A MADARAK ANTIGÉNPREZENTÁLÓ DENDRITIKUS SEJTJEI
|
|
|
- Piroska Szekeresné
- 5 évvel ezelőtt
- Látták:
Átírás

1 A MADARAK ANTIGÉNPREZENTÁLÓ DENDRITIKUS SEJTJEI Doktori értekezés Igyártó Botond-Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Dr. Magyar Attila egyetemi adjunktus, Ph.D. Hivatalos bírálók: Dr. Balogh Péter egyetemi docens, Ph.D. Dr. Krenács Tibor tudományos főmunkatárs, Ph.D. Szigorlati bizottság elnöke: Szigorlati bizottság tagjai: Dr. Spät András egyetemi tanár, az MTA tagja Rajnavölgyi Éva egyetemi tanár, Ph.D., D.Sc. Tóth Sára egyetemi docens, Ph.D. Budapest 2006.
2 AZ ÉRTEKEZÉSBEN HASZNÁLT RÖVIDÍTÉSEK JEGYZÉKE... 4 BEVEZETÉS... 6 A dendritikus sejtek A dendritikus sejtek életciklusa A dendritikus sejtek a klinikumban A Langerhans sejt eredetének és funkciójának alakulása A Langerhans sejtek molekuláris jellegzetességei A Langerhans sejtek eredete A Langerhans-sejtek repopulációs kinetikája Gyulladásos bőrben Normál állapotban A Langerhans sejtek kivándorlását elősegítő változások A Langerhans sejtek további sorsa Antigénspecifikus aktiváció Antigénspecifikus tolerancia A madarak Langerhans-szerű sejtjei A follikuláris dendritikus sejtek Az emlős follikuláris dendritikus sejtek jellegzetességei és szerepei A follikuláris dendritikus sejtek eredete Különböző sejttípus, mint lehetséges FDC-prekurzor Feltételezett FDC-prekurzorok a vérben és csontvelőben Adatok, amelyek az FDC-k csontvelő eredetét erősítik Az FDC-k differenciálódását befolyásoló citokinek Az emlős- és madár lép jellegzetességei A csirke follikuláris dendritikus sejtek jellegzetességei CÉLKITŰZÉSEK ANYAG ÉS MÓDSZER Anyagok Állatok Hámnyúzatok készítése A Langerhans sejtek kimutatása ATPáz hisztokémiával Immunohisztokémia Elektronmikroszkópia Az epidermális dendritikus sejtek turn-overe A epidermális dendritikus sejtek izolálása és tenyésztése A Langerhans sejtek in vivo és in vitro aktivációja β-galaktozidázzal történő immunizálás menete β-galaktozidáz elleni immunválasz kvalitatív vizsgálata A β-galaktozidáz in vitro felvételének tanulmányozása Az ellipszoid-asszociált sejtek izolálása Az ellipszoid-asszociált sejtek antigénfelvételének tanulmányozása β-galaktozidáz gyöngyök előállítása Radioaktív receptorjelölés és izolálás Sejtfelszíni jelölés biotinnal és immunprecipitáció Elektroforézis és Western-blot A β-galaktozidáz receptorának azonosítása Az E5G12 mea karakterizálása
3 19. Monoklonális ellenanyagok előállítása Monoklonális ellenanyagok izotipizálása EREDMÉNYEK I Az epidermális dendritikus sejtek fenotípusa Az epidermális dendritikus sejtek elektronmikroszkópiája Az epidermális dendritikus sejtek izolálása és tenyésztése Az epidermális dendritikus sejtek ontogenetikája Az epidermális dendritikus sejtek funkciója Az epidermális dendritikus sejtek vándorlási útvonalának feltérképezése EREDMÉNYEK II A véletlen a tudomány szolgálatában Az E5G12 monoklonális ellenanyag szelekciója és vizsgálata A bakteriális antigént szelektíven az ellipszoid-asszociált sejtek veszik fel Az ellipszoid-asszociált sejtek vándorlása A humorális immunválasz mérése Az antigén felvételének in vitro vizsgálata A β-galaktozidáz specifikus receptor(ok)hoz kötődik Az EAC-k szerepe a baktériumfelvételben Az EAC-k nemcsak a β-gal-t veszik fel MEGBESZÉLÉS A csirke epidermális dendritikus sejtjei Az ellipszoid-asszociált sejt, mint follikuláris dendritikus sejt prekurzor A follikuláris dendritikus sejtek tényleg annyira különböznek a többi dendritikus sejttől? FDC-k eredete Follikuláris dendritkus sejt és antigén lokalizáció Follikuláris dendritikus sejt-dendritikus sejt, hasonló életciklus? ÖSSZEFOGLALÓ A madár EDC-kkel kapcsolatos eredményeink A madár FDC-kkel kapcsolatos eredményeink SUMMARY IRODALOMJEGYZÉK A DISSZERTÁCIÓHOZ KAPCSOLÓDÓ PUBLIKÁCIÓK A DISSZERTÁCIÓHOZ NEM KAPCSOLÓDÓ PUBLIKÁCIÓK KÖSZÖNETNYILVÁNÍTÁS AZ ÉRTEKEZÉSBEN HASZNÁLT SAJÁT KÖZLEMÉNYEK KÜLÖNLENYOMATAI
4 AZ ÉRTEKEZÉSBEN HASZNÁLT RÖVIDÍTÉSEK JEGYZÉKE Ag APS ATC ATP BAFF BC BCR BLC BM BrdU BSA CA CLA CLTA CR CSS DAB DAPI DBF DC DMSO EAC EAM EDC EDTA ELC ELISA FACS FBS Fc FDC FITC GC GM-CSF HEPES HEV HSA Hsp IC ICAM-1 idc antigén ammónium-perszulfát antigén szállító sejt (antigen transporting cell) adenozin-trifoszfát B-sejt aktiváló faktor (B-cell activating factor) Billroth-köteg (Billroth cord) B-sejt receptor (B-cell receptor) B lymphocyte chemoattractant bazális membrán 5-bromo-2-deoxiuridin marha szérum albumin (bovine serum albumine) centrális arteriola bőr leukocita antigén (cutaneous lymphocyte antigen) citotoxikus T-limfocita asszociált antigén (cytolitic T-lymphocyteassociated antigen) komplement receptor (complement receptor) Schweigger-Seidel hüvely (Capsule of Schweigger-Seidel) 3,3 -diamino benzidin 4,6-diamidino-2-fenilindol dibutil-ftalát dendritikus sejt (dendritic cell) dimetil-szulfoxid ellipszoid-asszociált sejt (ellipsoid-associated cell) ellipszoid-asszociált makrofág epidermális dendritikus sejt (epidermal dendritic cell) etiléndiamin-tetraecetsav Epstein Barr virus-induced molecule 1 ligand chemokine enzyme-linked immunosorbent assay fluorescence activated cell sorter újszülött borjúsavó (fetal bovine serum) kristályosítható fragmentum (fragment crystallizable) follikuláris dendritikus sejt (follicular dendritic cell) fluoreszcein-izotiocianát csíracentrum (germinal center) granulocita-monocita kolóniastimuláló faktor (granulocyte-monocyte colony stimulating factor) N-(2-Hidroxietil) piperazin-n -(2-etán szulfon sav) magas endotélű venula (High-Endothelial Venule) humán szérum albumin hősokk fehérje (heat shock protein) immunkomplex (immunecomplex) sejtközötti adhéziós molekula-1 (intercellular adhesion molecule-1) éretlen dendritikus sejt (immature dendritic cell)
5 RÖVIDÍTÉSEK JEGYZÉKE IDC interdigitáló dendritikus sejt IL interleukin ILT immunglobulinszerű transzkriptum INF interferon L limfocita LC Langerhans sejt (Langerhans cell) LFA limfocita funkcióhoz-asszociált antigén (lymphocyte function-associated antigen) LPS lipopoliszacharid MCP-1 monocita kemoatraktáns fehérje-1 (monocyte chemoattractant protein 1) mdc érett dendritikus sejt (mature dendritic cell) MDC makrofág eredetű kemokin (macrophage-derived chemokine) ME merkapto-etanol mea monoklonális ellenanyag MHCII fő hisztokompatibilitási komplex (major histocompatibility homplex) MIP-1α makrofág inflammatórikus protein 1α (macrophage inflamatory protein 1α) MIP-3α makrofág inflammatórikus protein 3α (macrophage inflamatory protein 3α) MIP-3β makrofág inflammatórikus protein 3β (macrophage inflamatory protein 3β) MZ marginális zóna MZDC marginális zóna dendritikus sejt OD optical density PALS periarteriolás limfoid hüvely (periarteriolar lymphoid sheat) PBS foszfáttal pufferolt fiziológiás sóoldat (phosphate buffered saline) Pc penicilliform kapilláris (penicilliform capillary) pdc plazmacitoid dendritikus sejt (plasmacytoid dendritic cell) PRR mintázatfelismerő receptor (pattern recognition receptor) PWP periarteriolás fehér pulpa (periarterial white pulp) RANTES Regulated upon Activation, Normal T-cell expressed and Secreted S szinuszoid (sinusoid) SC támasztó sejt (suportive cell) SCID severe combined immunodeficiency disease SDS nátrium dodecilszulfát SLC secondary lymphoid tissue chemokine Tc citotoxikus T-sejt (T-cytotoxic) TEMED N,N,N,N -tetrametil etiléndiamin TGF-β Transforming Growth Factor- β Th segítő T-sejt (T-helper) TLR Toll-szerű receptor (Toll-like receptor) TNF-α Tumor nekrózis faktor α Tr trabekula VCAM vaszkuláris sejtadhéziós molekula (vascular cell adhesion molecule) VLA-4 very late after activation 4 5
6 BEVEZETÉS BEVEZETÉS A DENDRITIKUS SEJTEK Amikor 1868-ban Paul Langerhans arany-impregnációval nyúlványokban gazdag sejteket talált a humán epidermiszben, még nem sejtette, hogy az immunrendszerhez tartozó dendritikus sejtek (DC) egyik jellegzetes képviselőjét tette láthatóvá. Sőt, idegsejteknek vélte, amelyek feltehetőleg a külvilágból érkező ingerek percepciójában játszanak szerepet (Langerhans 1868). Majdnem egy évszázadnak kellett eltelnie, míg valós kilétükre fényderült. Ekkorra nyilvánvalóvá vált, hogy ezek a sejtek az immunrendszer szerves részét képezik (Silberberg és mtsai 1976). Ugyancsak a XX. század közepe táján kezdték felismerni a DC-k immunválaszban betöltött szerepét. A DC-k részletes leírása és jellemzése Steinman és Cohn nevéhez fűződik (Steinman és Cohn 1973). A DC-k a nevüket az idegsejtek bokorszerűen elágazó dendritnyúlványaihoz való hasonlóságuk alapján kapták. A DC-ket majdnem valamennyi szervben megtalálták, az eloszlásuk azonban nem egyenletes. A gyakori fertőzés veszélynek kitett helyeken jóval nagyobb gyakorisággal fordul elő DC. Így nagy számban találunk DC-ket a bőrben, valamint a nyálkahártyák szintjén, míg a szívben vagy a vesékben kisebb mértékben képviseltetik magukat. Az ún. immunprivilegizált helyeken egyáltalán nem találunk DC-ket (pl. a szaruhártya centrális része, a központi idegrendszer). Ez az eloszlás előrevetíti a DC-k szerepét az immunrendszeren belül. A többi nyúlványos sejttel ellentétben (pl. fibroblasztok) a DCket nagymértékű anyagfelvétel jellemzi. A DC-k receptoraik révén teszik hatékonnyá a környezetükből való mintavételt. Ezen receptorokon keresztül ellenőrzik a körülöttük zajló eseményeket, míg nyúlványaik révén kiterjedt érintkezési felületet alakítanak ki. Ha netán egy szövetsérülés során baktériumok, vírusok kerülnek a szervezetbe, azokat a védvonalat jelentő DC-k hatékonyan fagocitálják. A megszerzett információt továbbadják az immunrendszer többi sejtjének. Szolúbilis molekulák révén nagy távolságokra lévő sejtekre gyakorolnak hatást, és a fertőzés helyszínére vonzzák. Közben a felvett idegen anyagokat elkezdik lebontani, majd az ezekből származó darabkákat (antigén epitóp) mintaként kihelyezik a felszínükre. Ezen antigén prezentáció révén az immunrendszer többi sejtjei is tudomást szereznek a betolakodóról 6
7 BEVEZETÉS és annak összetételéről. A DC-k felszínén megjelenő antigén darabkákat a limfociták ismerik fel. A bemutatott antigén felismerése után a limfociták aktiválódnak, egyesek ellenanyagokat kezdenek termelni (B-limfociták), míg mások ölőképessége fokozódik (Tc-limfociták). Az ellenanyagok specifikusak a betolakodókra, így azok semlegesítését ill. kiürülését segítik elő. Az aktiválódott T-sejtek képesek lesznek elpusztítani a betolakodók által megfertőzött sejteket. Lássuk, hogyan is zajlik mindez részleteiben. 1. A dendritikus sejtek életciklusa A különböző szervekben jelenlevő DC-k a csontvelői előalakokból jönnek létre. (Steinman és Cohn 1974, Katz és mtsai 1979). A csontvelőből származó sejteket általában két csoportba szokták sorolni: ún. limfoid-, ill. mieloid -ba. A limfoid vonalba tartoznak a limfociták, valamint a limfoid DC-k, míg a mieloidba a granulociták, monociták, vérlemezkék, vörösvértestek és a mieloid DC-k. Ez a két teljesen különválasztott vonalelmélet alapjait a csontvelői progenitorokon végzett vizsgálatok adták. Találtak olyan progenitorokat, amelyek vagy az egyik, vagy a másik vonalhoz tartozó sejteket voltak képesek létrehozni. Így leírtak ún. közös limfoid- (Kondo és mtsai 1997), ill. közös mieloid progenitorokat (Akashi és mtsai 2000). A csontvelőből a véráram útján a DC előalakok különböző szövetekbe vándorolnak. Itt pedig a DC-ket az expresszált CD (cluster of differentiation) antigének, valamint szöveti lokalizációjuk alapján minősítették limfoid-, ill. mieloid DC-knek (Shortman és Liu 2002). Ezeket a sablonokat nem lehet véglegesnek tekinteni, hiszen újabb eredmények szerint a régen limfoid vagy mieloid DC-k közös előalakból képződnek (Martinez del Hoyo és mtsai 2002, Martin és mtsai 2000, Traver és mtsai 2000, Manz és mtsai 2001, León és mtsai 2004). Nemrég olyan sejtpopulációt izoláltak az egér véréből, amely valamennyi DC-alpopulációt képes volt létrehozni (Martinez del Hoyo és mtsai 2002). A progenitorok plaszticitását bizonyítják azon adatok is, hogy találtak egy olyan ún. közös limfo-mieloid előalakot, amely képes volt a mieloid és limfoid vonalakhoz tartozó valamennyi sejt létrehozására, kivéve a vörösvérsejteket és trombocitákat (Adolfsson és mtsai 2005). A fentebb ismertetett adatok is azt mutatják, hogy ezen rendszer sokkal tágabb határokkal rendelkezik, minthogy ezt korábban gondolták. Feltehetőleg nem annyira a limfoid vagy mieloid vonalhoz való tartozás a fontos, hanem 7
8 BEVEZETÉS azok a faktorok, amelyeket a benépesített szövetek termelnek. Ezek a faktorok hatására különböző limfoid ill. mieloid vonalra jellemző molekulák kerülhetnek expresszióra a DC-ken. A csontvelői fejlődés után a klasszikus mieloid DC-előalakok a véráram útján jutnak el a különböző nem limfoid szervekbe (pl. bőr, nyálkahártya stb.). A rendeltetésük helyére megérkezett DC-ket ún. éretlen DC-knek nevezik. Itt a már említett detektor funkciót töltik be. Az éretlen DC-k a különböző kórokozókat felismerő receptorok széles repertoárjával rendelkeznek. Ezek között említhetjük a mintázatfelismerő receptorokat. Amint azt a nevük is mutatja, csak a patogénekre jellemző egyedi molekuláris mintázatokat ismerik fel. Ide tartoznak pl. az ún. C-típusú lektinek (pl. mannóz receptor, langerin stb.), amelyek főleg cukor oldalláncokat ismernek fel és a Toll-szerű receptorok (TLR). A TLR-ek lipoproteineket, lipopoliszacharidokat (TLR4), nukleinsavakat (TLR3,7,8,9) és különböző fehérjéket ismernek fel. Ezek a motívumok konzervált és fontos mikrobiális struktúrák, melyek nélkülözhetetlenek a túléléshez (Banchereau és mtsai 2000a, Pulendran és Ahmed 2006). Ezen receptorok a leghatékonyabb patogén felismerő és felvevő sejtté teszik a DC-ket. Nemcsak extracelluláris, hanem intracelluláris kórokozók felismerését lehetővé tevő receptorokkal is rendelkeznek (Akira és mtsai 2006). Ezeket az ún. caterpiller családba sorolják (Ting és Davis 2005). A mintázatfelismerő receptorokon kívül a DC-k Fc- és komplement receptorokat is hordoznak (Banchereau és mtsai 2000a). Az éretlen DC-kre nagymértékű endocitózis/fagocitózis jellemző, ugyanakkor a hatékony antigénprezentációhoz szükséges molekulákat kevésbé expresszálnak. Antigénfelvétel után, valamint a fertőzés, nekrózis során felszabadult inflammatórikus citokinek, (pl. TNF-α), bakteriális toxinok (pl. LPS), ill. Hsp-k hatására az éretlen dendritikus sejtek belépnek az ontogenézisük következő fázisába. Ennek a folyamatnak a végére a DC-k érett formában jelennek meg, amikor is az aktív fagocitózissal jellemezhető éretlen DCből hatékony antigénprezentáló sejtté válnak. Ezen folyamat során az antigénfelvételben szerepet játszó molekulák expressziója lecsökken, helyüket kostimulátor (CD80, CD86, CD40)-, ill. adhéziós molekulák (pl. CD54) veszik át. Az eddig főleg intracelluláris vezikulumokba lokalizálódott MHCII molekulák antigénnel feltöltve a plazmamembránba kerülnek. Ezek azok a molekulák, amelyek révén a kórokozóból származó antigéndarabok bemutatásra kerülnek a limfocitáknak. Az érést jellemző 8
9 BEVEZETÉS molekuláris átalakulásokkal párhuzamosan a DC-k elkezdenek vándorolni a perifériás nyirokszervekbe (pl. nyirokcsomó, Peyer-plakk stb.). A célszerv(ek) felé vándorlást az ugyancsak megváltozott kemokinreceptor profil biztosítja. A kemokin receptorok ugyanis azzal a szereppel vannak felruházva, hogy érzékeljék a környezetükbe jelenlevő csalogató (kemokin) molekulák jelenlétét. A kemokinek vakvezetőként működve kijelölik a vándorlási útvonalat és a célállomást. Az aktivált DC-k az éretlen DC-kkel ellentétben ún. kemokin receptor 7-et (CCR7) kezdenek expresszálni, amely érzékennyé teszi őket a MIP-3β (CCL19) kemokinre, elősegítve a DC-k nyirokszervekbe való vándorlását (Banchereau és Steinman 1998, Lipscomb és Masten 2002, Steinman és mtsai 2003, Colino és Snapper 2003). A nyirokszervekbe vándorolt érett DC képes mind a CD4, mind pedig a CD8 T-sejtek, valamint a B-sejtek stimulálására, függetlenül attól, hogy az antigén extra- vagy intarcelluláris eredetű (Dubois 1998, Lipscomb és Masten 2002, Heath és mtsai 2004, Groothuis és Neefjes 2005, Inaba és Inaba 2005, Sergio Trombetta és Mellman 2005, Bevan 2006). Így joggal állíthatjuk, hogy a DC a leghatékonyabb antigénprezentáló sejt, hiszen azonkívül, hogy valamennyi leukocita szabályozásában szerepet játszik, a legváltozatosabb eredetű antigének bemutatására képes. A fenti adatokkal a nem limfoid szövetekbe fellelhető DC-alpopulációk életciklusát szokták jellemezni. Gyakran figyelmen kívül hagyják, hogy a lép MZ-jában található DC-k hasonló érési cikluson esnek át, annak ellenére, hogy ők már éretlen korukban a limfoid szövetben foglalnak helyet. Ezek a DC-k a vérben megjelenő kórokozók kiszűrésében segítkeznek. Az antigének felvétele után a PALS területére vándorolnak, ahol IDC-kké érnek. Az előbb tárgyalt DC-ktől nagymértékben eltér az ún. plazmacitoid DCalpopuláció (pdc). Ezeket a sejteket nemrég még plazmacitoid T-sejteknek, majd később plazmacitoid monocitáknak nevezték. A pdc-k jól fejlett endoplazmatikus retikulummal rendelkeznek, ami a citoplazma nagy részét kitölti. Ez a plazmasejtek egyik jellegzetessége, de itt nem ellenanyagszintézis zajlik, hanem citokintermelés. Ezek a sejtek ugyanis rengeteg I. típusú interferont (INF) termelnek (3-10 pg/sejt/24 óra). Ezek közül a legfontosabb az IFN-α és a β. A mieloid DC-kkel ellentétben főleg a vírusfertőzésekben játszanak fontos szerepet, erre utal az is, hogy a különböző vírusmotívumokat felismerő receptorkészlettel rendelkeznek. Az INF-szintézisét a 9
10 BEVEZETÉS vírussal való találkozás indítja el. Ezen citokineknek révén a szomszédos sejtek értesülnek a vírus jelenlétéről, így felkészülhetnek a védekezésre. A vírus felvétele után ugyanakkor a pdc-k hatékony antigénprezentáló DC-kké érnek, vírusspecifikus immunválaszt indukálva. Nemcsak funkciójuk tekintetében térnek el a pdc-k a többi DC-alpopulációtól, hanem az életciklusuk is más. A csontvelői képződés után nem a perifériás szövetekbe, hanem a szekunder nyirokszervek T-dependens területeire vándorolnak, ahol a vírusok elleni immunválasz koordinálói lesznek. Fontos megjegyezni, hogy a pdc-ket limfoid eredetűnek tartják (Banchereau és mtsai 2000b, Liu 2005, Magyarics és Rajnavölgyi 2005). A különböző DC-alpopulációk szerepét és életciklusát az 1. kép szemlélteti. 2. A dendritikus sejtek a klinikumban Nemrég vált nyilvánvalóvá, hogy a DC-k az immunválasz kimenetelét is eldöntik. Az antigén természetétől függően nemcsak aktiválni képesek az immunkompetens sejteket, hanem anergiát (válaszképtelenség egy adott antigénre), sőt apoptózist is előidézhetnek bennük. Megfigyelték, hogy pl. az apoptótikus sejtet felvett DC nem aktiválódik, nem fog gyulladáskeltő citokineket termelni, sem pedig kostimulátor molekulákat; ennek következtében az így bemutatott antigén anergiát okoz a T-sejtekben. Ezen eredmények fényében nem meglepő, hogy nagyméretű kutatások folynak a DC alapú vakcinák területén. Ilyen, ún. tolerogén DC-ket nyerhetünk, ha in vitro differenciáltatott DC-ket bizonyos antigén jelenlétében IL10, ill. TGF-β-val kezelünk (Steinman és mtsai 2003). Az in vitro tolerogénné tett DC-k képesek in vivo anergiát kiváltani a recipiens szervezetben. Ezen módszerek megoldást jelenthetnek az autoimmun betegségekben szenvedő egyéneknek. Ugyanakkor, ha a DC-ket tumor antigénekkel töltjük fel és megfelelően aktiváljuk (pl. LPS, TNFα), akkor olyan sejteket kapunk, amelyek hatékony tumorspecifikus immunválaszt indukálnak. A tumorbiológia ezen területén már számos biztató eredmény áll rendelkezésünkre (Fong és Engleman 2000, Colino és Snapper 2003). Az eddig említettek alapján joggal tarthatjuk a DC-t az immunrendszer karmesterének. 10
11 BEVEZETÉS A dolgozatnak nem célja valamennyi DC-alpopuláció eredetét és funkcióját ismertetni. A dolgozat a munkánk során vizsgált két DC-alpopuláció részletesebb bemutatását kísérli meg. Ezen két alpopuláció közül az egyik az immunológiában modellként említett Langerhans sejt alpopuláció, míg a másik alpopulációba, az igencsak vitatott eredettel és szereppel felruházott follikuláris dendritikus sejtek kerülnek diszkutálásra. 11

12 12 BEVEZETÉS
13 BEVEZETÉS 1. kép A dendritikus sejtek életciklusa. A fertőzés színhelyén levő éretlen dendritikus (idc) sejtek PRRek révén hatékonyan felveszik a bejutott kórokozókat. A bakteriális toxinok (pl. LPS), valamint a nekrózis során felszabadult anyagok (TNF-α, Hsp-k) elősegítik az idc-k aktivációját és érését. Az aktiválódott idc-k a környező sejtekkel együttesen inflammatórikus citokineket és -kemokineket termelnek. Ezek a citokinek egyrészt más idc-ket aktiválnak, másrészt a gyulladásos környezet kialakításában játszanak szerepet. A kemokinek pedig a vérben keringő csontvelői DC prekurzorokat és más gyulladásos sejteket vonzanak a tett színhelyére. Az antigént felvevő DC a kellő aktivációt követően CCR7-et kezd expresszálni, ami elősegíti a környező nyirokcsomóba való vándorlását. Amint már említettük, a nyirokcsomóba érkezett DC időközben hatékony antigénprezentáló sejtté érik (mdc), így képes az immunkompetens sejtek aktivációjára. Azt itt lezajló folyamatok során az antigénre specifikus ellenanyagok termelődnek, és T-sejt populációk aktiválódnak. Feltételezik, hogy a DC-k végül apoptózissal elpusztulnak. A klasszikus mieloid prekurzorokkal ellentétben a plazmacitoid DC (pdc) nem perifériás szövetekbe, hanem a nyirokszervekbe vándorol (HEV-en keresztül). A pdc-knek kiemelt szerepet tulajdonítanak a vírusfertőzésekben. Vírusok hatására ezek a sejtek érett DC-vé (mdc) érnek és közben nagy mennyiségű INFα/β-t termelnek, ami a hatékony vírusellenes immunválasz alapját szolgálja. Ha a kórokozó a véráramba kerül, akkor azt a lép marginális zóna DC-k (MZDC) veszik fel, amelyek az előbb említett idc-khez hasonló tulajdonságokkal rendelkeznek. Az MZDC-k hasonló aktiváción/maturación esnek át, és interdigitáló DC-kké (IDC) érnek. Csak hipotézis szintjén lett berajzolva, hogy a MZDC-k iterdigitáló DC-ken keresztül vagy direkt módon follikuláris dendritikus sejtekké (FDC) differenciálódhatnak. A LANGERHANS SEJT EREDETÉNEK ÉS FUNKCIÓJÁNAK ALAKULÁSA A Langerhans sejtek (LC) leírójuk nevét viselik. Annak ellenére, hogy Arstein 1876-ban és Herxeimer 1896-ban hipotézis szintjén az LC-ket átutazó leukocitáknak képzelték, a technika akkori fejlettsége nem tette lehetővé elméletük bizonyítását (Arstein 1876, Herxeimer 1896). Az elkövetkező években különböző feltételezett eredettel és funkcióval ruházták fel az LC-ket. Ezek között említhetjük azt az elképzelést is, hogy az LC-k elhalt melanociták lennének (Wolff 1991). Nem sokkal azután, hogy Steinman és Cohn leírják a lép dendritikus sejtjeit, bebizonyosodik, hogy az LC-k az immunrendszer szerves részét képezik. Az is nyilvánvalóvá válik, hogy az LC-k fontos szerepet játszanak a hiperszenzitivitás kialakulásában (Silberberg és mtsai 1976). 1. A Langerhans sejtek molekuláris jellegzetességei Az LC-k számos, a többi mieloid dendritikus sejtekhez hasonló molekuláris mintázattal rendelkeznek. Ezek között említhetjük az MHCI, ill. MHCII molekulák konstitutív expresszióját (Romani és Schuler 1992), az antigének felvételében szerepet játszó-, kostimulátor-, valamint adhéziós molekulákat (Larregina és Falo 2005). Ezen közös jellemvonások mellett léteznek egyedi molekuláris jellegzetességek is. Ezek 13
