöveti aggregátumok centrumában és külső rétegében egyaránt (n=3). Független mintás t-próbát alkalmaztunk statisztikai próbának, * p<
62 5. Megbeszélés A tüdőben zajló, cigarettafüst-indukálta krónikus gyulladásos folyamatok, a PPAR-γ aktivációja, a Wnt szignalizációjának szabálytalan működése és a karcinogenezis kialakulásának megnövekedett rizikója mind összeköttetésben állnak egymással (111) (112). Az általunk végzett kutatások elsőként világítottak rá egy lehetséges molekuláris mechanizmusra, mely az egymással összefüggésben álló változások mögött húzódhat. Eredményeink bizonyították, hogy a cigarettafüst a nem-kanonikus Wnt jelátvitel útvonal molekuláinak az emelkedését képes indukálni, mely párhuzamosan a kanonikus Wnt jelátviteli útvonal szupressziójával jár együtt. Ez a megfigyelés nagy jelentőséggel bír, mert tudjuk, hogy a kanonikus Wnt jelátvitel elengedhetetlen a tüdőben található őssejt populáció fenntartásához (113). Megállapíthatjuk, hogy a dohányfüst olyan szöveti mikrokörnyezet kialakítására képes, mely több szinten is hasonlóságot mutat az öregedésben megfigyelt szöveti változásokhoz. A dohányzókhoz hasonlóan az öregedésben látható mikrokörnyezetben is megfigyelhető az EpCAM + epiteliális sejtekkel dúsított sejtréteg szignifikáns hanyatlása, mely korrelál az emfizéma súlyosságával és a csökkent légzőfelülettel. Ezzel párhuzamosan a gyulladásos környezet is jelen van, melyet az immunsejtek CD45 + leukocita populációjának szaporulata, illetve az aktivált, nem-kanonikus Wnt szignalizáció jellemez a tüdőben (103). Amíg a tüdő fiatal és egészséges, addig a gyulladásos tüdőbetegségek szabályozása és kordában tartása az anti-inflammatorikus M2 makrofág alcsoport dominanciájának és ezzel egyidőben a pro-inflammatorikus M1 makrofág alcsoport szupressziójának dinamikus egyensúlyának fenntartásával történik (114). A PPAR-γ fontos és hasonlóan dinamikus anti-inflammatorikus szerepet tölt be az M2 típusú makrofág polarizáció indukálásával (115). A cigarettafüst elnyújtott expozíciója erősíti az öregedés folyamatát a PPAR-γ expressziójának redukálásával. A cigarettafüst ismételt expozíciója megváltoztatja a tüdő szövetének homeosztázisát és benne a Wnt mikrokörnyezetet is, mely szorosan interferál a PPAR-γ expressziós változásaival (116). Bár a pontos molekuláris szabályozás még nem ismert, az feltételezhető, hogy a PPAR-γ és a Wnt szignalizáció szabályozzák a COPD szisztémás természetét (117) (118) (119). Az öregedésben, karcinogenezisben és az M1 makrofág differenciáción keresztül a proinflammatorikus folyamatok aktivációjában szerepet játszó nem-kanonikus Wnt5a cigarettafüst hatására aktiválódik (103) (38). COPD betegekből izolált apoptotikus testekben/onkoszómákban detektált Wnt5a jelenléte is annak gyulladásban betöltött szerepét bizonyítja (120) (41) (121). Emellett a Wnt5a képes az anti-inflammatorikus PPAR-γ 62
63 szupressziójára a mir27b megnövelt szintjén keresztül, mely a PPAR-γ promoterének kötésével blokkolja annak transzkripcióját (39). Tekintve, hogy mind a Wnt5a, mind a gyulladásos citokinek becsomagolódhatnak az exoszómák kettős lipidrétegébe, a COPD során nem csak a gyulladás válhat szisztémás természetűvé, hanem ezzel egy időben a gyulladás ellen dolgozó PPAR-γ aktivitása is. Korábbi kutatások is feltételezték már, hogy a PPAR-γ agonista terápia modulálja és csökkenti a gyulladást és akár még a cigarettafüst indukálta emfizéma kialakulását is visszafordíthatja (64) (122). További kísérleteink rávilágítottak a PPAR-gammát célzó gyulladáscsökkentő monoterápiák hiányosságára és a terápia sikerét befolyásoló tényezőkre. A PPAR-γ agonista az M2 típusú makrofág differenciációt promotálja, amelyből arra következtethetünk, hogy a PPAR-γ agonista kezelés a karcinogenezis kialakulásában, promotálásában is szerepet játszhat (123). Összefoglalva, a jelen tanulmányban bizonyítottuk, hogy a Wnt5a dohányzás hatására indukálódik és ezzel egy időben a PPAR-γ gátlása is megtörténik, melyek gyulladásos folyamatokat indítanak be. Emellett eredményeink alapján feltételezhetjük, hogy a tüdőben lejátszódó gyulladásos folyamatokban aktivált pro-inflammatorikus citokinek különböző extracelluláris vezikulákban szállítódhatnak el az emberi szervezet bármelyik szervébe, kiváltva ezzel az egész szervezetet érintő szisztémás gyulladást, melynek következtében a krónikus obstruktív tüdőbetegség egy komplex és nehezen kontrollálható betegséggé válik. További kísérletek feltétlenül szükségesek, hogy az általunk tett megfigyeléseket alkalmazni tudjuk a differenciál diagnosztikában, illetve az eredményeink alapján azonosított prognosztikai markereket és extracelluláris vezikulákat hogyan alkalmazhatjuk a célzott terápiában a szisztémás gyulladás csökkentésére. 63

 (125).")
64 Wnt5a Wnt5a és pro-inflammatorikus citokin tartalmú exoszómák felszabadulása Szisztémás gyulladás Pro-inflammatorikus citokinek (IL-6, IL-8) 28. ábra: A dohányzás hatására indukálódó szisztémás gyulladás kialakulásának összefoglaló ábrája a folyamatban szerepet játszó Wnt5a és gyulladásos citokin szintek változásával. A tüdőben dohányzás hatására kialakult krónikus gyulladás esetében a légcserét végző alveolusok felszíne is súlyos mechanikai sérülést szenved, hasonlóan az öregedésben zajló folyamatokhoz, továbbá a tüdő regenerációs képességének csökkenéséhez (124) (125). A dohányzás okozta légzőfelület károsodás mellett nagyon gyakran okoz problémát az aneszteziológiai és intenzív terápiás osztályon fekvő lélegeztetett betegek légzőhámjának súlyos károsodása is (126). Ennek okán a tüdő regenerációjában zajló molekuláris mechanizmusoknak és a légzőfelület differenciálódásának pontosabb megismerése a tüdő gyulladásos folyamatainak terápiás megoldásaiban is kulcsfontosságú lehet. A tüdő alveolusait felépítő ATI és ATII típusú epitéliális sejtek érintettek az ATII-ATI TD-ban, mely folyamat molekuláris szabályozásának megismerése szerepet játszhat terápiásan indukált regenerációs folyamatokban. Munkánk során három Wnt molekula, a Wnt4, Wnt5a és a Wnt7a szerepét feltételeztük az ATII-ATI TD-ban. Eredményeink azt mutatták, hogy mind a három Wnt ligand negatív szabályozóként működik az SFTPC esetében, emellett a Wnt7a az ATI sejtek differenciációs markere, az AQP5 indukátoraként viselkedik. Korábbi eredmények is alátámasztják, hogy a Wnt4 és a Wnt5a a szörfaktáns termelés inhibitoraként (103) míg a Wnt7a a differenciáció és proliferáció negatív szabályozójaként funkcionál adenokarcinóma 64
65 sejtvonalakban (127). Az említett Wnt ligandok mellett az ATII-ATI TD folyamatban más gének is szerepet játszanak, melyek egy komplex hálózaton keresztül szabályozzák egymást. Ennek a bonyolult hálózatnak a feltérképezésére ANN analízist végeztünk. Az analízis során egy közvetlenül Wnt ligand által érintett gént, az ITGAV-ot detektáltuk. Az ITGAV gén a tumornövekedés, metasztázis és szöveti újjáépítés során játszik szerepet (128), emellett aktivációja növeli a sejtadhéziót és gátolja a szörfaktáns termeléshez elengedhetetlen lipid transzportot is (129). Eredményeink azt mutatják, hogy az ITGAV szignifikánsan emelkedik a TD folyamat során, ezzel szemben Wnt5a kezelés hatására csökkenést mutat, így feltételezzük, hogy aktivációja elősegíti az ATI differenciációs folyamatot a szörfaktáns termelés blokkolásán keresztül. További eredményeink az ITGAV fehérjeszint analízisével bizonyítják, hogy a Wnt5a sejtspecifikusan képes az ITGAV epresszióját módosítani. Mezenchimális fibroblasztokban emelkedett ITGAV expressziót mértünk, míg az epiteliális sejtrétegben ezzel szemben az ITGAV csökkénését detektáltuk, melyet mrns szinten is megfigyeltünk az aggregátum kultúrákban rekombináns Wnt5a kezelés hatására. Ezek az eredmények összefüggésbe hozhatóak korábbi eredményekkel, melyekben leírták, hogy a Wnt5a növeli az ITGAV expresszióját a mezenchimában és az SFTPC termelődése összefüggésbe hozható a fibroblaszt differenciációval (129) (103). Összességében feltételezhetjük, hogy nem önmagában a Wnt ligandok, hanem az ITGAV közvetlenül felelős az SFTPC szintjének szabályozásáért. A módosult Wnt mikrokörnyezet által érintett még a THBS1 szekretált glikoprotein, amely részt vesz a sebgyógyulásban, angiogenezisben és a gyulladásos válaszban (130). Eddigi tanulmányok leírták a THBS1-nek a tumornövekedésben, sejtmigrációban és neovaszkularizációban inhibitorként betöltött szerepét is (131) (132). A THBS1 emelkedett expressziója a karcinogenezis során értelmezhető egyfajta molekuláris védelmi mechanizmusként is. Az általunk vizsgált TGM2 funkciója egy olyan enzimatikus katalizáló fehérje kódolása, mellyel az extracelluláris mátrix integritásában játszik szerepet (133). Eredményeink alapján a TGM2 lecsökkent szintje a TD-ban az extracelluláris mátrix elvesztésén keresztül szükséges lehet az ATII differenciáció elősegítéséhez. Érdekes adat, hogy korábban már leírták a TGM2 alacsonyabb expressziójának védő szerepét gyulladásban (134). Ezek alapján a TGM1 expressziójának redukálódása javíthatja az alveoláris regenerációt. Az EMP2 emelkedett expresszióját detektálták már különböző szövetekben, amellyel a sejtmembrán összetételének és az erek növekedésének szabályozásával a tumor progresszióban játszik szerepet (135) (136) (137). Így az EMP2 emelkedett expressziója az ATII-ATI TD során a kapilláris erek 65
66 formációjában lehet fontos. A CYP4B1 is egy jellegzetes markere a tüdő celluláris és bronchiális differenciációjának (138). A CYP4B1 emelkedése az ATII-ATI differenciáció során a Wnt3a ligand által érintett, amely fontos szerepet játszik az epitéliális-mezenchimális tranzícióban (EMT) tüdőrák esetében (139). Továbbá elmondhatjuk, hogy az ATII-ATI TD során detektált Wnt ligandok mellett, más, Wnt jelátvitel asszociált gének expresszióját is vizsgáltuk, melyek fokozhatják az ATI típusú differenciációt és segíthetnek a károsodott légzőfelület gyógyulásában. Összefoglalhatjuk, hogy kísérleteink során rendelkezésünkre álló statisztikai adatokból highthroughput analízisek és adatkiértékelő szoftverek segítségével olyan specifikus molekulákat azonosítottunk, melyek a tüdő légzőhámjának regenerációs folyamatában játszanak szerepet. Ennek eredményeként a légcserét végző alveolusok sejtszintű megújulásában lehetséges terápiás célpontok kerültek előtérbe, megteremtve ezzel potenciális szövetspecifikus terápiás lehetőségeket a jövőben. 66
67 6. Új eredmények, összefoglalás Kiváltja-e a cigarettafüst a nem-kanonikus Wnt5a molekula szintjének emelkedését és a Wnt5aval kapcsolatosan COPD-re jellemző gyulladásos környezet kialakulását? 1. A cigarettafüst indukálja a nem-kanonikus Wnt5a emelkedését a tüdőben, párhuzamosan a kanonikus Wnt jelátviteli útvonal szupressziójával. Emellett megfigyelhető az EpCAM + epiteliális sejtekkel dúsított sejtréteg szignifikáns hanyatlása, mely korrelál az emfizéma súlyosságával és a csökkent légzőfelülettel. 2. Dohányzás hatására a Wnt5a aktiválódásával párhuzamosan a gyulladásos folyamatok is indukálódnak az M1 makrofág differenciáción keresztül, melyet az immunsejtek CD45 + leukocita populációjának szaporulata jellemez, illetve ennek a folyamatnak a részeként az anti-inflammatorikus PPAR-γ expressziója is redukálódik. Kimutatható-e a Wnt5a emelkedett szintje COPD-betegek szérumában, amely magyarázhatja az emberre, de a kísérleti állatokra nem jellemző szisztémás gyulladás kialakulását? 3. Kísérleteink során bizonyítottuk, hogy COPD betegek esetében a Wnt5a és proinflammatorikus citokinek becsomagolódhatnak és különböző extracelluláris vezikulákban szállítódhatnak, melynek következtében a COPD során nem csak a gyulladás válhat szisztémás természetűvé, hanem ezzel egy időben az antiinflammatorikus PPAR-γ aktivitása is. Szabályozza-e, és ha igen, milyen módon a Wnt jelátviteli útvonal a pulmonális regenerációt és az epitéliális transzdifferenciációs folyamatokat? 4. Az ATII-ATI TD folyamataiban eredményeink azt mutatták, hogy a Wnt4, Wnt5a és Wnt7a ligandok negatív szabályozóként működnek az SFTPC esetében, emellett a Wnt7a az AQP5 indukátoraként viselkedik. Összességében feltételezhetjük, hogy nem önmagában a Wnt ligandok, hanem egy közvetlenül Wnt ligand által érintett gén, az ITGAV felelős az SFTPC szintjének szabályozásáért. 67
68 7. Hivatkozások 1. Sopori M. Science and society: Effects of cigarette smoke on the immune system. Nature Reviews Immunology (2002) 2: doi: /nri Qiu F, Liang C-L, Liu H, Zeng Y-Q, Hou S, Huang S, Lai X, Dai Z. Impacts of cigarette smoking on immune responsiveness: Up and down or upside down? Oncotarget (2017) 8: doi: /oncotarget Gonçalves RB, Coletta RD, Silvério KG, Benevides L, Casati MZ, Da Silva JS, Nociti FH. Impact of smoking on inflammation: Overview of molecular mechanisms. Inflammation Research (2011) 60: doi: /s Kalra R, Singh SP, Savage SM, Finch GL, Sopori ML. Effects of Cigarette Smoke on Immune Response: Chronic Exposure to Cigarette Smoke Impairs Antigen-Mediated Signaling in T Cells and Depletes IP3-Sensitive Ca2+ Stores. J Pharmacol Exp Ther (2000) 293: doi: / Talhout R, Schulz T, Florek E, van Benthem J, Wester P, Opperhuizen A. Hazardous compounds in tobacco smoke. International Journal of Environmental Research and Public Health (2011) 8: doi: /ijerph Gualano RC, Hansen MJ, Vlahos R, Jones JE, Park-Jones RA, Deliyannis G, Turner SJ, Duca KA, Anderson GP. Cigarette smoke worsens lung inflammation and impairs resolution of influenza infection in mice. Respiratory Research (2008) 9: doi: / Lee J, Taneja V, Vassallo R. Cigarette smoking and inflammation: Cellular and molecular mechanisms. Journal of Dental Research (2012) 91: doi: / Moldoveanu B, Otmishi P, Jani P, Walker J, Sarmiento X, Guardiola J, Saad M, Jerry Y. Inflammatory mechanisms in the lung. Journal of Inflammation Research (2009) 2:1 11. doi: /jir.s Izquierdo JL, Resano P, El Hachem A, Graziani D, Almonacid C, Sánchez IM. Impact of COPD in patients with lung cancer and advanced disease treated with chemotherapy and/or tyrosine kinase inhibitors. International journal of chronic obstructive pulmonary disease (2014) 9: doi: /copd.s Durham AL, Adcock IM. The relationship between COPD and lung cancer. Lung cancer (Amsterdam, Netherlands) (2015) 90: doi: /j.lungcan