14 BEVEZETÉS között említhetjük az E-kadherin expressziót, amely lehetővé teszi az LC-k megmaradását a keratinociták között, valamint a jellegzetes dendritikus alak létrejöttét (Tang és mtsai 1993). Másik jellegzetessége ezen sejteknek, hogy specifikus mintázatfelismerő receptort hordoznak, az ún. langerint (C-típusú lektin) (Valladeau és mtsai 1999). Ezen molekula elősegíti az ugyancsak LC-k azonosítására használt Birbeck granulumok kialakulását (Valladeau és mtsai 2000). Ezek a teniszütő alakú, trilamináris strukturák az endoszomális rendszerhez tartoznak, de pontos szerepük még nem ismert. Előzetes adatok az antigénfelvételben való szerepüket valószínűsítik (Valladeau és mtsai 2000, McDermott és mtsai 2004). Az előbbi molekulák mellett még meg kell említenünk a CD1a expressziót, valamint az ekto-atpáz aktivitást (Maurer és Stingl 1999, Kimber és mtsai 1999, Larregina és Falo 2005, Kissenpfennig és Malissen 2006). Az előbbi molekula a lipidtermészetű antigének bemutatásában vesz részt, míg az ekto-atpáz (CD39) aktivitás védelmet nyújt a keratinociták által szecernált ATP-vel szemben (Mizumoto és mtsai 2003 és 2004). Fontos megjegyezni, hogy mindezen markerek csak bizonyos kompromisszumok mellett alkalmazhatók az LC-k azonosítására, ugyanis expressziójuk nem teljesen korlátozódik ezen sejtekre. A humán LC-k molekuláris fenotípusát az 1. táblázat foglalja össze. 1. Táblázat A nyugvó és aktivált Langerhans sejtek fenotípusa (Módosítva Maurer és Stingl 1999) A Langerhans sejtek fenotípusa Morfológiai jellegzetességek Enzim profil Antigénprezentációban szerepet játszó molekulák Adhéziós molekulák Birbeck granulumok Rezidens Aktiválódott/ vándorló +++ +/- Nyúlványok ATPáz (CD39) +++ +/- Nemspecifikus észteráz + - MHCI + ++ MHCII CD1a CD1b - - CD1c + - CD1d - - E-kadherin ++ +/- Beta-1-integrin + vagy ++? 14
15 BEVEZETÉS Antigénfelismerő receptorok Kostimulátor molekulák/aktivációs markerek Fc-receptorok Beta-2-integrin + vagy ++? CLA + + vagy ++ CD44 (pan) + ++ CD11b ++? CD11c ++? Langerin (CD207) +++ +/- Mannóz receptor (CD206) - - DEC205 (CD205) TLR 2, 4, 9 ++? TLR 7 -? CD CD40 +/- + CD54 (ICAM-1) +/- ++ CD CD CD CD CD FcγRI (CD64), FcγRIII (CD16) FcγRII (CD32), FcεRI B-sejt markerek CD19, CD T-sejt markerek Citokin receptorok CD4 +? CD3-TCR, CD8 - + (CD8; Anjuere és mtsai 2000) GM-CSFRα + ++ GM-CSFRβ +/- ++ TNF-RII + +/- IL-1RI, IL-1RII + +/- ill. ++ CCR6 + - CCR A Langerhans sejtek eredete 15
16 BEVEZETÉS Az epidermiszt az LC-k, mint éretlen dendritikus sejtek népesítik be. Az összepidermális sejtek 2-4 %-át teszik ki. Számuk humán epidermiszben /mm 2 között ingadozik (Chen és mtsai 1985). Szenzorként is felfoghatjuk őket, hisz folyamatosan válaszolnak a környezetükből érkező antigénekre, legyenek azok saját vagy idegen természetű anyagok. A felvett antigént a legközelebbi nyirokcsomóba szállítják, ahol bemutatják az immunkompetens sejteknek. Alapállapotban az LC-k számát főleg a lokális osztódások tartják megfelelő szinten. Ez megnehezítette az LC-k eredetének vizsgálatát, mivel nem lehetett megfigyelni az esetleges prekurzorok hámba való vándorlását. Ez a probléma azonban megoldódott, amikor a kutatók gyulladásos hám LC-ket vizsgálták. Ekkor ugyanis kiderült, hogy a gyulladásos folyamatok során kiürült hámot ezen osztódások nem képesek benépesíteni, ezért itt fontos szerephez jutnak az extra epidermális prekurzorok (Merad és mtsai 2002, Larregina és mtsai 2001). Csontvelőátültetéssel Katz és munkatársai igazolták először, hogy ezek a prekurzorok a csontvelőből származnak. (Katz és mtsai 1979). Ilyen előalakok a véráramban is jelen vannak, az egyik, már bizonyított állomásozó hely a dermisz (Larregina és mtsai 2001). Összefoglalva, a csontvelői eredetű LC-progenitorok a véráram útján eljutnak a dermiszbe. Itt egy olyan populációt alkotnak, amely a gyulladást követő fázisban nagyban hozzájárul az elvándorolt LC-k pótlásához. A progenitorok sejtvonal eredete még mindig éles viták tárgyát képezi. Nagy általánosságban elmondható, hogy az LC-k mind limfoid, mind pedig mieloid előalakokból létrejöhetnek, azonban az LC-k mintegy 90 %-a mieloid eredetű (Anjuére és mtsai 2000, Mende és mtsai 2006). Ami biztosra vehető az LC-k ontogenézisében, hogy az epidermisz által termelt TGF-β1 fontos szerepet játszik ezen sejtek érésében, hiszen TGF-β1 hiányos egerekben nem azonosíthatók LC-k az epidermiszben (Borkowski és mtsai 1996). A TGF-β1 egyrészt a Runx3 transzkripciós faktoron keresztül fejti ki hatását, így nem meglepő, hogy az LC-k Runx3 -/- egerekben is hiányoznak (Fainaru és mtsai 2004). A Runx3 -/- egerek dendritikus sejtjei rezisztensek a TGF-β1-re nézve. Érdekes módon az Ikaros és relb transzkripciós faktorokba bekövetkezett mutációk nem gyakorolnak jelentős hatást az LC-kre (Zenke és Hieronymus 2006) annak ellenére, hogy a többi dendritikus alpopulációba tartozó sejtek száma drasztikusan lecsökken. 16
17 BEVEZETÉS 3. A Langerhans-sejtek repopulációs kinetikája 3.1. Gyulladásos bőrben Az LC-k kinetikáját mind gyulladásos, mind pedig normális fiziológiás körülmények között vizsgálták. Legtöbb időt az LC-k mobilizációját kiváltó faktorok tanulmányozásának szenteltek. A gyulladásos folyamatokat proinflamatorikus citokinek és kemokinek szekréciója jellemzi. Ezek közül említésre méltók az IL-1β, a TNF-α és a MIP-3β, amelyek fontos szerepet játszanak az LC-k vándorlásának előidézésében (Enk és mtsai 1993a, Cumberbach és mtsai 1994, 1995, 1997, Schwarzenberger és Udey 1996, Charbonier és mtsai 1999). Kevésbé ismertek azok a mechanizmusok, amelyek az LC progenitoroknak a szövetspecifikus homingját teszik lehetővé. Eddigi kutatási eredmények a kemokinek fontos szerepét hangsúlyozzák. Ezek a kemoatraktáns citokinek G-proteinekhez kapcsolt receptorok révén fejtik ki hatásukat. Köztudott, hogy az in vitro differenciáltatott LC/DC-k nagyon érzékenyek a következő kemokinekre: MIP-1, MCP- 1, MDC, RANTES. Nemrég bizonyították a MIP-3α szerepét a dendritikus előalakok migrációjában. Ez a kemokin az LC/DC-ken jelenlevő CCR6 receptoron keresztül hat. Ezen kemokinnek fontos szerepet tulajdonítanak a dendritikus előalakok gyulladásos helyre való toborzásában. Továbbá, elősegítheti a gyulladásos folyamat során kiürült epidermisz LC előalakokkal való benépesülését (Greaves és mtsai 1997, Maurer és Stingl 1999, Charbonnier és mtsai 1999) Normál állapotban A gyulladásos esetekkel ellentétben, vajmi keveset tudunk azokról a szignálokról, amelyek alapállapotban szerepet játszhatnak az LC-k emigrációjában. Az LC-k elvándorlásának kinetikája nagymértékben különbözik ez előbb említett két esetben (Morelli és mtsai 2001). Míg gyulladás esetén az LC-k száma drasztikusan lecsökken, addigra normális állapotban a sejtek elvándorlásának foka az észlelhetőség határa alatt marad (Jakob és mtsai 2001). Az elvándorolt sejtek pótlása is 17
18 BEVEZETÉS különbözőképpen történik. Amint az már említésre került, normál állapotban az LC-k számát a lokális osztódások tartják megfelelő szinten. Ezzel ellentétben a gyulladásos folyamatok miatt elvándorolt LC-k pótlása a csontvelői progenitorok feladata. A legtöbb kutatás a gyulladásos folyamatok során fellépő prekurzor toborzást vizsgálta, amelyek többé-kevésbé feltárták ezen folyamat lépéseit. Ezzel ellentétben nemrég derült fény arra, hogy hogyan történhet a prekurzorok megfelelő helyre való csalogatása normális fiziológiás körülmények között. Kimutatták ugyanis, hogy az epidermisz sejtjei konstitutívan termelnek egy olyan kemokint, amely szelektív módon az LC-prekurzorokra hat, elősegítve azok hámba való vándorlását és érését. Ez a kemokin a CXCL14 nevet viseli, de receptora még nem ismert (Schaerli és mtsai 2005). 4. A Langerhans sejtek kivándorlását elősegítő változások A klasszikus modell szerint az LC-k járőrszolgálatot teljesítenek a bőrben. Az elágazó nyúlványaik révén nagy felületet képeznek, amely megnöveli a kórokozókkal való interakció esélyét. Mihelyt kontaktusba kerülnek bizonyos antigénnel és megfelelő vészjelzéseket is kapnak, a sejtek aktiválódnak. Az aktivációt követően az LC-k epidermiszből való kivándorlását bizonyos citokinek is segítik. Az LC-k epidermiszből való kivándorlásában két citokin hatását bizonyították (Cumberbatch és mtsai 1994 és 1997). Az egyik citokint, a TNF-α-t csak a keratinociták termelik, míg a IL-1β-t kizárólag az LC-k képesek termelni. Az IL-1β a szomszédos hámsejtekben TNF-α termelést képes indukálni. Az így nagy mennyiségben termelődött TNF-α számos molekula expresszióját befolyásolja. Az IL-1β-hoz hasonlóan az E-kadherin fehérjék szintézisét gátolja, elősegítve az LC-k elszakadását a környező hámsejtektől (Schwarzenberger és Udey 1996). Továbbá, más adhéziós molekulák fokozott megjelenését is a TNF-α hatásának tulajdonítják. Ilyen adhéziós molekula az ICAM-1, amely nemcsak a sejtek immunstimulátor képességét növeli, de kimutatták, hogy az LCk nyirokcsomókban való akkumulációjában is fontos szerephez jut (Cumberbatch és Kimber 1992, 1995, Ma és mtsai 1994). A másik adhéziós molekula, amelynek expressziós mintázatát némiképp a TNF-α befolyásolja, a CD44 (hialuronsav receptor). Nem teljesen tisztázott ezen molekula szerepe az LC-k kivándorlásában, de a TNF-α hatására a CD44 olyan izoformája (v9) kerül kifejezésre, amely részt vehet a sejtek 18
19 BEVEZETÉS hámból való kivándorlásában (Sallusto és Lanzavecchia 1994). Ezt bizonyítják azon vizsgálatok, ahol ezen izoformát felismerő ellenanyag kezelés esetén az LC-k kivándorlása elmaradt (Weiss és mtsai 1997). Bizonyították, hogy makrofágok esetében az előbb említett két citokin kollagenáz szintézist indukál (Saren és mtsai 1996). Így feltehetőleg az aktivált az LC-k által termelt mátrix metaloproteináz 9 (IV típusú kollagenáz) is e két citokin szabályozása alatt áll (Kobayashi 1997). Ez az enzim lehetővé teszi az LC-k bazális membránon való áthaladását, míg a lamininnel való kölcsönhatást a VLA-6 (α6β1) antigén biztosítja (Price és mtsai 1997). 5. A Langerhans sejtek további sorsa 5.1. Antigénspecifikus aktiváció Amint azt az előbbi részben láttuk, citokinek hatására az LC-k molekuláris mintázata megváltozik. Ez lehetővé teszi az LC-k elszakadását a környező keratinocitáktól, és a sejtek az életciklusuk következő fázisába lépnek. Ebben az ún. érési fázisban a sejtek elszakadva az epidermisztől a nyirokcsomók felé vándorolnak, miközben hatékony antigénprezentáló sejtekké érnek. Ezen folyamat során az éretlen LC-kben nagyrészt citoplazmatikus vezikulumok membránjába jelenlevő MHCII molekulák (Mommaas és mtsai 1995) parakrin vagy autokrin stimulusok hatására (GM- CSF, TNF-α, IL-1β, LPS és/vagy CD40 keresztkötése) a sejtfelszínen jelennek meg (Cella és mtsai 1997, Sallusto és mtsai 1995). A maturáció további jellegzetessége, hogy nagymennyiségű kostimulátor molekula kerül expresszióra. Itt említhetjük meg a CD80, CD86 antigéneket. Továbbá, a T-sejtek működését befolyásoló citokinek termelése is fokozódik (pl. IL-12), ugyanakkor az antigének felvételében szerepet játszó receptorok mennyisége csökken (Romani és mtsai 1989, Schuler és Steinman 1985, Steinman 1991, Schreiber és mtsai 1992, Kang és mtsai 1996, Koch és mtsai 2006). Ezen aktiváció során a sejtek új kemokin receptort (CCR7) kezdenek expresszálni, amely lehetővé teszi a sejtek nyirokcsomókba való vándorlását. Itt a felvett antigénre specifikus immunválaszt indukálnak (Streilein és mtsai 1979, Toews és mtsai 1980, Condon és mtsai 1996, Casares és mtsai 1997, Reis e Sousa 2001, Romani és mtsai 2003, Larregina és Falo 2005). 19
20 BEVEZETÉS Mindezen modellt megerősítő érvek mellett olyanok is napvilágot láttak, amelyek a bőrimmunitásról kialakult elképzelésünk továbbfejlesztését sürgetik. Kimutatták ugyanis, hogy olyan vírusok esetében, amelyek a bőrön vagy nyálkahártyákon keresztül fertőznek, nem az LC-k, hanem más DC populációk jelentik a hatékony védelmet (Allan és mtsai 2003, Zhao és mtsai 2003). Másrészt, genetikai módszereket használva, azt is bebizonyították, hogy az LC-k szelektív eliminációja esetén a kontakt hiperszenzitivitás kiváltható (Bennett és mtsai 2005, Kaplan és mtsai 2005). Arra is fény derült, hogy a bőr felületére juttatott haptén először a dermiszben levő DC-k közvetítésével jut el a nyirokcsomóba, míg a haptén pozitív LC-k majdnem egy nappal lemaradva jelennek meg a perifériás nyirokszervekben. Ez az időbeli eltolódás feltehetőleg azzal magyarázható, hogy az LC-k szorosabb kötelékből kell, hogy kiszabaduljanak, mint a dermisz DC-k. A két sejtpopuláció szöveti lokalizációja is különböző: míg a dermiszből származó DC-k a parakortex régió kéreg felé eső részében tömörülnek, addig az LC-k a velőhöz közelebbi területen csoportosulnak (Kissenpfennig és mtsai 2005, Kissenpfennig és Malissen 2006) (2. kép) Antigénspecifikus tolerancia Az LC-k tolerogén funkciója kevésbé ismert, de több adat is utal arra, hogy ezek a sejtek a gyulladás hiányában vészszignál nélkül- folyamatosan epidermális antigéneket (saját) vesznek fel (Parr és mtsai 1991). Egyik ilyen antigén a melanin, amelyet arra használtak, hogy azonosítsák a normál állapotban kivándorolt LC-ket (Hemmi és mtsai 2001, Yoshino és mtsai 2003). Ez a folyamatos saját antigén szállítás nem vezet autoimmun folyamatokhoz, ami felvetette annak lehetőségét, hogy az LC-k a tolerancia fenntartásában is fontos szerepet játszanak. Az apoptótikus sejtek felvétele után -még nem teljesen ismert módon- a DC-k tolerogén sejtekké alakulnak. Ahhoz, hogy egy DC tolerogénné váljon, megfelelő citokinekre is szükség van. Ilyen citokinek az IL-10 (Enk és mtsai 1993b) (a hámsejtek UVB hatására nagymértékben termelik) és TGF-β. Ezen citokinek egyrészt gátolják a proinflamatorikus citokinek termelését (IL-6, IL-1β, IL-12, silt4), valamint a kostimulátor molekulák (CD80, CD86) expresszióját, másrészt immunglobulinszerű transzkriptumok (pl. ILT4) fokozott kifejezését indukálják. Ezek a molekulák a T sejtek felé tolerogén szignált közvetítenek, és 20

21 BEVEZETÉS elősegítik a regulatorikus T sejtekké való differenciálódást. A regulatorikus T sejtekkel való kölcsönhatás is szerepet játszik a tolerancia kialakításában. A regulatorikus T sejtek membránjában jelenlevő CTLA-4 nagy affinitással kötődik a DC-k membránjában jelenlevő CD80 vagy CD86 molekulákhoz. Ezen kapcsolódás IFN-γ szekrécióhoz vezet, amely autokrin módon azonnal indolamin 2,3-dioxigenáz enzim termelését indukálja. Ez az enzim a környezetben levő triptofánt lebontja, és az így keletkező bomlástermékek végül a környező T sejtek apoptózisát okozza (Morelli és mtsai 2001, Steinman és mtsai 2003, Wallet és mtsai 2005). Egyes neuropeptidek (calcitonin gene related peptid, melanocita stimuláló hormon) is képesek befolyásolni a DC-k működését. Ezen neuropeptidek képesek az éretlen DC-ket toleranciát indukáló sejtekké differenciáltatni (Hosoi és mtsai 1993, Grabbe és mtsai 1996). 2. kép A bőr dendritikus sejtjeinek szerepe és a vándorlási célpontok. A legfrissebb eredmények szerint mind az LC-k, mind pedig a dermisz DC-k részt vesznek a bőrimmunitásban. Az intakt bőr felületére jutott hapténeket, kisebb molekulákat, vagy a sérülések árán bekerült antigéneket az előbb említett mindkét DC-alpopuláció hatékonyan felveszi. A felvételt követően a már többször említett érési folyamaton esnek át, míg specifikus immunválaszt indukálnak. A két DC-alpopuláció eltérő gyorsasággal kolonizálja a perifériás nyirokcsomókat. Először a dermisz DC-k jelennek meg és a nyirokcsomó B-sejtes régióhoz közelebb vernek tanyát, míg az LC-k a parakortex velőhöz közelebbi régióban tömörülnek (a képen nem látható). Ez az elhelyezkedés feltehetőleg magyarázza ezen sejtek limfocitákkal való 21
22 BEVEZETÉS kölcsönhatását is, hiszen kimutatták, hogy csak a dermisz DC-k képesek a B-limfociták aktivációjára is. Az elvándorolt DC-k alapállapotban lokális osztódással pótlódnak, míg nagyobb mértékű (pl. UVB, gyulladás hatására) elvándorlás esetén a dermiszben helyet foglaló csontvelői progenitorok állítják vissza az egyensúlyt. A nyirokcsomóban más DC-alpopuláció is megtalálható: DC(1), DC(2) és plazmacitoid dendritikus sejtek. (Kissenpfennig és Malissen 2006 után) 6. A madarak Langerhans-szerű sejtjei Sokáig az az elképzelés uralkodott, hogy a madarak epidermiszében nincsenek DC-k, hiszen hisztokémiai festésekkel nem tudták kimutatni jelenlétüket. Immunhisztokémiai festéseket meg nem végeztek. Ez azonban a 90-es évek elején megdőlt, amikor is egy mexikói munkacsoport ATPáz + dendritikus sejteket írt le a madár epidermiszben (Carrillo-Farga és mtsai 1991). Később ugyanez a csoport kimutatta, hogy ezek a sejtek MHCII-t expresszálnak (Akhter és mtsai 1993, Perez- Torres és mtsai 1994) és Birbeck-granulumokat (Perez-Torres és mtsai 2001) hordoznak. Ezek a sejtek fenotípusuk alapján hasonlítanak az emlősök epidermiszéből ismert Langerhans sejtekhez. Ezen hasonlóság alapján kapták a Langerhans-szerű elnevezést. Hasonló morfológiai bélyegekkel rendelkező sejteket a nyálkahártya szintjén is azonosítottak (Perez-Torres és mtsai 2002, Nagy és mtsai 2005a). Azonban mindezen morfológiai vizsgálatok nem adtak kielégítő választ ezen sejtek eredetét és funkcióját tekintve. Ahhoz, hogy ezeket a sejteket teljes mértékben LC-nek tekintsük, elengedhetetlen az alapos morfo-funkcionális jellemzésük. Ugyancsak érdekes lehet vándorlásuk célállomásának azonosítása, ha bebizonyosodik, hogy funkciójukat tekintve is hasonlítanak az emlős LC-khez, azon tény fényében, hogy a madarak nem rendelkeznek perifériás nyirokcsomókkal. A FOLLIKULÁRIS DENDRITIKUS SEJTEK 1. Az emlős follikuláris dendritikus sejtek jellegzetességei és szerepei A follikuláris dendritikus sejtek (FDC) a follikulusban (főleg a csíracentrumokba) foglalnak helyet. Jellegzetességei között említhetjük a dendritikus külalakot, valamint hogy immunkomplexeket (IC) kötnek a felszínükön (Tew és mtsai 1990, 1997). Ugyancsak tankönyvi adat, hogy ezek a sejtek nem fagocitálnak, 22
23 BEVEZETÉS szabálytalan alakú eukromatikus maggal rendelkeznek, komplement receptorokat (CR1, CR2), Fc-receptort (FcγRIIB) és adhéziós molekulát (ICAM-1/CD54) hordoznak (Hanna és Szakal 1969, Szakal és Hanna 1968, Gerdes és mtsai 1983, Schriever és mtsai 1989, Sellheyer és mtsai 1989, Petrasch és mtsai 1990, Maeda és mtsai 1992, Balogh és mtsai 2001). A humán FDC-k molekuláris jellegzetességeit a 2. táblázat foglalja össze. Az idők során számos névvel illették az FDC-ket, így ismeretes a dendritikus retikulum sejt, a follikuláris IC-kötő sejt vagy a dendritikus makrofágok elnevezés, mindaddig, amíg a jelenlegi nevet egyezményesen el nem fogadták (Tew és mtsai 1982). 2. Táblázat Az "érett" FDC-k fenotípusa (Schriever és mtsai 1989 után módosítva) A follikuláris dendritikus sejtek fenotípusa Humán (mandula) Morfológiai Dendritikus sejtalak ++ jellegzetességek IC ++ Hemopoetikus marker CD45 +/- MHCI ++ Antigénprezentációban szerepet játszó molekulák Adhéziós molekulák Mieloid markerek Fc-receptorok Komplement receptorok MHCII ++ CD1a - CD1b - CD1c - CD1d? ICAM-1 ++ VCAM-1 ++ VLA3,4,5,6 + CD11b ++ CD11c - CD14 + CD13 - CD33 - CD11b (CR3) + FcγRIIB ++ FcγRIII (CD16), FcεRII (CD23) + CR1(CD35) ++ CR2(CD21Long) ++ CR3(CD11b) ++ 23
24 BEVEZETÉS B-sejt markerek CD19, CD21 ++ T-sejt markerek Citokin receptorok Citokinek CD4 - CD3-TCR, CD8 - GM-CSFRα + TNF-RI + TNF-RII - IL-1RI, IL-2Rα - IL-2Rβ, IL-4R, IL-6R + TGF-β + BAFF ++ Már az 50-es években megfigyelték, hogy a nyiroktüszők csapdába ejtik az antigéneket (Hill és mtsai 1950). Ezt a folyamatot kezdetben a limfocitáknak tulajdonították, de a 60-as évek közepe felé autoradiográfiás módszerrel kimutatták, hogy a protein antigén a csíracentrumokra lokalizálódik, és a specifikus ellenanyag elősegíti a csapdába ejtést (Ada és mtsai 1964, Nossal és mtsai 1964). Előzetes adatok szerint az antigén olyan fagocitáló retikulumban foglal helyet, amelyet fagocita sejtek alkotnak (Miller és Nossal 1964). Ezen korai elméletet elvetették, amikor az ultarstrukturális vizsgálatok azt mutatták, hogy az antigén nem került felvételre, hanem az elágazó nyúlványok felszínéhez tapadt (Szakal és Hanna 1968). A tankönyvi modell szerint a FDC-k passzív szereppel vannak felruházva (3. kép). A tüszőbe szállított IC-ket csapdába ejtik (főleg a MZ B-sejtjeinek tulajdonítják ezt a feladatot) és fogvatartják a membránjukban levő komplement- és Fc-receptorok segítségével (Brown és mtsai 1970, Gray és mtsai 1984, Heinen és mtsai 1986, Oldfield és mtsai 1988, Ferguson és mtsai 2004). Ezen IC-k számos folyamatban szerepet játszanak, így pl. a csíracentrumképződésben, a B-sejt szelekcióban, izotípusváltásban, affinitásérésben stb. (Kosco és mtsai 1992, Liu és mtsai 1996, Tew és mtsai 1997, Tew és mtsai 2001). Számos, az általánosan elfogadott modellt feszegető adat látott és lát napvilágot. Egyik ilyen ellentmondó adatokat leíró közlemény a 80-as évek elején jelent meg. Ebben a közleményében Kroese normális csíracentrum képződést ír le, még mielőtt az FDC-k csapdába ejtenék a képződött IC-ket (Nieuwenhuis és mtsai 1984, Kroese és mtsai 1986). Az FDC-k felszínén megtartott antigén immunológiai memóriában játszott szerepe is megkérdőjeleződött. Maruyama és mtsai genetikailag módosított egereket használva, egy nagyon elegáns kísérlettel kimutatták, hogy az 24
. Egy másik munkacsoport is alátámasztotta ezeket az eredményeket.")
25 BEVEZETÉS antigén jelenléte nem szükséges a memória fenntartásához. A Cre-rekombináz adta lehetőséget kihasználva a memória B-sejtek BCR-jének specificitását megváltoztatták. Ez a változtatás nem vezetett a memória sejtek gyorsabb elpusztulásához (Maruyama és mtsai 2000). Egy másik munkacsoport is alátámasztotta ezeket az eredményeket. Munkájukhoz olyan egereket használtak, amelyek nem voltak képesek immunglobulin szekrécióra, ami lehetőséget adott számukra az IC-k szerepének vizsgálatára. Meglepő módon azt tapasztalták, hogy detektálható IC hiányában normális csíracentrum képződés megy végbe, sőt a BCR-ek affinitás érése is hasonló gyakorisággal fordul elő, mint a kontroll állatokban (Hannum és mtsai 2000, Haberman és Shlomchik 2003, Anderson és mtsai 2006). 3. kép. Az FDC B-limfocita, ill. B-limfocita T-sejt közötti kommunikációban résztvevő molekulák. Az FDC-k által fogvatartott IC-t a B-sejt BCR-je ismeri fel, míg a CD21 az IC-n jelenlevő ligandumát köti (CD21L). Ezek a kötődések a B-limfociták aktivációját és differenciálódását idézik elő. Az FDC-kből származó ún. iccosomákat a B-sejtek felveszik és a T-sejteknek mutatják be. A T-sejt a CD40L-en keresztül további stimuláló jeleket közvetít a B-sejt irányába. (Tew és mtsai 2001, után) 2. A follikuláris dendritikus sejtek eredete Nemcsak az FDC szerepe, de pontos eredete sem tisztázott kellőképpen. Felvetődik a kérdés, hogy tárgyalhatjuk-e úgy ezen sejteket, mint az immunrendszer 25
26 BEVEZETÉS dendritikus sejtjei közül valókat. Az immunológusok nagy része még ma sem tekinti valódi dendritikus sejteknek, ugyanis az FDC-k retikulumsejtekhez hasonló morfológiai jellegekkel rendelkeznek. Ugyanakkor számos olyan markert hordoznak, amelyek jelen vannak retikulumsejteken, endotélsejteken, pericitákon és monocitákon. Ezek a közös morfológiai bélyegek és/vagy markerek sugallták, hogy mindezen sejttípusok FDCprekuzorként szolgálhatnak. 3. Különböző sejttípus, mint lehetséges FDC-prekurzor 1980-ban Heusermann és munkatársai fény- és elektronmikroszkópos vizsgálatokkal olyan sejtalakokat azonosítottak nyúl lépben, amelyek feltehetőleg átmeneti formákat képviseltek a retikulumsejtek és az FDC-k között (Heusermann és mtsai 1980) ban Rademakers és kollégái kimutatták, hogy a periciták és FDC-k hasonló alkalikus foszfatáz aktivitással rendelkeznek (Rademakers és mtsai 1988). Castleman betegek nyirokcsomóiban FDC-szerű sejteken DRC-1- és endoteliális leukocita adhéziós I antigéneket detektáltak, amiből logikusan arra következtettek, hogy az FDC-k endotél sejtekből származnak (Ruco és mtsai 1991). A legújabb közlemény, amely összefüggésben hozza a fibroblasztokat az FDC-kkel, Bofill és kollégáitól származik. Itt egy újonnan leírt fibroblaszt markert vetettek be az FDC-k eredetének eldöntésére. Előzetes tanulmányok szerint az ún. AS02 ellenanyag csak a humán fibroblasztokat festette. Munkájuk során elszórtan találtak olyan FDC-ket, amelyek ezen ellenanyag által felismert antigént hordozták. Ezen eredményekkel alátámasztani vélték az FDC-k fibroblaszt eredetét (Bofill és mtsai 2000). Azonban fontos megjegyezni, hogy az előbb említett ellenanyag a Thy-1 (CD90) antigént ismeri fel, amely többek között a DC-ken is expresszióra kerül (Saalbach és mtsai 1998). Egy teljesen más irányból próbálták megfejteni az FDC-k eredetét azok, akik transzfer-kísérletekkel próbálkoztak. Kezdetben ezen transzfer-kísérletek, amelyek során csontvelő sejteket vittek át a recipiens állatokban, az FDC-k nem csontvelő eredetét támogatták. Humphrey és munkatársai besugarazott egereken allogén csontvelőtranszplantációt végeztek, és azt tapasztalták, hogy a lép FDC-k egy év múlva is recipiens MHCII-őt hordoztak a felszínükön. Ezzel ellentétben a limfociták és makrofágok donor eredetűek voltak. (Humphrey és mtsai 1984). Hasonló eredményeket 26
27 BEVEZETÉS kapotak Yoshida és munkatársai is. (Yoshida és mtsai 1995). Mindezen eredmények az FDC-k nem csontvelői eredetét látszanak bizonyítani. 4. Feltételezett FDC-prekurzorok a vérben és csontvelőben Parwaresch és munkacsoportja humán FDC-specifikus ellenanyagot (KiM4) használva KiM4 + sejteket találtak a vérben (Parwaresch és mtsai 1983). Hasonló eredményekkel zárultak azok a kísérletek, amelyek az egér vérben keresték az FDCprekurzorokat (Haley és mtsai 1995). Itt az egér FDC-kre specifikus ellenanyagot használtak (FDC-M1). Ezek a sejtek erősen pozitívak voltak a komplement receptorra nézve, akárcsak a fentebb említett antigén transzportáló sejtek és FDC-k. Hasonló sejteket nemcsak a vérben, hanem csontvelőben is azonosítottak, azonban ezen eredményeket elektronmikroszkópos vizsgálatokkal nem tudták igazolni. 5. Adatok, amelyek az FDC-k csontvelő eredetét erősítik Az FDC-k hosszú életű sejtek és nagyon erős (18,5 Gy) besugárzásnak is ellenállnak (Nettesheim és Hanna 1969). Ezt figyelembe véve nem csoda, hogy csontvelő átültetés után recipiens FDC-k vannak jelen. Hogy elkerüljék az FDC-k besugárzás elleni rezisztenciájából fakadó nehézségeket, egyes kutatók SCID egérmodellt használtak. Ezekben az egerekben specifikus ellenanyaggal nem lehet FDC-ket azonosítani, habár kimutatták, hogy ha ezeket az egereket B- vagy T-sejtekkel népesítik be, akkor az FDC-hálózat nagyon gyorsan létrejön. Mindez azt sugallja, hogy a SCID egerekben az FDC-prekurzorok jelen vannak, de a differenciálódásukhoz szükséges jelek/sejtek hiányoznak (Kapasi és mtsai 1993). Egereken és patkányokon végzett kísérletekből származó adatok szerint éretlen FDC-k a születést követő 3. napon megjelennek (FDC-M1 + ), amelyek fokozatosan tovább érnek és érett fenotípusukat a születést követően a második-harmadik hét körül érik el (Holmes és mtsai 1984, Dijkstra és mtsai 1984, Balogh és mtsai 2001). Ezen eredményeket figyelembe véve, SCID újszülött egerekbe más egerek (specifikus ellenanyagokkal megkülönböztethető) csontvelejét vitték át. Feltételezték, hogy a fejlődés során a két különböző eredetű FDCprekurzorok versenyezni fognak egymással. Az elvárásoknak megfelelően a vizsgált 27
28 BEVEZETÉS limfoid szövetek csíracentrumaiban donor eredetű FDC-ket is találtak (Kapasi és mtsai 1994, 1998). Egy másik, még meggyőzőbb kísérletsorozatban a SCID egerekbe -ezek az egerek xenograftokat is elfogadnak- patkány csontvelőt ültettek át. 6-8 héttel a rekonstituciót követően, patkány FDC-specifikus ellenanyaggal donor eredetű FDC-ket azonosítottak a limfoid szövetekben (nyirokcsomó és lép) (Kapasi és mtsai 1998). Az FDC-k csontvelői eredete mellett szólnak a SCID és Rosa-26 egérkimérákkal végzett kísérletek is. A Rosa-26 egerek valamennyi sejtje β-galaktozidázt expresszál. Az enzimaktivitás fluoreszcens szubsztrátummal könnyen kimutatható. Ezen egér csontvelő sejtjeit újszülött SCID egerekbe vitték át, majd a nyirokszervekből izolált sejteket FACS-szal és morfológiai módszerekkel vizsgálták. Az eredmények itt is a donoreredetű FDC-k jelenlétét igazolták (~21 %, nyirokcsomó és lép) (Kapasi és mtsai 1998). Az FDC-k csontvelői eredetét közvetett módon, azok az eredmények is alátámasztják, amelyek azt mutatják, hogy a csíracentrumokba az antigént ún. antigén transzportáló sejtek (ATC), ill. makrofágszerű DC-k szállítják. Erre mai tudásunk szerint csak a csontvelői eredettel rendelkező, az immunrendszerhez tartozó sejtek képesek. Az első FDC-k extra-follikuláris eredetét támogató eredmények White-tól származnak. Munkássága arra a periódusra tehető, amikor még őskorát élte a DCkutatás. Nem meglepő, hogy abban az időben White makrofágszerű dendritikus sejtekről beszél. Ő figyelte meg először, hogy az intravénásan beadott antigént a lépben az ellipszoid körüli sejtek felveszik, majd ezen sejtek elvándorolnak a felvétel színhelyéről, és ezek után csíracentrum-képződésben játszanak szerepet. Ez az előzetes adat abból a korból származik, amikor még nagyon keveset tudtak a DC-k funkciójáról, és az FDC-k még névleg sem léteztek. Így nem csoda, hogy felejtésbe merültek White madarakon végzett kísérleteiből származó adatai (White és mtsai 1969, 1975). Olyan emlős adatok, amelyek alátámasztják White eredményeinek helyességét csak a 80-as évek elején születtek. Ekkorra Szakál és munkatársai azonosítják az ATCket, amelyek az intradermálisan beadott IC-ket veszik fel, majd az afferens nyirokereken keresztül a nyirokcsomókba vándorolnak. Ezek a sejtek (molekuláris mintázatuk megegyezik az FDC-kével) végül bevándorolnak a nyiroktüszőkbe. Csak elméletek szintjén magyarázható, hogy mi történik a bevándorolt sejtekkel és a szállított 28
29 BEVEZETÉS antigénnel. A szerzők valószínűsítik, hogy FDC-kké érnek, ugyanakkor azt sem zárják ki, hogy az antigént átadják a már meglevő FDC-knek (Szakal és mtsai 1983). 6. Az FDC-k differenciálódását befolyásoló citokinek Az LT/TNF családba tartozó citokinek (nevezetesen: tumor nekrózis faktor: TNF- α, limfotoxin: LT α és β) és receptoraikról a 90-es évek elején derült ki, hogy fontos szerepet játszanak a csíracentrumképződésben és az FDC-k érésében, illetve vándorlásában. Kimutatták, hogy a TNF-α, LTα, LTβ citokinek, ill.tnfr-1 vagy LTβR receptorok hiányában a szekunder nyirokszervek rendellenesen fejlődnek és működnek (Matsumoto és mtsai 1997, Ettinger és mtsai 1998). A TNF -/-, de főleg a LTα -/- egérben a lép mikro-architektúrája nagymértékben megváltozik (Fu és mtsai 1997). A megváltozott kötőszöveti váz feltehetőleg nem tudja ellátni feladatát. Ezekben az egerekben nincs csíracentrumképződés és a humorális immunválasz is hibát szenved (pl. nincs izotípusváltás stb.). Ezen knock-out egerek normál egerek csontvelejének transzplantációja után képesek lesznek normális immunválaszt produkálni (Alexopoulou és mtsai 1998). Ezzel ellentétben a TNFR1 -/- egér lépének kötőszöveti váza teljesen ép a GC-k és az FDC-k itt is hiányoznak-, de csontvelő átültetésével nem lehet visszaállítani a GC-képződést. Ezek a kísérletek rámutatnak a nem hemopoetikus elemek által expresszált TNFR1 azon fontos szerepére, amelyet a GC-képződésében játszik (Le Hir és mtsai 1996, Tkachuk és mtsai 1998). Érdekes megfigyelésként könyvelhető el, hogy a fent említett valamennyi esetben bizonyos kemokinek szintézise is hibát szenved. Így pl. a B-limfocita kemoatraktáns (BLC vagy CXCL 13), a másodlagos nyirokszöveti kemokin (SLC: secondary lymphoid tissue chemokine), valamint az Epstein Barr vírus indukált molekula 1 ligand kemokin (ELC: Epstein Barr virus-induced molecule 1 ligand chemokine) mennyisége lecsökken. A BLC a B- limfociták CXCR5 receptorán keresztül hatva a follikulusokba történő vándorlását segíti elő. Hasonló szereppel bírnak az SLC és ELC kemokinek, de ezek a T-sejtekre és a DCkre hatnak (CCR7-en keresztül). Feltehetőleg az említett citokinek (TNF, LT) a stromális elemeken levő receptoraikon keresztül kemokinek (BLC, ELC, SLC) szintézisét indítják be, amelyek a sejteket (T, B, DC) a megfelelő helyre irányítják (Gunn és mtsai 1998, Ngo és mtsai 1999, Weih és mtsai 2001). Ezt az elméletet látszik 29
30 BEVEZETÉS alátámasztani az a megfigyelés is, miszerint a TNFR1 -/- egérben immunizációt követően FDC markerekre pozitív sejteket találtak a marginális zóna területén, amelyek kemokinek hiányában képtelenek voltak a follikulusokba vándorolni. Ez a megfigyelés viszont a nyirokcsomóra leírt ATC-khez hasonló mechanizmust feltételez (Pasparakis és mtsai 2000). AZ EMLŐS- ÉS MADÁR LÉP JELLEGZETESSÉGEI A madarak immunrendszerének működésével kapcsolatos tudásunk főleg a Bursaval és léppel kapcsolatos kísérleteken nyugszik. Az emlősökkel ellentétben a madarak bőrimmunitásáról vajmi keveset tudunk. Ez nem meglepő, hiszen a madarakban hiányoznak a perifériás nyirokcsomók. Ezek hiányában a kutatók nem nagy figyelmet szenteltek a bőrben zajló immunfolyamatok feltérképezésére, hiszen ha vannak is a bőrben DC-k, azoknak nincs egy jól körülhatárolt vándorlási végállomásuk (nyirokcsomó). Ezen adatok fényében mi sem a nem létező nyirokcsomókban vizsgáltuk az FDC-k eredetét, hanem egy, működésében részletesen tanulmányozott szekunder nyirokszervben, a lépben. A madárlép véreredetű antigének kiszűrésében játszott szerepe régóta bizonyított. Már a 10-es években Kyes bebizonyította a lép kiemelkedő fontosságát a Streptococcus elleni rezisztenciában (Kyes 1916). A 80-as évek során keletkezett adatok tovább hangsúlyozták a lép immunrendszerben betöltött szerepét. Az eredmények könnyebb értelmezését szeretnénk elősegíteni a két csoport lépe közötti hasonlóságok és különbségek kiemelésével. Mind az emlős, mind pedig a madár lép fontos feladatai közé tartozik a véreredetű antigének kiszűrése és hatékony immunválasz indukálása. Az emlős lép jellegzetessége, hogy a fehér- és a vörös pulpa határfelületén egy speciális sejtösszetétellel rendelkező régió alakul ki. Ezt nevezik marginális zónának (MZ), ahol a MZ B-sejtek, -makrofágok, -DC-k helyezkednek el, amelyek kiemelt szerepet játszanak az antigénfelvételben. A MZ a madarak lépéből hiányzik, szerepét az ellipszoidokat körülvevő ún. periarteriolás fehér pulpa (PWP) veszi át, amely összetételében a MZ-hoz hasonlít (Oláh és Glick 1982, Oláh és mtsai 1984, Jeurissen és mtsai 1994). Az ellipszoid felületén helyezkednek el (sejttestükkel a 30
31 BEVEZETÉS PWP-ben) azok az ún. ellipszoid-asszociált sejtek (EAC), amelyeket Oláh és Glick írt le (Oláh és Glick 1982). Ezek a rezidens lépsejtek nyúlványaikkal mélyen az ellipszoid belsejébe hatolnak, és a centrális arteriola bazális membránja közelében végződnek. Jellegzetes küllőszerű elrendeződést mutatnak, és a toluidin-kék festés affinitása miatt sötétebben festődnek, mint a környező sejtek (4. kép). Ezek a nyúlványos sejtek elektronmikroszkópos felvételeken jól megfigyelhető 2-3 specifikus szemcsét tartalmaznak (feltehetőleg primer lizoszómák). Leírásukat követően funkciójukra nézve is egyre több adat került publikálásra. Ezek a publikációk rámutattak arra, hogy az intravénásan bejutatott tus/antigének felvételében fontos szerepet játszanak az EAC-k (Oláh és Glick 1982, Eikelenboom és mtsai 1983, Olah és mtsai 1984, del Cacho és mtsai 1995, Gallego és mtsai 1997). Felvételt követően ezek a sejtek leválnak az ellipszoid felszínéről és vándorolni kezdnek a PALS-ban vagy a vörös pulpában. Bizonyos idő elteltével elszórtan tus szemcséket hordozó sejteket a csíracentrumokban is megfigyeltek. Ezen elektronmikroszkópos vizsgálatok vetették fel, annak a lehetőségét, hogy az EAC-k az IDC-k, ill. FDC-k prekurzorai lehetnek. Az EAC-k leírása előtt White és munkatársai figyelték meg, hogy fluoreszcens HSA felvétele az ellipszoidra lokalizálódik (White és mtsai 1969, 1975). Ő makrofágszerű dendritikus sejtekről beszélt -valószínűleg azonosak a később leírt EAC-kkal-, amelyek a felvétel után elvándorolva csíracentrumok képződését indítják el. Hasonló antigénfelvételt és csíracentrumképződést írt le egy másik munkacsoport is (Nagy és mtsai 1973). Ezek az eredmények is az előbb említett hipotézist erősítik. A felsoroltak alapján a madár lép PWP régióját mind morfológiailag, mind pedig funkcionálisan a MZ megfelelőjeként foghatjuk fel. 1. A csirke follikuláris dendritikus sejtek jellegzetességei Az emlős FDC-vel ellentétben vajmi keveset tudunk a csirke FDC-kről. Jelenlétüket Oláh és Glick bizonyították fény- és elektronmikroszkópos módszerekkel (Oláh és Glick 1982). Morfológiai jellegeken kívül, amelyek alapján nagymértékben hasonlítanak az emlős FDC-kre, funkcionális adatok nem állnak rendelkezésünkre. A vizsgálatukhoz szükséges ellenanyagok is hiányoznak. Egyetlen egy ellenanyag létezik, amely többé-kevésbé specifikus a csirke FDC-kre (Jeurissen és mtsai 1992). Nemcsak 31
- és egér(b) lép sémás rajza, valamint az ellipszoid szerkezete. A két lép szerkezeti különbségei közé tartozik a MZ hiánya a madár lépben, aminek feladatát a PWP veszi át.")