69 11. Yao H, Rahman I. Current concepts on the role of inflammation in COPD and lung cancer. Current Opinion in Pharmacology (2009) 9: doi: /j.coph Adeloye D, Chua S, Lee C, Basquill C, Papana A, Theodoratou E, Nair H, Gasevic D, Sridhar D, Campbell H, et al. Global and regional estimates of COPD prevalence: Systematic review and meta-analysis. Journal of global health (2015) 5: doi: /jogh Anthonisen NR, Connett JE, Murray RP. Smoking and lung function of Lung Health Study participants after 11 years. American Journal of Respiratory and Critical Care Medicine (2002) 166: doi: /rccm Raherison C, Girodet P-O. Epidemiology of COPD. European respiratory review : an official journal of the European Respiratory Society (2009) 18: doi: / Oh JY, Sin DD. Lung inflammation in COPD: why does it matter? F1000 medicine reports (2012) 4:23. doi: /m Warren CPW. The nature and causes of chronic obstructive pulmonary disease: A historical perspective. The Christie Lecture 2007, Chicago, USA. Canadian Respiratory Journal (2009) 16: doi: /2009/ Vestbo J, Hurd SS, Agustı AG, Jones PW, Vogelmeier C, Anzueto A, Barnes PJ, Fabbri LM, Martinez FJ, Nishimura MM, et al. Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease GOLD executive summary. American Journal of Respiratory and Critical Care Medicine (2013) 187: doi: /rccm pp 18. King PT. Inflammation in chronic obstructive pulmonary disease and its role in cardiovascular disease and lung cancer. Clinical and Translational Medicine (2015) 4:26. doi: /s z 19. Balkissoon R. Persistent systemic infl ammation is associated with poor clinical outcomes in COPD: A novel phenotype. COPD: Journal of Chronic Obstructive Pulmonary Disease (2012) 9:439. doi: / Pan Z, Yu H, Liao J Lou. Probing cellular and molecular mechanisms of cigarette smoke-induced immune response in the progression of chronic obstructive pulmonary disease using multiscale network modeling. PLoS ONE (2016) 11: doi: /journal.pone Cazzola M, Matera MG, Rogliani P, Page C. Treating systemic effects of COPD. 69
70 Trends in Pharmacological Sciences (2007) 28: doi: /j.tips Barnes PJ, Ito K, Adcock IM. Corticosteroid resistance in chronic obstructive pulmonary disease: Inactivation of histone deacetylase. Lancet (2004) 363: doi: /s (04)15650-x 23. Mirza S, Clay RD, Koslow MA, Scanlon PD. COPD Guidelines: A Review of the 2018 GOLD Report. Mayo Clinic Proceedings (2018) 93: doi: /j.mayocp Dijksterhuis JP, Petersen J, Schulte G. WNT/Frizzled signalling: receptor-ligand selectivity with focus on FZD-G protein signalling and its physiological relevance: IUPHAR Review 3. British journal of pharmacology (2014) 171: doi: /bph Torres MA, Yang-Snyder JA, Purcell SM, DeMarais AA, McGrew LL, Moon RT. Activities of the Wnt-1 class of secreted signaling factors are antagonized by the Wnt- 5A class and by a dominant negative cadherin in early Xenopus development. Journal of Cell Biology (1996) 133: doi: /jcb Ackers I, Malgor R. Interrelationship of canonical and non-canonical Wnt signalling pathways in chronic metabolic diseases. Diabetes & vascular disease research (2018) 15:3 13. doi: / Pongracz JE, Stockley RA. Wnt signalling in lung development and diseases. Respiratory research (2006) 7:15. doi: / Shao Y, Zheng Q, Wang W, Xin N, Song X, Zhao C. Biological functions of macrophage-derived Wnt5a, and its roles in human diseases. Oncotarget (2016) 7: doi: /oncotarget Sandsmark E, Hansen AF, Selnæs KM, Bertilsson H, Bofin AM, Wright AJ, Viset T, Richardsen E, Drabløs F, Bathen TF, et al. A novel non-canonical Wnt signature for prostate cancer aggressiveness. Oncotarget (2017) 8: doi: /oncotarget Stewart DJ. Wnt Signaling Pathway in Non Small Cell Lung Cancer. Journal of the National Cancer Institute (2013) 300:1 11. doi: /jnci/djt Liang H, Chen Q, Coles AH, Anderson SJ, Pihan G, Bradley A, Gerstein R, Jurecic R, Jones SN. Wnt5a inhibits B cell proliferation and functions as a tumor suppressor in hematopoietic tissue. Cancer Cell (2003) 4: doi: /s (03)00268-x 70
71 32. Bitler BG, Nicodemus JP, Li H, Cai Q, Wu H, Hua X, Li T, Birrer MJ, Godwin AK, Cairns P, et al. Wnt5a suppresses epithelial ovarian cancer by promoting cellular senescence. Cancer Research (2011) 71: doi: / can Kremenevskaja N, Von Wasielewski R, Rao AS, Schöfl C, Andersson T, Brabant G. Wnt-5a has tumor suppressor activity in thyroid carcinoma. Oncogene (2005) 24: doi: /sj.onc Kim J, Kim J, Kim DW, Ha Y, Ihm MH, Kim H, Song K, Lee I. Wnt5a Induces Endothelial Inflammation via -Catenin-Independent Signaling. The Journal of Immunology (2010) 185: doi: /jimmunol Popp T, Steinritz D, Breit A, Deppe J, Egea V, Schmidt A, Gudermann T, Weber C, Ries C. Wnt5a/β-catenin signaling drives calcium-induced differentiation of human primary keratinocytes. Journal of Investigative Dermatology (2014) 134: doi: /jid Weeraratna AT, Jiang Y, Hostetter G, Rosenblatt K, Duray P, Bittner M, Trent JM. Wnt5a signaling directly affects cell motility and invasion of metastatic melanoma. Cancer Cell (2002) 1: doi: /s (02) Bartis D, Csongei V, Weich A, Kiss E, Barko S, Kovacs T, Avdicevic M, D Souza VK, Rapp J, Kvell K, et al. Down-regulation of canonical and up-regulation of noncanonical wnt signalling in the carcinogenic process of squamous cell lung carcinoma. PloS one (2013) 8:e doi: /journal.pone Rapp J, Kiss E, Meggyes M, Szabo-Meleg E, Feller D, Smuk G, Laszlo T, Sarosi V, Molnar TF, Kvell K, et al. Increased Wnt5a in squamous cell lung carcinoma inhibits endothelial cell motility. BMC Cancer (2016) 16: doi: /s Rauner M, Stein N, Winzer M, Goettsch C, Zwerina J, Schett G, Distler JHW, Albers J, Schulze J, Schinke T, et al. WNT5A is induced by inflammatory mediators in bone marrow stromal cells and regulates cytokine and chemokine production. Journal of Bone and Mineral Research (2012) doi: /jbmr Whang YM, Jo U, Sung JS, Ju HJ, Kim HK, Park KH, Lee JW, Koh IS, Kim YH. Wnt5a is associated with cigarette smoke-related lung carcinogenesis via protein kinase C. PloS one (2013) 8:e doi: /journal.pone Zhao Y, Wang C-L, Li R-M, Hui T-Q, Su Y-Y, Yuan Q, Zhou X-D, Ye L. Wnt5a promotes inflammatory responses via nuclear factor κb (NF-κB) and mitogen-activated protein kinase (MAPK) pathways in human dental pulp cells. Journal of Biological 71
72 Chemistry (2014) doi: /jbc.m Naskar D, Maiti G, Chakraborty A, Roy A, Chattopadhyay D, Sen M. Wnt5a Rac1 NF-κB Homeostatic Circuitry Sustains Innate Immune Functions in Macrophages. The Journal of Immunology (2014) 192:4386 LP doi: /jimmunol Wojtan P, Mierzejewski M, Osiñska I, Domagala-Kulawik J. Macrophage polarization in interstitial lung diseases. Central European Journal of Immunology (2016) 41: doi: /ceji Lehtonen A, Ahlfors H, Veckman V, Miettinen M, Lahesmaa R, Julkunen I. Gene expression profiling during differentiation of human monocytes to macrophages or dendritic cells. Journal of Leukocyte Biology (2007) 82: doi: /jlb Blumenthal A, Ehlers S, Lauber J, Buer J, Lange C, Goldmann T, Heine H, Brandt E, Reiling N. The Wingless homolog WNT5A and its receptor Frizzled-5 regulate inflammatory responses of human mononuclear cells induced by microbial stimulation. Blood (2006) 108: doi: /blood Pukrop T, Dehghani F, Chuang HN, Lohaus R, Bayanga K, Heermann S, Regen T, Van Rossum D, Klemm F, Schulz M, et al. Microglia promote colonization of brain tissue by breast cancer cells in a Wnt-dependent way. GLIA (2010) 58: doi: /glia Menck K, Klemm F, Gross JC, Pukrop T, Wenzel D, Binder C. Induction and transport of Wnt 5a during macrophage-induced malignant invasion is mediated by two types of extracellular vesicles. Oncotarget (2013) 4: doi: /oncotarget Daynes RA, Jones DC. Emerging roles of PPARs in inflammation and immunity. Nature Reviews Immunology (2002) 2: doi: /nri Lakshmi SP, Reddy AT, Zhang Y, Sciurba FC, Mallampalli RK, Duncan SR, Reddy RC. Down-regulated peroxisome proliferator-activated receptor γ (PPARγ) in lung epithelial cells promotes a PPARγ agonist-reversible proinflammatory phenotype in chronic obstructive pulmonary disease (COPD). Journal of Biological Chemistry (2014) 289: doi: /jbc.m de sá PM, Richard AJ, Hang H, Stephens JM. Transcriptional regulation of adipogenesis. Comprehensive Physiology (2017) 7: doi: /cphy.c Tomas J, Mulet C, Saffarian A, Cavin J-B, Ducroc R, Regnault B, Kun Tan C, Duszka K, Burcelin R, Wahli W, et al. High-fat diet modifies the PPAR-γ pathway leading to 72
73 disruption of microbial and physiological ecosystem in murine small intestine. Proceedings of the National Academy of Sciences of the United States of America (2016) 113:E5934 E5943. doi: /pnas Mazzei JC, Zhou H, Brayfield BP, Hontecillas R, Bassaganya-Riera J, Schmelz EM. Suppression of intestinal inflammation and inflammation-driven colon cancer in mice by dietary sphingomyelin: Importance of peroxisome proliferator-activated receptor γ expression. Journal of Nutritional Biochemistry (2011) 22: doi: /j.jnutbio Huang THW, Razmovski-Naumovski V, Kota BP, Lin DSH, Roufogalis BD. The pathophysiological function of peroxisome proliferator-activated receptor-γ in lungrelated diseases. Respiratory Research (2005) 6: doi: / Huang W, Glass CK. Nuclear receptors and inflammation control: Molecular mechanisms and pathophysiological relevance. Arteriosclerosis, Thrombosis, and Vascular Biology (2010) 30: doi: /atvbaha Pascual G, Fong AL, Ogawa S, Gamliel A, Li AC, Perissi V, Rose DW, Willson TM, Rosenfeld MG, Glass CK. A SUMOylation-dependent pathway mediates transrepression of inflammatory response genes by PPAR-γ. Nature (2005) 437: doi: /nature Wahli W, Michalik L. PPARs at the crossroads of lipid signaling and inflammation. Trends in Endocrinology and Metabolism (2012) 23: doi: /j.tem Cabrero A, Laguna JC, Vazquez M. Peroxisome proliferator-activated receptors and the control of inflammation. Current Drug Targets: Inflammation & Allergy (2002) 1: doi: / (87)90285-x 58. Polvani S, Tarocchi M, Galli A. PPARγ and Oxidative Stress: Con(beta) Catenating NRF2 and FOXO. PPAR Res (2012) 2012: doi: /2012/ Staels B, Fruchart JC. Therapeutic roles of peroxisome proliferator-activated receptor agonists. Diabetes (2005) 54: doi:54/8/2460 [pii] 60. Chung SS, Kim M, Youn B-S, Lee NS, Park JW, Lee IK, Lee YS, Kim JB, Cho YM, Lee HK, et al. Glutathione peroxidase 3 mediates the antioxidant effect of peroxisome proliferator-activated receptor γ in human skeletal muscle cells. Molecular and cellular biology (2009) 29: doi: /mcb Takada I, Mihara M, Suzawa M, Ohtake F, Kobayashi S, Igarashi M, Youn MY, Takeyama KI, Nakamura T, Mezaki Y, et al. A histone lysine methyltransferase 73
74 activated by non-canonical Wnt signalling suppresses PPAR-γ transactivation. Nature Cell Biology (2007) 9: doi: /ncb Liu J, Wang H, Zuo Y, Farmer SR. Functional interaction between peroxisome proliferator-activated receptor γ and beta-catenin. Molecular and cellular biology (2006) 26: doi: /mcb Lecarpentier Y, Claes V, Vallée A, Hébert J-L. Thermodynamics in cancers: opposing interactions between PPAR γ and the canonical WNT/beta-catenin pathway. Clinical and Translational Medicine (2017) 6:14. doi: /s Yin Y, Hou G, Li E, Wang Q, Kang J. PPAR Γ agonists regulate tobacco smokeinduced toll like receptor 4 expression in alveolar macrophages. Respiratory Research (2014) 15: doi: / Belvisi MG, Hele DJ, Birrell MA. Peroxisome proliferator-activated receptor γ agonists as therapy for chronic airway inflammation. European Journal of Pharmacology (2006) 533: doi: /j.ejphar Shan M, You R, Yuan X, Frazier M V., Porter P, Seryshev A, Hong JS, Song LZ, Zhang Y, Hilsenbeck S, et al. Agonistic induction of PPAR?? reverses cigarette smokeinduced emphysema. Journal of Clinical Investigation (2014) 124: doi: /jci Malur A, McCoy AJ, Arce S, Barna BP, Kavuru MS, Malur AG, Thomassen MJ. Deletion of PPAR γ in alveolar macrophages is associated with a Th-1 pulmonary inflammatory response. Journal of immunology (2009) doi: /jimmunol Bouhlel MA, Derudas B, Rigamonti E, Dièvart R, Brozek J, Haulon S, Zawadzki C, Jude B, Torpier G, Marx N, et al. PPARγ Activation Primes Human Monocytes into Alternative M2 Macrophages with Anti-inflammatory Properties. Cell Metabolism (2007) doi: /j.cmet György B, Szabó TG, Pásztói M, Pál Z, Misják P, Aradi B, László V, Pállinger É, Pap E, Kittel Á, et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cellular and Molecular Life Sciences (2011) 68: doi: /s Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: A comprehensive review. Cancer and Metastasis Reviews (2013) 32: doi: /s McDonald MK, Tian Y, Qureshi RA, Gormley M, Ertel A, Gao R, Aradillas Lopez E, Alexander GM, Sacan A, Fortina P, et al. Functional significance of macrophage- 74