32 BEVEZETÉS funkciójuk nem ismert, de sejtes eredetük sem tisztázott. Előzetes, mikroszkópos megfigyelések felvetették annak lehetőségét, hogy a csirke FDC-k előalakjai az EAC-k lehetnek. 4. kép A madár(a)- és egér(b) lép sémás rajza, valamint az ellipszoid szerkezete. A két lép szerkezeti különbségei közé tartozik a MZ hiánya a madár lépben, aminek feladatát a PWP veszi át. Pc: penicilliform kapilláris, CA: centrális arteriola, BC: Billroth-köteg, GC: csíracentrum, Tr: trabekula, S: szinuszoid, SSS/CSS: Schweigger-Seidel hüvely. (c) A madár ellipszoidból készített keresztmetszet félvékony képe. Középen a penicilliform kapillárist körülvevő vastag bazális membrán (BM) figyelhető meg, amelyet támasztó sejtek vesznek körül (SC). A támasztó sejteket kívülről a CSS határolja, míg a sötéten festődő EAC-k (nyilak) küllőszerűen nyúlnak a PWP-ből az ellipszoid belsejébe. (d) Az ellipszoid sémás rajza. EAM: ellipszoid-asszociált makrofág, L: limfocita. 32
33 CÉLKITŰZÉSEK CÉLKITŰZÉSEK Az immunrendszer működésével kapcsolatos tudásunk javarészt rágcsálókon elvégzett kísérletek eredményein alapszik. Az évek során felhalmozódott adatok lehetővé tették, hogy betekintsünk az immunrendszer sejtjei között zajló kommunikációba, kölcsönhatásokba és szabályozásokba. Ezek fényében az immunrendszer működését tekintve többé-kevésbé működőképes modellek láttak napvilágot. Mindezek ellenére fontos hangsúlyozni, hogy a modellállatokra érvényes adatok legtöbb esetben csak fenntartásokkal extrapolálhatók a humán immunrendszerre. Ezért fontos, hogy a kutatók több modellállatra vonatkozó adatokat összesítve hozzanak döntéseket. Egy bonyolult rendszer működésének megértése mindig könnyebb, ha egy hasonló, de egyszerűbb rendszer működését megértettük. Egy ilyen egyszerűbb rendszernek fogható fel a házityúk (Gallus domesticus) immunrendszere. A háziállatok immunrendszerének vizsgálata mellett számos érv szól: gazdasági, állategészségügyi, jelentősége mellett a különböző humán megbetegedések patomechanizmusának modellezésére is alkalmas alanyok lehetnek. Az utóbbi években a DC kutatás soha nem látott méreteket öltött. Ezek a kutatások rengeteg adatot szolgáltattak a DC-k eredetével és funkciójával kapcsolatosan. A DC-k immunrendszerben betöltött szerepének részleges megismerése tette lehetővé egy új kutatási terület kialakulását, éspedig a DC-vakcinák gyógyászatban való használatát. Ahhoz viszont, hogy a DC-ket hatékonyan és biztonságosan lehessen alkalmazni a humán gyógyászatban, alaposabban meg kell, hogy ismerjük ezen sejtek működését. Ehhez próbálunk hozzájárulni a csirke DC-k tanulmányozásával. A csirke DC-alpopulációk eredetét és funkcióját tekintve vajmi keveset tudunk. Munkánk során két, nagyon eltérő DC-alpopuláció részletes karakterizálását vállaltuk fel. Ez a két alpopuláció az LC-k és az FDC-k. Reményeink szerint ezen alpopulációk részletes megismerése az eddigi emlős adatok kiegészítésével hozzájárulhat a DC-k szerepének tisztázásához. 1. Az emlős LC-k a legjobban ismert DC-alpopulációhoz tartoznak. Ezzel ellentétben egyáltalán nem léteznek adatok a csirke LC-k eredetét és funkcióját tekintve. Kísérleteinkkel ezen hiányosságok pótlását kíséreljük meg: 33
34 CÉLKITŰZÉSEK o részletes molekuláris fenotípus meghatározása; o ontogenetikus vizsgálat; o funkcionális vizsgálat. 2. A másik sejtpopuláció, mely vizsgálatát terveztük, még az emlősökben is a legvitatottabb eredettel és funkcióval rendelkező FDC-ké. Korábbi vizsgálatok felvetették annak a lehetőségét, hogy az EAC-k az IDC-k és FDC-k prekurzorai lehetnek. Ezen hipotézisből kiindulva, egy olyan rendszer kidolgozását tűztük ki célul, amely lehetővé tenné ennek vizsgálatát. Ahhoz, hogy mindez vizsgálható legyen, célul tűztük ki egy olyan monoklonális ellenanyag előállítását, amely specifikusan felismeri mind az EAC-kat, mind pedig az FDC-ket. Továbbá szükségünk volt egy olyan antigénre, amelyet szelektíven csak az EAC-k vesznek fel. Ez a rendszer lehetővé tenné az FDC-k valós eredetének és funkciójának megismerését. 34
35 ANYAG ÉS MÓDSZER ANYAG ÉS MÓDSZER 1. Anyagok Amersham Biosciences (Amersham Biosciences Trading Gmbh, Vienna, Austria): 0,45 µm pórusátmérőjű nitrocellulóz membrán, ECL Plus Western Blotting Detection System. Bio-Rad Laboratories (Bio-Rad, Budapest, Magyarorszag): Bis/Akrilamid oldat, SDS, APS, TEMED, 4-15 %-os kész gél, Coomassie 250, Tween 20, 0,45 µm pórusátmérőjű nitrocellulóz membrán. Calbiochem (Merck Kft, Budapest, Magyarország): ProteoExtract, Native Membrane Protein Extraction Kit. Corning-Costar (Corning Incorporated, Corning, NY, USA): egyszerhasználatos, steril műanyag tenyésztőeszközök, ELISA plate-ek. Dako (Dako A/S, Glostrup, Denmark): nyúl anti-humán CD3. Developmental Studies Hybridoma Bank (DSHB, Iowa, IA): hibridómák, felülúszók, ascitesszek. Fermentas (Fermentas international INC, Burlington, Canada): Prestained Protein Ladder. Millipore (Millipore Kft, Budapest, Magyarorszag): tápoldatok sterilizálásához használatos szűrők. Invitrogen (Invitrogen Corporation, Carlsbad, California, USA): sztreptavidin konjugált Alexa 488 ill. -594, valamint Alexa-val jelzett szekunder ellenanyagok, CM- DiI, nyúl anti-beta-galaktozidáz (Molecular Probes), FBS (Gibco), mágneses szemcsékhez (4,5 µm átmérő) kötött anti-egér Ig (Dynal). MTA Izotópkutató Intézet (Budapest, Magyarország): Na 125 I. Orange (Orange Scientific, Braine-l'Alleud, Belgium): egyszer használatos, steril műanyag tenyésztőeszközök. Pierce (Pierce Biotechnology Inc., Rockford, IL, USA) Sulfo-NHS-LC-biotin, IODO- BEAD, Protein-L. Polysciences (Polysciences Inc., Warrington, PA): osmium tetroxid, Polybed/Araldite
36 ANYAG ÉS MÓDSZER Reanal (Budapest, Magyarország): ammónium-szulfid, valamint más finomvegyszerek. Roche (Roche Magyarország, Budaörs, Magyarország): beta-galaktozidáz, X-Gal, proteáz inhibitor koktél tabletta, diszpáz II, DNázI, BrdU. Sigma-Aldrich (Sigma-Aldrich Kft, Budapest, Magyarország): RPMI 1640, DMEM, médiumkiegészítők (HEPES, MEM-vitaminok, MEM-nem esszenciális aminosavak, antibiotikumok, nátrium-piruvát, 2-merkaptoetanol), hibridóma indító kit, 0,25 %-os tripszin oldat, DMSO, DAB, 4-kloro-1-naftol, poli-l-lizin, izotipizáló ellenanyag kit, CnBr-sepharose, Protein G-Sepharose Histopaque-1077, Percoll, Blue-Gal, dextrán (500 kda) ill. dextrán-szulfát, ATP, FITC. Southern Biotechnology (Southern Biotechnology Associates, Inc., Birmingham, AL): izotípus alosztály specifikus ellenanyagok, biotinilált kecske anti-egér IgG1 ill. biotinilált kecske anti-egér IgG2a. Vector Labs (Vector Laboratories, Burlingame, CA): VECTASTAIN Elite ABC kit, NovaRed- és AEC előhívó rendszerek, Avidin-biotin blokkoló kit, biotinilált ló antiegér IgG, biotinilált kecske anti-egér IgM, biotinilált kecske anti-nyúl IgG. 2. Állatok A megtermékenyített SPF White Leghorn tojások a mohácsi BiOvo cégtől, míg a konvencionális New Hampshire tojások és állatok a gödöllői KÁTKI-tól kerültek megrendelésre. A tojásokat 37.7 C-on inkubáltuk. A különböző korú embriókból, ill. állatokból szövetmintákat vettünk. Valamennyi kísérlethez csoportonként legalább négy állatból származó mintát dolgoztunk fel. Ontogenetikus kísérleteink során mindkét fajta egyedeit megvizsgáltuk, míg a többi kísérlethez csak a New Hampshire állatokat használtuk. Az állatkísérletek a Semmelweis Egyetem Etikai Bizottságának jóváhagyásával történtek (Engedélyszám: ). 3. Hámnyúzatok készítése Munkánkhoz általában tollmentes területről származó bőrt használtunk. Ilyen bőrterületet főleg az állat szárnya által fedett, axilláris régióban találtunk. Az innen eltávolított bőrt szikével kisméretű darabokra vágtuk (kb. 0,5 cm2) és percre 2 M 36
37 ANYAG ÉS MÓDSZER CaCl 2 oldatba helyeztük. Ezután sztereómikroszkóp alatt a hámot az alatta levő dermisztől finom csipeszek segítségével választottuk el. Az így nyert hámot Trispufferben (0,1 M, ph 7,2) mostuk, hogy eltávolítsuk a felesleges CaCl 2 -ot, majd a szöveteket 4 C egy órát 4 %-os pufferolt formalinban fixáltuk. Alternatív nyúzatkészítési eljárás alapját szolgálta az a megfigyelésünk, hogy a madárbőr egyes rétegei (hám, ill. dermisz) elég nagy hatékonysággal szétválaszthatók egymástól csak mechanikai eszközöket használva. Ebben az esetben egyszerűen Dumont No. 5 csipeszeket használva lehúztuk az epidermiszt. Megjegyzendő, hogy ebben az esetben a szétválás az alaphártya mentén történt, de az így kapott nyúzatokon nyomokban dermális szennyeződés is jelen volt. Hámnyúzatokat enzimes kezeléssel is készítettünk. Ebben az esetben a kisméretű darabokra vágott bőrt 1 órát 37 C-on 2 mg/ml-es koncentrációjú diszpáz oldatban emésztettük, majd a fentebb említett módszerrel az epidermiszt elválasztottuk a dermisztől (részleteket lásd alább). 4. A Langerhans sejtek kimutatása ATPáz hisztokémiával Az ATPáz festést Robins és Brandon (Robins és Brandon 1981) leírása alapján végeztük el. Röviden, a formalinfixált epidermisz nyúzatokat, valamint az izolált sejteket 0,1 M Tris pufferrel (ph 7,2) mostuk, majd 1 órán keresztül 37 C-on ATPáz médiumban inkubáltuk (0,1 M tris-malát puffer ph 7,2, 10 mm MgSO 4, 6 mm Pb(NO 3 ) 2, 28 mm glükóz és 1,6 mm ATP, dinátrium só). Az inkubálást alapos desztillált vizes mosás követte. Az enzim aktivitásának helyét 1 % (NH 4 ) 2 S-dal tettük láthatóvá. Az előhívás 1 percig tartott szobahőmérsékleten. A megfelelő mosási lépések után az epidermisz nyúzatokat immunofluoreszcens módszerrel festettük meg. 5. Immunohisztokémia Immunohisztokémiai festéseket végeztünk epidermisz nyúzatokon, 10 µm vastag fagyasztott metszeteken, ill. izolált epidermális sejteken (citospin preparátumok). Röviden, PBS mosást követően a szöveteket 45 percig inkubáltuk primer ellenanyagokkal. A biotinilált szekunder ellenanyagokat az endogén peroxidáz blokkolás követte (3 % hidrogén-peroxid PBS-ben, 10 perc). A reakció felerősítéséhez 37
38 ANYAG ÉS MÓDSZER avidin biotinilált peroxidáz komplexet (ABC) használtunk. A primer ellenanyagok kötődési helyeit 4-chloro-1-naphthol vagy NovaRED kit segítségével tettük láthatóvá. Egyszeres immunofluoreszcens jelölés esetén streptavidin konjugált Alexa-488 vagy Alexa-594-et használtunk. Kettős jelölés esetén izotípus specifikus szekunder ellenanyagokat alkalmaztunk (biotinilált anti egér IgG1 vagy IgG2a, biotinilált anti egér IgM, vagy anti nyúl IgG). A két jelölés között avidin-biotin blokkot alkalmaztunk. A magfestés DAPI-val történt. β-gal kezelt állatok szerveiből készített kriosztátos metszeteken először az enzimet mutattuk ki. Ehhez X-gal oldatot használtunk, amelyből az enzimaktivitás során kék csapadék keletkezik. A metszeteket 1 órát inkubáltuk 37 C-on. A metszeteket PBS mosást követően a fentebb említett protokoll szerint immunhisztokémiai, ill. immunfluoreszcens festésekkel is megvizsgáltuk. össze. Munkánk során használatra került ellenanyagokat a 3. Táblázatban foglaltuk 6. Elektronmikroszkópia A szövetmintákat, ill. az izolált epidermális sejteket 0,1 %-os pufferolt glutáraldehidben fixáltuk egy éjszakán keresztül 4 C-on. PBS mosást követően egyes mintákon (pl. nyúzat) CD45 festést végeztünk. Ennek menete megegyezett a fentebb leírtakkal, annyi különbséggel, hogy az inkubációs lépések hosszabbak voltak (4-5 órás inkubációs lépések). A primer ellenanyag kötődési helyét 3,3 -diamino benzidin segítségével tettük láthatóvá. Az immunohisztokémiát követősen a mintákat 2 órát utófixáltuk 1 % ozmium tetroxid-dal. Az etanolos dehidrálást követően a mintákat Polybed/Araldite 6500 keverékbe ágyaztuk be. Az 1 µm vastag félvékony metszeteket toluidin kékkel festettük meg. Elektronmikroszkópiára az ultravékony metszeteket uranil-acetáttal és ólom-citráttal tettük kontrasztossá. A preparátumokat Hitachi H-7600 típusú elektronmikroszkóppal vizsgáltuk. β-gal kezelt állatokból származó lépdarabkákat 4 %-os pufferolt glutáraldehidben fixáltuk egy éjszakán keresztül 4 C-on. Alapos PBS mosást követően az enzimaktivitást X-gal-lal mutattuk ki. A beágyazás további lépései megegyeznek a fentebb leírtakkal. 38
39 ANYAG ÉS MÓDSZER 7. Az epidermális dendritikus sejtek turn-overe Az állatokat egyszeri alkalommal 5-bromo-2-deoxyuridinnel oltottuk be intravénásan (BrdU, 100 mg/kg testsúlykilogramm). Meghatározott időközönként (30 perc, 1, 2 és 4 óra) bőrbiopsziát vettünk nyúzatkészítés céljából (CaCl 2 módszer). Más esetekben a BrdU-t bőrön keresztül alkalmaztuk (0,5 mg/cm 2 ) és hasonló időközönként hámnyúzatokat készítettünk. Harmadik kezelési változat során kezeletlen állatokból mechanikai módszerrel nyert hámnyúzatokat BrdU-t tartalmazó médiumban tenyésztettük (10 µm/ml komplett médiumban; lásd alább). BrdU kezelés után a nyúzatokat pufferolt formalinban fixáltuk, majd PBS mosást követően 2 N HCl-dal denaturáltuk a szövetek DNS tartalmát (30 perc 37 C-on). A HCl neutralizációját követően kétszeres immunofluoreszcens festést végeztünk a nyúzatokon, hogy láthatóvá tegyük a BrdU beépülési helyeit. 8. A epidermális dendritikus sejtek izolálása és tenyésztése Az izolált bőrt vékony, kb. 0,2 x 2 cm csíkokra vágtuk és komplett RPMI médiumban inkubáltuk (10 % FBS, 2 mm L-glutamin, MEM nem-esszenciális aminosav oldat, MEM vitaminok, 100 IU/ml Penicillin, 100 µg/ml streptomicin, 10 mm Hepes, 1 mm nátrium-piruvát, 0,05 mm 2-ME) 1 órát 37 C-on. A médium 2 mg/ml-es koncentrációban diszpázt tartalmazott (grade II). A sejtek összecsapzódását DNázI alkalmazásával védtük ki (0,2 mg/ml). Inkubációt követően az epidermiszt elválasztottuk a dermisztől, majd a PBS-es mosást követően a nyúzatokat 37 C-on további 30 perces tripszines emésztésnek vetettük alá (0,25 %-os in PBS-EDTA). Az így kapott sejteket -mosás után- komplett RPMI médiumba vettük fel és rárétegeztük Histopaque 1077-re. Az 1700 g-s 15 perces centrifugálás után a médium és Ficoll határfelületén maradt sejteket begyűjtöttük, majd mosás után további vizsgálatokhoz használtuk őket. Az epidermális dendritikus sejtek más módszerrel is kinyerhetőek voltak. A bőr alapos alkoholos megtisztítása után, mechanikailag nyúzatokat készítettünk (3 cm 2 ), 39
40 ANYAG ÉS MÓDSZER amelyeket órán át komplett RPMI médiumban tenyésztettünk (37 C, 5% CO 2 ). Az in vitro tenyésztés után a nyúzatokat eltávolítottuk, és a kivándorolt sejteket tanulmányoztuk. 9. A Langerhans sejtek in vivo és in vitro aktivációja Különböző hapténekkel kezeltük a nyolchetes állatok tollmentes bőrfelületét (1 % FITC vagy CM-DiI amelyeket aceton:dibutil-ftalátban oldottunk fel (1:1 v/v). A kezelést követően 1, 2, 4, 12, 24 óra vagy 1 hét után bőrbiopsziákat vettünk a kezelt és kezeletlen területekről. Az epidermiszt a dermisztől CaCl 2 módszerrel választottuk el, majd immunohisztokémiai módszerekkel vizsgáltuk. Az adatokat (3 állat/csoport) páros t-teszttel hasonlítottuk össze (SPSS 13.0 verzió). In vitro kísérleteink során a bőrt hapténnel kezeltük (50 µl/3 cm 2 1 % steril FITC oldat, 30 perc). Alkoholos fertőtlenítés után mechanikailag nyúzatokat készítettünk, amelyeket a fentebb leírtak alapján tenyésztettünk. 10. β-galaktozidázzal történő immunizálás menete Az állatokat intravénásan immunizáltuk 100 U (~0,33 mg fehérje) β-gal oldattal. Az injektálást követően a kezelt állatok lépét meghatározott időközönként eltávolítottuk (45 perc, 2, 24 óra és 4 nap). A lépen kívül más limfoid- és nem limfoid szerveket/szöveteket is megvizsgáltunk (tímusz, bursa, coekális mandula, csontvelő, vér, tüdő, máj, bőr és vese). Ezeket a szerveket májszövetbe ágyaztuk és folyékony nitrogénbe fagyasztottuk le. Az így kapott blokkokat a feldolgozásig -80 C-on tároltuk. Az ellipszoid-asszociált sejtek vándorlási kinetikáját, ill. útvonalát ráoltás során is nyomonkövettük. Ezekben az esetekben a primer immunizált állatok a 14. napon kapták a második oltást. Az így kezelt állatokat 45 perc, ill. 24 óra múlva dolgoztuk fel, a fentebb leírtakhoz hasonlóan. 11. β-galaktozidáz elleni immunválasz kvalitatív vizsgálata 40
41 ANYAG ÉS MÓDSZER Kontroll, ill. kezelt állatokból a kezelést megelőzően, majd ezt követően meghatározott időközönként (4, 12 nap) vért vettünk. Az alvadást követően a szérumot ELISA-val vizsgáltuk. Costar 96 lyukú ELISA lemezeket Voller-pufferben (0,05 M, ph 9,6) hígított β-gal (10 µg/ml) oldattal fedtük be. A szilárd felület, nem telítődött, szabad aktív helyeit tween-es kezeléssel inaktiváltuk. A blokkoló mosásokat követően az állatok szérummintáival (feles hígításban, 50 µl/lyuk) 1 órát inkubáltuk 37 C-on a bevont lyukakat. Mosás után a lyukakat anti-csirke IgG, ill. anti-csirke IgM-el fedtük be (1 µg/ml), majd az egész lemezt peroxidázzal jelzett anti-egér IgG-vel inkubáltuk. Az előhívást ELISA-pufferben oldott OPD-vel végeztük, míg a leállítást 4 N-os H 2 SO 4 -val. A leolvasás ELISA-readerrel történt 492 nm-en. 12. A β-galaktozidáz in vitro felvételének tanulmányozása Kezeletlen állatok lépéből 0,5-1 mm vastag metszeteket készítettünk steril pengével. Az így nyert metszeteket steril PBS-ben alaposan átmostuk, majd termosztátban inkubáltuk különböző összetételű tenyésztőmédiumban. A kontroll metszeteket 1 ml PBS-ben vagy alapmédiumban (10 % FBS-t tartalmazó RPMI) inkubáltuk. Egyes estekben az alapmédiumot 10 % friss autológ vagy hőinaktivált szérummal, ill. heparin-kezelt plazmával egészítettük ki. Valamennyi tenyészethez β- gal-t adtunk (60 µm végkoncetráció) és a tenyészeteket 45 percig CO 2 inkubátorban tartottuk 37 C-on. Kompetíciós vizsgálataink során a metszeteket β-gal-t tartalmazó alapmédiumba helyeztük és heparinnal-, dextránnal- vagy dextrán-szulfáttal (500 kd) egészítettünk ki. A heparint (Ratiopharm, Hungária Kft., Budapest, Hungary) 5 különböző koncentrációban alkalmaztuk (500; 250; 125; 62,5; 31,25 IU/ml), míg a dextrán, ill. dextrán-szulfátot 10 és 20 µm töménységben. A megfelelő inkubálás után a metszeteket lefixáltuk és X-gal előhívó folyadékkal kezeltük. Az elhalt sejtek β-gal kötő képességét aceton fixált, fagyasztott lépmetszeteken vizsgáltuk. A metszeteket 45 percig β-gal-t tartalmazó alapmédiummal fedtük be, majd X-gal-lal hívtuk elő. 13. Az ellipszoid-asszociált sejtek izolálása 41
42 ANYAG ÉS MÓDSZER A lépet sterilezett eszközökkel távolítottuk el, majd PBS-be helyeztük. A lép tokját mechanikusan, csipeszek segítségével eltávolítottuk. Ezt követően egy steril 50 ml-es cső belső falán a lépet alaposan összedaraboltuk és 2 mg/ml-es koncentrációban diszpázt tartalmazó komplett DMEM-et adtunk hozzá (egy 4-6 hetes állat lépéhez 15 ml proteáz oldat kell). Gyakori keverés közepette a csövet 37 ºC-on vízfürdőben inkubáltuk, legalább egy órát. Az inkubálási idő felénél a lépszövet disszociációját erőteljes pipettázással segítettük elő. Ezután 0,2 mg/ml DNáz I-et adtunk a sejtszuszpenzióhoz, hogy megelőzzük a sejtek összecsapzódását. Az emésztést követően a sejtszuszpenziót különválasztottuk a törmeléktől. Ezt követően a sejteket kétszer mostuk 0,1 %-ban BSA-t és 1 mm EDTA-t tartalmazó PBS-sel. A sejteket fokozatosan 4 ºC-ra hűtöttük és Histopaque 1077-re rétegeztük g-n 4 ºC-on 15 percet centrifugáltuk. Ezzel elválasztottuk a vörös- és fehérvérsejteket. 2-3x10 7 fehérvérsejtet tettünk ki 10 cm átmérőjű Petri csészébe. A sejteket 15 percet CO 2 inkubátorba helyeztük. Ez a periódus elég volt ahhoz, hogy a letapadásra képes sejtek (köztük az EAC-k) az aljzathoz rögzüljenek. A le nem tapadt sejteket óvatos mosással eltávolítottuk, míg a kitapadt sejteket erőteljes pipettázással ill. EDTA-s kezelés után gyűjtöttük össze. Az EAC-k feldúsítására más módszert is alkalmaztunk. A vörös- és fehérvérsejtek szétválasztása, valamint mosása után a sejteket 35 %-os BSA oldatba vettük fel, majd DMEM-et rétegeztünk a BSA oldat fölé. A centrifugálást 9500 g-n 15 percen keresztül 4 ºC-on végeztük. A két réteg határfelületén összegyűlt sejtek kerültek további felhasználásra. 14. Az ellipszoid-asszociált sejtek antigénfelvételének tanulmányozása A fentebb említett módszerekkel izolált sejteket β-gal-t (60 µm) tartalmazó komplett DMEM-ben tenyésztettük 45 percen keresztül. Alapos mosást követően a sejteket tárgylemezre centrifugáltuk (citospin), majd X-gal-lal előhívtuk. Az EAC-kat immunhisztokémiai módszerekkel azonosítottuk. 42
43 ANYAG ÉS MÓDSZER 15. β-galaktozidáz gyöngyök előállítása A β-gal-t először 0,1 M-os NaHCO 3 puffer ellenébe dializáltuk, amely 0,5 M NaCl-ot tartalmazott (ph 8,5). Ezt követően a CNBr aktivált Sepharose 4B-t a gyártó által előírt módon kezeltük. A CNBr-Sepharose-t 1 mm-os HCl-ban áztattuk 30 percig, majd a 2 desztillált vizes mosást a dialízishez használt pufferrel való átmosás követte. Az így előkészített gyöngyöket azonnal összekevertük a dializált β-gal-lal (5 mg fehérje/1 ml bead), és miközben folyamatosan kevertettük 2 órát inkubáltuk szobahőmérsékleten. A nem kötődött fehérjéket karbonát pufferrel eltávolítottuk, míg a nem reagált csoportokat 1 M-os etanolaminnal blokkoltuk (ph 8,0) két órán keresztül. A blokkoló puffert PBS mosással távolítottuk el. A gyöngyök β-gal aktivitását X-gal-lal ellenőriztük 16. Radioaktív receptorjelölés és izolálás A feldúsított EAC-k membránfehérjéit IODO-BEADS segítségével radioaktív 125 I-dal jelöltük meg a cég leírása alapján. Ezután a sejtekből a Calbiochem cég Native Membrane Protein Extraction Kit-jét felhasználva követve a cég által közölt protokollt- membránfehérje kivonatot készítettünk. A membránfehérje kivonatot CNBraktivált Sepharose-hoz kötött β-gal gyöngyökkel hoztuk össze (50 µl bead/100 µl kivonat), és két órán keresztül szobahőmérsékleten inkubáltuk (folyamatos forgatás). A gyöngyöket hatszor mostuk PBS-sel (centrifugálás 400 g/15 ), majd redukáló puffer jelenlétében 4 percet főztük. Az így lefőzött fehérjéket SDS-PAGE-n 4-15 %-os gradiens gélen futtattuk meg. Szárítás után a gélt radiográfiás módszerrel előhívtuk (-70 ºC-on 2 napot). 17. Sejtfelszíni jelölés biotinnal és immunprecipitáció A fentebb említett módszerekkel nyert sejtek membránfehérjéit biotinnal is megjelöltük. Sejtfelszíni biotiniláláshoz a sejteket PBS-ben szuszpendáltuk (2,5x10 7 sejt/ml), és megjelölésüket szulfo-nhs-lc-biotinnal (a biotin vízoldékony formája) 43
44 ANYAG ÉS MÓDSZER végeztük. 1 ml szuszpenzióhoz 80 µl szulfo-nhs-lc-biotin törzsoldatot (10 mm) adtunk. Fél óra, 4 C-on történt inkubálás után a sejteket háromszor mostuk hideg PBSsel (300 g, 4 C, 6 perc). A mosást követően az üledéket lizáltuk. A sejtlizishez 1 ml lizispuffert használtunk (50 mm tris, 150 mm NaCl, ph 8,0; proteázgátló koktél). A sejtlízist olvadó jégen végeztük egy órán át. Ezt követően a lizátumot g-n 4 Con 10 percet centrifugáltuk. A tiszta felülúszót azonnal felhasználtuk immunprecipitációhoz vagy -20 C-on tároltuk. Kontrollként bursa sejteket is megjelöltünk. A fentebb leírt biotinilálást vékony lépmetszeteken is elvégeztük. Ebben az esetben mintegy 200 mg súlyú lépszövetet alapos PBS mosást követően- 1 mg/ml-es biotin oldattal inkubáltuk (5 ml). A jelölés után a szöveteket kétféleképpen lizáltuk: egyrészt a sejtes jelölésnél használt hagyományos módszert, másrészt a fentebb említett Calbiochem cég kit-jét alkalmazva nyertünk jelölt membránfrakciót. 18. Elektroforézis és Western-blot A β-galaktozidáz receptorának azonosítása A különböző módszerekkel izolált, feldúsított biotinilált fehérje frakciókat SDSpoliakrilamid-gélelektroforézissel vizsgáltuk meg. Kísérleteinkhez Bio-Rad cég MiniProtean II rendszerét használtuk. Egyes esetekben a futtatásokat gradiens géleken (4-15 %) végeztük, mivel ezek szétválasztási tartománya szélesebb, mint egy adott koncentrációjú poliakrilamid gélé. A nem-gradiens szeparáló gélek összakrilamid koncentrációja 10 % volt. Az immunprecipitációs kísérletek végén a β-gal gyöngyök (~50 µl) utolsó mosása után azokhoz µl redukáló- vagy nem-redukáló mintapuffert adtunk, és a csöveket 4 percig, forrásban levő vízfürdőben inkubáltuk. Ezt követően a bead-eket lecentrifugáltuk, és a tiszta felülúszót vittük fel a gél zsebeibe. A gélre felvitt immunprecipitátumok térfogata ~20 µl volt. Az elektroforézist követően a szétválasztott fehérjéket nitrocellulóz (0,45 µm pórusátmérő) membránra blottoltuk. Zsírmentes tejjel való blokkolást követően a membránokat avidin-biotin-peroxidáz komplex-szel 30 44
45 ANYAG ÉS MÓDSZER percet inkubáltuk. A komplex kötődési helyeit kloro-1-naphtol vagy NovaRed kit segítségével tettük láthatóvá Az E5G12 mea karakterizálása Potter homogenizátor segítségével egy felnőtt állat lépéből a lizátumot készítettünk. A lizispuffer, valamint a fehérjekivonás lépései megegyeztek a 17-es pontban leírtakkal. Az így nyert fehérjekivonatot gélelektroforézissel szétválasztottuk és nitrocellulóz membránra vittük át. Tejes blokkolást követően a membránt csíkokra vágtuk és a következő ellenanyagokkal inkubáltuk: E5G12 és HO-13-4 (indifferens ellenanyag; anti-thy 1.2; IgM). A kontroll membráncsíkot a primer ellenanyag helyett PBS-sel inkubáltuk. A PBS-Tween 20 (0,05 %) mosást követően a membránt biotinnal jelzett anti-egér IgG-vel inkubáltuk. Ezt követően jelerősítéshez avidin-biotin-peroxidáz komplexet alkalmaztunk. Az előhívást kemilumineszcens kit segítségével végeztük. A jeleket Agfa Ortho HT-G filmre rögzítettük. 19. Monoklonális ellenanyagok előállítása A Langerhans sejteknél említett módszerrel epidermális sejtszuszpenziót készítettünk. 8 hetes nőstény Balb/c egereket 2x10 6 sejttel intraperitoneálisan oltottuk be, komplett Freund adjuvánsban oldva. Az oltásokat 14 naponként megismételtük (háromszor, inkomplett Freund adjuváns), majd az utolsó oltást követően a harmadik napon az egereket cervikális díszlokációval megöltük. A monoklonális ellenanyag előállítását Magyar és mtsai által leírt módon végeztük (Magyar és mtsai 1994). A kinőtt klónok felülúszóit immunhisztokémiai módszerekkel, fagyasztott metszeteken teszteltük. Az érdekes festést adó klónokat többször újraklónoztuk és meghatároztuk izotípusokat. 20. Monoklonális ellenanyagok izotipizálása 45
46 ANYAG ÉS MÓDSZER A klónok által termelt ellenanyagok izotípusát ELISA-val határoztuk meg. Ehhez izotipizáló kitet használtunk. 3. Táblázat A munkánk során használt ellenanyagok jellegzetességei Felismert sejttípus (ellenanyag specificitása) Hámsejtek (α-humán pán citokeratin, keresztreagál) Csirke hámsejtek (csirke E-kadherin) Hemopoetikus sejtek (csirke CD45) pan T sejt (humán CD3, keresztreagál) T h sejt (csirke CD4) α/β T cell (csirke α/β (Vβ1) TCR) α/β T cell (csirke α/β (Vβ2) TCR) T c sejt (csirke CD8) γδ T sejt (csirke γδ TCR) B sejt és makrofág alpopuláció (csirke Bu-1b) Ellenanyagok Lu-5 (egér IgG1) 7D6 (egér IgG1) HIS-C7 (egér IgG2a) Poliklonális Anti-humán pan T sejt (nyúl IgG) CT-4 (egér IgG1) TCR-2 TCR-3 CT-8 (egér IgG1) TCR-1-FITC (egér IgG1) 5-11G2 (egér IgG1) Beszerzési hely BMA, Biomedicals AG Augst, Switzerland DSHB, Iowa, IA dr. H. Kurz szívességéből Albert-Ludwigs Univ. Freiburg, Németország Dako A/S, Glostrup, Denmark Southern Biotechnology Associates, Birmingham, AL dr. Thomas Göbel szívességéből, Ludwig Maximilian University, München dr. Thomas Göbel szívességéből, Ludwig Maximilian University, München Southern Biotechnology Southern Biotechnology Southern Biotechnology Valamennyi csirke dendritikus-, ill. ellipszoid-asszociált sejt Különböző sejttípusok (vimentin) Különböző sejttípusok (vimentin) E5G12 (egér IgM, κ) AMF17b (egér IgG1) VIM 3B4 (egér IgG2a) Laboratóriumunkban előállított DSHB, Iowa, IA Progen, Biotechnik GmbH, Heildelberg 46
47 ANYAG ÉS MÓDSZER Csirke IgG Csirke IgM Follikuláris dendritikus sejtek Csirke retikuláris ellipszoid sejtek (EAC) EAC és plazmasejtek Csirke makrofágok Csirke pán-mononukleáris fagocita specifikus Csirke mononukleáris fagocita B sejt, aktiválódott T sejt és dendritikus sejtek (csirke MHC II) Valamennyi magvas sejt (csirke MHCI) Csirke granulocita Csirke granulocita Lizoszómális membrán (csirke LEP 100) Különböző sejttípusok (csirke integrin α5 alegység) Különböző sejttípusok (csirke integrin α6 alegység) Különböző sejttípusok (csirke integrin α7 alegység) Különböző sejttípusok (csirke integrin β1 alegység) CG-106 (egér IgG1) M-1 (egér IgG2b) CVI-ChNL-74.3 (egér IgG1) CVI-ChNL-68.2 (egér IgG1) EIVE12 (egér IgG1) CVI-ChNL-74.2 (egér IgG1) CVI-ChNL-68.1 (egér IgG1) Kul01 (egér IgG1) 2D5 (egér IgG2b) C6B12 (egér IgG1) GRL1 (egér IgG3) GRL2 (egér IgG1) LEP 100 IgG (egér IgG1) D71E2 (egér IgG) P2C62C4 (egér IgG1) H1B4 (egér, ismeretlen izotípus) CSAT (egér IgG2b) Sigma-Aldrich Kft, Magyarország Southern Biotechnology CEDI-Diagnostics, Lelystad, Netherlands CEDI-Diagnostics Dr. Todd Pharr szívességéből, Mississippi, USA (Pharr és mtsai 1995) CEDI-Diagnostics CEDI-Diagnostics Dr. B.M. Goddeeris szívességéből, KU, Leuven, Belgium (Mast és mtsai 1998) DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA DSHB, Iowa, IA Dr. Kiss Anna Anti-kaveolin-1 (human poliklonális szívességéből, SE, kaveolin-1, keresztreagál) nyúl IgG Budapest, Magyarország Csirke melanocita MelEM DSHB, Iowa, IA 47
48 ANYAG ÉS MÓDSZER kollagén III Neuron Anti-β-galaktozidáz (Bakteriális β-galaktozidáz) 3B2 (egér IgG1) 8D9 (egér IgG1) nyúl IgG frakció DSHB, Iowa, USA DSHB, Iowa, USA Molecular Probes, Leiden, Netherlands 48

49 EREDMÉNYEK EREDMÉNYEK I. 1. Az epidermális dendritikus sejtek fenotípusa A bőrhám DC-jeinek a vizsgálatát először fiatal felnőtt (8 hetes) állatokon végeztük. A hám DC-jeinek a kimutatására az irodalomban leírt ATPáz reakciót 5. kép. Egy 8 hetes csirke epidermisz ATPáz festéssel. Bar: 30 µm. használtuk. Ezzel a módszerrel mintegy 2000 sejt jelölődött mm 2 -enként (5. kép). Valamennyi megvizsgált bőrterület hasonló sejtsűrűséget mutatott. További vizsgálatainkhoz a hónalji régióból származó, tollmentes hámot használtuk. Ezt követően kettős jelölési technikát (ATPáz + immunfluoreszcens) állítottunk be az ATPáz + sejtek fenotípusának vizsgálatára. Kísérleteink során két olyan ellenanyagot találtunk, amelyek hasonló festési mintázatot adott, mint az előbb említett ATPáz festés: a CD45 (6. kép: D)-, ill. a vimentin specifikus ellenanyagok. A festési mintázatok összehasonlítása érdekében az ATPáz reakciót követően a nyúzatokat CD45-, ill. vimentin specifikus ellenanyagokkal jelöltük. A két festés reakciói teljesen átfedtek, vagyis az ATPáz + sejtek CD45 + -ak és vimentin intermedier filamentumot tartalmaznak (6. kép: A, B). A vimentin/cd45 kettős immunfluoreszcens festés szintén teljesen átfedett, azaz nem találtunk egyszeres pozitív sejteket a keratinociták között (6. kép: C). Az ATPáz + sejtek részletes fenotípizálására a továbbiakban ezen ellenanyagokat használtuk. Ezek segítségével megállapítottuk, hogy az ATPáz/CD45/vimentin + sejteken kívül gyakorlatilag más (fenotípusú) nyúlványos sejt nem található az epidermiszben. Elenyésző mennyiségben (2-3 sejt/mm 2 ) találtunk melanocitákat is (nincs kép), de ezek CD45 festéssel kizárhatók voltak a DC-k közül. Amint azt a 6. I. képen láthatjuk, az epidermális DC-k (EDC) nem expresszálnak keratint, és a környező keratinociták közé ékelődve helyezkednek el. A csirke EDC-k mintegy %-a hordoz MHCII molekulát. Az MHCII expresszió nem volt 49
50 EREDMÉNYEK egyenletes, míg a csoportokba tömörülő EDC-k erősebben festődtek, addigra az elszórt sejtek kevésbé jelölődtek (6. kép: E, ill. betét 1,2). Az EDC-k %-a KUL01 (6. kép: F), ill monocita markereket hordoznak. Fontos kiemelni, hogy e két utóbbi ellenanyag csak a mechanikailag készített nyúzatokon működtek. A madár EDC-k E- kadherin expressziója nem annyira nyilvánvaló, mint az emlős LC-ké, itt ugyanis a CD45, ill. E-kadherin kettősen festett nyúzatok konfokális mikroszkópos vizsgálata azt mutatja, hogy az EDC-k csak elszórt foltok szintjén expresszálnak E-kadherint (6. kép: G). Az izolált epidermális sejteken végzett kettős festések szerint az EDC-k E-kadherinnegatívak, ellentétben a keratinocitákkal (6. kép: G betét). A madár EDC-k a vizsgált integrinek közül csak a β1-izoformát hordozzák (nincs kép). Az antigének felvételében szerepet játszó kaveoszóma alkotó fehérjéje, a kaveolin-1 nem volt detektálható az EDC-ken, ellentétben az erősen expresszáló hámsejtekkel (6. kép: J). Az EDC-kben ezzel ellentétben majdnem azonos mértékben, mint a keratinocitákban, Lep100 lizoszómális membránfehérjét mutattunk ki (6. kép: K). Folytatva a fenotípizálást, egy olyan DC alpopulációt azonosítottunk (5-10 %), amely CD3 pozitivitást mutatott. Konfokális mikroszkóppal megvizsgálva azt találtuk, hogy a CD3 pozitivitás a citoplazmába lokalizálódik (6. kép: H). Ezek a sejtek nem expresszáltak semmilyen TCR-t (α/β és γ/δ), sem pedig CD4 vagy CD8-at. Ugyanakkor MHCII negatívaknak bizonyultak. Stabilan előforduló sejtpopulációról van szó, amely morfológiai szempontból a többi EDC-től nem különbözik. Általában kisebb csoportosulásokat alkotnak, de elszórtan is előfordulnak. Az EDC-k a környező keratinocitáknál erősebben expresszálják az MHCI-et. A keratinociták határait enyhén kirajzoló MHCI pozitivitás mellett megfigyelhetők az erősen pozitív nyúlványos EDC-k (7. kép: B). Az EDC-ket egy jól fejlett, neurális elemekből álló hálózat veszi körül. Az általunk használt ellenanyag (8D9) főleg az axonokat festi, de jelen esetben feltehetőleg a dendriteket tünteti fel (7. kép: A). Nem sikerült immunglobulinokat kimutatni az EDC-k felszínén, de ez nem zárja ki az Fc-receptorok jelenlétét. Aggregált IgG-vel nem próbálkoztunk. A csirke monocitákat/makrofágokat felismerő ellenanyagokkal nem kaptunk festést az epidermiszben (74.2, K1). B-sejtek jelen vannak a dermiszben, de az epidermiszbe nem vándorolnak be. Az EDC-k ugyancsak Bu-1 negatívak. Az FDC-k azonosítására 50
51 EREDMÉNYEK használt marker a 74.3 nem mutat pozitivitást a hámban. A granulociták jelenlétét, pedig a GRL1,2 ellenanyagokkal kizártuk. Összefoglalva elmondhatjuk, hogy a fenotípikus vizsgálat során az ATPáz + /CD45 + /vimentin + DC populáción belül három DC alpopulációt sikerült azonosítanunk (4. táblázat): 1). MHCII + /KUL01 + / CD3 -. 2). MHCII - /KUL01 - / /CD3 -. MHCII - /KUL01 - / /CD Táblázat Csirke EDC-k fenotípusa EPIDERMÁLIS DENDRITIKUS SEJTEK Antigén expresszió I Típus II Típus III Típus ATPáz CD Vimentin MHC II mab KUL mab E-kadherin -/+ -/+ -/+ Integrin β1 alegység Integrin α5, α6, α7 alegységek anti-kaveolin Lep100 -/+ -/+ -/+ CD CD CD γ/δ TCR α/β TCR Bu-1b IgG and IgM Follikuláris dendritikus sejt marker mab monocita/makrofág markerek (mabs 74.2, K1) mab Granulocita markerek mabs GRL1 and GRL Százalékos arány 40-50% 35-45% 5-10% 51
52 EREDMÉNYEK 2. Az epidermális dendritikus sejtek elektronmikroszkópiája Az elektronmikroszkópos immunhisztokémiai vizsgálatok szerint az EDC-k egyrészt a germinális réteg sejtjei között, másrészt azok bazális felszínéhez tapadva fordulnak elő. Az EDC-k nyúlványaik révén a környező keratinociták közé ékelődnek, de sejtkapcsoló struktúrákat nem sikerült rajtuk azonosítani. A szomszédos sejtektől elérően mind a citoplazma, mind pedig a mag sötétebben festődik (kontrasztosabb). A citoplazmájukban tonofilamentum nem található, de intermedier filamentum kötegek gyakran láthatók. Magjuk lebenyezett, kiterjedt heterokromatikus részekkel, amelyek főleg a maghártyához asszociáltan jelentkeznek. Változó számú sejtmagvacska is megfigyelhető a magban. Nem mindegyik EDC-ben találtunk az LC-kre jellemző Birbeck-granulumot (8. kép: A). Az izolált EDC-k egy részében is fellelhetők a Birbeck granulumok (8. kép: B és betét). 3. Az epidermális dendritikus sejtek izolálása és tenyésztése Enzimatikus kezelést követően az epidermális sejtek között 5-8 %-ban találunk CD45 + EDC-t (8. kép: C). Az EDC-ket mintegy 50 %-ra dúsíthatjuk, ha a natív epidermális sejtszuszpenziót Ficoll-on szétválasztjuk (8. kép: D). Az EDC-k a Ficollmédium határfelületen koncentrálódnak. Az izolált sejtek fenotípusa nem változik meg. Valamennyi EDC-típus az in vivo nyert adatokhoz hasonló arányban képviseltette magát. Az így izolált sejtek azonban egy-két napon belül elpusztulnak. Jóval tisztább (~100 %) EDC populációt kapunk a nyúzatok in vitro tenyésztésével. Ebben az esetben nincs keratinocita szennyeződés. A tenyészetek befertőződése nagyon ritkán fordul elő. 1 mm 2 bőrből mintegy sejt vándorol ki 48 óra alatt. A kivándorolt EDC-k csak egy része képes a felszínhez erőteljesen letapadni. Ezek a sejtek számos nyúlványt képeznek és jellegzetes dendritikus morfológiát mutatnak, ami eltér a makrofágoknál vagy fibroblasztoknál tapasztaltaktól. Az EDC-k többi része kevésbé képes letapadni és jellegzetes az LC-kre jellemző fátyolos külalakkal jellemezhetők. Ezek a sejtek számos apró sejtnyúlvánnyal rendelkeznek. A kivándorolt sejtek fenotípusa nem változik az in situ tapasztaltakhoz 52
53 EREDMÉNYEK viszonyítva (8. kép: E, F). CD3 + populációhoz tartozó sejteket azonban nem találtunk a kivándorolt EDC-k között. Az EDC-k emigrációját haptén alkalmazásával gyorsíthatjuk. 4. Az epidermális dendritikus sejtek ontogenetikája Az EDC-k megjelenésének dinamikáját CD45 festéssel követtük nyomon. A New Hampshire (NH) fajtában a kikelést megelőzően (18 napos embrió) találtunk EDCket az epidermiszben (9. kép: A). Ezek száma 2-10 sejt/mm 2, amely fokozatosan emelkedett, és kéthetes korban megközelítette a felnőtt állatokra jellemző sejtsűrűséget (~2000 sejt/mm 2 ) (9. kép: B, C). Ezen fajta egyedeiben az EDC-szám felnőtt állatokban nem mutatott szignifikáns ingadozást. Ezzel ellentétben az EDC-ek a White Leghorn (WL) csirkékben embrionális korban nincsenek jelen (9. kép: D). Megjelenésük a kikelést követő második hétre tehető (9. kép: E). Az EDC-k száma fokozatosan nőtt, de még 4-5 hetes csirkékben is tág határok között ingadozott (9. kép: F). Gyakran találtunk olyan egyedeket, amelyekben a normális sejtszám (~2000 sejt/mm 2 ) helyett sejt volt mm 2 -enként. Intravénás BrdU adással vizsgáltuk az EDC-k proliferációs képességét. 30 perccel az intravénás BrdU adást követően a nyúzatokban a CD45 + sejtek mintegy 4-5 %-a mutatott pozitivitást. Ezen arány a következő 2 vagy 24 órán belül szignifikánsan nem változott (in vivo jelölés; 9. kép: G). Ahhoz, hogy kizárjuk az extraepidermálisan jelölődött, majd bevándorolt prekurzorokat in vitro kísérleteket folytattunk. A BrdU-t tartalmazó médiumban tenyésztett epidermális nyúzatokban hasonló számú EDC jelölődött. A perkután alkalmazott BrdU is felvételre került, itt az EDC-k ~5 %-a osztódott. Amint azt az 5. táblázat is mutatja az in vitro kísérletek az in vivo-hoz hasonló adatokat szolgáltattak. 5. Táblázat Az EDC-k proliferációs képessége BrdU adását követően eltelt idő In vivo BrdU jelölődés In vitro BrdU jelölődés 30 perc 4,86 % 4,42 % 2 óra 5,38 % 5,12 % 24 óra 3,99 % n.a. 53
 keverékében oldott haptént vittünk fel (FITC vagy DiI).")
54 EREDMÉNYEK 5. Az epidermális dendritikus sejtek funkciója Mind in vivo, mind pedig in vitro kísérleteket végeztünk az EDC-k funkcionális jellemzésére. In vivo kísérletekben a hónalji, tollmentes területekre dibutil-ftalát:aceton (1:1) keverékében oldott haptént vittünk fel (FITC vagy DiI). Ezt követően időközönként bőrmintákat vettünk, és CD45 festéssel követtük nyomon az EDC-k számát. A hapténfelvitelt követően 1-2 órán belül az EDC-k egy része a hámban lekerekedett, majd a következő órákban számuk az elvándorlás miatt fokozatosan csökkenni kezdett. Mintegy óra elteltével az EDC-k száma %-kal csökkent ( sejt/mm 2 ) a kontroll területhez képest (10. kép). Az oldószer önmagában nem befolyásolta az EDC-k számát. Az epidermiszben visszamaradt sejtek %-a CD3 + alpopulációba sorolható. A kezelt területen az EDC-k száma mintegy hét alatt 10. kép. Az EDC-k számának változása 24 órával a kezelést követően. 1-, 2-, 3- kezelt csoportok 3-3 egyeddel. normalizálódott. In vitro kísérleteink is azt bizonyítják, hogy az EDC-k képesek kivándorolni az epidermiszből. 30 perccel a FITC felvitelét követően a mechanikusan készített 54
55 EREDMÉNYEK nyúzatokat kitettük tenyészetbe. 24 óra elteltével a kivándorolt sejteket megvizsgálva azt tapasztaltuk, hogy ezek ATPáz +, CD45 +, vimentin +, MHCII + (~100 %), Lep vak. A sejtek dendritikus morfológiát mutattak és mintegy 90 %-a FITC + -nak bizonyult (9. kép: K). Ezekben a tenyészetekben soha nem találtunk kivándorolt CD3 + /FITC + sejtet. A haptén stimulált tenyészetekben nagyfokú EDC kivándorlás volt megfigyelhető ( sejt vándorolt ki 1 mm 2 bőrből 24 óra alatt). A kivándorolt sejtek hasonló morfológiát mutattak, mint amint azt fentebb az EDC izolálásánál láttuk. 6. Az epidermális dendritikus sejtek vándorlási útvonalának feltérképezése Következő lépés volt az EDC-k vándorlási útvonalának a feltérképezése. A hónalji területek FITC-ces kezelését követően 24 óra múlva immunfluoreszcens módszerekkel megvizsgálva a különböző limfoid szerveket/szöveteket (lép, tímusz, cökális mandula, bursa, csontvelő, vér) nem találtunk haptén hordozó sejteket. Ezen eredmények felvetették annak a lehetőségét, hogy a sejtek nem hagyják el a bőrt. Ennek eldöntésére a kezelt bőrterületeket sorozatmetszeteken vizsgáltuk meg. FITC + sejtek elszórtan a dermiszben, valamint az ún. dermális limfoid nodulusokban akkumulálódtak. Ezek a néhány 100 sejtből álló struktúrák az epidermisz közelében a dermiszben foglalnak helyet. Immunhisztokémiai festések azt mutatják, hogy kis kapillárisok és nyirokerek mentén alakulnak ki. A bőr felületével párhuzamosan készített metszetek alapján számuk 1-5 nodulus/cm 2 -re tehető. Ezen struktúrák központi szerkezeti vázát 74.3 pozitív DC-k alkotják, míg széli részüket KUL01 + sejtek határolják. A dendritikus retikulum közötti teret T- és B-sejtek töltik ki (9. kép: H). A T sejtek közül a Th típus van túlsúlyban. IgG pozitivitás csak elszórtan, ritkán figyelhető meg a nodulusban, így a B-limfociták jórészt IgM-et hordoznak (nincs kép). A nodulusok nagyon erős MHCII expressziót- és ekto-atpáz aktivitást mutatnak. A FITC pozitivitás a nodulusokban a KUL01 és 74.3 sejtekre korlátozódott (9. kép: I, J). 55
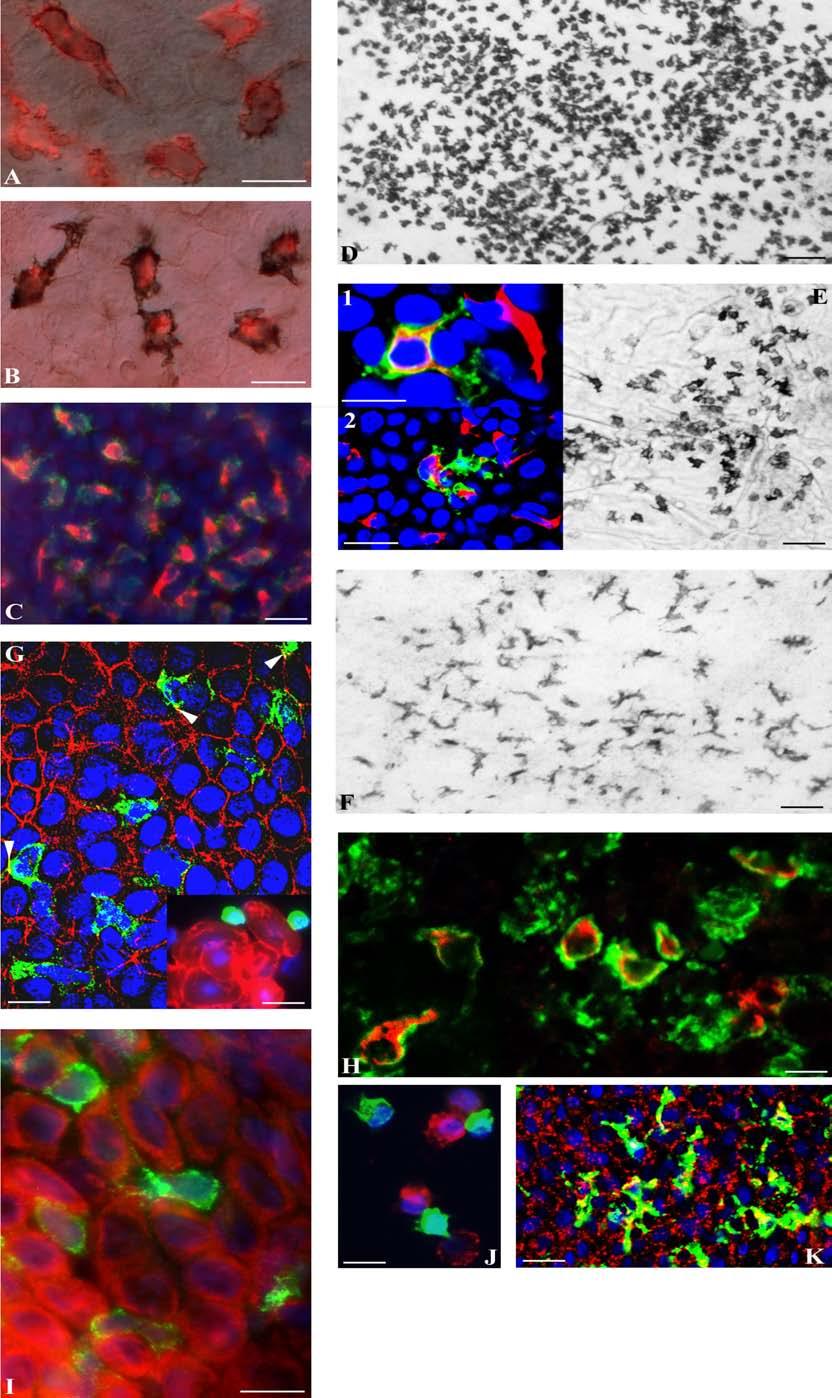
56 56 EREDMÉNYEK
 Az ekto- ATPáz aktivitással (barna) rendelkező EDC-k ugyanakkor vimentint tartalmaznak (piros). Bar: 10 µm. (C) Az újonnan azonosított EDC markerek teljes mértékben átfednek.")
 Az MHCII festés kevesebb sejtet jelölt. Bar: 30 µm. A betét 1, 2 vimentin (piros) MHCII (zöld) kettősfestés konfokális képét mutatja. A magfestés DAPI-val történt. Bar: 10 µm, ill. 20 µm.")