75 derived exosomes in inflammation and pain. Pain (2014) 155: doi: /j.pain Li JJ, Wang B, Kodali MC, Chen C, Kim E, Patters BJ, Lan L, Kumar S, Wang X, Yue J, et al. In vivo evidence for the contribution of peripheral circulating inflammatory exosomes to neuroinflammation. Journal of Neuroinflammation (2018) 15: doi: /s Kulshreshtha A, Ahmad T, Agrawal A, Ghosh B. Proinflammatory role of epithelial cell-derived exosomes in allergic airway inflammation. Journal of Allergy and Clinical Immunology (2013) 131: doi: /j.jaci Admyre C, Telemo E, Almqvist N, Lötvall J, Lahesmaa R, Scheynius A, Gabrielsson S. Exosomes - Nanovesicles with possible roles in allergic inflammation. Allergy: European Journal of Allergy and Clinical Immunology (2008) 63: doi: /j x 75. Serban KA, Rezania S, Petrusca DN, Poirier C, Cao D, Justice MJ, Patel M, Tsvetkova I, Kamocki K, Mikosz A, et al. Structural and functional characterization of endothelial microparticles released by cigarette smoke. Scientific Reports (2016) 6: doi: /srep Raposo G. B lymphocytes secrete antigen-presenting vesicles. Journal of Experimental Medicine (1996) 183: doi: /jem Denzer K, Kleijmeer MJ, Heijnen HF, Stoorvogel W, Geuze HJ. Exosome: from internal vesicle of the multivesicular body to intercellular signaling device. Journal of cell science (2000) 113 Pt 19: doi: /jcb Blanchard N, Lankar D, Faure F, Regnault A, Dumont C, Raposo G, Hivroz C. TCR Activation of Human T Cells Induces the Production of Exosomes Bearing the TCR/CD3/ Complex. The Journal of Immunology (2002) 168: doi: /jimmunol Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, Desaymard C. Accumulation of Major Histocompatibility Complex Class II Molecules in Mast Cell Secretory Granules and Their Release upon Degranulation. Molecular Biology of the Cell (1997) 8: doi: /mbc Van Niel G, Raposo G, Candalh C, Boussac M, Hershberg R, Cerf-Bensussan N, Heyman M. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology (2001) 121: doi: /gast Admyre C, Grunewald J, Thyberg J, Bripenäck S, Tornling G, Eklund A, Scheynius A, 75
76 Gabrielsson S. Exosomes with major histocompatibility complex class II and costimulatory molecules are present in human BAL fluid. European Respiratory Journal (2003) 22: doi: / Caby MP, Lankar D, Vincendeau-Scherrer C, Raposo G, Bonnerot C. Exosomal-like vesicles are present in human blood plasma. International Immunology (2005) 17: doi: /intimm/dxh Pisitkun T, Shen R-F, Knepper MA. Identification and proteomic profiling of exosomes in human urine. Proceedings of the National Academy of Sciences of the United States of America (2004) 101: doi: /pnas Admyre C, Johansson S, Qazi K, Filen J, Lahesmaa R, Norman M, Neve E, Scheynius A, Gabrielsson S. Exosomes with Immune Modulatory Features Are Present in Human Breast Milk. The Journal of Immunology (2007) 179: doi: /jimmunol Tan DBA, Armitage J, Teo TH, Ong NE, Shin H, Moodley YP. Elevated levels of circulating exosome in COPD patients are associated with systemic inflammation. Respiratory Medicine (2017) 132: doi: /j.rmed Kadota T, Fujita Y, Yoshioka Y, Araya J, Kuwano K, Ochiya T. Extracellular vesicles in chronic obstructive pulmonary disease. International Journal of Molecular Sciences (2016) 17: doi: /ijms Chen Y, Li G, Liu Y, Werth VP, Williams KJ, Liu ML. Translocation of Endogenous Danger Signal HMGB1 From Nucleus to Membrane Microvesicles in Macrophages. Journal of Cellular Physiology (2016) 231: doi: /jcp Cordazzo C, Petrini S, Neri T, Lombardi S, Carmazzi Y, Pedrinelli R, Paggiaro P, Celi A. Rapid shedding of proinflammatory microparticles by human mononuclear cells exposed to cigarette smoke is dependent on Ca2+mobilization. Inflammation Research (2014) 63: doi: /s Héliot A, Landkocz Y, Roy Saint-Georges F, Gosset P, Billet S, Shirali P, Courcot D, Martin PJ. Smoker extracellular vesicles influence status of human bronchial epithelial cells. International Journal of Hygiene and Environmental Health (2017) 220: doi: /j.ijheh Yang J, Chen J. Developmental programs of lung epithelial progenitors: A balanced progenitor model. Wiley Interdisciplinary Reviews: Developmental Biology (2014) 3: doi: /wdev Gotts JE, Matthay MA. Endogenous and exogenous cell-based pathways for recovery 76
77 from acute respiratory distress syndrome. Clinics in Chest Medicine (2014) 35: doi: /j.ccm IY A, DH B. Derivation of type 1 epithelium from type 2 cells in the developing rat lung. Laboratory Investigations (1975) 32: Available at: +type+2+cells+in+the+developing+rat+lung 93. Barkauskas CE, Cronce MJ, Rackley CR, Bowie EJ, Keene DR, Stripp BR, Randell SH, Noble PW, Hogan BLM. Type 2 alveolar cells are stem cells in adult lung. Journal of Clinical Investigation (2013) 123: doi: /jci Torday JS, Rehan VK. The evolutionary continuum from lung development to homeostasis and repair. American Journal of Physiology-Lung Cellular and Molecular Physiology (2007) 292:L608 L611. doi: /ajplung Bielefeld KA, Amini-Nik S, Alman BA. Cutaneous wound healing: Recruiting developmental pathways for regeneration. Cellular and Molecular Life Sciences (2013) 70: doi: /s Mucenski ML, Wert SE, Nation JM, Loudy DE, Huelsken J, Birchmeier W, Morrisey EE, Whitsett JA. β-catenin Is Required for Specification of Proximal/Distal Cell Fate during Lung Morphogenesis. Journal of Biological Chemistry (2003) 278: doi: /jbc.m Li C, Xiao J, Hormi K, Borok Z, Minoo P. Wnt5a participates in distal lung morphogenesis. Developmental Biology (2002) 248: doi: /dbio Weidenfeld J, Shu W, Zhang L, Millar SE, Morrisey EE. The WNT7b promoter is regulated by TTF-1, GATA6, and Foxa2 in lung epithelium. Journal of Biological Chemistry (2002) 277: doi: /jbc.m Kadzik RS, Cohen ED, Morley MP, Stewart KM, Lu MM, Morrisey EE. Wnt ligand/frizzled 2 receptor signaling regulates tube shape and branch-point formation in the lung through control of epithelial cell shape. Proceedings of the National Academy of Sciences of the United States of America (2014) 111: doi: /pnas Shiels MS, Pfeiffer RM, Hildesheim A, Engels EA, Kemp TJ, Park JH, Katki HA, Koshiol J, Shelton G, Caporaso NE, et al. Circulating inflammation markers and prospective risk for lung cancer. Journal of the National Cancer Institute (2013) 105: doi: /jnci/djt309 77
78 101. Sekine Y, Hata A, Koh E, Hiroshima K. Lung carcinogenesis from chronic obstructive pulmonary disease: Characteristics of lung cancer from COPD and contribution of signal transducers and lung stem cells in the inflammatory microenvironment. General Thoracic and Cardiovascular Surgery (2014) 62: doi: /s x 102. Kemény Á, Csekő K, Szitter I, Varga Z V, Bencsik P, Kiss K, Halmosi R, Deres L, Erős K, Perkecz A, et al. Integrative characterization of chronic cigarette smokeinduced cardiopulmonary comorbidities in a mouse model. Environmental Pollution (2017) 229: doi: Kovacs T, Csongei V, Feller D, Ernszt D, Smuk G, Sarosi V, Jakab L, Kvell K, Bartis D, Pongracz JE. Alteration in the Wnt microenvironment directly regulates molecular events leading to pulmonary senescence. Aging Cell (2014) 13: doi: /acel Perkins GD, Gao F, Thickett DR. In vivo and in vitro effects of salbutamol on alveolar epithelial repair in acute lung injury. Thorax (2008) 63: doi: /thx Mi H, Muruganujan A, Thomas PD. PANTHER in 2013: modeling the evolution of gene function, and other gene attributes, in the context of phylogenetic trees. Nucleic Acids Research (2012) 41:D377 D386. doi: /nar/gks Thomas PD, Campbell MJ, Kejariwal A, Mi H, Karlak B, Daverman R, Diemer K, Muruganujan A, Narechania A. PANTHER: a library of protein families and subfamilies indexed by function. Genome research (2003) 13: doi: /gr Khan J, Wei JS, Ringner M, Saal LH, Ladanyi M, Westermann F, Berthold F, Schwab M, Antonescu CR, Peterson C, et al. Classification and diagnostic prediction of cancers using gene expression profiling and artificial neural networks.[see comment]. Nature Medicine (2001) 7: doi: / Sarhan AM. Cancer Classification Based on Microarray Gene Expression Data Using DCT and ANN. Journal of Theoretical and Applied Information Technology (2009) Fujiwara N, Kobayashi K. Macrophages in Inflammation. Current Drug Target - Inflammation & Allergy (2005) 4: doi: / Porcheray F, Viaud S, Rimaniol AC, Léone C, Samah B, Dereuddre-Bosquet N, Dormont D, Gras G. Macrophage activation switching: An asset for the resolution of inflammation. Clinical and Experimental Immunology (2005) 142:
79 doi: /j x 111. Sakurai R, Cerny LM, Torday JS, Rehan VK. Mechanism for nicotine-induced upregulation of Wnt signaling in human alveolar interstitial fibroblasts. Experimental Lung Research (2011) 37: doi: / Rapp J, Jaromi L, Kvell K, Miskei G, Pongracz JE. WNT signaling - lung cancer is no exception. Respiratory Research (2017) 18: doi: /s Nabhan A, Brownfield DG, Harbury PB, Krasnow MA, Desai TJ. Single-cell Wnt signaling niches maintain stemness of alveolar type 2 cells. Science (2018)1 12. doi: /science.aam Yuan A, Hsiao YJ, Chen HY, Chen HW, Ho CC, Chen YY, Liu YC, Hong TH, Yu SL, Chen JJW, et al. Opposite Effects of M1 and M2 Macrophage Subtypes on Lung Cancer Progression. Scientific Reports (2015) 5: doi: /srep Lea S, Plumb J, Metcalfe H, Spicer D, Woodman P, Fox JC, Singh D. The effect of peroxisome proliferator-activated receptor-γ ligands on in vitro and in vivo models of COPD. The European respiratory journal (2014) 43: doi: / Ikemura S, Aramaki N, Fujii S, Kirita K, Umemura S, Matsumoto S, Yoh K, Niho S, Ohmatsu H, Kuwata T, et al. Changes in the tumor microenvironment during lymphatic metastasis of lung squamous cell carcinoma. Cancer Science (2016) doi: /cas Hammad H, de Heer HJ, Soullie T, Angeli V, Trottein F, Hoogsteden HC, Lambrecht BN. Activation of peroxisome proliferator-activated receptor-γ in dendritic cells inhibits the development of eosinophilic airway inflammation in a mouse model of asthma. AmJPathol (2004) 118. Solleti SK, Simon DM, Srisuma S, Arikan MC, Bhattacharya S, Rangasamy T, Bijli KM, Rahman A, Crossno JT, Shapiro SD, et al. Airway epithelial cell PPARγ modulates cigarette smoke-induced chemokine expression and emphysema susceptibility in mice. American Journal of Physiology - Lung Cellular and Molecular Physiology (2015) 309:L293 L304. doi: /ajplung Suryawanshi A, Tadagavadi RK, Swafford D, Manicassamy S. Modulation of inflammatory responses by Wnt/??-catenin signaling in dendritic cells: A novel immunotherapy target for autoimmunity and cancer. Frontiers in Immunology (2016) doi: /fimmu Bergenfelz C, Medrek C, Ekstrom E, Jirstrom K, Janols H, Wullt M, Bredberg A, 79
80 Leandersson K. Wnt5a Induces a Tolerogenic Phenotype of Macrophages in Sepsis and Breast Cancer Patients. The Journal of Immunology (2012) 188: doi: /jimmunol Yu CH, Nguyen TTK, Irvine KM, Sweet MJ, Frazer IH, Blumenthal A. Recombinant Wnt3a and Wnt5a elicit macrophage cytokine production and tolerization to microbial stimulation via Toll-like receptor 4. European Journal of Immunology (2014) 44: doi: /eji Youssef J, Badr M. Role of peroxisome proliferator-activated receptors in inflammation control. Journal of biomedicine & biotechnology (2004) 2004: doi: /s Bouhlel MA, Derudas B, Rigamonti E, Dièvart R, Brozek J, Haulon S, Zawadzki C, Jude B, Torpier G, Marx N, et al. PPARγ Activation Primes Human Monocytes into Alternative M2 Macrophages with Anti-inflammatory Properties. Cell Metabolism (2007) 6: doi: /j.cmet Pavone L, Albert S, DiRocco J, Gatto L, Nieman G. Alveolar instability caused by mechanical ventilation initially damages the nondependent normal lung. Critical Care (2007) 11: doi: /cc Sharma G, Goodwin J. Effect of aging on respiratory system physiology and immunology. Clinical interventions in aging (2006) 1: doi: /ciia Alexopoulos C, Jansson B, Lindholm CE. Mucus transport and surface damage after endotracheal intubation and tracheostomy. An experimental study in pigs. Acta anaesthesiologica Scandinavica (1984) 28: Winn RA, Marek L, Han SY, Rodriguez K, Rodriguez N, Haimnond M, Van Scoyk M, Acosta H, Mirus J, Barry N, et al. Restoration of Wnt-7a expression reverses non-small cell lung cancer cellular transformation through Frizzled-9-mediated growth inhibition and promotion of cell differentiation. Journal of Biological Chemistry (2005) 280: doi: /jbc.m Lu JG, Li Y, Li L, Kan X. Overexpression of osteopontin and integrin αv in laryngeal and hypopharyngeal carcinomas associated with differentiation and metastasis. Journal of Cancer Research and Clinical Oncology (2011) 137: doi: /s y 129. Morandi EM, Verstappen R, Zwierzina ME, Geley S, Pierer G, Ploner C. ITGAV and ITGA5 diversely regulate proliferation and adipogenic differentiation of human 80
81 adipose derived stem cells. Scientific Reports (2016) 6: doi: /srep Mikhailenko I, Krylov D, Argraves KMT, Roberts DD, Liau G, Strickland DK. Cellular internalization and degradation of thrombospondin-1 is mediated by the amino-terminal heparin binding domain (HBD). High affinity interaction of dimeric HBD with the low density lipoprotein receptor-related protein. Journal of Biological Chemistry (1997) 272: doi: /jbc Fleitas T, Martínez-Sales V, Vila V, Reganon E, Mesado D, Martín M, Gómez-Codina J, Montalar J, Reynés G. VEGF and TSP1 levels correlate with prognosis in advanced non-small cell lung cancer. Clinical and Translational Oncology (2013) 15: doi: /s Lawler J. Thrombospondin-1 as an endogenous inhibitor of angiogenesis and tumor growth. Journal of Cellular and Molecular Medicine (2002) 6:1 12. doi: /j tb00307.x 133. Ai L, Kim WJ, Demircan B, Dyer LM, Bray KJ, Skehan RR, Massoll NA, Brown KD. The transglutaminase 2 gene (TGM2), a potential molecular marker for chemotherapeutic drug sensitivity, is epigenetically silenced in breast cancer. Carcinogenesis (2008) 29: doi: /carcin/bgm Maiuri L, Luciani A, Giardino I, Raia V, Villella VR, D Apolito M, Pettoello- Mantovani M, Guido S, Ciacci C, Cimmino M, et al. Tissue Transglutaminase Activation Modulates Inflammation in Cystic Fibrosis via PPAR Down-Regulation. The Journal of Immunology (2008) 180: doi: /jimmunol Qin Y, Fu M, Takahashi M, Iwanami A, Kuga D, Rao RG, Sudhakar D, Huang T, Kiyohara M, Torres K, et al. Epithelial membrane protein-2 (EMP2) activates Src protein and is a novel therapeutic target for glioblastoma. Journal of Biological Chemistry (2014) 289: doi: /jbc.m Wadehra M, Iyer R, Goodglick L, Braun J. The tetraspan protein epithelial membrane protein-2 interacts with β1integrins and regulates adhesion. Journal of Biological Chemistry (2002) 277: doi: /jbc.m Gordon LK, Kiyohara M, Fu M, Braun J, Dhawan P, Chan A, Goodglick L, Wadehra M. EMP2 regulates angiogenesis in endometrial cancer cells through induction of VEGF. Oncogene (2013) 32: doi: /onc Hashimoto S, Chen H, Que J, Brockway BL, Drake JA, Snyder JC, Randell SH, Stripp BR. β-catenin SOX2 signaling regulates the fate of developing airway epithelium. 81