57 EREDMÉNYEK 6. kép. Az epidermális dendritikus sejtek fenotipizálása I. (A) Az ATPáz + EDC-k CD45-öt hordoznak felszínükön. ATPáz hisztokémia (barna) és CD45 immunfluoreszcencia (piros). Bar: 10 µm. (B) Az ekto- ATPáz aktivitással (barna) rendelkező EDC-k ugyanakkor vimentint tartalmaznak (piros). Bar: 10 µm. (C) Az újonnan azonosított EDC markerek teljes mértékben átfednek. CD45 (zöld) vimentin (piros) kettős immunfluoreszcens festés. A magokat DAPI-val tettük láthatóvá. Bar: 10 µm. (D) A CD45 immunhisztokémiai festés az EDC-k szabálytalan eloszlását mutatja. Bar: 30 µm. (E) Az MHCII festés kevesebb sejtet jelölt. Bar: 30 µm. A betét 1, 2 vimentin (piros) MHCII (zöld) kettősfestés konfokális képét mutatja. A magfestés DAPI-val történt. Bar: 10 µm, ill. 20 µm. (F) A monocita/makrofág marker a KUL01, -a D képhez hasonlítva- kevesebb sejtet fest. Bar: 30 µm. (G) A csirke EDC-k (CD45, zöld) majdnem teljesen negatívak E-kadherinre (piros) nézve. E két marker közötti átfedések elszórt sárga foltok (nyílhegyek) képében jelennek meg a konfokális képen. Bar: 10 µm. Betét: Hasonló festés, de enzimatikusan nyert epidermális sejteken. A keratinociták megtartották E-kadherin pozitivitásukat, míg az EDC-k teljesen negatívak voltak. Bar: 10 µm. (H) CD3 (piros) és CD45 (zöld) kettősfestés azt mutatja, hogy a CD3 pozitivitás intracitoplazmatikus. Konfokális kép. Bar: 10 µm. (I) A CD45 + (zöld) EDC-k a citokeratin + (piros) hámsejtek között helyezkednek el. Bar: 10 µm. (J) Az izolált epidermális sejtek közül csak a keratinociták expresszálnak kaveolin-1-et (piros) detektálható szinten. A CD45 + (zöld) sejtek negatívak erre a molekulára. Bar: 10 µm. (K) A CD45 + (zöld) EDC-k közel hasonló mennyiségű Lep100- at (piros) tartalmaznak, mint a keratinociták. A B 7. kép. Az epidermális dendritikus sejtek fenotipizálása II. (A) Az EDC-ket jól fejlett idegrendszeri elemekből álló hálózat veszi körül. 8D9 (szürke) - CD45 (piros) kettős festés. Bar: 10 µm. (B) Az EDC-k erősebben expresszálják az MHCI molekulát, mint a környező keratinociták. Bar: 10 µm. 57
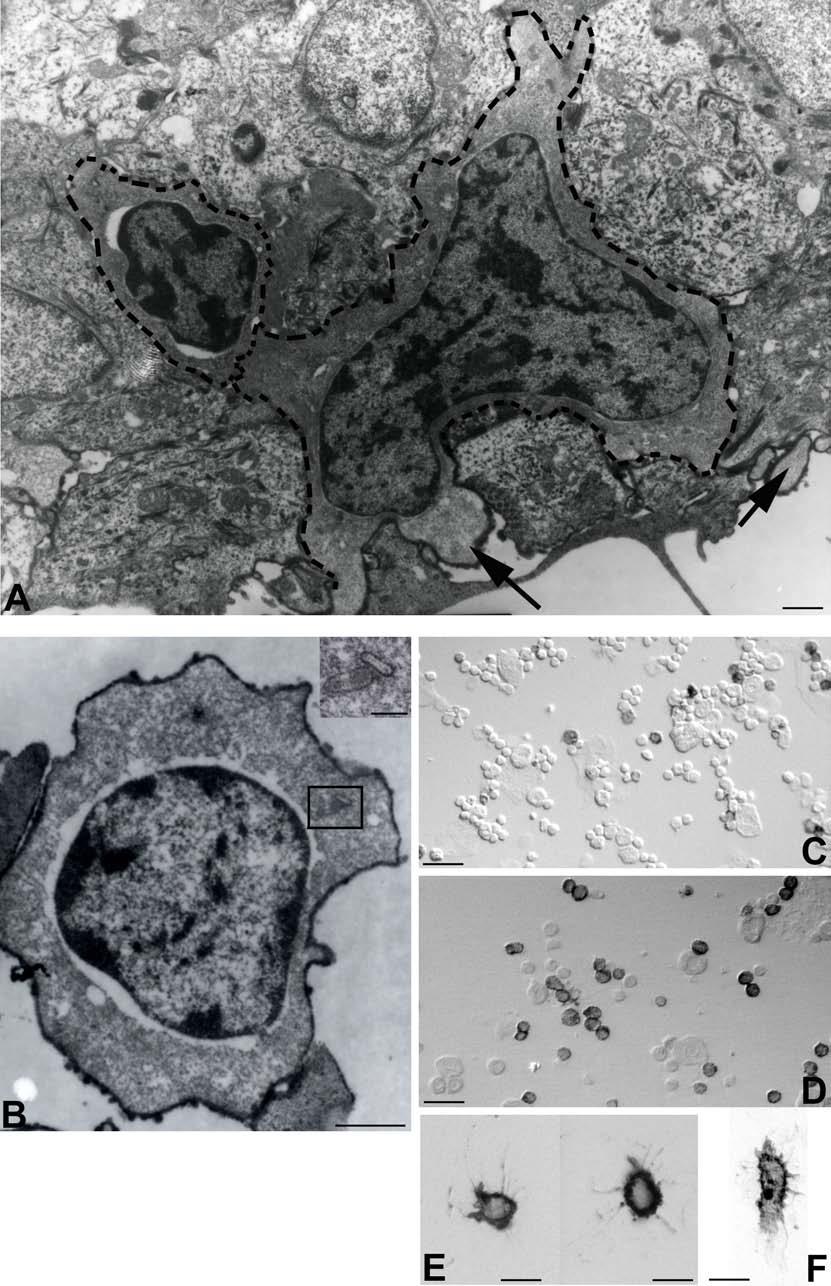
58 58 EREDMÉNYEK
59 EREDMÉNYEK 8. kép. Az epidermális dendritikus sejtek fenotipizálása III. (A) Elektronmikroszkópos vizsgálathoz az EDC-ket CD45-tel festettük meg. Az EDC-k határait szaggatott vonallal jelöltük. A nyilak a CD45 + nyúlványokra mutatnak Bar: 0,5 µm. (B) Ugyancsak CD45 festés, de izolált EDC-ken. A négyzet a Birbeck granulumokat emeli ki. Bar: 0,5 µm. Betét: A Birbeck granulumok kinagyított képe. Bar: 0,1 µm. (C-D) Az EDC-k izolálása diszpázos-tripszines emésztéssel. (C) Natív epidermális sejtszuszpenzióból készített citospin preparátum, megfestve CD45-re. A sejtek mintegy 5-8 % mutatott CD45 pozitivitást. (D) Ficoll-Histopaque-n való szétválasztás után az EDC-k feldúsíthatók mintegy 50 %-ra. (E-F) 24 óra in vitro kezeletlen hámnyúzatokból tenyésztés után kivándorolt dendritikus sejtek. (E) CD45. (F) MHCII. Bar: 8 µm. 9. kép. Az epidermális dendritikus sejtek ontogenetikus- és funkcionális vizsgálata. (A-F) Az EDC-k megjelenésének időbeli dinamikája CD45 festéssel nyomonkövetve. (A) Kikelést megelőzően a NH csirkeembriókban néhány pozitív sejtet találunk mm 2 -enként. (B) A 14 napos állatok bőre hasonló számú EDC-t tartalmaz, mint a felnőtt állaté. (C, F) 8 hetes állatokban az EDC-szám mindkét fajtában 2000 sejt/mm 2 körül stabilizálódik. (D) Embrionális stádiumban a WH csirkék bőrében nem találtunk CD45 + sejteket. (E) Ebben a fajtában az EDC-k megjelenése a kikelést követő második héten veszi kezdetét. Bar: 30 µm. (G) Két órával a BrdU kezelést követően a hámon BrdU (piros) - CD45 (zöld) kettősfestést végeztünk. A nyílhegyek a BrdU + EDC-kre mutatnak. Bar: 30 µm. Betét: egy osztódott EDC kinagyított képe látható. Bar: 10 µm. (H-K) Az EDC-k funkcionális tanulmányozása. (H) Egy nyolchetes csirke bőréből készített fagyasztott metszet. A dermális limfoid nodulusokat, amelyek közvetlenül az epidermisz alatt foglalnak helyet (nyílhegyek), B (Bu-1, szürkés-kék) ill. T-sejtekre (CD3, barna) specifikus ellenanyagokkal jelöltük. Bar: 50 µm. (I) FITC-cel való szenzitivizációt követően a FITC pozitivitás ezen nodulusok perifériás részét övező KUL01 sejtekre-, ill. a centrális részt alkotó sejtekre (J) lokalizálódik. Bar: 20 µm. (K) FITC-cel kezelt nyúzatokból 24 óra in vitro tenyésztés után kivándorolt CD45 + sejtek (piros). Bar: 10 µm. 59
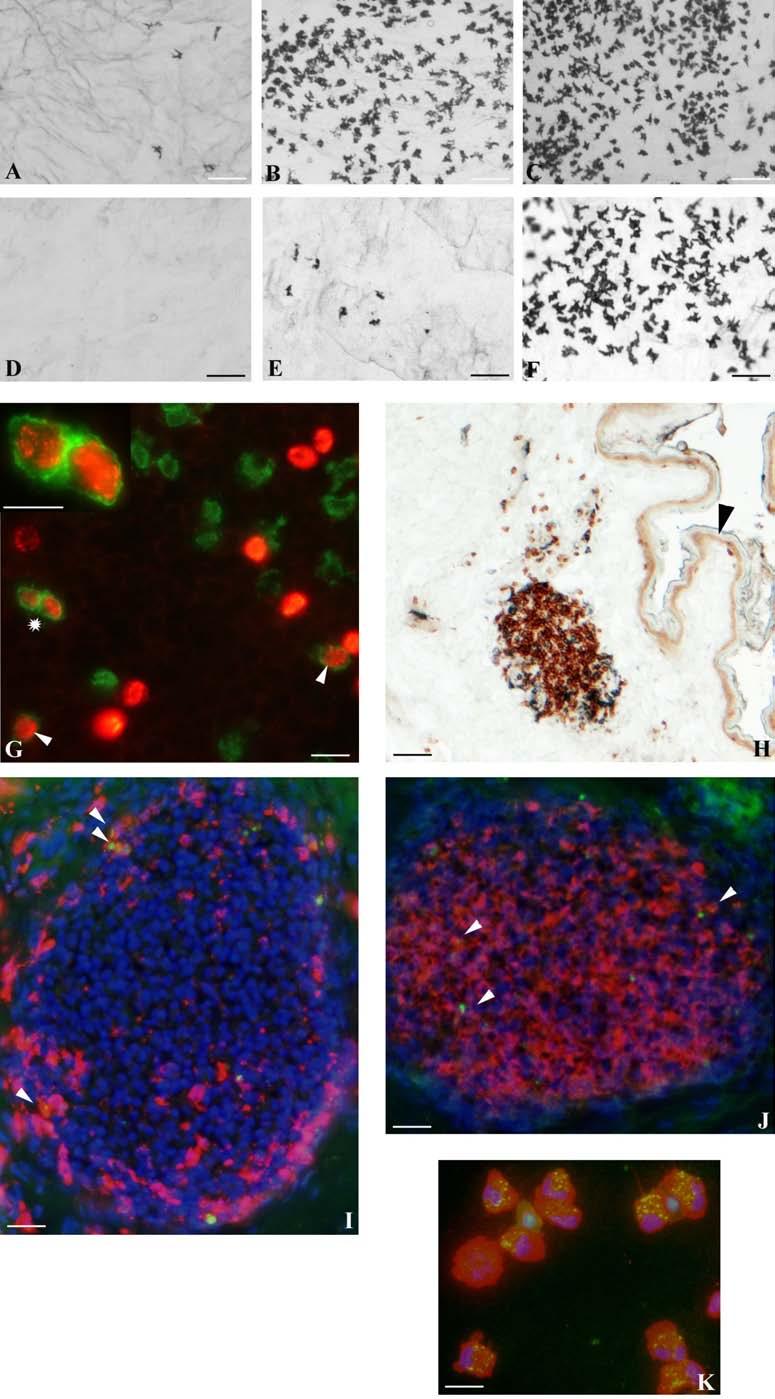
60 60 EREDMÉNYEK
61 EREDMÉNYEK EREDMÉNYEK II. 1. A véletlen a tudomány szolgálatában Az FDC-k differenciálódásának vizsgálata a lépben tulajdonképpen egy véletlennek köszönhető. Az LC-k antigénfelvételének tanulmányozásához egy könnyen kimutatható fehérjét kerestünk. A bakteriális béta-galaktozidáz (β-gal) erre a célra nagyon jól megfelelt, mert detektálását enzimreakciója révén- egyszerűen és gyorsan el lehet végezni. A csirkeszövetek ilyen típusú enzimaktivitással nem rendelkeznek, így gyakorlatilag háttérmentes környezetben egy nagyon érzékeny antigénkimutatási módszer áll a rendelkezésünkre. Habár ez a fehérje az eredeti célra: a LC-ek bőrön keresztüli antigénfelvételének tanulmányozására nem vált be, e kísérletek egyik kontrollja nagyon érdekes eredményt mutatott. A β-gal intravénás injekcióját (45 ) követően a kísérleti állatok (2 hetes kortól 8 hetes korig) gyakorlatilag összes szövetét átvizsgálva, csak a lépben, és itt is nagyon jellegzetes eloszlásban kaptunk enzimreakciót. Ez a melléklelet indította el a vizsgálatainkat, melyek során az EAC-k antigénfelvételét, elvándorlását és differenciálódását tanulmányoztuk. 2. Az E5G12 monoklonális ellenanyag szelekciója és vizsgálata Az E5G12 jelű monoklonális ellenanyagunk (IgM, κ) epidermiszsejtekkel (így LC-kel is) immunizált egérben készült. Az ellenanyag madárlépben jellegzetes lokalizációjú, nyúlványos sejteket fest. Egyrészt az ellipszoid körüli PWP nyúlványos sejtjeit jelöli meg, másrészt dendritikus sejteket fest a lép csíracentrumaiban és a fehér pulpa többi részében is. Kettős immunhisztokémiai festéssel kiderült, hogy az E5G12 monoklonális ellenanyag (mea) festése a PWP területén teljesen kolokalizál az EAC sejtekre specifikus 68.2 mea-éval (11. kép: B), valamint az ezeket a sejteket is felismerő EIVE12 ellenanyagéval. A 68.2 mea a CD45 + EAC-n (11. kép: A) kívül feltehetőleg az IDC-ket is felismeri. Másrészt az E5G12 csíracentrum festése kolokalizált az FDCspecifikus 74.3 mea-éval (nincs kép). Az ellenanyag izotípusa megnehezítette az ellenanyag további karakterizálását. Többféle antigénkivonási kísérlet után végül sikerült az ellenanyag által felismert antigén molekulatömegét meghatározni. Az 61
 Az EAC-k (68.")
62 EREDMÉNYEK előzetes vizsgálatok szerint az E5G12 mea egy ~45 kda-os fehérjét ismer fel. A nemredukált fehérje molekulatömege megegyezik a redukált fehérje tömegével. (nincs kép). Ugyanakkor az izotípus sugallja oligoszacharid oldalláncban jelenlévő epitóp felismerését. A B 11. kép. Az EAC-k jellegzetességei. (A) Az EAC-k (68.2 piros) CD45 (zöld) molekulát expresszálnak, amint az a konfokális képen is látható. Bar. 30 µm. Az új ellenanyag (E5G12, zöld) festési mintázata teljes mértékben átfed a 68.2 antigén expressziójával (piros). A csíracentrum FDC-it csak az E5G12 festi. E: ellipszoid; GC: csíracentrum. Bar: 30 µm 62

63 EREDMÉNYEK kép. Az E5G12 mea karakterizálása. 1. Molekulasúly standard. 2. PBSkontroll. 3. Az E5G12 mea egy ~45 kda tömegű fehérjét ismer fel. 4. Indifferens ellanyag (IgM izotípus). A gél felső részében látható, erősen pozitív csíkokat az endogén biotinilált fehérjék adták. 3. A bakteriális antigént szelektíven az ellipszoid-asszociált sejtek veszik fel Intravénás beadást követően különböző sejttípusokra specifikus ellenanyagokkal ellenőriztük a β-gal pontos lokalizációját a madárlépben. Közvetlenül a beadást követően (15-45 ) a β-gal pozitivitás 99 %-a a periellipszoidális fehérpulpában (PWP) volt kimutatható, pontosan ott, ahol az EAC-k találhatók (13. kép: A, B). A PWP az ellipszoidot körülvevő Schweigger-Seidel tokon (CSS) kívül helyezkedik el. Ezt kollagén III festéssel igazoltuk. A kollagén III-ra specifikus ellenanyag az ellipszoidot kettős gyűrűként tünteti fel, megfestve a kapilláris bazális membránját és a CSS-et (13. kép: C). E két gyűrű között elszórtan előfordult néhány β-gal + sejt is (13. kép: D). Kettősfestéssel igazoltuk, hogy a β-gal hisztokémia során képződött csapadék teljes mértékben a , ill. EIVE12 + sejtekre lokalizálódott (13. kép: E, F). Az előzetes vizsgálatok alapján nem meglepő, hogy az E5G12 mea is kolokalizációt mutatott a β- gal-t felvevő sejtekkel (13. kép: G). Csirke monocita-makrofág markerek, mint a 74.2 vagy a KUL01 a β-gal + sejteket egyáltalán nem festik (13. kép: H, I). Az ellipszoidokat határoló B-sejtes gyűrű nem vesz részt a β-gal felvételben (13. kép: J). A beadást követően a vérből nagymértékű granulocita beáramlás történik az ellipszoid belsejébe, de a β-gal-t ezek a sejtek nem veszik fel (β-gal pozitivitást soha nem találtunk GRL1, -2 63
 45 perccel a beadást követően a β-gal kirajzolja az ellipszoidokat.")
 a β-gal aktivitás pontos lokalizációját a kollagén IIIal (barna) való festés adja meg. A: centrális artéria.")
 A β-gal kolokalizációt mutat a 68.2-vel (F), valamint az EIVE12-vel. (G) Az E5G12 (szürke) is a β-gal-t felvevő sejteket festi.")
 A KUL01 ellenanyag is hasonló festést ad (barna).")
64 EREDMÉNYEK pozitív granulcitákban) (13. kép: L). Ugyanígy, sem az ellipszoid körül jelenlevő ra gyengén pozitív sejtek (13. kép: K). A B C D 13. kép. A β-gal lépen belüli lokalizációja. (A) 45 perccel a beadást követően a β-gal kirajzolja az ellipszoidokat. A β-gal aktivitás valamennyi képen kék színű csapadékként (hisztokémia) jelenik meg. (B) Az előbbi félvékony képe látható. (C) a β-gal aktivitás pontos lokalizációját a kollagén IIIal (barna) való festés adja meg. A: centrális artéria. (D) Nagyobb nagyításon megfigyelhetjük, hogy néhány β-gal + sejt (nyilak) a kollagén kettősgyűrű belsejében is előfordul. (E) A β-gal kolokalizációt mutat a 68.2-vel (F), valamint az EIVE12-vel. (G) Az E5G12 (szürke) is a β-gal-t felvevő sejteket festi. (H) A makrofág specifikus 74.2 ellenanyag (szürke) a β-gal + sejteken kívüli régióban fest. (I) A KUL01 ellenanyag is hasonló festést ad (barna). (J) A β-gal + sejtek B-sejtek között foglalnak helyet, de nem mutatnak kolokalizációt (barna). (K) A sejtek nem vesznek részt az antigénfelvételben (szürke). (L) Az odavándorolt granulociták (barna) sem veszik fel a β-gal-t. 64
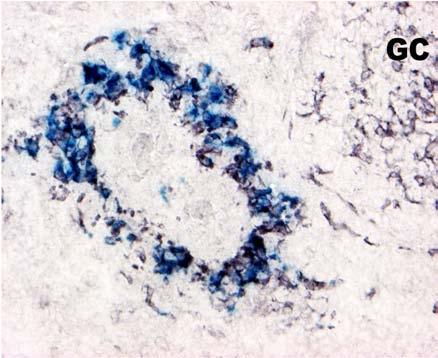
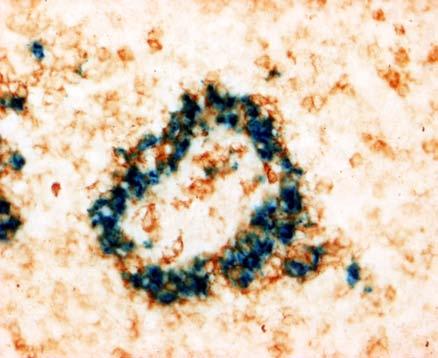
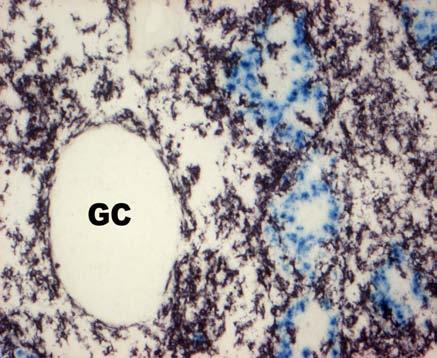
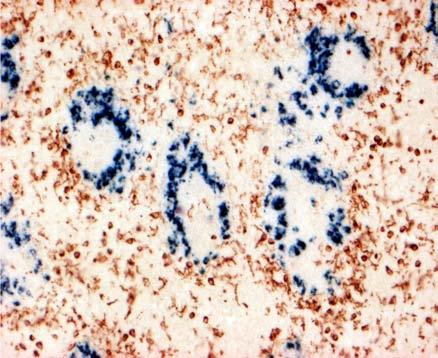
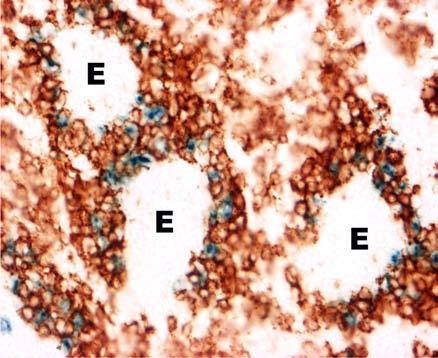
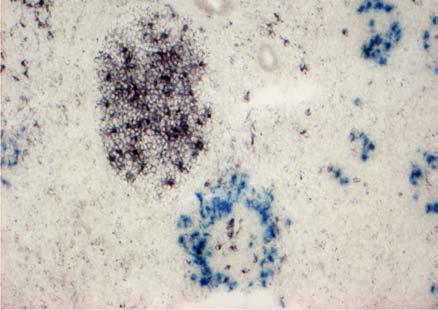
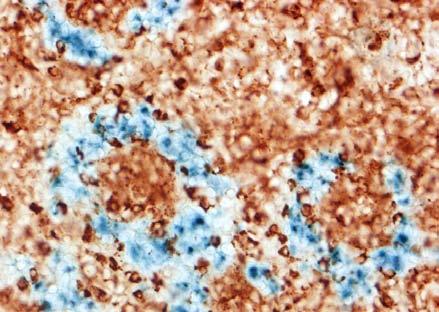
65 EREDMÉNYEK E F G H I J K L 65
66 EREDMÉNYEK 4. Az ellipszoid-asszociált sejtek vándorlása A β-gal-t az EAC-k endocitózissal veszik fel, amint az az elektronmikroszkópos képen is látható (14. kép: A). Az intravénás (i.v.) β-gal beadást követően akár 48 órával is azt tapasztaltuk, hogy a felvett antigén enzimaktivitása nem változott, vagyis nem bomlott le. 2 órával a beadás után az antigént felvett EAC-k kezdtek eltávolodni az ellipszoidtól (leválni az ellipszoid felszínéről). 24 óra elteltével az EAC-k elszórtan, ill. csoportokat alkotva a PALS-ban, elsősorban annak a széli részén, vagy a már meglevő csíracentrumokban voltak fellelhetők (14. kép: B, betét, C). A PWP-t elhagyó, citoplazmatikusan β-gal + EAC-k a vándorlás során fokozatosan elvesztették a 68.2 antigént, és a csíracentrumban lévő sejtek már ak (14. kép: D). A csíracentrumokban lévő β-gal + sejtek viszont expresszálják az FDC kre jellemző 74.3 markert (14. kép: E, betét). Az E5G12 kolokalizációja β-gal + sejtekkel ezen időszak alatt nem változott, azaz a β-gal pozitivitás mindig az E5G12 + sejtekben volt kimutatható (14. kép: F, betét). Négy nappal az antigén beadást követően a β-gal + EAC-k nagy része a csíracentrumokba lokalizálódott, amelyek száma időközben megduplázódott. Ekkor még találtunk kis számban β-gal + EAC-t az ellipszoid körül, az eddigre nagymértékben megvastagodott PWP-ben. 14 nappal a primer antigénbevitel után β-gal aktivitás egyes csíracentrumokban még kimutatható volt. Két héttel az első immunizálást követően a β-gal injekciót megismételtük. Az antigén megkötésében itt is kizárólag az EAC-k vettek részt. Hasonló időközökben mintát véve, azt tapasztaltuk, hogy a fent említett antigénszállítás felgyorsult. A β-gal-t felvevő EAC-k nagy része 24 óra elteltével már a csíracentrumokba vándorolt. 66
 β-gal és B-sejtes (Bu1, barna) kettősfestés. 24 órával az i.v. antigén beadást követően a β- gal + sejtek csoportokat alkotnak (nyíl), vagy bevándorolnak a csíracentrumokba (betét).")
67 EREDMÉNYEK 14. kép. Az EAC-k vándorlásának nyomonkövetése. (A) Az elektronmikroszkópos kép azt mutatja, hogy az X-gal csapadék intracelluláris vezikulumban található (nyíl). Betét: a vezikulumból kinagyított részlet. (B) β-gal és B-sejtes (Bu1, barna) kettősfestés. 24 órával az i.v. antigén beadást követően a β- gal + sejtek csoportokat alkotnak (nyíl), vagy bevándorolnak a csíracentrumokba (betét). (C) A CD3 + sejtek körbeveszik a β-gal + EAC-csoportot (nyíl). (D) Négy nappal az immunizálást követően a β-gal + EAC-k nagy része a csíracentrumokban található, és a vándorlás során elveszítik a 68.2 antigént (szürke). (E) A csíracentrumokba vándorolt EAC-k 74.3 antigént kezdenek expresszálni (szürke). Betét: 74.3 (zöld) és β-gal (piros) kettős immunfluoreszcens, konfokális képe. (F) Párhuzamos metszet az előbbivel, de E5G12-vel (szürke) festve. Betét: az E5G12 + sejtekre (zöld) lokalizálódik a β-gal pozitivitás (piros). Konfokális kép. 67
68 EREDMÉNYEK 5. A humorális immunválasz mérése A primer immunizálást követő 4. napon vett szérummintákban jelentősen megemelkedett a β-gal specifikus IgM mennyisége, kis mértékben az IgG-é is. A 12. napon ez az állapot teljes mértékben megváltozott: a szérum IgM titer lecsökkent a kontroll (nem immunizált) állatokban mért szintre, míg az IgG szint jelentősen megemelkedett (15. kép). 1,2 OD 492 (átlag+sd) 1 0,8 0,6 0,4 0,2 IgG IgM 0 kontroll 4 napos kezelt 4 napos kontroll 12 napos kezelt 12 napos 15. kép. A humorális immunválasz kvalitatív elemzése 6. Az antigén felvételének in vitro vizsgálata A lép volt az egyetlen szerv, ahol a keringésbe juttatott β-gal felvételre került. Ez felvetette annak a lehetőségét, hogy olyan specifikus receptor(ok) játszik szerepet, amely csak az EAC-kon van jelen. Kísérleteinkben először a humorális faktorok (természetes ellenanyagok, komplementek) potenciális szerepét vizsgáltuk az antigénfelvételben. Ezt in vitro túlélő lépszelet kultúrákban végeztük. Az izolált lépszeletek (átlagos vastagság kb. 500 µm) alapos mosásával távolítottuk el a bennük maradt szérumot. Ezt követően β-gal-t adtunk a szeletekhez. Az inkubáció után elvégzett enzimhisztokémiai reakció karakterisztikusan kirajzolta az ellipszoidokat 68
69 EREDMÉNYEK körülvevő PWP-t. Ezt követően a lépszeletek inkubálását β-gal-lal különböző módosításokkal végeztük. A β-gal felvétel akkor is zavartalan maradt, ha a lépszeleteket szérummentes médiumban tenyésztettük. Ugyanígy nem befolyásolta a normál és a hőinaktivált ortológ szérum jelenléte sem a β-gal felvételét. Abban az esetben viszont, amikor heparinizált plazmát adtuk a szeletekhez a β-gal-t, az antigén felvétele nagymértékben lecsökkent. Ezen eredmény arra késztetett, hogy kiderítsük a heparin gátló szerepét az antigénfelvételben. Ennek érdekében kompetíciós kísérleteket végeztünk, amelyben növekvő heparin koncentráció jelenlétében vizsgáltuk a β-gal felvételt. Negatív összefüggést találtunk a heparin koncentrációja és az antigén felvétel között, azaz minél nagyobb volt a heparin koncentrációja, annál inkább gátlódott az antigén felvétele; így 250 IU/ml heparin teljesen meggátolta a β-gal felvételt. A heparinhoz hasonlóan a dextrán-szulfát ugyancsak teljes gátlást okozott 10 µm koncentrációban. A semleges dextrán viszont mindezt nem befolyásolta. A kompetíciós vizsgálatok eredményeit a 6. táblázat összesíti. 6. Táblázat. A β-gal in vitro felvételének tanulmányozása Kontroll ++++ PBS , IU 125 IU 62,5 IU Dextrán Dextrán IU heparin heparin heparin 20 µm 10 µm heparin Dextrán Dextrán -szulfát - szulfát 20 µm 10 µm - -/ : Az enzimhisztokémiai reakció erősségét jelenti. ++++: erős festődés, +/-: még észlelhető festődés, -: nincs festődés. 7. A β-galaktozidáz specifikus receptor(ok)hoz kötődik Előzetes vizsgálatok szerint a β-gal megkötésében specifikus receptor(ok) vesz részt. A biotin és a radioaktív jóddal végzett kísérletek hasonló eredményeket szolgáltattak. β-gal gyöngyökkel precipitált fehérjék molekulatömege mindkét esetben 52-53, ill. ~43-45 kda volt (16. kép: 4-5 csík, nyílak; redukált környezetben). Nem redukált mintapufferrel nem sikerült a precipitált fehérjéket a β-gal gyöngyökről 69
. 2. Molekulasúly standard. 3. Biotinilált fehérjekivonat+β-gal+β-gal gyöngy (20 µl). 4. Biotinilált fehérjekivonat+β-gal gyöngy (20 µl). 5.")
70 EREDMÉNYEK eltávolítani (nincs kép). Szolúbilis β-gal hozzáadással gátolható volt a receptorok β-gal beadekhez való kötődése (3. csík) kép. A β-galaktozidáz megkötésében szerepet játszó fehérjék azonosítása. 1. Biotinilált membránfrakció (5 µl). 2. Molekulasúly standard. 3. Biotinilált fehérjekivonat+β-gal+β-gal gyöngy (20 µl). 4. Biotinilált fehérjekivonat+β-gal gyöngy (20 µl). 5. Biotinilált fehérjekivonat+β-gal bead (10 µl). 6. Üres sepharose gyöngy+biotinilált fehérjekivonat (10 µl). A nyílak a receptorokra mutatnak. 10 %-os redukált SDS-PAGE. Előhívás kloro-1-naphtol kittel történt. 8. Az EAC-k szerepe a baktériumfelvételben Az EAC-k nemcsak a bakteriális antigén-, hanem a teljes baktérium felvételére is képesek. 45 perccel a hővel elölt Streptococcus pneumoniae beadását követően az EAC-k aktívan fagocitálják őket (17. kép: A, B). Immunfluoreszcens festéssel, illetve elektronmikroszkóposan bizonyítottuk, hogy az EAC-kon kívül sem a lép makrofágjai, sem pedig az odavándorló granulociták nem veszik fel a baktériumokat. A β-galtól 70
 A véráramba került baktériumokat (zöld) szelektíven az EAC-k (68.2, piros) veszik fel. Konfokális kép. (B) Elektronmikroszkópos képen is jól látható, hogy az EAC (nyíl) veszi fel a baktériumot.")

71 EREDMÉNYEK eltérően a baktérium felvétele azonban nem lokalizálódott csak a lépre. A máj sejtjei is nagymértékű baktérium fagocitózist mutattak. 17. kép. EAC-k baktérium felvétele. (A) A véráramba került baktériumokat (zöld) szelektíven az EAC-k (68.2, piros) veszik fel. Konfokális kép. (B) Elektronmikroszkópos képen is jól látható, hogy az EAC (nyíl) veszi fel a baktériumot. Betét: a fagocitált baktérium kinagyított képe. A szaggatott vonal a Pc-t határolja. 9. Az EAC-k nemcsak a β-gal-t veszik fel Az EAC-k hatékony és szelektív β-gal felvételét hisztokémiai módszerekkel könnyen bizonyítani lehetett, míg más antigén esetében, pl. sztreptavidin, ez csak elektronmikroszkópiával sikerült. Az arannyal jelzett sztreptavidin is az EAC-k endocitotikus vezikulumaira lokalizálódott (18. kép). 18. kép. Az EAC mint univerzális antigénfelvevő sejt. Az EAC-kban az arannyal jelzett sztreptavidin is endocitotikus vezikulumba kerül (lásd a kinagyított részletet). 71
72 MEGBESZÉLÉS MEGBESZÉLÉS A CSIRKE EPIDERMÁLIS DENDRITIKUS SEJTJEI Doktori munkám első részében elvégeztem a madárbőr epidermális dendritikus sejtjeinek (EDC) részletes jellemzését egy nagy ellenanyagpanel segítségével. Az ATPáz hisztokémiát immunfluoreszcens festésekkel kombinálva bebizonyítottuk, hogy a madár epidermális dendritikus sejtjei, akárcsak az emlős LC-k, CD45 és vimentin pozitívak (Romani és mtsai 1989, de Waal és mtsai 1984). Ez a három marker teljes mértékben átfed egymással. A CD45 pozitivitás az EDC-k hemopoetikus (csontvelői) eredetét támasztja alá. Az EDC-kkel ellentétben a környező keratinociták nem expresszálják az előbb említett markereket. Az epidermiszben ugyanakkor találtunk melanocitákat (melyek kimutatását a MelEM mea segítségével végeztük; ezek a sejtek CD45 negatívak) is, de ezek száma még a pigmentált New Hampshire törzsben ismeglehetősen alacsony volt (2-3 sejt/mm 2 ). Ezek alapján a későbbiekben az EDC-k azonosítására vagy a CD45, vagy a vimentin festést használtuk azokban a kettősfestésekben, melyekkel az EDC-kre jellemző további markereket kerestük. A következő vizsgált markerek az antigénprezentációban fontos MHCI és II molekulák voltak. Az MHCII-festések alapján elmondható, hogy az EDC-k közel fele (átlagosan 40%-a) hordozza ezt a molekulát. Ez ellentmond Perez-Torres és kollégái által közölt adatoknak, miszerint valamennyi ATPáz + sejt egyúttal MHCII + is. Az ellentmondás hátterében feltehetőleg az áll, hogy ők nem használtak szimultán kettősfestést. Az MHCII immunfluoreszcens festést lefényképezték, majd ezt követően ATPáz hisztokémiát alkalmaztak a nyúzatokon (ami eltakarta a fluoreszcens jelet). Az itt kapott festődést újra lefényképezték és a két képet egymásra vetítették. Az általunk alkalmazott kettős immunfluoreszcencia a két marker egyidejű vizsgálatát teszi lehetővé. Az EDC-k csoportosítása szempontjából fontos ellenanyagnak bizonyult a CD3. A hámban lévő CD45 + dendritikus sejtek körülbelül 5-10%-a hordozza ezt az antigént. A különböző kettősfestési kombinációkkal kiderítettük, hogy a hámban három EDC alpopuláció különíthető el: az I. típus MHCII + /CD3 -, a II. típus MHCII - /CD3 - és a III. típus MHCII - /CD
73 MEGBESZÉLÉS Az MHCII + CD3 - EDC-ket az I. csoportba soroltuk. Az MHCII expressziója ezen populáción belül sem volt egyenletes. A kisebb csoportokat, tömörüléseket alkotó sejtek erősebb MHCII expressziót mutattak. Hasonló expressziós mintázatbeli különbségeket emlős LC-kre is leírtak (Dezutter-Dambuyant és mtsai 1984). Az ebbe a csoportba tartozó sejtek fenotípusa áll a legközelebb az emlős LC-khez. A monocita-dendritikus sejt differenciálódás lehetőségét húzza alá, hogy az MHCII + EDC alpopuláció további két monocita-makrofág markerre, a KUL01-re és a 68.1-re volt pozitív. A II. csoportba soroltuk az MHCII - /CD3 - sejteket. Ezekről a sejtekről azt feltételezzük, hogy a megfelelő stimulus(ok) hatására MHCII + EDC-vé érnek, azaz az I. típusú EDC-k prekurzorai vagy nyugvó formái. Ezt támasztja alá az in vitro tenyészeten tett megfigyelésünk is, hogy a perkután immunizálás során kivándorolt FITC + sejtek ~100 %-a MHCII + volt. A harmadik csoportba sorolt sejtek (III. típus) az összes EDC-k mintegy 5-10 %- át teszik ki. A CD3 expresszió ezeken a sejteken nem jár sem a T sejt receptorok (α/β vagy γ/δ), sem pedig a CD4, CD8 expressziójával. Így ezeket a sejteket nem tekinthetjük T sejteknek. Ugyanakkor a madarak NK sejtjei intracitoplazmatikus CD3 pozitivitással és a fenti markerek hiányával jellemezhetők (Göbel és mtsai 1996). Ezeket a sejteket tehát intraepiteliális NK sejteknek tekinthetjük. A madarakban a bőr hámjában még nem írták le NK sejtek jelenlétét. Emlősök esetében (elsősorban az emberben) nagyon kevés adat áll rendelkezésünkre (Cameron és mtsai 2002). Ezen közleményekben elszórt NK sejtek jelenlétét mutatták ki normál humán epidermiszben. Ezzel ellentétben mi egy konstans, eléggé számottevő NK sejtpopulációt azonosítottunk a csirke epidermiszben. Az itt található NK sejtek szerepet játszhatnak az esetleges vírusfertőzések leküzdésében. Érdekes összefüggés lehet, hogy emlős adatok szerint a vírusfertőzésekben a LC-k nem biztosítanak megfelelő védelmet, a feladatot egy más DC alpopuláció kell elvégezze. Funkcionális kísérleteinkben azt találtuk, hogy perkután alkalmazott haptén (FITC) hatására a CD3 pozitív sejtek nem vándorolnak ki az epidermiszből. Ez arra utal, hogy másféle stimulust igényelnek az aktivációjukhoz. A dendritikus morfológia feltehetőleg a nagyobb érintkezési felület létrejöttét szolgálja. Madarakban eddig a nyálkahártyákban igazolták NK és T-sejtek együttes jelenlétét a hámsejtek között (Göbel és mtsai 2001). Ugyanakkor mi semmilyen T-sejtet nem 73
74 MEGBESZÉLÉS találtunk a csirkebőr hámjában (szemben az egér epidermális γ/δ TCR + T sejtjeivel, az ún. DETC-kel). Érdekes különbséget találtunk a madár EDC-k és az emlős LC-k adhéziós molekuláit tekintve. Az emlősökben a LC-k erőteljesen expresszálják az E(epiteliális)- kadherint, amely szerepet játszik a hámsejtek között tartásukban (Tang és mtsai 1993). A csirkében az EDC-k E-kadherin expressziója nagyon kismértékű (elszórtan látható egy-egy folt a CD45 + sejteken), és ez a sejtek enzimatikus izolálása után már nem is detektálható. A madárbőr jóval vékonyabb az emlős bőrénél. A bazális (germinális) sejtsoron kívül csak egy-két tüskés (spinosum) réteg különíthető el benne. Emiatt gyakorlatilag minden EDC kisebb-nagyobb felületen érintkezik a bazális membránnal. Ezen elhelyezkedés magyarázattal szolgálhat arra, hogy a madár EDC-k miért csak néhány folt szintjén hordoznak E-kadherint. Az EDC-k epidermiszben való maradását, ill. kivándorlásukat elősegítheti a β1-integrin, az egyedüli integrin alegység, amelyet az EDC-ken azonosítani tudtunk. A β1-integrin a bazális membrán lamininját ismeri fel (Staquet és mtsai 1991). Mindez nem zárja ki, hogy más kadherinek expresszálódjanak az EDC-ken. Ezek kimutatására nem állt rendelkezésünkre ellenanyag. A mai napig semmilyen adat nincs a madár EDC-k ontogenézisét tekintve. Az emlős LC-kkel ellentétben, ahol a felnőtt állatra jellemző sejtszám a késői embrionális élet során kialakul (Foster és Holbrook 1989), a NH csirkék epidermiszében kikelés előtt csak néhány EDC-t találtunk. Ez a szám fokozatosan nő és az 5-8 hetes csirkékben stabilizálódik ~2000 sejt/mm 2 körül. A kísérleteinkben használt másik csirkefajta, a WL, nagymértékben eltért a NH-től. A WL epidermiszében az EDC-k a kikelést követő második hét végén kezdenek csak megjelenni (2-3 sejt/mm 2 ). A másik fontos eltérés, hogy ebben a törzsben az EDC-szám még felnőtt állatokban is nagy ingadozást mutatott. Ezt a különbséget egyrészt a genetikai háttér, másrészt a tojások eltérő higiénés státusa adhatja (SPF, illetve konvencionális). A LC-k kinetikáját alaposan tanulmányozták mind egér-, mind pedig humán bőrben (Czernielewski és mtsai 1985, Larregina és mtsai 2001, Merad és mtsai 2002). Ezek a vizsgálatok bebizonyították, hogy a hámban lévő LC-k osztódnak. Normális körülmények között (azaz ha a hámot nem éri immunológiai stimulus, pl. fertőzés), az LC-k mennyiségét elsősorban az osztódás és a konstitutív elvándorlás határozza meg, a 74
75 MEGBESZÉLÉS folyamatos pótlódás a dermis felől nagyon kismértékű. Ha a hámot valamilyen inger éri, a LC-ek elvándorlása fokozódik, és az utánpótlásuk most már főleg a bevándorlásból valósul meg, és csak kisebb mértékben az in situ osztódásból. Intravénásan adott BrdU-t használva elsőként bizonyítottuk, hogy a csirkebőr hámjában az EDC-k osztódásra képesek. A BrdU adást követő 30 perc és 24 óra közti időszakban az EDC-k viszonylag állandó jelölődést mutattak. Hasonló eredményeket kaptunk az in vitro BrdU-jelöléses kísérleteinkben, amellyel a már a dermiszben osztódott prekurzorok bevándorlása a hámba kizárható. Ezen adatok alapján végzett becsléseink szerint a csirke bőrében az EDC-k mintegy nap alatt újulnak meg teljesen (beleszámítva az EDC-k konstitutív emigrációját, amely a perifériás tolerancia fenntartásáért felelős). A munkám következő részében azt vizsgáltam, hogy képesek-e az EDC-k antigének felvételére és ezt követően a hámból való kivándorlásra. Funkcionális vizsgálatainkkal bebizonyítottuk, hogy az EDC-k részét képező madár LC-szerű sejtek képesek válaszolni antigénstimulusra. Bebizonyítottuk, hogy nemcsak a fenotípusuk hasonlít az emlős LC-khez, hanem funkcionálisan is teljesítik az LC kritériumot. Ezért az EDC-k két típusát (I-II) az LC-szerű elnevezés helyett csirke Langerhans sejt (csls) elnevezés illeti meg. A hám felületére juttatott haptén hatására a csls-ek kivándorolnak az epidermiszből, amint azt a kezelt területről vett epidermisz nyúzatokban tapasztaltuk. Ez késztetett arra minket, hogy nyomonkövessük ezen sejtek vándorlását. Mivel a csirkékben hiányoznak a perifériás nyirokcsomók, ezért először a távoli nyirokszerveket vizsgáltuk meg, az antigénkezelést követő 24 óra múlva. Egyetlen megvizsgált primerés szekunder limfoid szervben, sem pedig a vérsejtek között nem találtunk haptén pozitív sejtet. Az eredménytelenségnek oka az lehet, hogy a kivándorolt sejtek 24 óra alatt nem érték el a célszervet. Ezek az eredmények azonban arra késztettek, hogy tovább vizsgálódjunk. A FITC-cel kezelt bőrből készített sorozatmetszetek vizsgálata során FITC + cslc-ek voltak láthatók elszórtan a dermiszben, valamint nyiroksejtcsoportokban voltak azonosíthatók. Ezen eredmény fordulópontot jelentett a cslc-k vándorlási útvonalának feltérképezésében. Felvetődött annak a lehetősége, hogy a cslc-k nem hagyják el a bőrt, hanem azt itt jelenlevő nyiroksejt akkumulációkba vándorolnak. Ezeknek az általunk limfoid nodulusoknak nevezett struktúráknak a sejtes összetételéről semmilyen irodalmi adat nem állt rendelkezésünkre. Ezen nodulusok jelenlétét az 50-es években többféle szövetben (bőr, máj, tüdő stb.) leírták, de nem sok 75
76 MEGBESZÉLÉS figyelmet szenteltek nekik (Biggs 1957). A nodulusok néhány száz nyiroksejtből és járulékos sejtekből álló, kerekded képletek a dermiszben. Legtöbbször nyirokerek és vénák mentén jelennek meg. Eddigi adataink szerint (axilláris bőr) 1 cm 2 területen 1-5 közötti a számuk. A nodulusok átlagosan 100 µm-rel a hám alatt, a dermisz kötőszövetében találhatók. A nodulusok további vizsgálata az azóta eltelt időszakban nem folytatódott, lényegében véletlenszerűen aggregálódott nyiroksejtcsoportnak tekintették ezeket. Minthogy kapcsolatba voltak hozhatóak a bőrön keresztüli immunizálással (lásd alább) ezért szükségessé vált a szerkezetük, összetételük alaposabb vizsgálata. Immunhisztokémiai festéseink szerint a nodulusok vázát egy nyúlványos sejtekből álló retikulum alkotja. A nodulus központi részén ezek a nyúlványos sejtek a csíraközpontokra jellemző FDC markert (74.3) hordozzák. A retikulum perifériáján lévő nyúlványos sejtek ezzel szemben KUL01 + -ak. A hálózat belső terét T- és B-sejtek töltik ki. Ez a sejtösszetétel emlékeztet a szekunder nyirokszervek csíracentrumára, de az utóbbiakban elszórtan vagy egyáltalán nem találunk KUL01 pozitivitást. A nodulusokra nagymértékű MHCII expresszió jellemző, amit feltehetőleg a DC-retikulum és a B-limfociták kölcsönöznek. Hapténnal (FITC) történt szenzitizáció után, a kezelt bőrterülethez tartozó nodulusok metszetén mindig található FITC jel. A FITC szemcsék citoplazmatikusan jelennek meg, méghozzá kizárólag a retikulum sejtjeiben: a KUL01 + -, és kevésbé a sejtekben. Ez a lokalizáció felveti annak a lehetőségét, hogy a bőrből kivándorló cslc-ek, amelyek KUL01 pozitívak, vándorolnak a nodulusok perifériájára. Másik feltételezésünk, hogy az ide homingoló" LC-ek, továbbdifferenciálódnak, és sejtekké alakulnak. Ennek bizonyítására mindenképpen szükséges lesz a továbbiakban megismételni ezeket a kísérleteket fehérjeantigénekkel és izolált LC-kkel. A használt módszerrel nem lehet teljes mértékben kizárni azt sem, hogy egyes haptén + sejt eljut a lépbe vagy más nyirokszervbe. Ugyanakkor még azonosításra váró végállomása is létezhet a cslc-knek. Ha azonban bizonyíthatóvá válik, hogy ezek a képletek valóban befogadják a hámból kivándorolt LC-eket (azaz kialakulásuk antigéningerhez kötött), abban az esetben a nodulusokat az emlősök nyirokcsomójához hasonló funkciót ellátó szerkezeteknek (analógjainak) tekinthetjük. Ebben az esetben szerveződésük vizsgálata a nyirokcsomóképződés mechanizmusait tárhatja fel. Ugyanakkor további kísérletek szükségesek azon kérdés eldöntésére, hogy ezek a nodulusok genetikailag 76
77 MEGBESZÉLÉS determináltak-e vagy sem. Mindezek alaposabb megismerése nagyban elősegítheti a csirke bőrimmunitásának megértését, és egyben megkönnyítheti az emlősökben lezajló folyamatok értelmezését is. A cslc-k vándorlási útvonalát a 19. kép foglalja össze. AZ ELLIPSZOID-ASSZOCIÁLT SEJT, MINT FOLLIKULÁRIS DENDRITIKUS SEJT PREKURZOR Doktori munkám második részét egy másik járulékos sejtféleség, az ellipszoidasszociált sejt (EAC) vizsgálata jelentette. Ez a sejttípus a madárlép ellipszoidját körülvevő PWP alkotóeleme. Az ellipszoid, mely a penicilliform kapillárisokat veszi körül, egy főleg támasztó sejtekből álló hüvely az ér körül. Ezen kívül helyezkedik el, attól bazális lamina-szerű mátrix-szal elválasztva a PWP, mely az emlőslép marginális zónájával rokonítható terület. A PWP-t három fő sejttípus építi fel: EAC, limfociták és makrofágok. Mind elektronmikroszkóposan, mind immunhisztokémiai vizsgálatokkal egyértelműen kiderült, hogy az EAC-k nem limfociták és nem makrofágok. Az irodalom sokszor ellipszoid-asszociált retikulumsejteknek is hívja őket. Leírásukat követően több dolgozat foglalkozott az EAC-k antigénfelvételben játszott szerepével, és az esetleges EAC-FDC differenciációval. Ezek a kísérletek igazolták, hogy az EAC-k különféle anyagok/antigének felvételére képesek. Antigén felvételt követően különböző időközökben vett minták elektronmikroszkópos vizsgálata azt mutatta, hogy leválnak az ellipszoid felszínéről és a lépen belül vándorolni kezdenek. A PALS területén, ill. esetenként a csíracentrumokban azonosítottak hasonló morfológiájú sejteket. Ekkor még nem álltak rendelkezésre EAC, ill. FDC specifikus ellenanyagok, ami megnehezítette ezen sejtek nyomonkövetését. Azóta rendelkezünk olyan monoklonális ellenanyagokkal, melyek vagy az EAC-kre (68.2), vagy az FDC-kre (74.3) specifikusak. Olyan ellenanyag azonban nem ismert, amely mind a két sejttípust felismerné. A későbbi vizsgálatok során, pedig nem használtak kettős jelöléseket az EAC-FDC probléma megoldására. A másik probléma, hogy az intravénásan adott antigéneket több, különböző sejttípus vette fel, nem volt ismert olyan antigén, amelyet csak az EAC-k vesznek fel. Ezen hiányosságok pótlásával sikerült alaposan megvizsgálni a csirke lép FDC-inek differenciálódását. 77
 is létezhet (kérdőjel). Feltehetőleg itt is létezik az emlős DC-knél tapasztalt DC érési folyamat (idc-mdc).")
78 MEGBESZÉLÉS 19. kép A csirke dendritikus sejtek életciklusa. Az antigént/haptént felvevő cslc-k az aktivációt követően a környező dermális limfoid nodulusokba vándorolnak. Ezen az azonosított vándorlási végállomáson kívül más terminális(ok) is létezhet (kérdőjel). Feltehetőleg itt is létezik az emlős DC-knél tapasztalt DC érési folyamat (idc-mdc). Feltehetőleg a nodulusokba érkezett cslc-k hatékony immunválaszt indukálnak. Ha a kórokozó a véráramba kerül, akkor azt a lép EAC-k veszik fel. Bebizonyítottuk, hogy az EAC-k (mint éretlen DC) az antigénfelvételt követően két útvonalon keresztül differenciálódhatnak FDCkké (EAC-IDC-FDC, ill. EAC-FDC). Kérdőjelek: egyáltalán nem ismert útvonalak. 78
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE
 Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer sejtjei, differenciálódási antigének
 Az immunrendszer sejtjei, differenciálódási antigének Immunológia alapjai 2. hét Immunológiai és Biotechnológiai Intézet Az immunrendszer sejtjei Természetes/Veleszületett Immunitás: Granulociták (Neutrofil,
Az immunrendszer sejtjei, differenciálódási antigének Immunológia alapjai 2. hét Immunológiai és Biotechnológiai Intézet Az immunrendszer sejtjei Természetes/Veleszületett Immunitás: Granulociták (Neutrofil,
Az adaptív immunválasz kialakulása. Erdei Anna Immunológiai Tanszék ELTE
 Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Immunológia alapjai előadás. A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és
 Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Antigén, Antigén prezentáció
 Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Az immunológia alapjai
 Az immunológia alapjai 8. előadás A gyulladásos reakció kialakulása: lokális és szisztémás gyulladás, leukocita migráció Berki Timea Lokális akut gyulladás kialakulása A veleszületeh és szerzeh immunitás
Az immunológia alapjai 8. előadás A gyulladásos reakció kialakulása: lokális és szisztémás gyulladás, leukocita migráció Berki Timea Lokális akut gyulladás kialakulása A veleszületeh és szerzeh immunitás
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok.
 Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
Immunológia alapjai 7-8. előadás Adhéziós molekulák és ko-receptorok. Az immunválasz kezdeti lépései: fehérvérsejt migráció, gyulladás, korai T sejt aktiváció, citokinek. T sejt receptor komplex ITAMs
Immunológia I. 4. előadás. Kacskovics Imre
 Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Az immunrendszer sejtjei, differenciálódási antigének
 Az immunrendszer sejtjei, differenciálódási antigének Immunológia alapjai 2. hét Immunológiai és Biotechnológiai Intézet Az immunrendszer sejtjei Természetes/Veleszületett Immunitás: Granulociták (Neutrofil,
Az immunrendszer sejtjei, differenciálódási antigének Immunológia alapjai 2. hét Immunológiai és Biotechnológiai Intézet Az immunrendszer sejtjei Természetes/Veleszületett Immunitás: Granulociták (Neutrofil,
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek
 Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek Dr. Németh Péter PTE-KK Immunológiai és Biotechnológiai Intézet Mi az immunrendszer? Az immunrendszer a szervezet
Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek Dr. Németh Péter PTE-KK Immunológiai és Biotechnológiai Intézet Mi az immunrendszer? Az immunrendszer a szervezet
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia alapjai előadás. Sej-sejt kommunikációk az immunválaszban.
 Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
Immunológia alapjai 7-8. előadás Sej-sejt kommunikációk az immunválaszban. Koreceptorok és adhéziós molekulák. Cytokinek, chemokinek és receptoraik. A sejt-sejt kapcsolatok mediátorai: cross-talk - Szolubilis
Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek
 Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek Dr. Németh Péter PTE-KK Immunológiai és Biotechnológiai Intézet Mi az immunrendszer? Az immunrendszer a szervezet
Az immunrendszer ontogenezise, sejtjei, differenciálódási antigének és az immunszervek Dr. Németh Péter PTE-KK Immunológiai és Biotechnológiai Intézet Mi az immunrendszer? Az immunrendszer a szervezet
B-sejtek szerepe az RA patológiás folyamataiban
 B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
INTRACELLULÁRIS PATOGÉNEK
 INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia. Hogyan működik az immunrendszer? http://www.szote.u-szeged.hu/mdbio/oktatás/immunológia password: immun
 Immunológia Hogyan működik az immunrendszer? http://www.szote.u-szeged.hu/mdbio/oktatás/immunológia password: immun Hogyan működik az immunrendszer? Milyen stratégiája van? Milyen szervek / sejtek alkotják?
Immunológia Hogyan működik az immunrendszer? http://www.szote.u-szeged.hu/mdbio/oktatás/immunológia password: immun Hogyan működik az immunrendszer? Milyen stratégiája van? Milyen szervek / sejtek alkotják?
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Mikrogliák eredete és differenciációja
 Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
KLINIKAI IMMUNOLÓGIA I.
 Kórházhigienikus képzés, DE OEC KLINIKAI IMMUNOLÓGIA I. AZ IMMUNRENDSZER MŰKÖDÉSE Dr. Sipka Sándor DE OEC III. sz. Belgyógyászati Klinika Regionális Immunológiai Laboratórium A főbb ábrák és táblázatok
Kórházhigienikus képzés, DE OEC KLINIKAI IMMUNOLÓGIA I. AZ IMMUNRENDSZER MŰKÖDÉSE Dr. Sipka Sándor DE OEC III. sz. Belgyógyászati Klinika Regionális Immunológiai Laboratórium A főbb ábrák és táblázatok
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása. Uher Ferenc, PhD, DSc
 A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői.
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Doktori értekezés tézisei
 Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai előadás MHC. szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Az ellenanyagok szerkezete és funkciója. Erdei Anna Immunológiai Tanszék ELTE
 Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Immunológia alapjai előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok:
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek
 A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
A kemotaxis kiváltására specializálódott molekula-család: Cytokinek Cytokinek - definíció Cytokin (Cohen 1974): Sejtek közötti kémi miai kommunikációra alkalmas anyagok; legtöbbjük növekedési vagy differenciációs
http://www.rimm.dote.hu Tumor immunológia
 http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
Lukácsi Szilvia
 Immunológia II GY 2018.04.13. Lukácsi Szilvia 7. Sterilitás, in vitro sejttenyésztés Az immunrendszer sejtjeinek izolálása, azonosítása és funkcionális vizsgálata: I. Monociták, makrofágok, dendritikus
Immunológia II GY 2018.04.13. Lukácsi Szilvia 7. Sterilitás, in vitro sejttenyésztés Az immunrendszer sejtjeinek izolálása, azonosítása és funkcionális vizsgálata: I. Monociták, makrofágok, dendritikus
Immunológia 4. A BCR diverzitás kialakulása
 Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
1. előadás Immunológiai alapfogalmak. Immunrendszer felépítése
 1. előadás Immunológiai alapfogalmak. Immunrendszer felépítése Vér alakos elemei: 1mm3 vérben: 4-5 millió vörövértest 6000-9000 fehérvérssejt 200-400 ezer thrombocyta(vérlemezke) Fehérvérsejtek: agranulocyták:
1. előadás Immunológiai alapfogalmak. Immunrendszer felépítése Vér alakos elemei: 1mm3 vérben: 4-5 millió vörövértest 6000-9000 fehérvérssejt 200-400 ezer thrombocyta(vérlemezke) Fehérvérsejtek: agranulocyták:
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Preeclampsia-asszociált extracelluláris vezikulák
 Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
1. Az immunrendszer működése. Sejtfelszíni markerek, antigén receptorok. 2. Az immunrendszer szervei és a leukociták
 Sejtfelszíni markerek, antigén receptorok A test őrei 1. Az immunrendszer működése Az individualitás legjobban az immunitásban mutatkozik meg. Feladatai: - a saját és idegen elkülönítése, felismerése -
Sejtfelszíni markerek, antigén receptorok A test őrei 1. Az immunrendszer működése Az individualitás legjobban az immunitásban mutatkozik meg. Feladatai: - a saját és idegen elkülönítése, felismerése -
Irányzatok a biológiában: IMMUNOLÓGIA
 Irányzatok a biológiában: IMMUNOLÓGIA Dr. Kacskovics Imre tszv. egy. tanár Immunológiai Tanszék ELTE http://immunologia.elte.hu/ Medicina Kiadó 2012. Az Immunológiai Tanszék kutatási témái: http://immunologia.elte.hu/
Irányzatok a biológiában: IMMUNOLÓGIA Dr. Kacskovics Imre tszv. egy. tanár Immunológiai Tanszék ELTE http://immunologia.elte.hu/ Medicina Kiadó 2012. Az Immunológiai Tanszék kutatási témái: http://immunologia.elte.hu/
A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet
 A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet Mi a feladata az Immunrendszernek? 1. Védelem a kórokozók ellen 2. Immuntolerancia fenntartása Mik is azok a kórokozók? Kórokozók alatt
A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet Mi a feladata az Immunrendszernek? 1. Védelem a kórokozók ellen 2. Immuntolerancia fenntartása Mik is azok a kórokozók? Kórokozók alatt
Immunológia I. 2. előadás. Kacskovics Imre (imre.kacskovics@ttk.elte.hu)
") Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Ajánlott tankönyvek: Gergely János, Erdei Anna: Immunbiológia. Falus András: Az immunológia élettani és molekuláris alapjai
 Ajánlott tankönyvek: Gergely János, Erdei Anna: Immunbiológia Falus András: Az immunológia élettani és molekuláris alapjai Janeway, Travers, Walport, Schlomchik: Immunobiology Paraziták Virulencia egyes
Ajánlott tankönyvek: Gergely János, Erdei Anna: Immunbiológia Falus András: Az immunológia élettani és molekuláris alapjai Janeway, Travers, Walport, Schlomchik: Immunobiology Paraziták Virulencia egyes
Szervezetünk védelmének alapja: az immunológiai felismerés
 Szervezetünk védelmének alapja: az immunológiai felismerés Erdei Anna ELTE, TTK, Biológiai Intézet Immunológiai Tanszék ELTE, Pázmány-nap, 2012. Az immunrendszer fő feladata a gazdaszervezet védelme a
Szervezetünk védelmének alapja: az immunológiai felismerés Erdei Anna ELTE, TTK, Biológiai Intézet Immunológiai Tanszék ELTE, Pázmány-nap, 2012. Az immunrendszer fő feladata a gazdaszervezet védelme a
2016. nov. 8. Bajtay Zsuzsa
 6. Komplementreceptorok fajtái és szerepük az immunválasz során 2016. nov. 8. Bajtay Zsuzsa A komplementrendszer - Vérben, testnedvekben inaktív állapotban jelenlévő - egymást láncreakcióban aktiváló faktorok
6. Komplementreceptorok fajtái és szerepük az immunválasz során 2016. nov. 8. Bajtay Zsuzsa A komplementrendszer - Vérben, testnedvekben inaktív állapotban jelenlévő - egymást láncreakcióban aktiváló faktorok
Immunbiológia 4. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe Antigénprezentáció Engelmann Péter
 Immunbiológia 4. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe Antigénprezentáció Engelmann Péter MHC A saját és idegen antigének a gazdaszervezet specializált
Immunbiológia 4. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe Antigénprezentáció Engelmann Péter MHC A saját és idegen antigének a gazdaszervezet specializált
Kutatási beszámoló ( )
") Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Természetes immunitás
 Természetes immunitás Ősi: Gyors szaporodású mikroorganizmusok ellen azonnali védelem kell Elterjedés megakadályozása különben lehetetlen Azonnali reakciónak köszönhetően a fertőzést sokszor észre sem
Természetes immunitás Ősi: Gyors szaporodású mikroorganizmusok ellen azonnali védelem kell Elterjedés megakadályozása különben lehetetlen Azonnali reakciónak köszönhetően a fertőzést sokszor észre sem
AZ IMMUNRENDSZER VÁLASZAI A HPV FERTŐZÉSSEL KAPCSOLATOS KÉRDÉSEINKRE RAJNAVÖLGYI ÉVA DE OEC Immunológiai Intézet
 AZ IMMUNRENDSZER VÁLASZAI A HPV FERTŐZÉSSEL KAPCSOLATOS KÉRDÉSEINKRE RAJNAVÖLGYI ÉVA DE OEC Immunológiai Intézet A méhnyak rák előfordulása / év / 100 000 nő WHO 2005 A KÓROKOZÓK ÉS AZ IMMUNRENDSZER KÉTIRÁNYÚ
AZ IMMUNRENDSZER VÁLASZAI A HPV FERTŐZÉSSEL KAPCSOLATOS KÉRDÉSEINKRE RAJNAVÖLGYI ÉVA DE OEC Immunológiai Intézet A méhnyak rák előfordulása / év / 100 000 nő WHO 2005 A KÓROKOZÓK ÉS AZ IMMUNRENDSZER KÉTIRÁNYÚ
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Immunológia Világnapja
 a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
Az immunológia alapjai (2018/2019. II. Félév)
") Az immunológia alapjai (2018/2019. II. Félév) A CELLULÁRIS IMMUNVÁLASZ ÉS EFFEKTOR FOLYAMATAI http://www.nobelprize.org/ Az adaptív immunválasz során a B- limfocitákból plazmasejtek keletkeznek, melyek
Az immunológia alapjai (2018/2019. II. Félév) A CELLULÁRIS IMMUNVÁLASZ ÉS EFFEKTOR FOLYAMATAI http://www.nobelprize.org/ Az adaptív immunválasz során a B- limfocitákból plazmasejtek keletkeznek, melyek
A T sejtes immunválasz egy evolúciós szempontból váratlan helyzetben: Szervtranszplantáció
 A T sejtes immunválasz egy evolúciós szempontból váratlan helyzetben: Szervtranszplantáció Autotranszplantáció: saját szövet átültetése, pl. autológ bőrtranszplantáció, autológ őssejt-transzplantáció.
A T sejtes immunválasz egy evolúciós szempontból váratlan helyzetben: Szervtranszplantáció Autotranszplantáció: saját szövet átültetése, pl. autológ bőrtranszplantáció, autológ őssejt-transzplantáció.
4. A humorális immunválasz október 12.
 4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
Immunológia alapjai
 Immunológia alapjai 2011.11.03. A sejt-mediálta immunválasz effektor mechanizmusai (CMI): 1. Citotoxicitás 2. T H sejt mediálta makrofág aktiváció (Késői típusú hyperszenzitivitás = DTH.) Az adaptív immunválasz
Immunológia alapjai 2011.11.03. A sejt-mediálta immunválasz effektor mechanizmusai (CMI): 1. Citotoxicitás 2. T H sejt mediálta makrofág aktiváció (Késői típusú hyperszenzitivitás = DTH.) Az adaptív immunválasz
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI. - autokrin. -neurokrin. - parakrin. -térátvitel. - endokrin
 A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
TÚLÉRZÉKENYSÉGI I. TÍPUSÚ TÚLÉRZÉKENYSÉGI REAKCIÓ 2013.04.21. A szenzitizáció folyamata TÚLÉRZÉKENYSÉGI REAKCIÓK ÁTTEKINTÉSE TÚLÉRZÉKENYSÉGI REAKCIÓK
 TÚLÉRZÉKENYSÉGI REAKCIÓK TÚLÉRZÉKENYSÉGI REAKCIÓK Ártalmatlan anyagok bejutása egyes emberekben túlérzékenységi reakciókat válthat ki Nemkívánatos gyulladáshoz, sejtek és szövetek károsodásához vezet Az
TÚLÉRZÉKENYSÉGI REAKCIÓK TÚLÉRZÉKENYSÉGI REAKCIÓK Ártalmatlan anyagok bejutása egyes emberekben túlérzékenységi reakciókat válthat ki Nemkívánatos gyulladáshoz, sejtek és szövetek károsodásához vezet Az
Immunológia alapjai. T-sejt differenciálódás és szelekció a tímuszban: a mikrokörnyezet és szolubilis faktorok szabályozó szerepe
 Immunológia alapjai T-sejt differenciálódás és szelekció a tímuszban: a mikrokörnyezet és szolubilis faktorok szabályozó szerepe A limfocita fejlődés lépései Recirkuláció a periférián Korai érés és növekedési
Immunológia alapjai T-sejt differenciálódás és szelekció a tímuszban: a mikrokörnyezet és szolubilis faktorok szabályozó szerepe A limfocita fejlődés lépései Recirkuláció a periférián Korai érés és növekedési
Környezetegészségtan 2016/2017. Immunológia 1.
 Környezetegészségtan 2016/2017 Immunológia 1. 2016. XI.11. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@freemail.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
Környezetegészségtan 2016/2017 Immunológia 1. 2016. XI.11. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@freemail.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
Hogyan véd és mikor árt immunrendszerünk?
 ERDEI ANNA Hogyan véd és mikor árt immunrendszerünk? Erdei Anna immunológus egyetemi tanár Az immunrendszer legfontosabb szerepe, hogy védelmet nyújt a különbözô kórokozók vírusok, baktériumok, gombák,
ERDEI ANNA Hogyan véd és mikor árt immunrendszerünk? Erdei Anna immunológus egyetemi tanár Az immunrendszer legfontosabb szerepe, hogy védelmet nyújt a különbözô kórokozók vírusok, baktériumok, gombák,
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Igyártó Botond-Zoltán
 A MADARAK ANTIGÉNPREZENTÁLÓ DENDRITIKUS SEJTJEI Doktori tézisek Igyártó Botond-Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Magyar Attila, egyetemi adjunktus, D.V.M.,
A MADARAK ANTIGÉNPREZENTÁLÓ DENDRITIKUS SEJTJEI Doktori tézisek Igyártó Botond-Zoltán Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Magyar Attila, egyetemi adjunktus, D.V.M.,
Túlérzékenységi reakciók Gell és Coombs felosztása szerint.
 Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
Túlérzékenységi reakciók Gell és Coombs felosztása szerint. A felosztás mai szemmel nem a leglogikusabb, mert nem tesz különbséget az allergia, az autoimmunitás és a a transzplantációs immunreakciók között.
Allergia immunológiája 2012.
 Allergia immunológiája 2012. AZ IMMUNVÁLASZ SZEREPLŐI BIOLÓGIAI MEGKÖZELÍTÉS Az immunrendszer A fő ellenfelek /ellenségek/ Limfociták, makrofágok antitestek, stb külső és belső élősködők (fertőzés, daganat)
Allergia immunológiája 2012. AZ IMMUNVÁLASZ SZEREPLŐI BIOLÓGIAI MEGKÖZELÍTÉS Az immunrendszer A fő ellenfelek /ellenségek/ Limfociták, makrofágok antitestek, stb külső és belső élősködők (fertőzés, daganat)
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
EXTRACELLULÁRIS PATOGÉNEK
 EXTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet AZ EXTRACELLULÁRIS BAKTÉRIUMOKKAL SZEMBENI IMMUNVÁLASZOK A bőr és a nyálkahártyák elhatárolják
EXTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet AZ EXTRACELLULÁRIS BAKTÉRIUMOKKAL SZEMBENI IMMUNVÁLASZOK A bőr és a nyálkahártyák elhatárolják
Az immunológia alapjai
 Az immunológia alapjai Kacskovics Imre Eötvös Loránd Tudományegyetem Immunológiai Tanszék Budapest Citokinek Kisméretű, szolubilis proteinek és glikoproteinek. Hírvivő és szabályozó szereppel rendelkeznek.
Az immunológia alapjai Kacskovics Imre Eötvös Loránd Tudományegyetem Immunológiai Tanszék Budapest Citokinek Kisméretű, szolubilis proteinek és glikoproteinek. Hírvivő és szabályozó szereppel rendelkeznek.
Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése.
 F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban
F38347 Dendritikus sejtek differenciálódásának szabályozása hormonreceptorok által című OTKA pályázat záró jelentése. A kutatás időtartama: 2002-2005 Témavezető neve: Dr. Szatmári István Jelen OTKA pályázatunkban
Sejtfelszíni markerek és antigén csoportok
 Sejtfelszíni markerek és antigén csoportok Markerek A sejthártya aszimmetrikus: extracellulárisan oligoszacharidokban gazdag (glikoprotein és glikolipid oldalláncai révén) Sejteket jelölő anyagok markerek
Sejtfelszíni markerek és antigén csoportok Markerek A sejthártya aszimmetrikus: extracellulárisan oligoszacharidokban gazdag (glikoprotein és glikolipid oldalláncai révén) Sejteket jelölő anyagok markerek
Környezetegészségtan 2018/2019. Immunológia 1.
 Környezetegészségtan 2018/2019 Immunológia 1. 2018. XI.12. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@ttk.elte.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
Környezetegészségtan 2018/2019 Immunológia 1. 2018. XI.12. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@ttk.elte.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
Környezetegészségtan 2016/2017. Immunológia 1.
 Környezetegészségtan 2016/2017 Immunológia 1. 2016. XI.11. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@freemail.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
Környezetegészségtan 2016/2017 Immunológia 1. 2016. XI.11. Józsi Mihály ELTE Immunológiai Tanszék http://immunologia.elte.hu email: mihaly.jozsi@freemail.hu Az Immunológia tankönyv elérhető: http://www.tankonyvtar.hu/hu/tartalom/tamop425/2011_0001_524_immunologia/adatok.html
A veleszületett és az adaptív immunválasz áttekintése
 A veleszületett és az adaptív immunválasz áttekintése Erdei Anna ELTE, TTK Biológiai Intézet Immunológiai Tanszék Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február
A veleszületett és az adaptív immunválasz áttekintése Erdei Anna ELTE, TTK Biológiai Intézet Immunológiai Tanszék Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február
A hisztamin és hisztamin H4 receptor hiányának hatása a dendritikus sejtek működésére
 A hisztamin és hisztamin H4 receptor hiányának hatása a dendritikus sejtek működésére Doktori tézisek Jelinek Ivett Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Dr. László
A hisztamin és hisztamin H4 receptor hiányának hatása a dendritikus sejtek működésére Doktori tézisek Jelinek Ivett Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Dr. László
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A szervezet védekező reakciói II. Adaptív/szerzett immunitás Emberi vércsoport rendszerek
 A szervezet védekező reakciói II. Adaptív/szerzett immunitás Emberi vércsoport rendszerek Tanulási támpontok: 19. és 20. Sántha Péter 2017. 10. 09 A veleszületett (természetes) és a szerzett (adaptív)
A szervezet védekező reakciói II. Adaptív/szerzett immunitás Emberi vércsoport rendszerek Tanulási támpontok: 19. és 20. Sántha Péter 2017. 10. 09 A veleszületett (természetes) és a szerzett (adaptív)
3. Az alábbi citokinek közül melyiket NEM szekretálja az aktivált Th sejt? A IFN-γ B interleukin-10 C interleukin-2 D interleukin-1 E interleukin-4
 A Név: Csoportszám: EGYSZERŰ VÁLASZTÁS 1. Mi atlr-5 legfontosabb ligandja? A endospóra B flagellin C poliszacharid tok D DNS E pilus 2. Mi alkotja az ellenanyag antigénkötő helyét? A a H és L láncok konstans
A Név: Csoportszám: EGYSZERŰ VÁLASZTÁS 1. Mi atlr-5 legfontosabb ligandja? A endospóra B flagellin C poliszacharid tok D DNS E pilus 2. Mi alkotja az ellenanyag antigénkötő helyét? A a H és L láncok konstans
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogénekimmunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Regulátoros T sejtek és sejtes környezetük immunmediált gyermekkori gasztroenterológiai kórképekben
 Regulátoros T sejtek és sejtes környezetük immunmediált gyermekkori gasztroenterológiai kórképekben Doktori értekezés Dr. Cseh Áron Semmelweis Egyetem Klinikai Orvostudományok Doktori Iskola Témavezető:
Regulátoros T sejtek és sejtes környezetük immunmediált gyermekkori gasztroenterológiai kórképekben Doktori értekezés Dr. Cseh Áron Semmelweis Egyetem Klinikai Orvostudományok Doktori Iskola Témavezető:
A C3 komplementfehérje hatása humán monocita eredetű dendritikus sejtek funkcióira
 A C3 komplementfehérje hatása humán monocita eredetű dendritikus sejtek funkcióira Sándor Noémi Témavezető: Bajtay Zsuzsa PhD, habilitált adjunktus ELTE TTK Biológiai Intézet Immunológia Tanszék ELTE TTK
A C3 komplementfehérje hatása humán monocita eredetű dendritikus sejtek funkcióira Sándor Noémi Témavezető: Bajtay Zsuzsa PhD, habilitált adjunktus ELTE TTK Biológiai Intézet Immunológia Tanszék ELTE TTK
A vér élettana III. Fehérvérsejtek és az immunrendszer
 A vér élettana III. Fehérvérsejtek és az immunrendszer Prof. Kéri Szabolcs SZTE ÁOK Élettani Intézet, 2015 ELLENSÉG AZ IMMUNOLÓGIÁBAN: - VÍRUS, BAKTÉRIUM, GOMBA, PARAZITÁK - IDEGEN SEJTEK - SAJÁT SEJTEK
A vér élettana III. Fehérvérsejtek és az immunrendszer Prof. Kéri Szabolcs SZTE ÁOK Élettani Intézet, 2015 ELLENSÉG AZ IMMUNOLÓGIÁBAN: - VÍRUS, BAKTÉRIUM, GOMBA, PARAZITÁK - IDEGEN SEJTEK - SAJÁT SEJTEK
Immunkomplexek által elindított gyulladási folyamatok követésére alkalmas mikrofluidikai rendszer fejlesztése
 1 Immunkomplexek által elindított gyulladási folyamatok követésére alkalmas mikrofluidikai rendszer fejlesztése Fürjes Péter MTA Energiatudományi Kutatóközpont MEMS Lab és Papp Krisztián MTA-ELTE Immunológia
1 Immunkomplexek által elindított gyulladási folyamatok követésére alkalmas mikrofluidikai rendszer fejlesztése Fürjes Péter MTA Energiatudományi Kutatóközpont MEMS Lab és Papp Krisztián MTA-ELTE Immunológia
T sejtek II Vizler Csaba 2010
 T sejtek II Vizler Csaba 2010 DENDRITIKUS SEJT Tc CD8+ Th CD4+ B SEJT CTL DTH Ab T sejtek és B sejtek - az elnevezés eredete A T sejt receptor (TcR) kialakulása T sejt érés és szelekció a tímuszban A T
T sejtek II Vizler Csaba 2010 DENDRITIKUS SEJT Tc CD8+ Th CD4+ B SEJT CTL DTH Ab T sejtek és B sejtek - az elnevezés eredete A T sejt receptor (TcR) kialakulása T sejt érés és szelekció a tímuszban A T
ELMÉLETI ÖSSZEFOGLALÓ
 ELMÉLETI ÖSSZEFOGLALÓ Előzetes ismeretek: a sejt felépítése sejtalkotók szerepe a sejtmembrán szerkezete sejtfelszíni molekulák szerepe (marker-receptor) fehérjeszintézis alapja, folyamata Megjegyzés:
ELMÉLETI ÖSSZEFOGLALÓ Előzetes ismeretek: a sejt felépítése sejtalkotók szerepe a sejtmembrán szerkezete sejtfelszíni molekulák szerepe (marker-receptor) fehérjeszintézis alapja, folyamata Megjegyzés:
A hízósejtek szerepe az immunológiai folyamatokban
 A hízósejtek szerepe az immunológiai folyamatokban Berki Timea Boldizsár F, Bartis D, Talabér G, Szabó M, Németh P, University of Pécs, Department of Immunology & Biotechnology, Pécs, Hungary Additon of
A hízósejtek szerepe az immunológiai folyamatokban Berki Timea Boldizsár F, Bartis D, Talabér G, Szabó M, Németh P, University of Pécs, Department of Immunology & Biotechnology, Pécs, Hungary Additon of
Az immunrendszer alapjai, sejtöregedés, tumorképződés. Biológiai alapismeretek
 Az immunrendszer alapjai, sejtöregedés, tumorképződés Biológiai alapismeretek Az immunrendszer Immunis (latin szó): jelentése mentes valamitől Feladata: a szervezetbe került idegen anyagok: 1. megtalálása
Az immunrendszer alapjai, sejtöregedés, tumorképződés Biológiai alapismeretek Az immunrendszer Immunis (latin szó): jelentése mentes valamitől Feladata: a szervezetbe került idegen anyagok: 1. megtalálása
Doktori értekezés tézisei
 Doktori értekezés tézisei Az FcRn overexpresszió hatása a humorális immunválaszra Dr. Schneider Zita Témavezető: Dr. Kacskovics Imre ELTE TTK Biológiai Doktori Iskola Doktori Iskola Vezető: Prof. Dr. Erdei
Doktori értekezés tézisei Az FcRn overexpresszió hatása a humorális immunválaszra Dr. Schneider Zita Témavezető: Dr. Kacskovics Imre ELTE TTK Biológiai Doktori Iskola Doktori Iskola Vezető: Prof. Dr. Erdei
Immunológiai alapfogalmak. Immunrendszer felépítése
 Immunológiai alapfogalmak. Immunrendszer felépítése Immunsejtek: Immunválaszban szereplő legfőbb sejtek: Limfociták: Csontvelői eredetű erősen festődő heterokromatinnal rendelkező sejtek. Antigénreceptorok
Immunológiai alapfogalmak. Immunrendszer felépítése Immunsejtek: Immunválaszban szereplő legfőbb sejtek: Limfociták: Csontvelői eredetű erősen festődő heterokromatinnal rendelkező sejtek. Antigénreceptorok
A sejtfelszíni FasL és szolubilis vezikulakötött FasL által indukált sejthalál gátlása és jellemzése
 A sejtfelszíni FasL és szolubilis vezikulakötött FasL által indukált sejthalál gátlása és jellemzése Doktori értekezés tézisei Hancz Anikó Témavezetők: Prof. Dr. Sármay Gabriella Dr. Koncz Gábor Biológia
A sejtfelszíni FasL és szolubilis vezikulakötött FasL által indukált sejthalál gátlása és jellemzése Doktori értekezés tézisei Hancz Anikó Témavezetők: Prof. Dr. Sármay Gabriella Dr. Koncz Gábor Biológia
Intézeti Beszámoló. Dr. Kovács Árpád Ferenc
 Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Allergológia Kurzus 2011
 Allergológia Kurzus 2011 Kedd, 14.00-15.30 1 kreditpont Hepreszenzitivitás A normális immunrendszer által adott nemkívánatos reakció Ezek a reakciók szövetkárosodást, kellemetlen tüneteket okoznak, ritkán
Allergológia Kurzus 2011 Kedd, 14.00-15.30 1 kreditpont Hepreszenzitivitás A normális immunrendszer által adott nemkívánatos reakció Ezek a reakciók szövetkárosodást, kellemetlen tüneteket okoznak, ritkán
Kacsa IMMUNOLÓGIA. A jobb megértés alapjai. S. Lemiere, F.X. Le Gros May Immunrendszer. Saját, veleszületett immunitás. Szerzett immunitás
 Kacsa IMMUNOLÓGIA Immunrendszer Saját, veleszületett immunitás A jobb megértés alapjai Szerzett immunitás S. Lemiere, F.X. Le Gros May 2016 Háziasított viziszárnyasok Cairina moschata dom. Anas platyrhyncos
Kacsa IMMUNOLÓGIA Immunrendszer Saját, veleszületett immunitás A jobb megértés alapjai Szerzett immunitás S. Lemiere, F.X. Le Gros May 2016 Háziasított viziszárnyasok Cairina moschata dom. Anas platyrhyncos
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
A veleszületett (természetes) immunrendszer. PAMPs = pathogen-associated molecular patterns. A fajspecifikus szignálok hiányának felismerése
 immunrendszer. PAMPs = pathogen-associated molecular patterns. A fajspecifikus szignálok hiányának felismerése") A veleszületett (természetes) immunrendszer PAMPs = pathogen-associated molecular patterns PRRs = pattern recognition receptors A fajspecifikus szignálok hiányának felismerése Eukariota sejtmembrán Az
A veleszületett (természetes) immunrendszer PAMPs = pathogen-associated molecular patterns PRRs = pattern recognition receptors A fajspecifikus szignálok hiányának felismerése Eukariota sejtmembrán Az
A veleszületett és az adaptív immunválasz áttekintése
 A veleszületett és az adaptív immunválasz áttekintése Erdei Anna ELTE, TTK Biológiai Intézet Immunológiai Tanszék Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2017. január
A veleszületett és az adaptív immunválasz áttekintése Erdei Anna ELTE, TTK Biológiai Intézet Immunológiai Tanszék Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2017. január