82 Journal of Cell Science (2012) 125: doi: /jcs Wang S, Qiu M, Xia W, Xu Y, Mao Q, Wang J, Dong G, Xu L, Yang X, Yin R. Glypican-5 suppresses Epithelial-Mesenchymal Transition of the lung adenocarcinoma by competitively binding to Wnt3a. Oncotarget (2016) 7: doi: /oncotarget
83 8. Köszönetnyilvánítás Elsőként témavezetőimnek Prof. Dr. Pongrácz Juditnak és Prof. Dr. Helyes Zsuzsannának szeretnék köszönetet mondani, akiknek nagyon hálás vagyok, hogy a kezdetektől fogva sok éven át támogattak és kiemelkedő szakmai tudásukkal és tapasztalatukkal hozzásegítettek kutatómunkám elkészüléséhez. Köszönettel tartozom a Gyógyszerészi Biotechnológia Intézet minden volt és jelenlegi dolgozójának, hogy az ott töltött éveim során mindig biztosították számomra a szakmai tudást és hátteret, illetve a jó munka hangulatát. Nagy köszönettel tartozom Dr. Rapp Juditnak szakmai tanácsaiért, melyekkel az évek során ellátott és a kezdetektől rám fordított rendkívül sok időért, amit arra szánt, hogy a kísérletekhez szükséges technikákat maradéktalanul elsajátítsam. Rendkívül hálás vagyok továbbá kollégáimnak és egyben barátaimnak, Dr. Kovács Tamásnak és Ernszt Dávidnak, hogy a nehéz pillanatokban bíztattak és szakmai tanácsaikkal mindig átlendítettek a nehézségeken és a hosszú szortolások alkalmával olyan sok időt szántak kísérleti mintáim elkészítésére. Emellett Steib Anitának és Bánfai Krisztinának tartozom köszönettel az exoszómás kísérletek során nyújtott hasznos szakmai tanácsokért. Továbbá köszönetet szeretnék mondani a PTE ÁOK Farmakológiai és Farmakoterápiai Intézet dolgozóinak, ahol külön szeretném megköszönni Dr. Kun Józsefnek a molekuláris biológiai technikákban nyújtott sok segítségét és Hírné Perkecz Anikónak a festésekben nyújtott segítségét. Nagyon köszönöm Dr. Meggyes Mátyásnak, a PTE ÁOK Orvosi Mikrobiológiai és Immunitástani Intézet munkatársának az áramlási citometriás mérésekben vállalt sok segítségét. Továbbá Dr. Pál Szilárdnak a Gyógyszertechnológiai és Biofarmáciai Intézetből a mesterséges neuronális hálózatok analízisében nyújtott segítségéért tartozunk köszönettel, illetve az Orvosi Biológiai Intézet és Központi Elektronmikroszkópos Laboratóriumnak, hogy kísérleteink során rendelkezésünkre bocsátották az elektronmikroszkóp használatát. Köszönet illeti a PTE Klinikai Központ I.sz. Belgyógyászati Klinika Pulmonológiai Osztályán dolgozó Dr. Sárosi Veronika osztályvezető főorvosnőt és Dr. Ruzsics Istvánt, hogy az osztályon fekvő betegekből származó mintákat biztosították számunkra. Végül, de nem utolsó sorban szeretném hálámat és köszönetemet kifejezni családomnak és páromnak, hogy mindig, minden pillanatban számíthattam támogatásukra. 83
84 ORIGINAL RESEARCH published: 25 July 2018 doi: /fimmu Cigarette Smoke-Induced Pulmonary Inflammation Becomes Systemic by Circulating Extracellular Vesicles Containing Wnt5a and Inflammatory Cytokines Diana Feller 1,2,3, Jozsef Kun 2,3, Istvan Ruzsics 4, Judit Rapp 1,3, Veronika Sarosi 4, Krisztian Kvell 1,3, Zsuzsanna Helyes 2,3 and Judit E. Pongracz 1,3 * 1 Department of Pharmaceutical Biotechnology, School of Pharmacy, University of Pecs, Pecs, Hungary, 2 Department of Pharmacology and Pharmacotherapy, Medical School, University of Pecs, Pecs, Hungary, 3 Szentagothai Research Center, University of Pecs, Pecs, Hungary, 4 Department of Internal Medicine, Clinical Centre and Medical School, University of Pecs, Pecs, Hungary Edited by: John William Christman, The Ohio State University, United States Reviewed by: Maryna Skok, Palladin Institute of Biochemistry (NAS Ukraine), Ukraine Sushil Kumar Mahata, University of California, San Diego, United States *Correspondence: Judit E. Pongracz pongracz.e.judit@pte.hu Specialty section: This article was submitted to Inflammation, a section of the journal Frontiers in Immunology Received: 26 April 2018 Accepted: 12 July 2018 Published: 25 July 2018 Citation: Feller D, Kun J, Ruzsics I, Rapp J, Sarosi V, Kvell K, Helyes Z and Pongracz JE (2018) Cigarette Smoke-Induced Pulmonary Inflammation Becomes Systemic by Circulating Extracellular Vesicles Containing Wnt5a and Inflammatory Cytokines. Front. Immunol. 9:1724. doi: /fimmu Chronic obstructive pulmonary disease (COPD) is a devastating, irreversible pathology affecting millions of people worldwide. Clinical studies show that currently available therapies are insufficient, have no or little effect on elevated comorbidities and on systemic inflammation. To develop alternative therapeutic options, a better understanding of the molecular background of COPD is essential. In the present study, we show that noncanonical and pro-inflammatory Wnt5a is up-regulated by cigarette smoking with parallel up-regulation of pro-inflammatory cytokines in both mouse and human model systems. Wnt5a is not only a pro-inflammatory Wnt ligand but can also inhibit the anti-inflammatory peroxisome proliferator-activated receptor gamma transcription and affect M1/M2 macrophage polarization. Both Wnt5a and pro-inflammatory cytokines can be transported in lipid bilayer sealed extracellular vesicles that reach and deliver their contents to every organ measured in the blood of COPD patients, therefore, demonstrating a potential mechanism for the systemic nature of this crippling disease. Keywords: chronic obstructive pulmonary disease, Wnt5a, extracellular vesicles, peroxisome proliferatoractivated receptor gamma, inflammatory cytokines INTRODUCTION Chronic obstructive pulmonary disease (COPD) is an irreversible condition of airflow limitation characterized by inflammation and impaired respiratory gas exchange (1). The prevalence of COPD is approximately 10% among individuals older than 40 years of age (2). Furthermore, based on recent statistics, COPD is the seventh and fourth highest cause of disability and death worldwide, respectively (3). The main cause of COPD is tobacco smoking, but other factors, such as air pollution, occupational hazards, and infections can also initiate the disease (4, 5). Despite all the therapeutic efforts, COPD is not just irreversible but is also progressive in nature (6). Although COPD is recognized primarily as a lung disease, it has systemic manifestations and the comorbidities affect the heart, pancreas, skeletal muscle, and other organs (7). To suppress systemic inflammation, glucocorticoid therapy is used to treat acute exacerbations of the disease (8 10). Unfortunately, apart from the limited effcacy, unwanted side effects occur including histone deacetylase regulated resistance to corticosteroids (11, 12). Frontiers in Immunology 1 July 2018 Volume 9 Article 1724
85 Feller et al. Systemic Inflammation Induced by Cigarette Smoke To find a permanent cure for COPD, more research is required into both drug target identification and disease pathomechanism. In recent years, peroxisome proliferator-activated receptor (PPAR)- gamma was identified as a central molecule in inflammation (13, 14). PPAR gamma, a member of the ligand-activated nuclear hormone receptor super family, controls gene transcriptions associated with lipid metabolism, adipogenesis, inflammation, and metabolic homeostasis (15, 16), making PPAR gamma an emerging anti-inflammatory and anti-oxidative target gene (13, 17). PPAR gamma agonists exert strong anti-atherogenic, anti-inflammatory (18 21), and anti-oxidant effects by inhibiting several inflammatory mediators (22 25). Regulation of PPAR gamma, however, is essentially Wnt signal ing dependent (26). Mostly, if the canonical or beta-catenin-dependent Wnt pathway is up-regulated, PPAR gamma is down-regulated (27 29). Additionally, PPAR gamma is also a non-canonical Wnt pathway target and is involved in the regulation of blood vessel growth via a non-canonical Wnt5a-dependent mechanism (30). Interestingly, Wnt5a is also a strong pro-inflammatory molecule that has been reported as a potent activator of cigarette smoke-induced inflammatory cytokine expression (30 32) and is characteristically up-regulated in squamous cell carcinomas (30). Cigarette smoke (CS) is also the initial trigger of macrophage activation (33) and macrophage polarization toward the classical M1 and the alternative M2 lineages (34). The anti-inflammatory activity of PPAR gamma is associated with inflammatory disorders of the lung leading to the hypothesis that treatment with PPAR gamma agonists can suppress inflammation (35, 36) and reverse CS-induced emphysema (37). Such theories have been substantiated by studies where PPAR gamma-deficient macrophages contributed toward spontaneous lung inflammation (38) and that PPAR gamma agonists promoted differentiation of the immune suppressive M2 macrophage phenotype (39). It is accepted that macrophage polarization is a multifactorial process and is associated with PPAR gamma in COPD. Based on the literature and our previous studies we hypothesized that although both canonical and non-canonical Wnt signaling regulate PPAR gamma, the non-canonical Wnt signaling pathway turns into dominant upon CS exposure. We also theorized that CS exposure induced local lung inflammation which becomes systemic via delivery of inflammatory messages in extracellular vesicles. To investigate our hypotheses, first, mice were exposed to CS. Then, mrna and protein expressions of the most characteristic, pro-inflammatory Wnt ligand Wnt5a was measured. Next, expression of Wnt signaling, PPAR gamma and pro-inflammatory cytokines were investigated in aggregate cultures of primary human lung cell types and monocyte-derived macrophages. Finally, extracellular vesicles (EV-s) were isolated from sera of control volunteers and COPD patients to measure pro-inflammatory molecules. MATERIALS AND METHODS Materials Cigarette smoke extract (CSE) was diluted in cell culture media to a final concentration of 0.5%. Recombinant human Wnt5a (rhwnt5a) (Chinese Hamster Ovary Cell Line, CHO-derived Gln38-Lys380) protein was obtained from R&D Systems (Minneapolis, MN, USA) and used at a concentration of 1 µg/ml. PPAR gamma agonist, rosiglitazone (RSG), and antagonist GW9662 were purchased from Sigma-Aldrich (St. Louis, USA) and used at 10 µm concentrations each. Collection of Human Serum Samples Human blood samples were collected from the Division of Pulmonology, 1st Department of Internal Medicine, Medical School, University of Pecs. A total of 5 COPD patients and 5 age-matched control samples were collected in blood collection tubes (BD Vacutainer, SST: BD SST Tubes with Silica Clot Activator and Polymer Gel, Franklin Lakes, NJ, USA). Blood samples were clotted for 30 min at room temperature. Samples were then centrifuged at 1,500 rpm for 10 min. Serum samples were stored at 80 C until used for further analysis. Animals CS Exposure of Mouse Lungs C57BL/6 male mice (20 25 g; n = 4 5/group) were housed in the animal facility of the Department of Pharmacology and Pharmacotherapy at the University of Pecs. Water and food was supplied ad libitum. Mice were exposed to CS in a whole-body smoke exposure system using 3R4F research cigarettes (College of Agriculture, University of Kentucky; Lexington, KY, USA). Passive smoking was performed in a manual smoking system (TE2; Teague Enterprises, Woodland, CA, USA). The exposures were performed 2 30 min/day with two cigarettes per occasion. The average concentration of the total suspended particles in the chamber was 70 mg/m 3. Microvesicle/Exosome Homing Study BALB/C mice (20 25 g; n = 3/group) were housed in the animal facility of the Department of Immunology and Biotechnology at the University of Pecs. Water and food was supplied ad libitum. Microvesicles/exosomes were isolated from supernatants of Wnt5a-A549 cell lines and were fluorescently labeled with DiI (see below in detail). Native and fluorescently labeled microvesicles/exosomes were resuspended in 200 µl of phosphate-buffered saline (PBS) and injected into the tail veins of BALB/C mice. Control mice were injected with non-fluorescent exosomes. Mouse Lung Cell Isolation Mice were anesthetized intraperitoneally with ketamine-xylazine (5 mg/kg), sacrificed by cervical dislocation, and their lungs were excised. Lung tissues were divided into three groups and were stored in RNA-later (Qiagen; Hilden, Germany) for molecular biological studies, in 1 ml lysis buffer for Western blot analysis or in 4% buffered formaldehyde (Szkarabeusz Ltd., Pecs, Hungary) for immunohistochemical examinations. For the cell sorting procedure, 3 mice/group were anesthetized. Abdominal aorta was intersected, and mice were perfused with 10 ml of PBS through right ventricle to reduce blood content of the lung. To initiate the fine digestion 3 ml trypsin was applied, 10 ml PBS was used to wash out trypsin thereafter. 3 ml collagenase-dispase 3 mg/ml collagenase (Sigma-Aldrich, St. Louis, USA), 1 mg/ml dispase (Roche F. Hoffmann-LaRoche Ltd., Basel, Switzerland), and 1 u/μl DNase I (Sigma-Aldrich, Frontiers in Immunology 2 July 2018 Volume 9 Article 1724
86 Feller et al. Systemic Inflammation Induced by Cigarette Smoke St. Louis, USA) were used to fill up the lungs through the trachea. Pulmonary lobes were dissected into smaller pieces and digested in 10 ml collagenase-dispase for 50 min with continuous stirring. Digested lung cells were filtered with 70 µm cell-strainer (BD Becton, Dickinson and Company, Franklin Lakes, NJ, USA). Cell Sorting Single cell suspensions of mouse lungs were labeled with anti-mouse CD45-FITC produced at the University of Pecs, Department of Immunology and Biotechnology and anti-mouse EpCAM1 (G8.8 anti-rat-pe) (40). Cell sorting was performed in a FACSAria III (BD Biosciences, San Jose, CA, USA) cell sorter. The following populations were collected: EpCAM CD45, EpCAM + CD45, EpCAM + CD45 +, and EpCAM + CD45. The purity of sorting was above 99%. Preparation of CSE Cigarette smoke extract solution was prepared using a modification of the method described earlier (41). CS of two 3R4F research grade cigarettes (Kentucky Tobacco Research and Development Center at the University of Kentucky, Lexington, KY, USA) were bubbled into 10 ml of RPMI without FBS (Lonza, Basel, Switzerland) using a Harvard peristaltic pump (Harvard Apparatus, P-1500, Holliston, USA) at a 550 ml/min flow rate for approximately 3 min. CSE solution was filtered through a 0.22 µm filter (TPP, Trasadingen, Switzerland). This solution was considered as 100% stock and diluted with culture media to 0.5% for further experiments. The ph of each CSE solution was measured as a mean ph The prepared CSE solution was used within 30 min of preparation. The OD of the inner particles of the CSE was measured spectrophotometrically at the wavelength of 360 nm and compared to the control media. Cell Lines and Primary Cells Human adenocarcinoma A549 cell line was obtained from American Type Culture Collection (Rockville, MD, USA). Transgenic Wnt5a overexpressing-a549 (Wnt5a-A549) cell line was created in our laboratory using lentiviral transgenesis (42). A549 cell lines were cultured in complete Dulbecco s Modified Eagle Medium (DMEM) supplemented with 10% FBS. Primary human small airway epithelial cells (SAEC), normal human lung fibroblast cells (NHLF), and normal human lung microvascular endothelial cells (HMVEC-L) were cultured according to the manufacturers recommendations (Lonza, Basel, Switzerland). Cells were cultured in small airway epithelial cell growth medium (SAGM), fibroblast growth medium, and endothelial growth medium, respectively. All the above cell cultures were incubated at 37 C in humidified atmosphere containing 5% CO 2. Isolation of Primary Human Monocytes Macrophages were obtained from healthy blood donors (Hungarian National Blood Transfusion Service of Pecs, Pecs, Hun gary). Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized whole peripheral blood by standard density Ficoll-Paque (GE Healthcare, Sigma-Aldrich, St. Louis, USA) gradient centrifugation. Positive selection of monocytes was performed using CD14 + MACS colloidal superparamagnetic microbeads (Miltenyi Biotec, GmbH, Bergisch Gladbach, Germany) conjugated with monoclonal anti-human CD14 anti bodies. Cell suspensions were mixed with the beads then CD14 + cells were separated by high gradient magnetic separation using MACS MultiStand, OctoMACS (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). Monocyte purity was regularly >85%. Macrophage differentiation was induced by incubation with 10 nm PMA, for 5 h, at 37 C in humidified atmosphere containing 5% CO 2. Three-Dimensional (3D) Human Lung Aggregates Three-dimensional lung aggregate tissues were generated as described in previous studies (30, 43, 44). Briefly, NHLF, SAEC, and HMVEC-L all purchased from Lonza (Lonza, Basel, Switzerland) were mixed (40% NHLF, 30% SAEC, and 30% HMVEC-L) in the absence or presence of peripheral monocytederived macrophages (30% NHLF, 25% SAEC, 25% HMVEC-L, and 20% macrophages) as described, then the cell suspensions were dispensed onto a low attachment 96-well U-bottom plate (Corning, New York, USA) and were centrifuged at 600 g for 10 min at room temperature. Tissue aggregates were maintained in 2:2:1 ratio of endothelial cell growth medium, SAGM, and fibroblasts growth medium. In the various experiments, aggregates were treated with 0.5% CSE or 1 µg/ml recombinant human Wnt5a (R&D Systems, Minneapolis, USA) for 48 h. RNA Isolation, cdna Synthesis, and Quantitative RT-PCR Total RNA was prepared from 3D cell cultures using NucleoSpin RNAII kit (Macherey-Nagel, Düren, Germany) with on-column DNase digestion. Random hexamer primers of the high capacity RNA to cdna kit (Thermo Fisher Scientific, Waltham, MA, USA) was applied. For cdna synthesis, 1 µg of total RNA was used according to the manufacturers protocol. For gene expression analysis, quantitative RT-PCR was performed using SensiFAST SYBR Green reagent (BioLine, London, UK) on ABI StepOne and StepOnePlus instruments (Thermo Fisher Scientific, Waltham, MA, USA). Data were analyzed using StepOne software and expression changes were normalized to beta-actin housekeeping gene. TaqMan Array Total RNA was isolated from human monocytes then cdna was synthesized. TaqMan master mix was combined with the cdna samples then the mixed solutions were loaded onto the Human WNT Pathway, Fast 96-well TaqMan Array (Thermo Fisher Scientific, Waltham, MA, USA) plate. Gene expression was analyzed using ABI StepOnePlus instrument (Thermo Fisher Scientific, Waltham, MA, USA). Immunohistochemistry and Fluorescent Staining Mice were anesthetized with ketamine-xylazine and their lungs were excised. After a formaldehyde fixation of the lung tissue, paraffn embedded sections were made using a microtome. Frontiers in Immunology 3 July 2018 Volume 9 Article 1724
87 Feller et al. Systemic Inflammation Induced by Cigarette Smoke Before proceeding with the staining protocol, the slides had to be deparaffnized by xylene and rehydrated. Antigen retrieval was performed in citrate buffer (10 mm citric acid, 0.05% Tween 20, ph 6.0). Fluorescent staining was performed using rat antihuman Wnt5a (1:100, Clone , R&D Systems, Minneapolis, USA) monoclonal antibody as primary, then Alexa Fluor 488 donkey anti-rat IgG polyclonal antibody (1:200, Thermo Fisher Scientific, Waltham, MA, USA) as secondary antibodies. The 3D human lung tissue aggregates were carefully removed from the 96-well plate and embedded into Cryomount (Histolab, Finland) embedding medium then immediately frozen to 80 o C. Sections were made using a Leica CM1950 cryostat (Leica, Wetzlar, Germany). After a drying step, sections were fixed in cold acetone (Molar Chemicals Kft., Hungary) for 10 min. Fixed slides were blocked in PBS containing 5% BSA (Sigma Aldrich, St. Louis, USA) for 20 min. Wnt5a protein expression was detected by rat anti-human Wnt5a (1:100, Clone , R&D Systems, Minneapolis, USA) monoclonal antibody as primary antibody. The secondary antibody was Alexa Fluor 488 donkey anti-rat IgG antibody (1:200, Thermo Fisher Scientific, Waltham, USA). Nuclei were counterstained with DAPI (1:1000, Serva, Heidelberg, Germany). Fluorescent images were captured using Olympus IX81 fluorescence microscope (Shinjuku, Tokyo, Japan) equipped with CCD camera and analysis software. Images were processed and analyzed with ImageJ. Flow Cytometric Bead Array (CBA) Cytokine Assay Production of pro-inflammatory cytokines IL-6 and IL-8 were measured in cell culture medium using the multiplexed flow CBA Human Inflammatory Cytokines Kit (BD Biosciences, San Jose, CA, USA). For this assay, 50 µl culture medium/each sample of 3D aggregate cell cultures were collected at 48 h time points. The CBA kits were used according to the manufacturer s instructions. The collected samples were diluted four times in the kit s assay buffer. Diluted samples (50 µl) were added to equal amounts of fluorescent cytokine capture bead suspension (50 µl), and human inflammatory cytokine-phycoerythrin detection reagent (50 µl). Samples were then incubated for 3 h at room temperature. Labeled beads were measured using FACS Canto II flow cytometer (BD Immunocytometry Systems, Erembodegen, Belgium) with BD FACS DIVA software V6 and data were analyzed by FCS Express V3 software. EV Isolation From Human Serum Samples Extracellular vesicles were isolated from human serum samples of 5 COPD patients and 5 healthy controls using the Total Exosome Isolation kit (from serum) (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturer s protocol. Frozen serum samples were thawed at room temperature. Samples were then centrifuged at 300 g for 30 min at 4 o C to remove cells and cell debris. Then the supernatant was centrifuged with 2,000 g for 30 min at 4 o C to collect megasomes/oncosomes in the pellet. The supernatant was then mixed with total exosome isolation reagent and vortexed. Samples were incubated at 2 8 C for 30 min, then centrifuged at 14,000 g for 10 min at room temperature. The pellets containing EV-s (microvesicles/oncosomes) were collected and resuspended in cold EV resuspension buffer and stored at 80 o C until further analysis. EV Isolation From A549 and Wnt5a-A549 Cell Lines Extracellular vesicles were isolated from cell culture media following the protocol of the total exosome isolation (from cell culture media) kit (Invitrogen, Cat. number: ). A549 and Wnt5a-A549 cell lines were cultured in DMEM supplemented with 10% FBS at 37 C in humidified atmosphere containing 5% CO 2 until confluent. The medium was then removed, the cell layer washed in PBS, and FBS-free medium was added to the cultures. After 2 days at 37 C, 5% CO 2, the FBS-free culture medium was harvested and centrifuged at 300 g for 10 min at 4 o C to remove cells and cell debris. Then the supernatant was centrifuged with 2,000 g for 30 min at 4 o C to collect megasomes/oncosomes in the pellet. Finally, total exosome isolation (for cell culture media) reagent was added in 0.5 volumes. Samples were incubated at 4 C overnight, and then centrifuged at 10,000 g for 1 h at 4 C. The pellets containing EV-s were collected after centrifugation and resuspended in cold PBS until further analysis. Isolated EV-s were stored at 80 C. Staining of EV-s (Microvesicles or Exosomes) Isolated microvesicles (exosomes) were centrifuged for 1 h at 4 C. Pellets were resuspended in PBS containing DiI (Molecular Probes, Thermo Fisher Scientific, Waltham, MA, USA) (dilution 1:10,000). Samples were incubated for 30 min at 37 C, then centrifuged with 10,000 g for 1 h at 4 C. Supernatant was then removed and the pellet was resuspended in PBS. Fluorescence Analysis of Microvesicle (Exosome) Localization Balb/c mice were injected with equal amount of stained or unstained exosome suspension in 200 µl of PBS. Mice were sacrificed after 20 h of exosome injection. Lung, liver, thymus, and the spleen were removed for fluorescence imaging of extracellular vesicle accumulations in mouse organs by the IVIS Lumina III In Vivo Imaging System using the Living Image Software (IVIS Imaging Systems) (PerkinElmer, Waltham, MA, USA). Fluorescence intensity was compared to fluorescence intensity of unstained control organs. Electron Microscopic Detection of Microvesicles (Exosomes) Microvesicles isolated from cell lines were visualized by electron microscopy. First, microvesicles were resuspended in PBS and then fixed in 2.5% glutaraldehyde aqueous solution. 5 µl of fixed exosomes were embedded in the EM grid, which was flushed by water and blown dry for 24 h. Samples were treated with 5% uranyl-acetate at ph 7 for 5 min. The grid was examined using Morgagni 268D transmission electron microscope at 80 kv. Images were acquired using an integrated MegaView III digital camera (Olympus Soft Imaging Solutions GmbH; Munster, Germany). Frontiers in Immunology 4 July 2018 Volume 9 Article 1724
88 Feller et al. Systemic Inflammation Induced by Cigarette Smoke Sandwich Enzyme-Linked Immunosorbent Assay (ELISA) Mega EV (oncosome) and micro EV (exosome) samples were resuspended in PBS containing 0.5% Tween µl of each sample was loaded to wells in the Aviva Systems Biology WNT5A ELISA Kit (Human) (OKEH00723). The plate was pre-coated with a Wnt5a-specific antibody (96-well plate, 12 8 well strips) and blocked. Standards were also provided by the manufacturer. Samples were incubated for 2 h at 37 o C, then the wells were washed and incubated further with a biotinylated Wnt5a-specific detector antibody for further 1 h at 37 o C. After a washing step, incubation continued with Avidin-HRP conjugate for 1 h at 37 C. The wells were then washed and TMB (3,3,5,5 -Tetramethylbenzidine) substrate was added for 15 min at 37 C in the dark. The reaction was then stopped and the color reaction was measured at 450 nm absorbance. Results were calculated against the standard curve. Statistical Analysis Statistical analysis was performed with GraphPad version 6 software. Data are presented as mean ± SEM, and statistical analysis was performed using the independent samples t-test and one-way ANOVA with Bonferroni correction. p < 0.05 was considered as significant. RESULTS CS and CSE Trigger Wnt5a Up-Regulation in Both Mouse and Human Lung Tissues To investigate the effects of CS as a systemic inducer of inflammation, in vivo and in vitro studies were performed. First, C57BL/6 mice were exposed to CS for 2 30 min each day over 2 months. The test animals developed emphysema-like tissue damage (Figure 1A) that correlated with decreased epithelial cell number (Figure 1B). Epithelial cells were also separated from the CD45 + leukocyte population (Figure 1B). Both the epithelial cell enriched and the CD45 + cell populations had elevated Wnt5a mrna levels (Figure 1C). Wnt5a protein expression levels were also increased (Figures 1D,E). Different isoforms of CD45 are expressed in many leukocyte populations including T and B lymphocytes and antigen-presenting cells, no effect of these immune cells, except macrophages, were investigated further. To test whether CSE can trigger Wnt5a up-regulation in human lung tissues, in vitro human lung aggregate cultures were used. The aggregate cultures contained SAEC, mesenchyme (NHLF), and endothelial cells (HMVEC-L) in the presence or absence of human blood monocyte-derived macrophages and were treated with CSE for a maximum of 48 h. The CSE treated aggregates have shown increased levels of Wnt5a both at mrna and protein levels but only if monocytes were present in the aggregate tissues (Figures 2A D) (45, 46). Increased Inflammatory Cytokine Production in the Lung After CS Exposure Is Associated With the Presence of Macrophages After the recruitment of immune cells to help in eliminating foreign materials and to aid tissue repair after damage or infection, immune reactions are resolved and the tissue homeostasis is normalized. Macrophages are involved in self-limiting, regulated resolution of inflammatory processes at different levels. Tissue-resident macrophages and activated monocytes can adopt different activation status, and although the process is still not fully characterized, M1 and M2 type polarization can provide an outline of how macrophage activity can change during inflammation. To test the effects of CS on the human lung, aggregate cultures of primary human lung cell types were exposed to CSE for 48 h and pro-inflammatory cytokine gene expressions were measured. Both in the presence or absence of macrophages, significant increase of IL6 peptide levels were detected compared to untreated controls (Figure 3A). If the human lung aggregate cultures were exposed to rhwnt5a (1 µg/ml, for 48 h) then both IL-6 and IL-8 peptide levels increased significantly in the monocyte containing tissue cultures indicating a sequential activation of pro-inflammatory cytokine and pro-inflammatory Wnt5a production (Figure 3B). To attempt to dissect such a dynamic process, blood monocytes were exposed to CSE, then, Wnt signaling TaqMan Array was performed (Figures 3C,D). Compared to untreated controls, up-regulation of characteristic non-canonical Wnt signaling molecules were detected, including Wnt5a, Wnt5b, Wnt11 as ligands and an inhibitor of the canonical Wnt pathway, Dickkopf 2 (DKK-2) (Figures 3C,D). In addition to specific molecules of the Wnt pathway, increased transcription of CXXC4 a member of the zinc finger domain-containing protein family was detected. Up-regulation of CXXC4 that functions as an antagonist of the canonical Wnt/integrated signaling pathway indicates active suppression of the canonical Wnt pathway. Furthermore, increased mrna levels of CSNKD1 a casein kinase I gene family member, transducin-like enhancer of split 4 (TLE4), and Wnt10a were also detected. While CSNKD1 is an essential serine/threonine-protein kinase and regulator of diverse cellular growth and survival; TLE4 is a transcriptional corepressor. Overexpression of the Wnt10a gene may be of particular interest as Wnt10a can play key roles in carcinogenesis through activation of the Wnt-beta-catenin-TCF signaling pathway. Simultaneously, reduced transcription of Axin2 that plays an important role in the regulation of beta-catenin stability and a characteristic canonical Wnt ligand, Wnt1, were down-regulated. Reduced transcription of TLE2 and secreted frizzled-related protein 5 was also observed, demonstrating deregulation of Wnt signaling in macrophages as a direct effect of CS exposure. Deregulation of either the canonical or the non-canonical Wnt signaling results in modification of PPAR gamma expression and activity (31). To investigate how such changes in the Wnt signaling pathway can affect the dynamic polarization of macrophages, pro-inflammatory cytokines characteristic to the pro-inflammatory M1, and the anti-inflammatory but pro-tumorigenic M2 phenotypes (47) were measured in association with PPAR gamma expression and activity. To study how PPAR gamma is involved in the CS induced Wnt5a up-regulation as well as consequent pro-inflammatory cytokine production, human lung aggregate cultures were exposed to Wnt5a and CSE in the presence or absence of PPAR gamma agonist, rosiglitazone (RSG), and inhibitor (GW9662) (Figures 4A,B). The M1 phenotype is mostly characterized by pro-inflammatory cytokines IL-6, IL-8, IL-23, and IL-1β, while M2 by anti-inflammatory cytokines IL-10 and TGF-beta (34). In the studied time frame, inhibition Frontiers in Immunology 5 July 2018 Volume 9 Article 1724

89 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 1 Continued Frontiers in Immunology 6 July 2018 Volume 9 Article 1724
 Hematoxylin-eosin staining of paraffin embedded sections of healthy control and CS exposed lungs. Mice were exposed to CS for 1 h/day for 2 months. (Scale bars, 100 µm.")
90 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 1 Effects of cigarette smoke (CS) on C57BL/6 mouse lungs. (A) Hematoxylin-eosin staining of paraffin embedded sections of healthy control and CS exposed lungs. Mice were exposed to CS for 1 h/day for 2 months. (Scale bars, 100 µm.) (B) Changes in EpCAM + and CD45 + cell populations in control and CS exposed lungs determined by flow cytometry. (C) Wnt5a mrna expression changes in sorted EpCAM + and CD45 + cells following 1 and 2 months of CS exposure (n = 3). (D) Western blot analysis of Wnt5a protein expression in control and CS exposed lung tissue extracts. (E) Detection of Wnt5a protein expression in control and CS exposed lung tissue sections [EpCAM + (green)/wnt5a (red)/nucleus (blue), Scale bars, 200 µm]. FIGURE 2 Effects of cigarette smoke extracts (CSE) on primary human 3D lung aggregate cultures. (A) Wnt5a mrna was measured by qrt-pcr in control and CSE treated (48 h) aggregate cultures containing and not containing macrophages (M). (B) Fluorescence intensity of Wnt5a protein was analyzed by ImageJ. (C) Immunofluorescence staining of Wnt5a (green) and Cytokeratin7 (red) proteins in frozen sections of control aggregates and CSE (48 h) exposed aggregates not containing M. (D) Immunofluorescence staining of Wnt5a (green) and Cytokeratin7 (red) proteins in frozen sections of control aggregates and CSE (48 h) exposed aggregates containing M. Nuclei were stained by TO-PRO-3 Iodide (1:1,000, pseudo-blue). [Scale bars: 100 µm (magnification, 20 ), 20 µm (magnification, 63 )]. of the anti-inflammatory PPAR gamma by GW9662 reduced the gene expression of IL-10 the characteristic cytokine marker of the anti-inflammatory M2 phenotype (Figure 4B). Cytokine production, however, is a dynamic process. In order to investigate it, human blood monocyte-derived macrophages were exposed to CSE for two different time points 3 and 48 h, then PPAR gamma transcription was measured. Interestingly, at the 3 h time point the anti-inflammatory PPAR gamma transcription significantly increased, but by 48 h the transcript level reduced to control levels (Figure 4C) indicating that expression of the anti-inflammatory PPAR gamma would not keep up with the prolonged inflammatory stimuli. Frontiers in Immunology 7 July 2018 Volume 9 Article 1724
 after cigarette smoke")
91 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 3 Inflammatory cytokine and Wnt mrna levels in primary human 3D lung aggregate cultures and human macrophages (M) after cigarette smoke extracts (CSE) exposure. (A) IL-6 and IL-8 inflammatory cytokine mrna levels in control and CSE exposed (48 h) lung aggregate cultures, containing and not containing M. (B) IL-6 and IL-8 inflammatory cytokine mrna levels in control and rwnt5a (1 µg/ml) treated lung aggregate cultures, containing and not containing M. (C) Taqman array heat map analysis of pooled (n = 4) cdna of human M compared with control. M were treated with CSE for 3 h. (D) Changes in mrna levels of Wnt signaling pathway genes measured by Taqman Array analysis using pooled (n = 4) cdna samples of human M compared with control. Chronic CS Exposure Increases Wnt5a Protein Levels in EV-s of COPD Patients Sera The lipo-glycoprotein Wnt ligands are highly hydrophobic (48), therefore, their transport in aqueous body fluids is facilitated by EV-s (49). EV-s (both megasomes/oncosomes or microsomes/ exosomes) are now widely accepted as a novel route of cellular communication that can influence the long-range Wnt signal gradients (50, 51). Based on the above, we hypothesized that Wnt5a and low serum levels of pro-inflammatory cytokines can be measured in EV-s isolated from human sera (52, 53). Frontiers in Immunology 8 July 2018 Volume 9 Article 1724
92 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 4 Changes of peroxisome proliferator-activated receptor (PPAR) gamma mrna levels in M1 and M2 M type differentiation in primary human 3D lung aggregate cultures and M monocultures. (A) qrt-pcr analysis of M1 (differentiation marker IL-23) and M2 (differentiation marker IL-10) after human rwnt5a (1 µg/ml) treatment for 48 h. (B) Effects of cigarette smoke extracts (CSE), PPAR gamma agonist (RSG), and antagonist (GW9662) treatment on expression of M1 (IL-23) and M2 (IL-10) differentiation markers. CSE treatment (48 h) and also CSE + RSG (10 µm) and CSE + GW9662 (10 µm) treatment on human M. (C) PPAR gamma mrna levels in CSE treated human macrophage cultures after 3 and 48 h of CSE treatment compared with controls. If EV-s can travel from the organ of origin and reach any cells and organs of the human body safely delivering their contents, such transport system would explain the systemic nature of COPD. To test our theory, the larger megavesicles/oncosomes and the smaller microvesicles/exosomes were isolated from sera of COPD patients as well as age-matched healthy controls. The non-small cell carcinoma cell line A549 and its engineered Wnt5a overexpressing sister cell line Wnt5a-A549 were used as positive controls. Wnt5a was measured by ELISA (Figure 5A), whereas pro-inflammatory cytokines were measured by a bead assay using flow cytometry (Figures 5B,C). While Wnt5a was not detectable in sera of neither COPD patients nor age-matched healthy volunteers, megavesicles/oncosomes isolated from sera of COPD patients contained similar levels of Wnt5a as measured in the Wnt5a overexpressing Wnt5a-A549 cell line (Figure 5A). Similarly, while many pro-inflammatory cytokines remained undetectable in pure human sera (data not shown), exosomes contained high levels of such cytokines (Figure 5C) with an extremely high level detected for IL-8 (Figure 5C). To investigate whether these concentrated cellular messages can reach other organs, extracellular vesicles were isolated from the supernatant of the Wnt5a-A549 cell line and the lipid bilayer was stained with a DiI fluorescent dye to be injected into the tail veins of Balb/c mice. Mice were sacrificed after 20 h and the organs were removed to detect fluorescence. The experiments provided evidence that inflammatory cytokine containing microvesicles/ exosomes can reach and become concentrated in the liver, lungs, spleen, and thymus of test animals (Figures 5D F). DISCUSSION Cigarette smoke induced chronic lung inflammation, PPAR gamma activity, Wnt signaling deregulation and increased overall risk of carcinogenesis are all interconnected (54, 55). Our report Frontiers in Immunology 9 July 2018 Volume 9 Article 1724

93 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 5 Continued Frontiers in Immunology 10 July 2018 Volume 9 Article 1724
 Wnt5a protein levels were measured in EV-s in sera of healthy donors and chronic obstructive pulmonary disease (COPD) patients as well as EV-s of A549 and Wnt5a-A549 cell lines as controls using")
94 Feller et al. Systemic Inflammation Induced by Cigarette Smoke FIGURE 5 Wnt5a and proinflammatory cytokine levels in extracellular vesicles (EV-s). (A) Wnt5a protein levels were measured in EV-s in sera of healthy donors and chronic obstructive pulmonary disease (COPD) patients as well as EV-s of A549 and Wnt5a-A549 cell lines as controls using a Wnt5a-specific enzyme-linked immunosorbent assay kit. (B) Inflammatory cytokine levels were measured in pooled megavesicles/oncosome samples of five healthy and five COPD patients isolated from human sera measured by cytometric bead array Flex kit using flow cytometry. (C) Inflammatory cytokine proteins were detected in pooled microvesicle/exosome samples of five healthy and five COPD patients isolated from human sera. (D) Fluorescent staining (Dil) labeled Wnt5a-containing EV-s are detectable in various organs (representative image of n = 3). (E) Fluorescence intensity of Wnt5a containing EV-s in various organs (representative data of n = 3). (F) Electron microscopic image of purified EV-s (microvesicle/exosome population) purified from the Wnt5a-A549 cell line. Scale bar is 100 nm. FIGURE 6 Summary figure of cigarette smoke induced local inflammation becoming systemic by the contribution of Wnt5a and inflammatory cytokines. is the first to shed light on a potential mechanism. We have shown that CS induces the expression of molecules of the non-canonical Wnt signaling pathway that simultaneously suppress the canonical Wnt pathway. This observation is significant as the canonical Wnt signaling is essential to maintain stem cell niches in the lung (56). In fact, CS creates a tissue microenvironment that is similar to the ones observed during aging (43). Similarly to smokers, the aging microenvironment is associated with a significant decrease of the EpCAM + epithelial cell layer that correlates with emphysema and decreased gas exchange surface (43). In parallel, inflammatory environment is present marked by the increased number of the CD45 + leukocyte population of immune cells in the tissue of the lung and up-regulated non-canonical Wnt signaling (43). While the lung is young and healthy, inflammatory respiratory diseases are kept in check by dynamic polarization of macrophages into the anti-inflammatory M2 subsets that suppress the proinflammatory M1 macrophage phenotype (57). PPAR gamma plays an important and similarly dynamic anti-inflammatory role by inducing M2 type polarization (39, 58). Prolonged exposure to CS, however, re-enforces the aging process by reducing PPAR gamma expression. Repeated exposure to CS also alters the lung tissue homeostasis and the Wnt microenvironment that closely interacts with PPAR gamma (59). Although the precise molecular regulation remains largely unclear, PPAR gamma (60, 61) and the Wnt signaling pathways (62) have been suspected to somehow regulate the systemic nature of COPD. The non-canonical Wnt5a that is associated with aging (43), carcinogenesis (30), as well as with activation of pro-inflammatory processes during M1 phenotype macrophage differentiation and pro-inflammatory cytokine production (63 65) was up-regulated upon CS exposure and was found in megavesicles/oncosomes of COPD patients. Wnt5a can also suppress levels of the anti-inflammatory PPAR gamma via increasing mir27b that binds to the PPAR gamma promoter and blocks its transcription (31). As both pro-inflammatory cytokines and Wnt5a can be wrapped and protected by the lipid bilayer of EV-s then not just the inflammation can become systemic but also the inhibition of the anti-inflammatory activity of PPAR gamma (Figure 6). Although previous studies hypothesized that PPAR gamma agonist treatments can modulate and suppress inflammation (35, 66) or even reverse CS-induced emphysema (37), our experiments have highlighted the diffculty of achieving such anti-inflammatory therapy targeting solely PPAR gamma. Furthermore, as PPAR gamma agonists can promote M2 phenotype differentiation of macrophages (39), such PPAR gamma agonist therapy can even lead to unintended promotion of carcinogenesis. In summary, in the present study we demonstrated that during cigarette smoking the inflammation inducer and PPAR Frontiers in Immunology 11 July 2018 Volume 9 Article 1724
Doktori (PhD) értekezés tézisei FELLER DIÁNA. Elméleti Orvostudományok Doktori Iskola Az Immunológia Alapjai Program
 értekezés tézisei FELLER DIÁNA. Elméleti Orvostudományok Doktori Iskola Az Immunológia Alapjai Program") CIGARETTAFÜST-INDUKÁLTA GYULLADÁSOS FOLYAMATOK COPD-BEN SZISZTÉMÁS GYULLADÁS KIALAKULÁSA A WNT5A ÉS A GYULLADÁSOS CITOKIN TARTALMÚ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL Doktori (PhD) értekezés tézisei FELLER
CIGARETTAFÜST-INDUKÁLTA GYULLADÁSOS FOLYAMATOK COPD-BEN SZISZTÉMÁS GYULLADÁS KIALAKULÁSA A WNT5A ÉS A GYULLADÁSOS CITOKIN TARTALMÚ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL Doktori (PhD) értekezés tézisei FELLER
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Immunológia I. 4. előadás. Kacskovics Imre
 Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Intézeti Beszámoló. Dr. Kovács Árpád Ferenc
 Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
Gyógyításra váró pulmonológiai betegségek
 Gyógyításra váró pulmonológiai betegségek Herjavecz Irén Országos Korányi Tbc és Pulmonológiai Intézet 2009.április 20. Népbetegségek a pulmonológiában Epidemiológiai és morbiditási súlyukat tekintve a
Gyógyításra váró pulmonológiai betegségek Herjavecz Irén Országos Korányi Tbc és Pulmonológiai Intézet 2009.április 20. Népbetegségek a pulmonológiában Epidemiológiai és morbiditási súlyukat tekintve a
Preeclampsia-asszociált extracelluláris vezikulák
 Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE
 Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
INTRACELLULÁRIS PATOGÉNEK
 INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN
 2016. 10. 14. KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN Kovács Árpád Ferenc 1, Pap Erna 1, Fekete Nóra 1, Rigó János 2, Buzás Edit 1,
2016. 10. 14. KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN Kovács Árpád Ferenc 1, Pap Erna 1, Fekete Nóra 1, Rigó János 2, Buzás Edit 1,
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Szőlőmag extraktum hatása makrofág immunsejtek által indukált gyulladásos folyamatokra Radnai Balázs, Antus Csenge, Sümegi Balázs
 Szőlőmag extraktum hatása makrofág immunsejtek által indukált gyulladásos folyamatokra Radnai Balázs, Antus Csenge, Sümegi Balázs Pécsi Tudományegytem Általános Orvostudományi Kar Biokémiai és Orvosi Kémiai
Szőlőmag extraktum hatása makrofág immunsejtek által indukált gyulladásos folyamatokra Radnai Balázs, Antus Csenge, Sümegi Balázs Pécsi Tudományegytem Általános Orvostudományi Kar Biokémiai és Orvosi Kémiai
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata
 sejtek mikro RNS (mir) profiljának vizsgálata") Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Áramlási citometria, sejtszeparációs technikák. Immunológiai és Biotechnológiai Intézet ÁOK, PTE
 Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Retinoid X Receptor, egy A-vitamin szenzor a tüdőmetasztázis kontrolljában. Kiss Máté!
 Retinoid X Receptor, egy A-vitamin szenzor a tüdőmetasztázis kontrolljában Kiss Máté! Magreceptor Kutató Laboratórium Biokémiai és Molekuláris Biológiai Intézet Debreceni Egyetem, Általános Orvostudományi
Retinoid X Receptor, egy A-vitamin szenzor a tüdőmetasztázis kontrolljában Kiss Máté! Magreceptor Kutató Laboratórium Biokémiai és Molekuláris Biológiai Intézet Debreceni Egyetem, Általános Orvostudományi
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogénekimmunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Immunológia I. 2. előadás. Kacskovics Imre (imre.kacskovics@ttk.elte.hu)
") Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
3. Főbb Jelutak. 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3.
 Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Doktori értekezés tézisei
 Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Áramlási citometria 2011. / 4. Immunológiai és Biotechnológiai Intézet PTE KK
 Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Allergia immunológiája 2012.
 Allergia immunológiája 2012. AZ IMMUNVÁLASZ SZEREPLŐI BIOLÓGIAI MEGKÖZELÍTÉS Az immunrendszer A fő ellenfelek /ellenségek/ Limfociták, makrofágok antitestek, stb külső és belső élősködők (fertőzés, daganat)
Allergia immunológiája 2012. AZ IMMUNVÁLASZ SZEREPLŐI BIOLÓGIAI MEGKÖZELÍTÉS Az immunrendszer A fő ellenfelek /ellenségek/ Limfociták, makrofágok antitestek, stb külső és belső élősködők (fertőzés, daganat)
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Az immunológia alapjai
 Az immunológia alapjai 8. előadás A gyulladásos reakció kialakulása: lokális és szisztémás gyulladás, leukocita migráció Berki Timea Lokális akut gyulladás kialakulása A veleszületeh és szerzeh immunitás
Az immunológia alapjai 8. előadás A gyulladásos reakció kialakulása: lokális és szisztémás gyulladás, leukocita migráció Berki Timea Lokális akut gyulladás kialakulása A veleszületeh és szerzeh immunitás
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogének immunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogének immunmoduláns hatásai Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun betegségek,
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogének immunmoduláns hatásai Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun betegségek,
A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban között. Dr. Hortobágyi Judit
 A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban 2007-2011 között Dr. Hortobágyi Judit Pikkelysömörre gyakorolt hatása 2007-től 2009-ig 1. Lokális hatása 2. Szisztémás hatása 3. Állatkísérlet
A harkányi gyógyvízzel végzett vizsgálataink eredményei psoriasisban 2007-2011 között Dr. Hortobágyi Judit Pikkelysömörre gyakorolt hatása 2007-től 2009-ig 1. Lokális hatása 2. Szisztémás hatása 3. Állatkísérlet
A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN. Doktori (Ph.D.) értekezés tézisei
 értekezés tézisei") A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN Doktori (Ph.D.) értekezés tézisei RAPP JUDIT Pécsi Tudományegyetem Gyógyszerésztudományi Kar Gyógyszerészi Biotechnológia Tanszék
A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN Doktori (Ph.D.) értekezés tézisei RAPP JUDIT Pécsi Tudományegyetem Gyógyszerésztudományi Kar Gyógyszerészi Biotechnológia Tanszék
Spondylitis ankylopoeticahoz társuló osteoporosis
 Spondylitis ankylopoeticahoz társuló osteoporosis Szántó Sándor DE OEC, Reumatológiai Tanszék 2013.11.05. Szeminárium Csontfelszivódás és csontképzés SPA-ban egészséges előrehaladott SPA Spondylitis ankylopoetica
Spondylitis ankylopoeticahoz társuló osteoporosis Szántó Sándor DE OEC, Reumatológiai Tanszék 2013.11.05. Szeminárium Csontfelszivódás és csontképzés SPA-ban egészséges előrehaladott SPA Spondylitis ankylopoetica
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Kutatási beszámoló ( )
") Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
http://www.rimm.dote.hu Tumor immunológia
 http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
A DOHÁNYZÁS ÉS C.O.P.D.
 A DOHÁNYZÁS ÉS C.O.P.D. Dr. Kardos Kálmán (főorvos, az orvostudományok kandidátusa) 2015 (ÁNTSZ-OTH-MFF) EPIDEMIOLÓGIA ( dohányzás ) A Föld felnőtt lakosságának 1/3 a dohányzik rendszeresen Korunkban évente
A DOHÁNYZÁS ÉS C.O.P.D. Dr. Kardos Kálmán (főorvos, az orvostudományok kandidátusa) 2015 (ÁNTSZ-OTH-MFF) EPIDEMIOLÓGIA ( dohányzás ) A Föld felnőtt lakosságának 1/3 a dohányzik rendszeresen Korunkban évente
1. Bevezetés. Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban
 Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban Doktori tézisek Dr. Jakus Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori
Integrinek, Fc-receptorok és G-fehérje-kapcsolt receptorok jelátvitelének mechanizmusa neutrofil granulocitákban Doktori tézisek Dr. Jakus Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL
 Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 7. előadás Immunizálás. Poliklonális és monoklonális ellenanyag előállítása, tisztítása, alkalmazása Az antigén (haptén + hordozó) sokféle specificitású ellenanyag
Immunológiai módszerek a klinikai kutatásban 7. előadás Immunizálás. Poliklonális és monoklonális ellenanyag előállítása, tisztítása, alkalmazása Az antigén (haptén + hordozó) sokféle specificitású ellenanyag
B-sejtek szerepe az RA patológiás folyamataiban
 B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológia 4. A BCR diverzitás kialakulása
 Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Sejtkultúra előállítása, steril munka feltételei Immunsejtek izolálása és funkcionális vizsgálata április 15. Kremlitzka Mariann, PhD
 Sejtkultúra előállítása, steril munka feltételei Immunsejtek izolálása és funkcionális vizsgálata 2019. április 15. Kremlitzka Mariann, PhD Miért tartunk fenn sejtkultúrákat? Egy sejt vizsgálat Kontrollált
Sejtkultúra előállítása, steril munka feltételei Immunsejtek izolálása és funkcionális vizsgálata 2019. április 15. Kremlitzka Mariann, PhD Miért tartunk fenn sejtkultúrákat? Egy sejt vizsgálat Kontrollált
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Mikrogliák eredete és differenciációja
 Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A COPD és foglalkozásegészségügyi
 A COPD és foglalkozásegészségügyi vonatkozásai Foglalkozás-egészségügyi Fórum 2011. szeptember 21. Dr. Kardos Kálmán A COPD-t, mint önálló betegségcsoportot először 1965-ben Briscoe említi. Kezdetben különböző
A COPD és foglalkozásegészségügyi vonatkozásai Foglalkozás-egészségügyi Fórum 2011. szeptember 21. Dr. Kardos Kálmán A COPD-t, mint önálló betegségcsoportot először 1965-ben Briscoe említi. Kezdetben különböző
A COPD gyógyítása, ápolása, gondozása a járóbeteg szakellátásban
 A COPD gyógyítása, ápolása, gondozása a járóbeteg szakellátásban Dr. Alföldi Ibolya Bp. XV.ker. TG 2014. 09. 12. Gordon Snider (1989): Chronic obstructive pulmonary disease a definition and implications
A COPD gyógyítása, ápolása, gondozása a járóbeteg szakellátásban Dr. Alföldi Ibolya Bp. XV.ker. TG 2014. 09. 12. Gordon Snider (1989): Chronic obstructive pulmonary disease a definition and implications
BD Vacutainer Molekuláris Diagnosztikai termékei
 BD Vacutainer Molekuláris Diagnosztikai termékei Andrea Süle, PhD Termékspecialista BD Diagnostics, Preanalytical Systems MOLSZE XI. Nagygyőlés, Pécs, 2009 augusztus 27-29. BD A BD egy orvostechnológiai
BD Vacutainer Molekuláris Diagnosztikai termékei Andrea Süle, PhD Termékspecialista BD Diagnostics, Preanalytical Systems MOLSZE XI. Nagygyőlés, Pécs, 2009 augusztus 27-29. BD A BD egy orvostechnológiai
ANTICANCER RESEARCH [Rákkutatás]
![ANTICANCER RESEARCH [Rákkutatás]](/thumbs/39/19250750.jpg "ANTICANCER RESEARCH [Rákkutatás]") ANTICANCER RESEARCH [Rákkutatás] Nemzetközi rákkutatási és rákgyógyászati folyóirat Az MGN-3/Biobran rizskorpából kivont módosított arabinoxilán in vitro növeli a metasztatikus emlőráksejtek érzékenységét
ANTICANCER RESEARCH [Rákkutatás] Nemzetközi rákkutatási és rákgyógyászati folyóirat Az MGN-3/Biobran rizskorpából kivont módosított arabinoxilán in vitro növeli a metasztatikus emlőráksejtek érzékenységét
Túlérzékenységi reakciók Gell és Coombs felosztása szerint.
 Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek
 A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
Antigén, Antigén prezentáció
 Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
4. A humorális immunválasz október 12.
 4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése.
 F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban
F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az anti-apoptózis mechanizmus vizsgálata agyi ischaemia/hypoxia modellekben
 OTKA T-037887 zárójelentés Az anti-apoptózis mechanizmus vizsgálata agyi ischaemia/hypoxia modellekben Az ischaemias stroke-ot követően az elzáródott ér ellátási területének centrumában percek, órák alatt
OTKA T-037887 zárójelentés Az anti-apoptózis mechanizmus vizsgálata agyi ischaemia/hypoxia modellekben Az ischaemias stroke-ot követően az elzáródott ér ellátási területének centrumában percek, órák alatt
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
GYERMEK-TÜDŐGYÓGYÁSZAT
 GYERMEK-TÜDŐGYÓGYÁSZAT I. 1. Légutak és a tüdő fejlődése 2. Légzőrendszer élettani működése 3. Újszülöttkori légzészavarok 4. Bronchopulmonalis dysplasia 5. A gége veleszületett és szerzett rendellenességei
GYERMEK-TÜDŐGYÓGYÁSZAT I. 1. Légutak és a tüdő fejlődése 2. Légzőrendszer élettani működése 3. Újszülöttkori légzészavarok 4. Bronchopulmonalis dysplasia 5. A gége veleszületett és szerzett rendellenességei
A psoriasis biológia terápiájának jelene és jövője. Holló Péter dr
 A psoriasis biológia terápiájának jelene és jövője Holló Péter dr Merre halad a psoriasis biológiai terápiája? Meglévő biológiai szerekkel új eredmények Hosszú távú hatásossági biztonságossági metaanalízisek
A psoriasis biológia terápiájának jelene és jövője Holló Péter dr Merre halad a psoriasis biológiai terápiája? Meglévő biológiai szerekkel új eredmények Hosszú távú hatásossági biztonságossági metaanalízisek
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok.
 Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA. Dr. Kurgyis Zsuzsanna
 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA PhD tézis Dr. Kurgyis Zsuzsanna Bőrgyógyászati és Allergológiai Klinika Szegedi Tudományegyetem Szeged 2017 2 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA
TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA PhD tézis Dr. Kurgyis Zsuzsanna Bőrgyógyászati és Allergológiai Klinika Szegedi Tudományegyetem Szeged 2017 2 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA
Az adaptív immunválasz kialakulása. Erdei Anna Immunológiai Tanszék ELTE
 Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Immunológia Világnapja
 a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN. DOKTORI (PhD) ÉRTEKEZÉS
 ÉRTEKEZÉS") A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN DOKTORI (PhD) ÉRTEKEZÉS RAPP JUDIT Pécsi Tudományegyetem Gyógyszerésztudományi Kar Gyógyszerészi Biotechnológia Tanszék Elméleti
A WNT JELÁTVITEL SZEREPE NEM KISSEJTES TÜDŐRÁK KIALAKULÁSÁBAN ÉS TERÁPIÁJÁBAN DOKTORI (PhD) ÉRTEKEZÉS RAPP JUDIT Pécsi Tudományegyetem Gyógyszerésztudományi Kar Gyógyszerészi Biotechnológia Tanszék Elméleti
A COPD keringésre kifejtett hatásai
 A COPD keringésre kifejtett hatásai Dr. Habil. Varga János Tamás Országos Korányi Pulmonológiai Intézet MTT Továbbképzés 2019 2019. január 25. A FEV1 csökkenés következményei Young R 2008 COPD-cluster
A COPD keringésre kifejtett hatásai Dr. Habil. Varga János Tamás Országos Korányi Pulmonológiai Intézet MTT Továbbképzés 2019 2019. január 25. A FEV1 csökkenés következményei Young R 2008 COPD-cluster
LÉGZÕRENDSZER. (Management of stable chronic obstructive pulmonary disease COPD) European Respiratory Society. Megjegyzések
 European Respiratory Society. Megjegyzések") A stabil krónikus obstruktív tüdôbetegség (COPD) kezelése (Management of stable chronic obstructive pulmonary disease COPD) European Respiratory Society A stabil COPD kezelése A kezelés céljai A tünetek
A stabil krónikus obstruktív tüdôbetegség (COPD) kezelése (Management of stable chronic obstructive pulmonary disease COPD) European Respiratory Society A stabil COPD kezelése A kezelés céljai A tünetek
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Immunológia alapjai előadás. A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és
 Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
PhD thesis. A Wnt mikrokörnyezet közvetlenül szabályozza a tüdő öregedéséhez vezető molekuláris eseményeket
 PhD thesis A Wnt mikrokörnyezet közvetlenül szabályozza a tüdő öregedéséhez vezető molekuláris eseményeket Kovács Tamás PhD témavezetők: Prof. Dr. Pongrácz E. Judit Dr. Kvell Krisztián PhD Program Vezető:
PhD thesis A Wnt mikrokörnyezet közvetlenül szabályozza a tüdő öregedéséhez vezető molekuláris eseményeket Kovács Tamás PhD témavezetők: Prof. Dr. Pongrácz E. Judit Dr. Kvell Krisztián PhD Program Vezető:
METASZTÁZISKÉPZÉS. Láng Orsolya. Kemotaxis speciálkollégium 2005.
 METASZTÁZISKÉPZÉS Láng Orsolya Kemotaxis speciálkollégium 2005. TUMOROK ÉS MIGRÁCIÓ PRIMER TUMOR METASZTÁZIS Angiogenezis Adhézió SEJT - SEJTCIKLUS Apoptózis Kemokinek Növekedési faktorok Szabályozó fehérjék
METASZTÁZISKÉPZÉS Láng Orsolya Kemotaxis speciálkollégium 2005. TUMOROK ÉS MIGRÁCIÓ PRIMER TUMOR METASZTÁZIS Angiogenezis Adhézió SEJT - SEJTCIKLUS Apoptózis Kemokinek Növekedési faktorok Szabályozó fehérjék
Az Oxidatív stressz hatása a PIBF receptor alegységek összeszerelődésére.
 Újabban világossá vált, hogy a Progesterone-induced blocking factor (PIBF) amely a progesteron számos immunológiai hatását közvetíti, nem csupán a lymphocytákban és terhességgel asszociált szövetekben,
Újabban világossá vált, hogy a Progesterone-induced blocking factor (PIBF) amely a progesteron számos immunológiai hatását közvetíti, nem csupán a lymphocytákban és terhességgel asszociált szövetekben,
Új terápiás lehetőségek (receptorok) a kardiológiában
 a kardiológiában") Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
Rövid és hosszútávú sejtkultúrák, funkcionális tesztek. Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem
 Rövid és hosszútávú sejtkultúrák, funkcionális tesztek Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem Bevezetés Definíció: az élő szervezeten kívül (in( vitro) ) tartunk élő sejteket Cél:
Rövid és hosszútávú sejtkultúrák, funkcionális tesztek Immunológiai és Biotechnológiai Intézet Pécsi Tudományegyetem Bevezetés Definíció: az élő szervezeten kívül (in( vitro) ) tartunk élő sejteket Cél:
FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a PD-L1 és PD-1 fehérjék túlélésre gyakorolt hatása
 Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
HLA-B27 pozitivitás vizsgálati lehetőségei
 HLA-B27 pozitivitás vizsgálati lehetőségei Pálinkás László, Uherkovichné Paál Mária, Berki Timea Pécsi Tudományegyetem Klinikai Központ Immunológiai és Biotechnológiai Intézet HLA-B27 Humán Leukocita Antigén
HLA-B27 pozitivitás vizsgálati lehetőségei Pálinkás László, Uherkovichné Paál Mária, Berki Timea Pécsi Tudományegyetem Klinikai Központ Immunológiai és Biotechnológiai Intézet HLA-B27 Humán Leukocita Antigén
Az ellenanyagok orvosbiológiai. PhD kurzus 2011/2012 II. félév
 Az ellenanyagok orvosbiológiai alkalmazása PhD kurzus 2011/2012 II. félév Ellenanyaggal működő módszerek Analitikai felhasználás Analitikai felhasználás Ellenanyag / antigén kapcsolódás Az Ab/Ag kapcsolat
Az ellenanyagok orvosbiológiai alkalmazása PhD kurzus 2011/2012 II. félév Ellenanyaggal működő módszerek Analitikai felhasználás Analitikai felhasználás Ellenanyag / antigén kapcsolódás Az Ab/Ag kapcsolat
Transzporterek vizsgálata lipidmembránokban Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest
 Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
Szakmai zárójelentés
 Szakmai zárójelentés A témavezető neve: dr. Antus Balázs A téma címe: A bronchiolitis obliterans szindróma pathomechanizmusa OTKA nyilvántartási szám: F 046526 Kutatási időtartam: 2004-2008. A kutatási
Szakmai zárójelentés A témavezető neve: dr. Antus Balázs A téma címe: A bronchiolitis obliterans szindróma pathomechanizmusa OTKA nyilvántartási szám: F 046526 Kutatási időtartam: 2004-2008. A kutatási
Monoklonális antitestek előállítása, jellemzői
 Monoklonális antitestek előállítása, jellemzői 2011. / 3 Immunológiai és Biotechnológiai Intézet PTE KK Antitestek szerkezete IgG Monoklonális ellenanyag előállítás: Hybridoma technika Lehetővé teszi kezdetben
Monoklonális antitestek előállítása, jellemzői 2011. / 3 Immunológiai és Biotechnológiai Intézet PTE KK Antitestek szerkezete IgG Monoklonális ellenanyag előállítás: Hybridoma technika Lehetővé teszi kezdetben
Zárójelentés. A) A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.
 A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.") Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
BETEGTÁJÉKOZTATÓ RHEUMATOID ARTHRITISBEN SZENVEDŐ BETEGEK SZÁMÁRA I. RHEUMATOID ARTHRITIS. origamigroup. www.origami.co.hu
 BETEGTÁJÉKOZTATÓ RHEUMATOID ARTHRITISBEN SZENVEDŐ BETEGEK SZÁMÁRA I. RHEUMATOID ARTHRITIS origamigroup www.origami.co.hu I. Rheumatoid arthritis Sokat hallunk napjainkban az immunrendszernek az egészség
BETEGTÁJÉKOZTATÓ RHEUMATOID ARTHRITISBEN SZENVEDŐ BETEGEK SZÁMÁRA I. RHEUMATOID ARTHRITIS origamigroup www.origami.co.hu I. Rheumatoid arthritis Sokat hallunk napjainkban az immunrendszernek az egészség
In vivo szövetanalízis. Különös tekintettel a biolumineszcens és fluoreszcens képalkotási eljárásokra
 In vivo szövetanalízis Különös tekintettel a biolumineszcens és fluoreszcens képalkotási eljárásokra In vivo képalkotó rendszerek Célja Noninvazív módon Biológiai folyamatokat képes rögzíteni Élő egyedekben
In vivo szövetanalízis Különös tekintettel a biolumineszcens és fluoreszcens képalkotási eljárásokra In vivo képalkotó rendszerek Célja Noninvazív módon Biológiai folyamatokat képes rögzíteni Élő egyedekben
Immunológia alapjai előadás. Sej-sejt kommunikációk az immunválaszban.
 Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
Genetikai panel kialakítása a hazai tejhasznú szarvasmarha állományok hasznos élettartamának növelésére
 Genetikai panel kialakítása a hazai tejhasznú szarvasmarha állományok hasznos élettartamának növelésére Dr. Czeglédi Levente Dr. Béri Béla Kutatás-fejlesztés támogatása a megújuló energiaforrások és agrár
Genetikai panel kialakítása a hazai tejhasznú szarvasmarha állományok hasznos élettartamának növelésére Dr. Czeglédi Levente Dr. Béri Béla Kutatás-fejlesztés támogatása a megújuló energiaforrások és agrár
TÁMOP /1/A
 Előadás száma Előadás címe Dia sorszáma Dia címe 1. Bevezetés 1. 2. Bevezetés 1. (Cím) 3. Történet 4. Jelátvitel 5. Sejt kommunikációs útvonalak 1. 6. Sejt kommunikációs útvonalak 2. 7. A citokinek hatásmechanizmusai
Előadás száma Előadás címe Dia sorszáma Dia címe 1. Bevezetés 1. 2. Bevezetés 1. (Cím) 3. Történet 4. Jelátvitel 5. Sejt kommunikációs útvonalak 1. 6. Sejt kommunikációs útvonalak 2. 7. A citokinek hatásmechanizmusai
Katasztrófális antifoszfolipid szindróma
 Katasztrófális antifoszfolipid szindróma Gadó Klára Semmelweis Egyetem, I.sz. Belgyógyászati Klinika Antifoszfolipid szindróma Artériás és vénás thrombosis Habituális vetélés apl antitest jelenléte Mi
Katasztrófális antifoszfolipid szindróma Gadó Klára Semmelweis Egyetem, I.sz. Belgyógyászati Klinika Antifoszfolipid szindróma Artériás és vénás thrombosis Habituális vetélés apl antitest jelenléte Mi
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai
 Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak