A nukleinsavkémiai kisszótár: aminoacil-trns szintetáz: adott aminosavat a megfelelő trns-hez kapcsoló enzim DNS: dezoxiribonukleinsav, DNA <ang.
|
|
|
- Nóra Gáspár
- 5 évvel ezelőtt
- Látták:
Átírás




1 1
2 A nukleinsavkémiai kisszótár: aminoacil-trs szintetáz: adott aminosavat a megfelelő trs-hez kapcsoló enzim DS: dezoxiribonukleinsav, DA <ang.>: olyan nukleotidegységekből felépülő nukleinsavak gyűjtőneve, amelyek dezoxiribóz cukorrészt tartalmaznak. A gének kódolására, és továbbadására szolgálnak. RS: ribonukleinsav, RA <ang.>: olyan nukleotid egységekből felépülő nukleinsav, amely cukorrészként ribózt tartalmaz, előfordul minden élő sejtben, valamint egyes vírusokban. ukleotidok: (= bázis + cukor + foszfát) a nukleozidok foszforsav-észtereinek gyűjtőneve. A bennük szereplő cukorrész (D-ribóz v. 2-dezoxi-D-ribóz) alapján megkülönböztetik a ribonukleotidok és a dezoxiribonukleotidok csoportját. ukleozidok: (= bázis + cukor) szűkebb értelemben a nukleinsavakban előforduló pirimidin- és purinbázisok -ribozid, ill. -2 -dezoxiribozid típusú glikozidjainak gyűjtőneve (citidin, uridin, timidin, adenozin, guanozin). A ~okban a cukorrész mindig furanóz szerekezetű, az -glikozidos szénatom pedig -konfigurációjú. ukleinsavak (DS, RS): <lat. nucleus mag >: (= bázis+cukor+foszfát -> polimer) nukleotidokból felépülő óriásmolekulák (biopolimerek), amelyek minden sejtben és a virusokban is megtalálhatók, azoknak esszenciális alkotórészei. ~akat először F. Miescher (1869) különített el a genny fehérvérsejtjeiből. Watson Crick-modell: a DS térszerkezetét leíró térszerkezeti modell. A ~ szerint a DS-molekula kettős hélixét két, ellentétes lefutású (antiparalel) polinukleotid-lánc alkotja. A két láncot a komplementer bázispárok (adenin timin, guanin citozin) között létrejött hidrogénkötések tartják össze. A kettőshélix-szerkezetet J. D. Watson és F. Crick javasolta 1953-ban. A ~ szerint a polinukleotid-láncokat helikális szerkezetű cukorfoszfátváz építi fel; a hélix tengelyére merőleges irányban, a hélix belseje felé helyezkednek el a nukleinsavbázisok. A ~ alapján jól magyarázható a genetikai információ megőrzése és átadása. DS kettős hélix/spirál: Watson Crick-modell DS-replikáció: A DS mindkét szálának megduplázódása, amely során egy DS kettős spirálból létrejön kettő, az eredetivel azonos DS kettős spirál. Transzkripció: Atírás (DS->RS) amely során a DS-függő RS polimeráz a rendelkezésére álló nukleozid 5 -trifoszfátokból RS-t hoz létre. Fontosabb lépései: iniciáció, elongáció és termináció. Transzláció: Fehérje bioszintézis (RS->Fehérje), amely során a riboszóma az RS alapján, a rendelkezésre 2 álló (és megfelelő aminosavat hordozó) trs-ek segítségével megszintetizálja az adott fehérjét.
3 Mutáció: Az eredetihez képesti elváltozás a DS-ben (más vagy hiányzó nukleotid(ok)) Riboszóma: A fehérjeszintézist = transzlációt végző ribozim, mely 2 rrs és több fehérjéből áll. Ribozim: sejtszervecske, ribonukleo-protein komplex: egy bizonyos enzimatikus feladatot ellátó (egy bizonyos reakciót katalizáló) RS. Enzim: Egy bizonyos enzimatikus feladatot ellátó (bizonyos reakciót katalizáló) fehérje. mrs: hírvivő vagy messenger RS: átmeneti adathordozóként szolgál (DS -> mrs -> Fehérje) feladata: a fehérje aminosavsorrendjét a genetikai kód által meghatározó RS, amely a megfelelő DS-ről szintetizálódik és a riboszóma fordítja le ezt az információt fehérjévé. rrs: riboszomális RS: a riboszóma fő alkotóeleme, a katalitikusan aktív része. trs: átvivő vagy transzfer RS: Antikodont és az annak megfelelő aminosavat (ill. kötőhelyét) tartalmazó RS. Az aminosavakat a riboszómához szállítja, az mrs-en lévő kodonokat antikodonja segítségével ismeri fel (bázispárosodás elve) aminoacil-trs szintetáz: adott aminosavat a megfelelő trs-hez kapcsoló enzim genetikai kód: a nukleinsavszekvenciák aminosavszekvenciára fordítását meghatározó szabályrendszer, az mrs 3 bázisa alkot egy kodont, ami egy aminosavat határoz meg oly módon, hogy adott antikodonú trs ismeri fel a transzláció során Kodon: ukleotid-hármas a DS/RS-en, mely egy-egy aminosavat kódol Kromoszóma: (a görög chroma=színes és soma=test) egyetlen hosszú DS, amely számos gént, szabályozó és egyéb szekvenciákat tartalmaz. Kromatin: a kromoszóma anyaga, mely eukarióta sejtekben DS (rendszerint kettős szálú) és fehérje komplexe. (Festhető, innen a neve.) Antikodon: ukleotid-hármas a trs-en, mely reverz-komplementje a kodonnak (tehát Watson- Crick párosítással illeszkedik a kodonhoz transzláció közben) Gén: Az öröklődő DS egy fehérjét kódoló része. genetikai kód: a nukleinsavszekvenciák aminosavszekvenciára fordítását meghatározó szabályrendszer, ahol az mrs 3 bázisa alkot egy kodont, amely egy fehérjeépítő aminosavat határoz meg oly módon, hogy azt a transzláció során egy adott antikodonú trs felismeri. 3
, mely(ek) tényleges fehérjeszekvenciát kódól(nak) Intron: Egy gén/mrs azon része(i), mely(ek) nem kódolnak fehérjeszekvenciát, és az RS érés során (transzkripció és")
4 Genom: A gének összesége (egy egyedben/fajban), pl. a humán (emberi) genom. Exon: Egy gén/mrs azon része(i), mely(ek) tényleges fehérjeszekvenciát kódól(nak) Intron: Egy gén/mrs azon része(i), mely(ek) nem kódolnak fehérjeszekvenciát, és az RS érés során (transzkripció és transzláció között) kivágódnak, pre-mrs -> m-rs. Restrikciós endonukleáz: DS-hasító enzim, mely csak egy meghatározott szekvencia egy meghatározott pontján hasítja a DS-t. Palindrom szekvencia: DS szekvencia, mely megegyezik a reverz-komplementjével, pl. AATT, GGATCC, stb. Antiszensz oligonukleotid: A kódoló DS szálhoz kapcsolódó oligonukleotid, amely speciális szakaszhoz kötődve gátolhatja a DS átíródását (transzkipciót). PCR: Polimerase-Chain-Reaction: polimeráz-láncreakció, egy eljárás a sejten-kívüli (in vitro) DS sokszorosítására. (templát DS + primer + polimeráz + nukleotidok(monomerek) + hőmérsékletváltakozás = sokszorozódás) RS- polimeráz: egy enzim amely széttekeri a DS kettős spirált Promoter: a DS-nek az a része ahova az RS-polimeráz kötődik és ahol megkezdi a transzkripciót riboszóma: a fehérjeszintézist végző sejtszervecske, egy ribonukleoprotein komplex RS nukleotidok, mint építőelemek Terminátor: prokarióták azon DS része ami jezi a transzkripció végét Transzkripciós egység: Az a DS darab ami RS-é átíródik. Transzkripcós factorok: az eukarióta sejtek azon fehérjéi amelyek az RS-polimeráz kötését, illetve a transzkripció megkezdését elősegítik. Transzkripciós Iniciációs Komplex: A transzkripciós faktorok és a promoter régióhoz kapcsolódó RS-polimeráz II együttese. TATA Box: A DS promoter része 4


 1957")




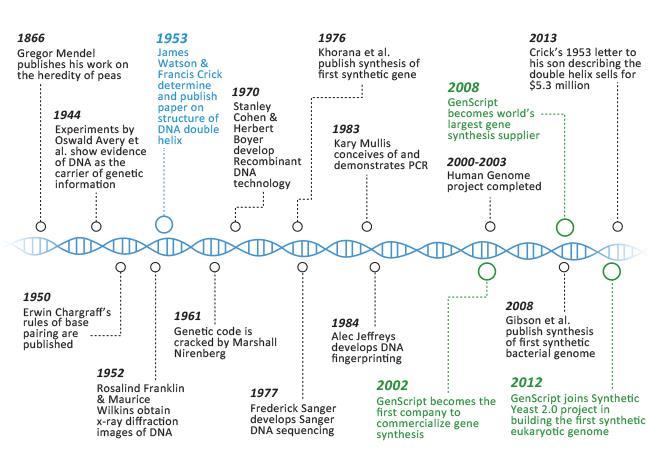
5 A nukleinsavkémia koronázatlan királyai: Johannes Friedrich Miescher svájci orvos-kémikus 1869-ban felfedezi és izolálja a DS-t. Lord Alexander R. Todd (angol biokémikus) 1957 obel-díj A nukleotidok és a nukleotid koenzimek felfedezéséért Francis arry Compton Crick James Dewey Watson 1962 obel-díj a DS molekuláris szerkezetének felismeréséért Maurice ugh Frederick Wilkins Picture 51 Rosalind Franklin 5 swald T. Avery (DS a kromószóma anyaga)



 1970 obel-díj a cukor nukleotidok")
 1982")

6 A nukleinsavkémia koronázatlan királyai, kémiai és orvosi obel-díjak: Stanford Moore és William. Stein amerikai biokémikusok 1972 obel-díj a ribonukleáz aktív centrumának katalitikus aktivitása és kémiai szerkezete közötti kapcsolat feltárásáért 1980 Paul Berg Walter Gilbert rekombináns-ds Frederick Sanger A nukleinsavak szekvenálásáért Luis Federico Leloir (argentín biokkémikus) 1970 obel-díj a cukor nukleotidok felfedezéséért és a szénhidrátok bioszintézise kapcsán elért eredményeiért Aaron Klug (angol kémikus ) 1982 obel-díj a krisztallográfiai elektronmikroszkóp kifejlesztéséért illetve a bilológiai szempontból fontos nukleinsavfehérjekomplexek 6 szerkezetfelderítéséért







 Michael Smith")

7 Sir James W. Black Gertrude B. Elion George. itchings 1988 obel-díj a purin alapú kemoterápiás gyógyszerek kifejlesztéséért Sidney Altman és Thomas R. Cech Amerikai/kanadai és amerikai kémikus 1989 obel-díj az RS katalitikus tulajdonságainak felfedezéséért. Kary B. Mullis 1993 obel-díj Polimeráz láncreakció (PCR) Michael Smith Irányított mutagenezis Venkatraman Ramakrishnan Thomas Steitz 2009 obel-díj a riboszóma szerkezetének és funkciójának tanulmányozásáért Ada Yonath 7



8 Tomas Lindahl, Paul Modrich és Aziz Sancar 2015 obel-díj A DS-javítás mechanizmusának tanulmányozásáért
9 9
10 ukleinsavak: 1) A kromoszóma és felépítése 2) DS- RS építőelemek: ukleozidok, nukleotidok, nukleinsavak em-rs és -DS alkotó purinszármazékok A DS térszerkezete, a bázispárok 3) DS nanorendszerek 4) DS (RS) reaktivitása: hidrolízise az építőelemek kémiai szintézise kémiai (szilárdfázisú) szintézis enzimatikus szintézis: a replikáció 5) Transzkripció 6) Transzláció 7) DS analitika: szekvenálás 8) Bioenergetika 10

")


11 1) A kromoszóma és felépítése A prokarióták, körülhatárolt sejtmag nélküli élőlények, tipikusan egysejtűek, ilyenek az archeák és a baktériumok Az eukarióták olyan élőlények, amelyek valódi sejtmaggal (a mag anyagát a citoplazmától egy maghártya választja el) rendelkező sejtekből épülnek fel (eu = valódi, karyon = sejtmag). Mind a prokatiota-, mind az eukariótasejtekben az információtárolásra a DS hivatott. Escherichia coli baktérium papucsállatka Egy eukarióta sejt genetikai anyagának többsége 11 a sejtmagban, kromoszómák formájában van tárolva.

 diploid (két kromoszómakészlet; 44+2) vagy poliploid (több készlet;")
 testi, mind 2) haploid (egy készlet; 22+1) ivari sejtekkel rendelkeznek.")
12 Fajfüggő az eukarióta sejtek kromoszómáinak a száma: rozs 14 galamb 16 giliszta 36 mezei nyúl 46 ember 46 csimpánz 48 kutya 78 ponty 104 pillangó 380 páfrány 1200 Egy férfi diploid sejt kariogramja: 22*2 testi kromoszóma = 44 2 nemi kromoszóma: X és Y = 2 Σ= 46 A szexuális úton szaporodó fajok mind 1) diploid (két kromoszómakészlet; 44+2) vagy poliploid (több készlet; triploid magnélküli dinnye, stb.) testi, mind 2) haploid (egy készlet; 22+1) ivari sejtekkel rendelkeznek. Egy női diploid sejt kariogramja: 22*2 testi kromoszóma = 44 2 nemi kromoszóma: X + X = 2 Σ= 46

 231 50 961 097 összesen = 46 ~30.")
13 Egy női diploid sejt kariogramja: 22*2 testi kromoszóma = 44 2 nemi kromoszóma (X és X) = 2 örökségünk : minden diploid sejt kromoszóma-párja egy anyai és egy apai kromoszómát tartalmaz apai anyai kromoszóma gének száma bázispárok száma X (ivari kromoszóma) Y (ivari kromoszóma) összesen = 46 ~ ~3, ,2 gigabyte-nyi betűt A biblia 3,5 millió karakter aza kb 800 biblia a genetikai kód 13


 - G")

14 Egy eukarióta sejtciklusa: a sejtosztódás lépései: A G 0 fázisban a sejtek változatlan állapotba kerülnek, várva a sejtciklusba vezető jelre. - G 1 fázis: első növekedési szakasz, - S fázis: a DS replikálódik (46-ból 92 kromószóma lesz) - G 2 fázis: második növekedési szakasz, és felkészülés az osztódásra, - M fázis (vagy mitózis + citokinezis): a sejt két utódsejtre válik szét replikáció 14


15 0,2-2 μm A kromoszómapár hierarchikus szerkezeti elrendeződése. A feladat: az eukarióta sejtek mintegy 3.2 milliárdnyi bázispárját feltekerve, azt néhány mikrométerbe tömöríteni. histon
 A")

 az egészet a -hidak")

16 centromer kromatin telomer régiók megkettőződött kromoszóma a metafázisban A kódolo DS aránya a teljes genomhoz viszonyított (%) A supercoiling szeres kondenzációt tesz lehetővé a nukleoszóma a 8 histon fehérjét körülölelő 146 bázispárnyi DS-ből áll. A komplexet a 1 linker fehérjék stabilizálják. mindig jobb menetes, antiparallel, (kivéve a Z-DS) az egészet a -hidak tartják egyben. Eukarióta sejtek génjében van nemkódoló rész; intronok. (prokarióta sejtekben tipikusan nincs intron.) Az intron a pre-mrs-be átíródik, majd az RS érés során az kivágódik. 16
 Giemsa-festés: a bázikus festék a P- gerinchez kötődik, de különösen az A-T bázispárban gazdag régiókhoz (A-T résznél sötétebb a szín) 2) Quinacrine-festés fluoreszcens festés")
17 Kromoszóma: (a görög chroma=színes és soma=test) egyetlen hosszú DS, amely számos gént, szabályozó és egyéb szekvenciákat tartalmaz. 1) Giemsa-festés: a bázikus festék a P- gerinchez kötődik, de különösen az A-T bázispárban gazdag régiókhoz (A-T résznél sötétebb a szín) 2) Quinacrine-festés fluoreszcens festés Gustav Giemsa ( ), német mikrobiológus 17
 és átfordítására (transzláció) alkalmasak, aminek köszönhetően a sejt összes fehérjéje meg tud")
18 2) DS- RS építőelemek memo: nucleus = sejtmag A nukleinsavak két fő fajtája a - dezoxiribonukleinsavak (DS) - ribonukleinsavak (RS) - savanyú jellegű (híg lúgban oldódó), nagy molekulasúlyú polimer vegyület - molekulák, amelyek az átörökítendő információ tárolására, annak átírására (transzkripció) és átfordítására (transzláció) alkalmasak, aminek köszönhetően a sejt összes fehérjéje meg tud szintetizálódni. (Alkalmasak továbbá reakciók katalízisére is.) Kémiai építőelemek: nukleotidok és nukleozidok P 4' 5' 3' 2' heterociklusos bázis --glikozidos kötés 1' egy nukleotid hidrolízis heterociklusos bázis: purin vagy pirimidin ötszénatomos monoszacharid: D-ribóz vagy 2-dezoxi-D-ribóz foszfátion 18
 + 3 P 4 purin nukleozid foszforiláz* + ukleotidbázis + pentóz (nucleobase) *")
19 Foszforsavészterek hidrolízise: ukleinsav (polinukleotid) nukleáz enyhe degradáció 1 a, 25 C ukleotid (monomer) 2 nukleotidáz cc. 3, 180 C Q= Q= A -glikozidkötés hidrolízise: Q adenozin dezoxiadenozin ukleozid (glikozid) + 3 P 4 purin nukleozid foszforiláz* + ukleotidbázis + pentóz (nucleobase) * ez a foszforiláz valójában egy hidroláz
20 ukleinsav építőelemek; a D-ribóz D-ribóz D-arabinóz D-xilóz D-lixóz memo: pirán furán tetrahidro-2 -pirán tetrahidrofurán Ciklusos félacetál képződése, avagy hogyan rajzoljuk a furanózokat: C C C C C 2 A nyílt láncú D-fruktóz különbözö konformációi 4 C 3 C 5 C 2 nyílt láncú D-ribóz 2 C C 1 C C 3 C 2 C C nyílt láncú D-ribóz C 2 C1 4 C C C -D-ribofuranóz C 2 C C C 1 C -D-ribofuranóz 2 ukleozidokban mindig 5 tagú (furanóz) gyűrű formájában van a ribóz vagy dezoxi-ribóz jelen. 20
21 ukleinsav építőelemek; a heterociklus Aromás vegyületek: két heteroatomos hattagú gyűrűk (Bruckner. III/1.) em aromás származékok memo: gyenge bázisok: a piperazin 10%-os vizes oldatának p-ja
22 a barbitursav fontosabb tautomerjei Pirimidinszármazékok, nukleinsav építőelemek; Az uracil keto-enol tautomer egyensúlyi formái: memo: csak a laktimlaktám vagy a dilaktám forma jöhet szóba, az -glikozid kötés miatt. 22
23 ukleinsav építőelemek; purinszármazékok A guanin keto-enol tautomer egyensúlyi formái: Melyik tautomer forma a stabilabb? 23
24 2 2 adenozin adenozin 9-( -D-ribofuranozil)-adenin op= 235 o C RS építőelem 2 2 adenozid - P - adenozin-5'-foszfát 5'-AMP P - - adenozin-3'-foszfát 3'-AMP dezoxiadenozin 2 2 dezoxiadenozin 9-(2'-dezoxi- -D-ribofuranozil)-adenin op= 190 o C 2 2 dezoxiadenozid DS építőelem - P - dezoxiadenozin-5'-foszfát 5'-dAMP P dezoxiadenozin-3'-foszfát 3'-dAMP
25 guanozin guanozin 9-( -D-ribof uranozil)-guanin op= 240 o C 2 2 RS építőelem guanozid P - guanozin-5'-foszfát 5'-GMP P - - guanozin-3'-foszfát 3'-GMP dezoxiguanozin 2 2 dezoxiguanozin 9-(2'-dezoxi- -D-ribofuranozil)-guanin DS építőelem dezoxiguanozid - P - dezoxiguanozin-5'-foszfát 5'-dGMP 2 2 P dezoxiguanozin-3'-foszfát 3'-dGMP
26 2 citidin 2 RS építőelem citidin 3-( -D-ribof uranozil)-citozin op= 230 o C citidinf oszfátok P - citidin-5'-foszfát 5'-CMP P - - citidin-3'-foszfát 3'-CMP dozoxicitidin 2 2 dezoxicitidin 3-(2'-dezoxi- -D-ribofuranozil)-citozin dezoxicitidinfoszfátok 2 2 DS építőelem - P - dezoxicitidin-5'-foszfát 5'-dCMP P dezoxicitidin-3'-foszfát 3'-dCMP
27 RS építőelem timidin 3 C timidin 3-(2'-dezoxi- -D-ribofuranozil)-timin 3 C op= 183 o timidinfoszfátok 3 C 3 C DS építőelem - P - timidin-5'-foszfát 5'-TMP P timidin-3'-foszfát 3'-TMP


28 em-rs és -DS alkotó purinszármazékok: A purin degradációs út: PPáz:= Purin nukleozid foszforiláz AMP nukleotidáz GMP Adenozin dezamináz PPáz memo: purinvázas alkaloidok: di- és trimetil xantinok: 28
29 A DS kémiai szerkezete foszforsavdiészter típusú, nem-elágazó, lineáris polimer, amit az 5 3 irányba írunk:..-5 -C-G-T-A Az RS kémiai szerkezete
 (G+A)/(C+T) 1 és A/T 1 és G/C 1 memo: (G+C)/(A+T) változik: ez adja a DS termostabilitását (lásd később) 5 Erwin Chargaff")
30 A Chargaff-szabály megadja a nukleotidok belső arányait a DS-ben: Σ(pur.)/Σ(pir.) (G+A)/(C+T) 1 és A/T 1 és G/C 1 memo: (G+C)/(A+T) változik: ez adja a DS termostabilitását (lásd később) 5 Erwin Chargaff P P P P 3 C G T A T A C G 3 P P P P 5 30



31 A DS térszerkezete, a bázispárok A DS jobbmenetes kettős hélix téralkatú, körülbelül 10 nukleotidpárral hélix menetenként. A spirálokat -hidak tartják össze, az egyik a kódoló a másik a templát szál. Az adenint és a timint 2, míg a guanint és a citozint 3- híd köti össze. Ezt a téralkatot először James Watson and Francis Crick becsülték (határozták) meg helyesen 1953-ban Rosalind Franklin diffrakciós méréseinek köszönhetően. 31
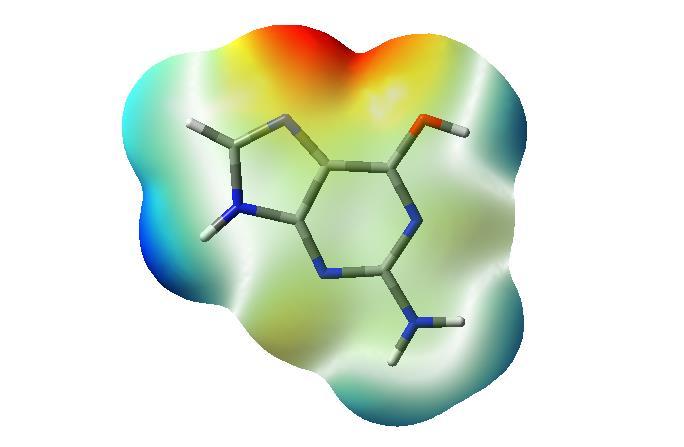
32 oogsteen párképzés Watson-Crick párképzés A DS térszerkezete, a bázispárok interface: kedvező elektrosztatikus kcs. parciális töltés színkódja: d- d+ G C A T interface: kedvezőtlen elektrosztatikus kcs. interface: kedvező elektrosztatikus kcs. 32
33 Watson-Crick oogsteen 33
34 Watson-Crick oogsteen 34


 DS elektronsörűségeloszlása Rosalind Franklin A kettős hélix (double helix, duplex)")
35 Watson-Crick párképzés: fontos, ez található elsősorban a duplexekben G is C A amino (aromás) míg T dilaktám forma is amino-laktám forma - a hidrofil és negatív töltésű foszfodezoxiribóz gerinc kifelé mutat - a bázisok befelé állnak, intermolekuláris -hidakkal stabilizáltak, - a cukor-foszforsav diészter gerinc teljesen konzervatív, míg a heterociklusok permutálódnak, - a bázispárok miatt a DS két szála komplementer jellegű - a heterociklusok pontos szekvenciája hordozza az információt. krisztallográfia (1952) DS elektronsörűségeloszlása Rosalind Franklin A kettős hélix (double helix, duplex) 1953 Francis arry James 35 Dewey Compton Crick Watson
36 Mutáció I: ormális és abnormális bázispár képzés Replikáció #1 Replikáció #2 -G- -Cszülő gyerek unoka -G- -C- -G- -C- mutáció -G*- -T- memo: ugyanúgy megvan a 3. -híd is, ám míg a normál esetben (felső) a purin bázis a 3. - hídban az akceptor, addig a mutációra vezető esetben ugyanitt a -híd donor szerepét tölti be. A két 36 tautomer eltérő -híd mintázatú, s ezért eltérő partnerrel (pirimidin bázissal) képez WC párt. -G- -C- -G- -C- -G- -C- -G- -C- -G- -C- -A- -T-

37 37
38 Mutáció II: a kémiai reagensek kiváltotta mutációk: Pl. salétromossav (a 2 + Cl) magyarázat: mutáció -*- -G- -C- -C- -A- -A- -T- -T- gyerek unoka -A- -A- -T- -T- indukált mutáció: memo: ugyanúgy megvan a 2. -híd, ám míg a normál esetben ( =C) a purin bázis a 2. -hídban donor, addig a mutációra vezető esetben az akceptor (C= ) szerepét tölti be. A két purin származék eltérő -híd mintázatú, s ezért eltérő partnerrel (pirimidin bázissal) képez WC párt. -A- -T- -A- -Tszülő -A- -T- -A- -T- 38
, akkor megjelenhet az A-forma.")
39 A DS térszerkezete, a bázispárok A leggyakoribb forma a B-DS, amely jobbmenetes, a két szál antiparallel elhelyezkedésű 3 nagyárok kisárok 5 P P P P A C C C G G G T P P P P 5 memo: Legtöbb DS téralkata a B-forma amikor vízzel egyensúlyt tart a molekula. a lecsökken a víz tenziója (pl. tömény acl oldattal tart egyensúlyt a DS, s ezért kisebb a tenzió), akkor megjelenhet az A-forma. kérdés: mi lehet a helyzet a kromoszómába becsomagolt DS esetén? 39 memo: Az A olyan mint a B-forma, csak tömörebb. 3
.")
40 jobbmenetes hélix jobbmenetes hélix Az A-DS hélix tömzsibb és rövidebb, mint a B-DS, a főtengelyhez képest a bázispárok síkja döntött ( képest 19 o ). R C-3' a sík felett C-3' endo balmenetes hélix Az Z-DS hélix karcsúbb, főtengelyhez képest a bázispárok síkja döntött ( képest 9 o ). Az B-DS hélixben a főtengelyhez képest a bázispárok síkja merőleges ( képest 1 o ). R 40 C-2' a sík felett C-2' endo
 összesen mintegy 3,2 milliárd bázispárból épül fel: 3,2 10 9 * 3.")
41 A DS 3 fő formájának szerkezeti jellemzői: kérdés: milyen hosszú az emberi genom (kihúzva és egyesítve )? válasz: a teljes genom (44 +2 kromoszóma) összesen mintegy 3,2 milliárd bázispárból épül fel: 3, * m = 1, 06 m gemetriai jellemzők: A-DS B-DS Z-DS hélix típús jobbmenetes jobbmenetes balmenetes ismétlödő egység 1 bp 1 bp 2 bp bp-onkénti elcsvarodás /2 1 menetre eső bp száma főtengelytől vett eltérés emelkedés/bp 2.3 Å (0.23 nm) 3.32 Å (0.332 nm) 3.8 Å (0.38 nm) menethossz 28.2 Å (2.82 nm) 33.2 Å (3.32 nm) 45.6 Å (4.56 nm) átmérő 23 Å (2.3 nm) 20 Å (2.0 nm) 18 Å (1.8 nm) 41
 kérdés: mennyi a T m -pont ha GC = 0,4, de teszünk mellé acl?")
42 A DS hőstabilitása / denaturálása tétel: A DS T m -pontja nő a GC-frakció növekedésével; T m1 < T m2 ha GC 1 /total<gc 2 /total! indoklás: a GC-ben 3, míg az AT-párban csak 2 - híd van bázispáronként. kérdés: mennyi a T m -pont ha GC= 0,4? válasz: T m 340K ha c(acl= 0) kérdés: mennyi a T m -pont ha GC = 0,4, de teszünk mellé acl? válasz:t m 360 K ha c(acl= 150 mmol) kérdés: miért nő a T m -pont ugyanolyan GC frakció mellett akkor, ha magasabb az oldat só koncentrációja? válasz: a DS felszíni negatív töltései P- kompenzálódnak. megjegyzés: - Minnél több a belső -híd a DS 2 szála között az annál stabilabb dimert képez. (Fehérjék esetében ez nem igaz, ott a belső -hidakkal nem nő lineárisan a fehérje termostabilitása!) - A DS T m (60-80 o C) pontja lényegesen magasabb mint a testhőmérsékletünk, és jelentősen magasabb mint a legtöbb fehérjénkké! Atkins de Paula 118 Tm (K) ribóz citozin oxo-f orma guanin oxo-f orma 0 acl 150mM acl Lineáris (0 acl) Lineáris (150mM acl) ribóz 3 C ribóz timin oxo-f orma adenin 42 ribóz DS termostabilitás y = 39,7x R 2 = 0,996 y = 39,7x R 2 = 0,996 0,3 0,4 0,5 0,6 0,7 0,8 GC- frakció A-T gazdag régió memo: - magas p - magas T (K)
 4xG: négy guanidin helyezkedik el egy síkban, egy kation + 4x 2 hidrogénhíd (a purinváz 7-es -jével")
43 em-klasszikus DS szerkezetek: triplex DS: háromszálú DS quadruplex DS: négyszálú DS előfordul: pl. telomerek (kromoszómavégek) 4xG: négy guanidin helyezkedik el egy síkban, egy kation + 4x 2 hidrogénhíd (a purinváz 7-es -jével is) stabilizálja a szerkezetet. 43
44 ukleinsavak: 1) A kromoszóma és felépítése 2) DS- RS építőelemek: ukleozidok, nukleotidok, nukleinsavak em-rs és -DS alkotó purinszármazékok A DS térszerkezete, a bázispárok 3) DS nanorendszerek 4) DS (RS) reaktivitása: hidrolízise az építőelemek kémiai szintézise kémiai (szilárdfázisú) szintézis enzimatikus szintézis: a replikáció 5) Transzkripció 6) Transzláció 7) DS analitika: szekvenálás 8) Bioenergetika 44
 vegyünk egy tompa végű DS kettős hélixet: 5 3 5 180 3 helikáz 5 5 3 3 olliday, R. (1964). A mechanism for gene conversion in fungi. Genetical Research, 5, 282 304.")
45 ) DS nanorendszerek I: olliday-junction, -elágazás ( kocka csúcsa ) hogyan csináljunk ilyen alakzatot? 1) vegyünk egy tompa végű DS kettős hélixet: helikáz olliday, R. (1964). A mechanism for gene conversion in fungi. Genetical Research, 5, azonos helikáz TCTGCACATA AGACGTGTAT memo: a kétszálú hélix és a kereszt forma egyensúlyban lehet, (DS!) 45

 Készíthetünk olyan DS-t")


 12 AT=2 6*(AT =2)")
46 ) A DS két szála nem feltétlenül egyforma hosszú: ilyenkor van túlnyúló szakasz (3 overhang, 5 overhang): Az ilyen DS is továbbalakítható: 3) Készíthetünk olyan DS-t melynek 1) egy részében a két szál egymásnak nem komplementere, a szétszedéshez tehát nem kell a helikáz,valamint 2) amelynek mindkét oldalán van egy-egy túlnyúló szakasz: 5 3 Az ilyen DS is továbbalakítható: 5 Az összes -hidak száma: CG =3 4*(CG =3) 12 AT=2 6*(AT =2) *(CG =3) 30 10*(AT =2) 20 46
 barna-barna (a 8 -híd összetartja majd a ligáz összeköti) 4.2) lila-lila (a 12 -híd összetartja majd a ligáz összeköti) 4.")
47 4) Készítsünk olyan DS szálakat, ahol a túlnyúló szakaszok szekvenciáit alkalmasan választjuk meg: barna-tompavég barna-hegyesvég 4.1) barna-barna (a 8 -híd összetartja majd a ligáz összeköti) 4.2) lila-lila (a 12 -híd összetartja majd a ligáz összeköti) 4.3) fekete-fekete (a 10 -híd összetartja, ligáz összeköti) 47
48 5) Készítsünk ennek fényében olyan DS szálakat amelyek spontán módon, s a túlnyúló szakaszaik révén egyértelműen asszociálódnak: 6) Készíthetünk ilyen módon kockát, vagy komplexebb idomokat, minden csak tervezés kérdése ezután? : a) DS nano-kocka, b) poliéder, c) csomó, stb..c.seeman, 48 MolBiotechnol, 2007,37,246-57
49 DS nanorendszerek II: T-elágazás ( kocka csúcsa ) 5 3 TAGCAGTTCCTATGAG 5 3 ACGACGTCAACTGCTA 5 TAGCAGTT AACTGCTA 3 5 TAGCAGTT AACTGCTA CTCATAGGGACGTCGT 5 3 CTCATAGGGACGTCGT CTCATAGGGACGTCGT 5 3 Összefoglalva: 3 CCTATGAG CTCATAGG 5 49



50 50
51 DS és RS hidrolízise: 1) Savas hidrolízis az eredmény: depurinálás (A v. G) memo: az RS és a DS kb. egyforma sebességgel hidrolízál savban kérdés: a 2-deoxi D-ribóz félacetálja savban felnyílik-e? 51
52 2) Lúgos hidrolízis (az RS lúgos hidrolízise): eredmény: lánchasadás memo: ezért az RS kevésbé, míg a DS jobban ellenáll a lúgos hidrolízisnek memo: azért tárol a DS és nem csak RS alapinformációt, mert az előbbi jobban ellenáll a lúgos hidrolízisnek, azaz stabilabb! 52
53 DS, RS építőelemek kémiai szintézise: 1) ukleozid szintézis I: a ilbert-johnson-nukleozid szintézis T.B. Johnson, G. E. ilbert, Science 69, 579 (1929) G. E. ilbert, T. B. Johnson, J. Am. Chem. soc. 52, 2001, 4489 (1930). 53
54 2) ukleozid szintézis II: kapcsolás ribozilaminnal A reakció javasolt mechanizmusa: 54
55 3) Szubsztituált nukleozid (purin) származékok továbbalakítása: Cl 3 2 S 2 / i 2 S R R R adenozin R: -D-ribofuranozil 55
56 4) ukleotid szintézis: nukleozidok foszforilezése (pl. dibenzil-foszfokloridáttal) rvosi alkalmazások: S 6-merkapto-purin Gyerekeknél akut leukémia kezelésére 80%-os gyógyulás 2 C P Cl 2 C dibenzil-foszfokloridát 2 allopurinol köszvény kezelésére aciklovir erpes virus kezelésére 2 C-atom hiányzik a ribózból 56
57 ligonukleotid szintézis:foszforamidit kapcsolási módszer McBridge és Caruthers Dmtr hasítás (dimetoxi-tritil) 2. Aktiválás és kapcsolás xidálás I2 (2 vagy TF) Lánczárás (az elreagálatlan 5 --t acetilezzük) 3.
58 DS kémiai (szilárdfázisú) szintézise: észter kötés(ek) kialakítása, S foszfor centrumon Védőcsoportok I: a bázisok védelme permanens csoportokkal Cél: a primer aminok védelme, avagy a nukleofil csoportok álcázása, hogy a majdani kapcsolásnál már csak a ribóz -5 legyen csupán az egyetlen u. A G C Rib Rib 4 -benzoilezés C Rib C C 6 -benzoilezés vagy 2 -izobutiril C C3 4 -acetilezés Rib T C 3 nem kell védeni Rib 58
59 Védőcsoportok II: a foszfodiészter védelme permanens csoporttal B C cianoetilészter formában CE P B Mindkét típusú permanens védőcsoport típus (-benzoil vagy -acetil és a CE) is eltávolítható: vizes 3 -val memo: a szintézis végén az összes állandó védőcsoport vizes ammóniával kvantitatíve eltávolítható 59
60 Védőcsoportok III: az 5 - csoport ideiglenes védelme Me Dmtr Me (enyhén savas rendszer) C P B C 2 Cl 2 + 3% CCl 3 -C Me C P B narancs szín Me 60
61 A szilárd hordozó: CPG (controlled pore glass) kontrolált pórusú üveg -kémiailag inert -nem duzzad -amino funkcionalizált CPG Me Si (C 2 ) n 2 Me A linker : Pl. borostyánkősav 2 C C C C (C 2 ) n Me Si 3 / 2 hasító hely Me CPG 2 C C 61
62 A DS szintézis lépésről lépésre : -szilárdfázisú szintézis 3 -től 5 irányba halad a Caruthers -módszer Dmtr B 1 memo: a bioszintézis menete az 5 -től 3 irányba halad 1. Dmtr hasítás (dimetoxi-tritil) C C (C 2 ) n Me Si Me CPG B 1 enyhe savas detritilezés (3% TCA) 2. Aktiválás és kapcsolás C C (C 2 ) n Me Si Me CPG 62
63 2. Aktiválás és kapcsolás B 1 C C (C 2 ) n Me Si CPG Me Dmtr foszforamidit B 2 P CE MeC + S (5' S reakciója) tetrazol (gyenge sav) 63
64 3. Lánczárás (az elreagálatlan 5 --t acetilezzük) 4. xidálás I 2 ( 2 vagy TF) Ciklus végén 4 -val: - minden permanens védőcsoport eltávolítása - gyantáról való lehasítás Legvégén: kromatográfiás tisztítás 64
65 Összefoglalás: oligonukleotidok laboratóriumi szintézisére Foszforamidit kapcsolási módszer DMTr B 3 R P B 2 CPG B 1 DMTr R P i Pr 2 B 3 tetrazol 1. lépés Kapcsolás R P R P B 2 CPG B 1 B 1,B 2,B 3 : védett bázisok R: C C 2 C 2 -cianoetil egy savra stabil, de bázisra érzékeny védõcsoport DMTr : tetrazol: CPG: "controlled pore glass" (porózus üveg) dimetoxitritil egy savra érzékeny védõcsoport 65
66 DMTr B 3 B 3 R P B 2 R P B 2 I 2 R P B 1 A R P B 1 2. lépés xidáció CPG 3. lépés Védõcsoport eltávolítása CPG memo: A PCR-től eltérően a kémiai szintézis nem igényel templátot 66



67 A szintetikus biológia hajnala J. Craig Ven Science 2 July 2010: Vol. 329 no pp Creation of a Bacterial Cell Controlled by a Chemically Synthesized Genome íme az első olyan élő szervezet amely atyja egy számítógép, s amely megalkotása az emberi elme és egynéhány robot műve volt 2003-ban még csak egy alig több mint 5000 bázispárt, 2010-ben már 1 millió bázispárt lehet szintetikusan összerakni! Aldous uxley Szép új világ 67






68 A DS enzimatikus szintézis: a replikáció cél: a genetikai kód megkettőződése: A kettős hélix az egyik pontján kitekeredik, majd a templátok mentén a komplementer szálak megszintetizálódnak. 68
 DS polimeráz II: 5 => 3 irányba szintetizál (alig ismert még) DS polimeráz III: 5 3 irányban szintetizálja az új láncot, ATP, GTP, CTP és TTP melyek magukon")
")
69 A DS enzimatikus szintézis: a replikáció DS polimeráz I: 5 => 3 irányba szintetizál (lassabb, de pontosabb, javítani is tud.) DS polimeráz II: 5 => 3 irányba szintetizál (alig ismert még) DS polimeráz III: 5 3 irányban szintetizálja az új láncot, ATP, GTP, CTP és TTP melyek magukon hordozzák az aktiváláshoz szükséges energiát. Leading strand = vezető szál, folyamatos szálszintézis Lagging strand = követő, sántikáló szál, darabonkénti szálszintézis, majd utólagos összekötés (DS ligáz) Primer: alkalmas RS Primase: RS primert hozza létre amely kell ahhoz hogy a DS polimerázok el tudjanak kezdeni dolgozni. DS ligáz: csak összekapcsol két láncot 69
 A feladat: [ ] n 1.")
70 PCR vagy Polymerase Chain Reaction (magyar polimeráz láncreakció) egyszerű és hatékony enzimatikus módszer a DS-molekulák számának - adott láncvégek (primerek ) közötti növelésére. (VIDE) A feladat: [ ] n 1. ciklus fütés T=95 C és alkalmas 2 primer hozzáadása T=55 C: a primerek helyrekerülnek (letapadnak, annelláció) 5 5 T=72 C: nukleotidok egyesévél bekapcsolódnak, majd a polimeráz enzim egy lánccá kapcsolja össze, a szintézis iránya 5 3 csak! 70
71 2. ciklus fütés T=95 C és alkalmas primerek hozzáadása T=55 C (helyre kerülnek az új primerek) T=72 C-on a nukleotidok bekapcsolódnak, s a polimeráz összeligálja az új szálakat memo: 4 kópia van de ebből még mind csak közti termék, tehát egyik sem jó még! memo: azért lesz rövidülés, mert a polimeráz szintézis iránya 5 3 csak! 71
72 3. ciklus végén 8 kópia van s ebből már 2 jó! 4. ciklus végén 16 kópia van, ebből 8 rossz, de 8 jó! 5. ciklus végén 32 kópia van, ebből 10 rossz, 22 jó! 6. ciklus végén 64 kópia van, ebből 12 rossz 52 jó! 10. ciklus végén 1024 kópia van, ebből 20 rossz, 1004 jó! összes kópiák száma: 2 n, ebből rossz 2n, jó (2 n 2n) Kary B. Mullis USA biokém ban kifejleszti a PCR-t, ban megkapja a obel-d.
73 30. ciklus után a helyes kópiák száma (a várt termék) >1 milliárd tehát: *30 [ ] n>10 9 Lényegében tiszta terméket kapunk, mert 2n << (2 n 2n) 73
74 ukleinsavak: 1) A kromoszóma és felépítése 2) DS- RS építőelemek: ukleozidok, nukleotidok, nukleinsavak em-rs és -DS alkotó purinszármazékok A DS térszerkezete, a bázispárok 3) DS nanorendszerek 4) DS (RS) reaktivitása: hidrolízise az építőelemek kémiai szintézise kémiai (szilárdfázisú) szintézis enzimatikus szintézis: a replikáció 5) Transzkripció 6) Transzláció 7) DS analitika: szekvenálás 8) Bioenergetika 74

 A")
 Folyamata: a bioszintézis menete az 5 -től 3")
75 5) Transzkripció A DS-ről mint információhordozóról a gének leolvasása, a hírvivő RS szintézise: DS (transzlokáció és mrs érés) mrs transzkripció átírás, másolás RS polimeráz (enzim) fehérje transzláció fordítás, dekódolás riboszóma (ribozim) A DS-ről RS-re átírás kezdete: RA-Polimerase Transzkripciós faktor (fehérje complex) Folyamata: a bioszintézis menete az 5 -től 3 irányba halad 75


76 RS polimeráz Transzkripció lépései: iniciáció: promoter régió elongáció terminálás 5 CATCCAATTGG 3 kódoló DS szál 5 CAUCCAAU 3 születő RS szál 3 GTAGGTTAACC 5 templát DS szál A transzkripció iránya 76


77 Ribonukleinsav (RS) és fehérjeszintézis gén: olyan DS szegmens amely tartalmazza mindazon információk összességét amely az adott polipeptid vagy fehérje szintéziséhez szükséges. (pl. egy egyszerű bacilus DS-e is legalább 3000 különböző enzimfehérjét kódol.) A transzkipció folyamatának szereplői: RS-polimeráz: egy enzim amely széttekeri a DS kettős spirált Promoter: a DS-nek az a része ahova az RS-polimeráz kötődik és ahol megkezdi a transzkripciót RS nukleotidok, mint építőelemek Terminátor: prokarióták azon DS része ami jelzi a transzkripció végét Transzkripciós egység: az a DS darab amely RS-é átíródik. Transzkripcós faktorok: az eukarióta sejtek azon fehérjéi amelyek az RS-polimeráz kötését, illetve a transzkripció megkezdését elősegítik. Transzkripciós iniciációs komplex: a transzkripciós faktorok és a promoter régióhoz kapcsolódó RS-polimeráz II együttese. TATA box : A DS promoter része 77

78 RS polimeráz láncszétválás A hírvivő-rs (messenger-rs, röviden mrs) szintézise: etc. P P P DS A C C G dezoxiribóz: P G P C P U P G ribóz: etc. P P P DS A C C G U G G C etc. P P P komplementer RS etc. P P P DS A C C G U G G C etc. P P P komplementer RS. Transzkripció a sejtmagban történik, a szintetizálódott RS-nek van fehérjét kódoló (exon: expressed sequence ) és nem-kódoló része (intron: intervening sequence ). Az intron a pre-mrs-be átíródik, majd az RS érés során kivágódik. A kész m-rs elvándorol a citoplazmába ahol a fehérjeszintézis templátjaként szerepel. 78
79 fehérje DS: - nem kódoló részek - kódoló részek (gének) >>>>>> mrs: - nem-kódoló részek (intron: intervening sequence ) - kódoló részek (exon: expressed sequence ) >>>>>>> fehérje 79
80 RS fajták és funkciójuk: kódoló RS: mrs: hírvivő (angol: messenger) RS: DS-ből másolt RS, mely a genetikai információt hordozza, és ezt a riboszómához szállítja, ahol a rajta lévő információ alapján a fehérjék szintetizálódnak. árom egymás melletti bázis alkot egy-egy kodon-t, amely egy-egy aminosavat kódol. pre-mrs: éretlen mrs. Az érés során a nem-kódoló részek (intronok) kivágódnak, csak a kódoló részek (exonok) maradnak az érett mrs-ben. nem-kódoló RS: trs: transzfer-rs: segít az mrs-en található kód (kodon) leolvasásában. Minden létező kodonhoz külön trs tartozik, mely a kodonnak megfelelő aminosavat hordozza, és a riboszómához szállítja, majd ott részt vesz a fehérjeszintézisben rrs: riboszomális RS: a riboszóma alkotóeleme, mely a fehérjeszintézis katalitikus centrumát alkotja. léteznek további nem-kódoló RS-ek is, mint például: sirs: (small interfering RA): gének kikapcsolásáért felelős rövid RS részek, amelyek a DS-hez kötődve meggátolják az adott rész átíródását. 80


81 Az RS térszerkezete: - általában egyszálú, de - Watson-Crick-féle párképzés az egy szálon belül, azaz önmagával is kialakulhat: egy hairpin RS téralkat trs térszerkezete E.coli rrs kisalegységének téralkata 81
82 Szállító RS (transzfer-rs, trs) feladata: hogy a megfelelő helyre eljuttassa az aminosavat, felépítése: A,U,C,G mellett módosított nukleotidok is: PSU, RT, stb. 3 A trs aminosav akceptor karja antikodon hurok 82
83 Szállító RS (transzfer-rs, trs) jellemzői: nukleotidból áll, - mindig a CCA- véghez kapcsolódik észterkötéssel az aminosav (pl. Tyr), - minden aminosavnak saját trs-e van, amelyen az annak az aminosavnak megfelelő antikodon található (egy aminosavhoz egy vagy akár több trs is tartozhat) - például: Met UAC.tRS: antikodon (CAU) AUG.mRS: kodon 83
84 A mrs tripletjei a genetikai kód: egy aminosavhoz egynél több kodon (egynél több antikodonnal ellátott t-rs) tarozik: As. m-rs As. m-rs As. m-rs Ala GCA is CAC Ser AGC GCC CAU AGU GCG Ile AUA UCA GCU AUC UCG Arg AGA AUU UCC AGG Leu CUA UCU CGA CUC Thr ACA CGC CUG ACC CGG CUU ACG CGU UUA ACU Asn AAC UUG Trp UGG AAU Lys AAA Tyr UAC Asp GAC AAG UAU GAU Met AUG Val GUA Cys UGC Phe UUU GUG UGU UUC GUC Gln CAA Pro CCA GUU CAG CCC Glu GAA CCG lánckezdő GAG CCU fmet AUG Gly GGA GGC lánczárás UAA GGG UAG GGU UGA kodon X Y Z antikodon 84

85 Transzláció, az információ lefordítása fehérje nyelvre, mely lépés során a citoszólban a ribószómán megtörténik a fehérjeszintézis. A szintézis - résztvevői: mrs (kodon) + trs (antikodon + aminosav) - helyszíne: a riboszóma - eredménye: a fehérje 85

86 Összefoglaló: az információáramlás hierarchiája: DS kodon mrs kodon trs antikodon aminosav 5 ACCTTTCTAAAG ACCTTTCTAAAG UGGAAAGAUUUC 5 UGGAAAGAUUUC 5 ACCUUUCUAAAG ACCUUUCUAAAG TrpLysAspPhe Kodon táblázat az mrs tripletjeinek kódját rendeli az aminosavakhoz 86
,")
, MW~ 4MDa, Prokarióta riboszóma térszerkezete")
 szintézis sebessége: 15-40 amidkötés/sec szintézis megbízhatósága: ~1 hiba/10 6 amidkötés")
87 a riboszóma egy RS-fehérje komplex: két alegység funkciója: kisalegység: köti az mrs-t nagy alegység: katalitikus aktivitás, fehérje szintézis mérete: - prokarióta esetén 70S (30S és 50S), MW~ 2.6MDa, - eukarióta esetén 80S (40S és 60S), MW~ 4MDa, Prokarióta riboszóma térszerkezete összetétele: RS és fehérje (L1,..L34) szintézis sebessége: amidkötés/sec szintézis megbízhatósága: ~1 hiba/10 6 amidkötés memo: nem az L1,..L34 fehérjék hanem az RS végzi a katalízist, ezért a riboszóma nem enzim hanem ribozim. 30S 50S S: (Svedberg egység) centrifugálás során a szedimentációs sebesség 87
88 Transzláció 1. lépés: (itt kezdődjön a transzláció) mindig az AUG mrs triplet. 2. ide mindig az -For-Met aminosav kerül 3. szintézis irány -term.-től a C-term. Felé ( C) 4. lánczáró kodonok az mrs-en: UAA, UAG és UGA. ( a mondat végi pontot jelölik ) 5. a szintézis legvégén a For-csoportot egy enzim lehidrolizálja. 88
89 Az amid szintézis mechanizmusát lásd később 89
90 90
91 A ribozim katalizálta peptidszintézis elemi kémiai lépései: az 50S alegység 2486 adeninje segíti a nukleofil addiciót : 1) Az adenin nemkötő elektronpárjának protonvonzása részlegesen deprotonálja az aminocsoportot, ezzel fokozva annak nukleofil jellegét. 2) A tertaéderes köztitermék elbomlását a proton visszaadása iniciálja, kialakítva így a peptidkötést. (Science 289; , 2000) 91



92 Transzláció lépéseinek áttekintése: - ciklusonként 3 db. GTP-t használ fel 92

93 Kérdés: mi a következő DS templáthoz szekvenciához tartozó fehérje aminosav-sorrendje? DS: 5 TGA TTC GTA TTG CTC GTG 3 DS: 5 TGA TTC GTA TAG CTC GTG 3 mrs:3 ACU AAG CAU AUC GAG CAC 5 Peptid: Ser -Glu -Tyr - Leu - Glu - is S - E - Y - L - E - kérdés: helyes-e ez az átírás? DS mrs 5 A 3 T 5 U T A A G C C 3 C 5 G 3 G memo1: Ügyeljünk tehát a bázissorrend irányultságára ( ) memo2: minden enzim az 5 oldal felől olvas! 93




94 Az antibiotikumok egy jelentős hányada a riboszómához kötve fejti ki szelektív hatását: Az Erythromycin A kölcsönhat az 50S (nagy alegység) 2058-as adeninjével, s igy beül és blokkolja azt a csatornát amelyen keresztűl bújik ki a fehérje. Meggátolja tehát a bakteriális fehérje szintézisét és a baktérium elpusztúl. A Eukarióták riboszómájának 2058 nukleotidja nem A hanem G, s ehhez nem köt az antibiotikum, igy lesz szelektív a hatás A Clindamycin az amidkötés kialakítását gátolja meg a ribószóma felületén erre a helyre kötődve ature 413, (2001) 94
 DS nanorendszerek 4) DS (RS)")
 Transzkripció 6) Transzláció 7) DS analitika: szekvenálás 8) Bioenergetika")
95 ukleinsavak: 1) A kromoszóma és felépítése 2) DS- RS építőelemek: ukleozidok, nukleotidok, nukleinsavak em-rs és -DS alkotó purinszármazékok A DS térszerkezete, a bázispárok 3) DS nanorendszerek 4) DS (RS) reaktivitása: hidrolízise az építőelemek kémiai szintézise kémiai (szilárdfázisú) szintézis enzimatikus szintézis: a replikáció 5) Transzkripció 6) Transzláció 7) DS analitika: szekvenálás 8) Bioenergetika 95
96 7) DS analitika: a szekvenálás cél egy DS fragmens bázissorrendjének meghatározása: megoldás: A lánczáró festékkel jelölt didezoxynukleotid (ddtp) vagy Sanger-féle módszer alapgondolata: a 3'-hidroxi-csoport hiánya megakadályozza a PCR-ezés során a lánc továbbépülését: lánctörést okoz. Frederick Sanger Chain Termination Method (Sanger, F., et al: PAS, 1977, 74, 5463.) A szekvenálandó (templát) DS 3 C-G-A-A-T-A-T-G-C-G-A-G-T-C-T-G-G-C-A-A-C-T 5 amelyre mint templátra PCR rel komplementer szálakat fogunk előállítani, de szintézis során nem csak a szokásos ATP, GTP, CTP és TTP nukleotidokat használunk, hanem ddatp, ddgtp, ddctp és ddttp-t is. 96
97 templát DS és alatta a különböző PCR termékek amelyek rendre a ddtp-k beépülése miatt terminálódtak: 3 C-G-A-A-T-A-T-G-C-G-A-G-T-C-T-G-G-C-A-A-C-T 5 G-C-T-T-A-T-A-C-G-C-T-C-A-G-A G-C-T-T-A-T-A-C-G-C-T-C G-C-T-T-A-T-A-C-G-C-T-C-A-G-A-C-C-G-T G-C-T G-C-T-T-A G-C-T-T-A-T-A-C G-C-T-T-A-T-A-C-G-C-T-C-A-G-A-C-C-G G-C G-C-T-T G-C-T-T-A-T-A-C-G-C G-C-T-T-A-T-A-C-G-C-T-C-A-G-A G-C-T-T-A-T-A-C-G-C-T-C-A-G-A-C G-C-T-T-A-T-A-C-G-C stb. minden lehetséges fragmenst előállítunk agaróz gélen (oszlopon) megfuttatjuk, amely méret szerint szeparál, s az oszlop alján az éppen kifolyó fragmenst színe alapján azonosítjuk G-C-T-T-A-T-A-C-G-C-T-C-A-G-A-C-C-G-T-T-G-A bizonytalan szegmens 97

98 egy valódi szekvenálás végeredménye: 98
99 8) Bioenergetika Elektromágneses energia forrása hőerőgép és belsőégésű motor lehet, amelyekben a hőenergia égésből, azaz kémiai átalakulásból származik. Ezek rendre az atomok és molekulák elektronszerkezetéhez köthető átalakulások, azaz elektromágneses folyamatok. Az élőlények számára is kémiai folyamatok, azaz az elektromágneses kölcsönhatás biztosítja az energiatárolást és energiafelhasználást, tehát a biológiai energiák is elektromágneses eredetűek. memo: Az energiaformákat vissza lehet vezetni a fizika négy alapvető kölcsönhatásának valamelyikére. A négy alapvető kölcsönhatás 1) a gravitációs, 2) az elektromágneses, 3) a gyenge (β-bomlás (elektron, ill. pozitron emisszió), illetve az elektronbefogás) 4) az erős kölcsönhatás. 99
 ΔG = 45.6 kj/mol ( 10.")
100 8) Bioenergetika ATP: adenozin -5 - trifoszfát foszforsavanhidrid foszfátészter DG~ 30.5 kj DG~ mol kj mol -1 ATP + 2 ADP + P i ATP + 2 AMP + PP i ΔG = 30.5 kj/mol ( 7.3 kcal/mol) ΔG = 45.6 kj/mol ( 10.9 kcal/mol) 100
101 8) Bioenergetika ATP és GTP mint energiahordozók: elem / akkumulátor (újrafeltölthető!) (ATP) felhasználás: direkt módon: kapcsolt reakció A sejtben igényelt anyagok szintézisében az endoterm ill. endergonikus reakciókhoz a kapcsolódó ATP/GTP hidrolízis adja a megfelelő hajtóerőt a reakció lefutásához. pl.a felhasználásra: - transzláció (fehérjeszintézis) - kreatin (energiaraktár az izomban) - (ATP) feltöltés: direkt módon: szubsztrát szintű foszforiláció: a metabolizmus (ételek, raktározott zsírsavak, egyéb anyagok lebontása) során az exoterm ill. exergonikus reakciókban történő ADP/GDP foszforilációja. indirekt módon: oxidatív foszforiláció a metabolizmus során redox-reakciókban redukált koenzimek keletkeznek: AD + és FAD 2 Ezek a redukált koenzimek elektrontranszporttal egybekötött, szabályozott regenerálása (oxidációja) során keletkezik ATP. 101
 szabadentalpia (DG) felszabadulást eredményez.")
102 8) Bioenergetika Atkins & de Paula 99 kérdés: mennyi glükóz elégetése teszi lehetővé hogy egy 30 g össztömegű madár számára, hogy 10 m magasra repüljön? tapasztalat: 1 mol szilárd glükóz x C 2 gázzá és y folyékony 2 való oxidációja 25 C-on ~ 2828 kj (2,8 MJ) szabadentalpia (DG) felszabadulást eredményez. válasz: az elvégezendő munka nagysága mgh= (30*10-3 kg)*(9,81*ms -2 )*(10 m) = 2,943 kg m 2 s -2 = 2,943 J mivel 1 mol glükóz DG ~ 2, J munka végzéséhez elegendő, 2,943 J munka elvégzéséhez 2,943 / 2, mol glükózra van szükség, ami 1,04 μmol cukrot jelent. Mivel a glükóz M W -je ~180 g/mol ezért ez hozzávetőlegesen (180 g/mol * 1,04*10-6 mol) = 0,19 mg kérdés: a koncentráló emberi agy ~25 J energiát igényel másodpercenként. Mennyi cukor (glükóz) elégetése szükséges ehhez óránként? tapasztalat: 1 mol glükóz oxidációja 25 C-on ~ 2828 kj szabadentalpia (DG) felszabadulását eredményezi. válasz: az elvégezendő munka nagysága óránként = (25 Js -1 )* (3,6*10 3 s) = J = 90 kj mivel 1 mol glükóz DG ~ 2, kj munka végzéséhez elegendő, 90 kj munka elvégzéséhez 90 / 2, mol glükózra van szükség, ami 32 mmol glükózt jelent. Mivel a glükóz M W -je ~180g mol -1 ezért ez hozzávetőlegesen (180 * 32*10-3 g ) = 5,7 g/óra Tehát az emberi agy naponta ~ 24 * 5,7g ~140g glükózt igényel. 102
/ M w (glükóz) 507/180 = 2,82 ezért 2,82 * 38 * 160g 17,1 kg ATP Ténylegesen egy 70 kg-os ember napi ATP fogyasztása ~ 145 kg (nem csak szénhidrátot, de zsírt és fehérjét is")
103 A napi g glükóz többsége ATP-vé alakul és így hasznosul a sejtekben. Aerob körülmények között 1 mol Glc (glükóz) mintegy 38 mol ATP eredményez. mivel M w (ATP)/ M w (glükóz) 507/180 = 2,82 ezért 2,82 * 38 * 160g 17,1 kg ATP Ténylegesen egy 70 kg-os ember napi ATP fogyasztása ~ 145 kg (nem csak szénhidrátot, de zsírt és fehérjét is fogyasztunk!) Ám a szervezetben egyszerre kb. 51 g össz ATP áll rendelkezésre, ami a teljes szükséglet (51 g /1, g) kb. 0,035% -a. Ez az tartalékolt ATP mennyiség tehát kb. (24* 3600s) * (3, ) ~ 30 s-ra elegendő mindössze! Tehát az ember ATP szükséglete ~ 100g / perc (aktív mozgás esetén 500g /perc) ADP/ATP ciklus (a foszforiltranszfer) A kémiai reakció: ATP 4 (aq) + 2 ADP 2 (aq) +P 4 2 (aq) DG~ 30 kj mol -1 A biokémiai rész ciklus: ½ 2 2 ADP 2 + P 4 2 ATP 4 DG 103
 DG elektron-átvitel ½ 2 + DG 2 ADP 2 + P 4 2 ATP 4 foszfor-átvitel DG memo: a félkövér résztvevők ( C, AD+ +, FAD 2, ATP) azok a")
104 A teljes biokémiai ciklus: C 2 C +2 2 DG égetés pl. D-Glc + DG AD + + AD + FAD 2 FAD AD + (ikotinamid-adenin-dinukleotid) DG elektron-átvitel ½ 2 + DG 2 ADP 2 + P 4 2 ATP 4 foszfor-átvitel DG memo: a félkövér résztvevők ( C, AD+ +, FAD 2, ATP) azok a nagyobb energiájú molekulák a megfelelő párokban. memo: A negatív DG-s reakciók hajtják a velük összekapcsolt ( csatolt ) pozitív DG-s (piros nyíl) reakciókat. Tűz, a levessel a levegőn keresztül csatolódik FAD (Flavin-adenin-dinukleotid)
105 éhány fontosabb foszfátvegyület foszfátcsoport(jai) hidrolízisének DG- értékei 37 o C-on: memo: foszfoenol-piruvát DG~ 62 kj mol -1 ATP ADP DG~ 31 kj mol -1 ADP AMP DG~ 28 kj mol -1 AMP DG~ 14 kj mol -1 glükóz-1-foszfát DG~ 21 kj mol -1 glükóz-6-foszfát DG~ 14 kj mol -1 fruktóz-6-foszfát DG~ 16 kj mol -1 kérdés: mit takar az ATP DG~ 31 kj mol -1 információ? A hidrolízis exergonikus DG< 0 és éppen 31 kj mol -1 energiát ad át más vele csatolt - esetleg endergonikus - reakciók lefolytatásához. Ezért becézzük a megfelelő savanhidrid kötést gyakran nagyenergiájú kötésnek. memo: vegyük észre hogy DG sorozatban az ATP és ADP középen helyezkednek el, ezért lehet foszfát donor és akceptor is. 105
106 ATP 4 (aq) + 2 ADP 2 (aq) +P 4 2 (aq) tény: az ATP DG~ 31 kj mol -1 valamint a D~ 20 kj mol -1 és DS~ +34 JK -1 mol -1 memo: mivel DG = D TDS TDS~ 310 K *34 JK -1 mol -1 ~ +11 kj mol -1 ) memo: az 1 mol víz a hidrát burok része, mivel a reakció nem vákuumban megy (ezért nem számít ). Tehát formálisan mólszám növekedés van (1-ből 2 mol lesz), s emiatt jár ez a reakció entrópia növekedéssel, avagy a komplexitás csökkenésével, amely egy további kedvező tény. A DG értéke ( 31 kj mol -1 ) ezért is ilyen kedvező. Az ATP hidrolízise felhasználható olyan csatolt endergonikus reakciókhoz, amelyek DG-je nem nagyobb mint +31 kj mol -1. pl. egy peptidkötés szintézise amely erősen endergonikus: DG~ +17 kj mol -1 memo: az 1 mol víz a hidrát burok része lesz, mivel a reakció nem vákuumban megy. Innen a nagy entrópia csökkenés (2-ből 1-re), a komplexitás növekedés. DG ezért is ilyen kedvezőtlen. memo: nem csak az entalpiaváltozás, de a szintézis entrópia csökkenése (komplexitás növekedése) is jelentős! kérdés: hogy mehet végbe a reakció 37 o C-on? válasz: úgy hogy ATP csatolt ez a folyamat. memo: a csatolt rendszer értelmében a két reakció ATP hidrolíziseé és amidkötés kialakítása össze van kapcsolva! (a e két reakció elkülönítve (2 edényben) menne végbe, vagy ha csak úgy összeöntenénk a reagenseket, akkor a folyamat nem menne végbe.) Csatolást végzi a ribozim (riboszóma)
 szükséges.")

~ 2,2 kg cukor kell 1 mol (16,7 kda) fehérje")
107 megfigyelés: Itt nem részletezett okok miatt 1 peptidkötés kialakításához 3 ATP (v. GTP) szükséges. kérdés: hány gramm glükóz kell 1 mol hemoglobin bioszintéziséhez, ha az 153 aminosavból épül fel? válasz: 153*3 = 459 ATP szükséges. 1 mol glükóz 38 ATP eredményez, tehát 459/38~ 12 mol glükóz szükséges. tehát: (12*180 g)~ 2,2 kg cukor kell 1 mol (16,7 kda) fehérje szintéziséhez (~16,7 kg) A vörösvértest szárazanyag-tartalmuk 90%-a hemoglobin 107
 is organized in different chromosomes and protectively packaged.")
108 Érdekes nano-rendszerek ow to safely store genetic material: cable reels in the cell nucleus... Legend: The nucleosome ucleosomes are cable reels for genetic material. In the cell s nucleus, the genetic material (DA) is organized in different chromosomes and protectively packaged. In this storage packing, the DA is wound around nucleosomes, just as a cable on a reel. Each nucleosome consists of four pairs of proteins called histones. Apart from their role as cable reels, nucleosomes are also involved in gene regulation as they prevent DA from being read at wrong positions. 108

109 Érdekes nano-rendszerek I am not giving away the original data! A copy is good enough... Legend: RA polymerase RA polymerase is essential for each cell. This protein reads the genetic code from the genetic material in the cell nucleus and copies it onto so-called mra molecules. These copies are then shipped to the ribosomes where they serve as blueprints for proteins. Thus, the precious original always stays in the vault of the nucleus, and only copies are shipped. Because RA polymerase is so important, its function is constantly monitored and controlled by a multitude of other molecules. 109
 in yellow The repetition of the amino acids threonine (T) and alanine (A) T-A-T-A marks the beginning of a gene.")
110 Érdekes nano-rendszerek DA is long where does a gene begin and how to find it? Legend: The TATA-box binding protein (TBP) in yellow The repetition of the amino acids threonine (T) and alanine (A) T-A-T-A marks the beginning of a gene. The TATA-box binding protein (TBP) recognizes this specific genetic start sequence and allows correct positioning of RA polymerase. Without TBP, genes could not be copied to mra and would, hence, not be read out. If the tail of TBP is too long, it causes 110 some of the most severe neurodegenerative diseases

111 Replikáció Ismétlés Szemidiszkontinuus Azaz a DS megkettőződése mindkét fonal mellett, ugyanabba az irányba történik Ez azt feltételezi, hogy a replikációs mechanizmus mind 5 ->3, mind 3 ->5 irányban képes DS-t szintetizálni De a DS-polimeráz csak 5 ->3 irányban képes haladni, vagyis elsőként az 5 helyen levő nukleotidot építi be, majd 3 irányba folytatja a láncot Vezető szál: folyamatosan szintetizálódik Követő szál: szakaszosan szintetizálódik (kazaki-fragmentumok) Itt a szintézis iránya ellentétes a replikációs villa mozgásának irányával A követő szál a villától távolodva szintetizálódhat 111

112 Transzkripció Ismétlés Iniciáció Az RS polimeráz felismeri a DS promoter szakaszát, hozzákötődve szétválaszt 12bp-nyi szakaszt, és megkezdi az RS szál felépítését: ATP, UTP, GTP, CTP Elongáció Az RS-polimeráz 5 ->3 irányban építi be a ribonuleotidokat, működése során a DS-lánc mentén mozog Termináció A terminátor szakaszok lényegében lazítják a DS és RS közötti kapcsolatot memo: a folyamat aszimmetrikus: csak az egyik szál másolódik, a DS két szála csak rövid ideig távolodik el egymástól 112

113 Transzláció Ismétlés Iniciáció Egy riboszóma, amelyik már befejezte egy másik mrs transzlációját, alegységeire disszociál (IF-3) IF-3 kötődik 30S-hez és egy új RS-hez IF-2 hozzákapcsolja 30S-hez az első aminoacil-trs-t Ehhez az iniciációs komplexhez kötődik GTP, majd 30S, ezután IF-3 leválik GTP -> GDP: IF-2 is leválik 113

 A P-helyen levő dezacilált trs leválik a riboszómáról Az A-helyen levő dipeptidil-trs az A-helyre lép A következő kodon az")
114 Transzláció Ismétlés Elongáció A transzláció iránya = transzkripció iránya = 5 -> 3 Az aa-trs minőségét az mrs második kodonja határozza meg, amely az üres A-helyen van (Katalizátor: EF-Tu, Energia: GTP A peptidil transzferáz az fmet-t a P-helyen levő trs-ről az A-helyen levő aa-trs-re kapcsolja, így egy dipeptid alakul ki az A hely trs-én A transzlokáció során az mrs az A-helyre kapcsolódó peptidil-trs-sel együtt egy kodonnal továbblép balra (EF-G) A P-helyen levő dezacilált trs leválik a riboszómáról Az A-helyen levő dipeptidil-trs az A-helyre lép A következő kodon az A-helyre kerül 114

115 Transzláció Ismétlés Termináció RF-1 és RF-2 ismeri fel a STP kodonokat RF-3-mal és GTP-vel leállítják a transzlációt 115
CHO H H H OH H OH OH H CH2OH HC OH HC OH HC OH CH 2
 4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense F. Miescher (Svájc) 1882 Flemming: Chromatin elnevezés Waldeyer:
4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense F. Miescher (Svájc) 1882 Flemming: Chromatin elnevezés Waldeyer:
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
CHO H H H OH H OH OH H CH2OH CHO OH H HC OH HC OH HC OH CH 2 OH
 4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense nuclein Friedrich Miescher (Svájc, 1844-1895) 1970: FM Insitute
4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense nuclein Friedrich Miescher (Svájc, 1844-1895) 1970: FM Insitute
A nukleinsavkémiai kisszótár:
 A nukleinsavkémiai kisszótár: DS: dezoxiribonukleinsav, DA : olyan nukleotidegységekből felépülő nukleinsavak gyűjtőneve, amelyek dezoxiribóz cukorrészt tartalmaznak. A gének kódolására, és továbbadására
A nukleinsavkémiai kisszótár: DS: dezoxiribonukleinsav, DA : olyan nukleotidegységekből felépülő nukleinsavak gyűjtőneve, amelyek dezoxiribóz cukorrészt tartalmaznak. A gének kódolására, és továbbadására
4. Előadás. Nukleozidok, nukleotidok, nukleinsavak
 4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense nuclein Friedrich Miescher (Svájc, 1844-1895), izolálás 1970: FM
4. Előadás ukleozidok, nukleotidok, nukleinsavak Történeti háttér Savas karakterű anyagok a sejtmagból 1869-71 DS a sejtmag fő komponense nuclein Friedrich Miescher (Svájc, 1844-1895), izolálás 1970: FM
A nukleinsavkémia koronázatlan királyai, kémiai és orvosi Nobel-díjak:
 A nukleinsavkémia koronázatlan királyai, kémiai és orvosi obel-díjak: Francis arry James Dewey ompton rick Watson Maurice ugh Frederick Wilkins 1962 DS molekuláris szerkezetének felismeréséért aul Berg
A nukleinsavkémia koronázatlan királyai, kémiai és orvosi obel-díjak: Francis arry James Dewey ompton rick Watson Maurice ugh Frederick Wilkins 1962 DS molekuláris szerkezetének felismeréséért aul Berg
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Gergely Pál 2009
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Bay Péter
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Nukleinsavak építőkövei
 ukleinsavak Szerkezeti hierarchia ukleinsavak építőkövei Pirimidin Purin Pirimidin Purin Timin (T) Adenin (A) Adenin (A) Citozin (C) Guanin (G) DS bázisai bázis Citozin (C) Guanin (G) RS bázisai bázis
ukleinsavak Szerkezeti hierarchia ukleinsavak építőkövei Pirimidin Purin Pirimidin Purin Timin (T) Adenin (A) Adenin (A) Citozin (C) Guanin (G) DS bázisai bázis Citozin (C) Guanin (G) RS bázisai bázis
NUKLEINSAVAK. Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag
 NUKLEINSAVAK Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag RNS = Ribonukleinsav DNS = Dezoxi-ribonukleinsav A nukleinsavak
NUKLEINSAVAK Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag RNS = Ribonukleinsav DNS = Dezoxi-ribonukleinsav A nukleinsavak
Biológus MSc. Molekuláris biológiai alapismeretek
 Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Nukleinsavak. Szerkezet, szintézis, funkció
 Nukleinsavak Szerkezet, szintézis, funkció Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
Nukleinsavak Szerkezet, szintézis, funkció Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
CIÓ A GENETIKAI INFORMÁCI A DNS REPLIKÁCI
 A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak?
 A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód)
, és az öröklődés molekuláris alapjai (gén, genetikai kód)") 2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód) 2.1 Nukleotidok, nukleinsavak Információátadás (örökítőanyag) Információs egység
2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód) 2.1 Nukleotidok, nukleinsavak Információátadás (örökítőanyag) Információs egység
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
Nanotechnológia. Nukleinsavak. Készítette - Fehérvári Gábor
 Nanotechnológia Nukleinsavak Készítette - Fehérvári Gábor Bevezető A nukleinsavak az élő anyag alapvetően fontos komponensei. Meghatározó szerepet töltenek be az átöröklésben, a fehérjék szintézisében
Nanotechnológia Nukleinsavak Készítette - Fehérvári Gábor Bevezető A nukleinsavak az élő anyag alapvetően fontos komponensei. Meghatározó szerepet töltenek be az átöröklésben, a fehérjék szintézisében
TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak?
 TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis,
TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis,
Mária. A pirimidin-nukleotidok. nukleotidok anyagcseréje
 Prof.. Sasvári Mária A pirimidin-nukleotidok nukleotidok anyagcseréje 1 A nukleobázisok szerkezete Nitrogéntartalmú, heterociklusos vegyületek; szubsztituált purin- és pirimidin-származékok purin Adenin
Prof.. Sasvári Mária A pirimidin-nukleotidok nukleotidok anyagcseréje 1 A nukleobázisok szerkezete Nitrogéntartalmú, heterociklusos vegyületek; szubsztituált purin- és pirimidin-származékok purin Adenin
DNS replikáció. DNS RNS Polipeptid Amino terminus. Karboxi terminus. Templát szál
 DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
9. Szilárdfázisú szintézisek. oligopeptidek, oligonukleotidok
 9. Szilárdfázisú szintézisek oligopeptidek, oligonukleotidok Peptidszintézis Amidkötés kialakítása R H + H 2 Q R Q + H 2 H R H + H 2 Q R + H 3 Q sav-bázis reakció már nem nukleofil Amidkötés kialakítása
9. Szilárdfázisú szintézisek oligopeptidek, oligonukleotidok Peptidszintézis Amidkötés kialakítása R H + H 2 Q R Q + H 2 H R H + H 2 Q R + H 3 Q sav-bázis reakció már nem nukleofil Amidkötés kialakítása
BIOLÓGIA ALAPJAI. Anyagcsere folyamatok 2. (Felépítő folyamatok)
") BIOLÓGIA ALAPJAI Anyagcsere folyamatok 2. (Felépítő folyamatok) A molekuláris biológiai alapjai DNS replikáció RNS transzkripció Fehérje szintézis (transzláció) (Az ábrák többsége Dr. Lénárd Gábor Biológia
BIOLÓGIA ALAPJAI Anyagcsere folyamatok 2. (Felépítő folyamatok) A molekuláris biológiai alapjai DNS replikáció RNS transzkripció Fehérje szintézis (transzláció) (Az ábrák többsége Dr. Lénárd Gábor Biológia
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét. Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet
 Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ
 Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
A METABOLIZMUS ENERGETIKÁJA
 A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2014.10.01. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2014.10.01. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA
 A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2013.10.02. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2013.10.02. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód)
, és az öröklődés molekuláris alapjai (gén, genetikai kód)") 2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód) 2.1 Nukleotidok, nukleinsavak Információátadás (örökítőanyag) Információs egység
2. Sejtalkotó molekulák II. Az örökítőanyag (DNS, RNS replikáció), és az öröklődés molekuláris alapjai (gén, genetikai kód) 2.1 Nukleotidok, nukleinsavak Információátadás (örökítőanyag) Információs egység
Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ
 Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
Nukleinsavak SZERKEZET, SZINTÉZIS, FUNKCIÓ Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A bioenergetika a biokémiai folyamatok során lezajló energiaváltozásokkal foglalkozik.
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek
 Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
3. Sejtalkotó molekulák III.
 3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
CHO CH 2 H 2 H HO H H O H OH OH OH H
 2. Előadás A szénhidrátok kémiai reakciói, szénhidrátszármazékok Áttekintés 1. Redukció 2. xidáció 3. Észter képzés 4. Reakciók a karbonil atomon 4.1. iklusos félacetál képzés 4.2. Reakció N-nukleofillel
2. Előadás A szénhidrátok kémiai reakciói, szénhidrátszármazékok Áttekintés 1. Redukció 2. xidáció 3. Észter képzés 4. Reakciók a karbonil atomon 4.1. iklusos félacetál képzés 4.2. Reakció N-nukleofillel
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Kémiai Intézet Kémiai Laboratórium. F o t o n o k k e r e s z tt ü z é b e n a D N S
 Szalay SzalayPéter Péter egyetemi egyetemi tanár tanár ELTE, ELTE,Kémiai Kémiai Intézet Intézet Elméleti ElméletiKémiai Kémiai Laboratórium Laboratórium F o t o n o k k e r e s z tt ü z é b e n a D N S
Szalay SzalayPéter Péter egyetemi egyetemi tanár tanár ELTE, ELTE,Kémiai Kémiai Intézet Intézet Elméleti ElméletiKémiai Kémiai Laboratórium Laboratórium F o t o n o k k e r e s z tt ü z é b e n a D N S
12/4/2014. Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció. 1952 Hershey & Chase 1953!!!
 Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció 1859 1865 1869 1952 Hershey & Chase 1953!!! 1879 1903 1951 1950 1944 1928 1911 1 1. DNS szerkezete Mi az örökítő anyag? Friedrich Miescher
Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció 1859 1865 1869 1952 Hershey & Chase 1953!!! 1879 1903 1951 1950 1944 1928 1911 1 1. DNS szerkezete Mi az örökítő anyag? Friedrich Miescher
11. előadás: A génektől a fehérjékig A genetikai információ áramlása
 11. előadás: A génektől a fehérjékig A genetikai információ áramlása A DNS információtartalma specifikus nukleotidsorrend formájában van jelen Az átörökített DNS fehérjék szintézisét szabályozva tulajdonságok
11. előadás: A génektől a fehérjékig A genetikai információ áramlása A DNS információtartalma specifikus nukleotidsorrend formájában van jelen Az átörökített DNS fehérjék szintézisét szabályozva tulajdonságok
A genetikai lelet értelmezése monogénes betegségekben
 A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
A tananyag felépítése: A BIOLÓGIA ALAPJAI. I. Prokarióták és eukarióták. Az eukarióta sejt. Pécs Miklós: A biológia alapjai
 A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
TÉMAKÖRÖK. Ősi RNS világ BEVEZETÉS. RNS-ek tradicionális szerepben
 esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
RNS SZINTÉZIS ÉS ÉRÉS
 RNS SZINTÉZIS ÉS ÉRÉS A genom alapvetõ funkciója, hogy a sejt mûködéséhez esszenciális gépek (fehérjék) elõállí tására vonatkozó információt tartalmazza. A DNS-ben rejlõ információ egy kétlépéses folyamatban
RNS SZINTÉZIS ÉS ÉRÉS A genom alapvetõ funkciója, hogy a sejt mûködéséhez esszenciális gépek (fehérjék) elõállí tására vonatkozó információt tartalmazza. A DNS-ben rejlõ információ egy kétlépéses folyamatban
15. Fehérjeszintézis: transzláció. Fehérje lebontás (proteolízis)
") 15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
A gyakorlat elméleti háttere A DNS molekula a sejt információhordozója. A DNS nemzedékről nemzedékre megőrzi az élőlények genetikai örökségét.
 A kísérlet megnevezése, célkitűzései: DNS molekula szerkezetének megismertetése Eszközszükséglet: Szükséges anyagok: színes gyurma, papírsablon Szükséges eszközök: olló, hurkapálcika, fogpiszkáló, cérna,
A kísérlet megnevezése, célkitűzései: DNS molekula szerkezetének megismertetése Eszközszükséglet: Szükséges anyagok: színes gyurma, papírsablon Szükséges eszközök: olló, hurkapálcika, fogpiszkáló, cérna,
DER (Felületén riboszómák találhatók) Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.
 Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.") Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
MOLEKULÁRIS GENETIKA A DNS SZEREPÉNEK TISZTÁZÁSA
 MOLEKULÁRIS GENETIKA A DNS SZEREPÉNEK TISZTÁZÁSA A DNS-ről 1869-ben Friedrich Mischer német orvos írt először. A gennyben talált sejtekben egy foszforban gazdag, de ként nem tartalmazó anyagot talált.
MOLEKULÁRIS GENETIKA A DNS SZEREPÉNEK TISZTÁZÁSA A DNS-ről 1869-ben Friedrich Mischer német orvos írt először. A gennyben talált sejtekben egy foszforban gazdag, de ként nem tartalmazó anyagot talált.
Poligénes v. kantitatív öröklődés
 1. Öröklődés komplexebb sajátosságai 2. Öröklődés molekuláris alapja Poligénes v. kantitatív öröklődés Azok a tulajdonságokat amelyek mértékegységgel nem, vagy csak nehezen mérhetők, kialakulásuk kevéssé
1. Öröklődés komplexebb sajátosságai 2. Öröklődés molekuláris alapja Poligénes v. kantitatív öröklődés Azok a tulajdonságokat amelyek mértékegységgel nem, vagy csak nehezen mérhetők, kialakulásuk kevéssé
9. Szilárdfázisú szintézisek. oligopeptidek, oligonukleotidok
 9. Szilárdfázisú szintézisek oligopeptidek, oligonukleotidok Peptidszintézis Amidkötés kialakítása R O OH + H 2 N Q R O Q N + H 2 O H R O OH + H 2 N Q R O O + H 3 N Q sav-bázis reakció már nem nukleofil
9. Szilárdfázisú szintézisek oligopeptidek, oligonukleotidok Peptidszintézis Amidkötés kialakítása R O OH + H 2 N Q R O Q N + H 2 O H R O OH + H 2 N Q R O O + H 3 N Q sav-bázis reakció már nem nukleofil
Glikolízis. Csala Miklós
 Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Transzláció. Leolvasás - fehérjeszintézis
 Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
Semmelweis Egyetem / Élettani Intézet / Budapest. Bioinformatika és genomanalízis az orvostudományban. Szekvenciaelemzés. Cserző Miklós 2017
 Bioinformatika és genomanalízis az orvostudományban Szekvenciaelemzés Cserző Miklós 2017 A mai előadás Szekvencia analízis statisztikus szempontból Annotálás homológia alapján Az annotálás szempontjai
Bioinformatika és genomanalízis az orvostudományban Szekvenciaelemzés Cserző Miklós 2017 A mai előadás Szekvencia analízis statisztikus szempontból Annotálás homológia alapján Az annotálás szempontjai
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak
 BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
1. jelentésük. Nevüket az alkotó szén, hidrogén, oxigén 1 : 2 : 1 arányából hajdan elképzelt képletről [C n (H 2 O) m ] kapták.
![1. jelentésük. Nevüket az alkotó szén, hidrogén, oxigén 1 : 2 : 1 arányából hajdan elképzelt képletről [C n (H 2 O) m ] kapták.](/thumbs/55/37234755.jpg "1. jelentésük. Nevüket az alkotó szén, hidrogén, oxigén 1 : 2 : 1 arányából hajdan elképzelt képletről [C n (H 2 O) m ] kapták.") Összefoglalás II. Szénhidrátok 1. jelentésük Nevüket az alkotó szén, hidrogén, oxigén 1 : 2 : 1 arányából hajdan elképzelt képletről [C n (H 2 O) m ] kapták. Ha ezeket az anyagokat hevítjük vizet vesztenek
Összefoglalás II. Szénhidrátok 1. jelentésük Nevüket az alkotó szén, hidrogén, oxigén 1 : 2 : 1 arányából hajdan elképzelt képletről [C n (H 2 O) m ] kapták. Ha ezeket az anyagokat hevítjük vizet vesztenek
A replikáció mechanizmusa
 Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
I. A sejttől a génekig
 Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Kromoszómák, Gének centromer
 Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei)
") Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
 Az élő szervezetek felépítése I. Biogén elemek biomolekulák alkotóelemei a természetben előforduló elemek közül 22 fordul elő az élővilágban O; N; C; H; P; és S; - élő anyag 99%-a Biogén elemek sajátosságai:
Az élő szervezetek felépítése I. Biogén elemek biomolekulák alkotóelemei a természetben előforduló elemek közül 22 fordul elő az élővilágban O; N; C; H; P; és S; - élő anyag 99%-a Biogén elemek sajátosságai:
AZ ÉLET KÉMIÁJA... ÉLŐ ANYAG SZERVEZETI ALAPEGYSÉGE
 AZ ÉLET KÉMIÁJA... ÉLŐ ANYAG SZERVEZETI ALAPEGYSÉGE A biológia az élet tanulmányozásával foglalkozik, az élő szervezetekre viszont vonatkoznak a fizika és kémia törvényei MI ÉPÍTI FEL AZ ÉLŐ ANYAGOT? HOGYAN
AZ ÉLET KÉMIÁJA... ÉLŐ ANYAG SZERVEZETI ALAPEGYSÉGE A biológia az élet tanulmányozásával foglalkozik, az élő szervezetekre viszont vonatkoznak a fizika és kémia törvényei MI ÉPÍTI FEL AZ ÉLŐ ANYAGOT? HOGYAN
Hamar Péter. RNS világ. Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. www.meetthescientist.hu 1 26
 Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
Purin nukleotidok bontása
 Dr. Sasvári MáriaM Purin nukleotidok bontása 24 1 Purin nukleotidok bontása AMP B r -p 5 nukleotidáz GMP P i adenozin (6-amino) ADA 2 adenozin deamináz 3 B r guanozin P i inozin (6-oxo) P i PP P i purin
Dr. Sasvári MáriaM Purin nukleotidok bontása 24 1 Purin nukleotidok bontása AMP B r -p 5 nukleotidáz GMP P i adenozin (6-amino) ADA 2 adenozin deamináz 3 B r guanozin P i inozin (6-oxo) P i PP P i purin
folsav, (a pteroil-glutaminsav vagy B 10 vitamin) dihidrofolsav tetrahidrofolsav N CH 2 N H H 2 N COOH
 dihidrofolsav tetrahidrofolsav N CH 2 N H H 2 N COOH") folsav, (a pteroil-glutaminsav vagy B 10 vitamin) 2 2 2 2 pirimidin rész pirazin rész aminobenzoesav rész glutaminsav rész pteridin rész dihidrofolsav 2 2 2 2 tetrahidrofolsav 2 2 2 2 A dihidrofolát-reduktáz
folsav, (a pteroil-glutaminsav vagy B 10 vitamin) 2 2 2 2 pirimidin rész pirazin rész aminobenzoesav rész glutaminsav rész pteridin rész dihidrofolsav 2 2 2 2 tetrahidrofolsav 2 2 2 2 A dihidrofolát-reduktáz
Fehérjeszerkezet, és tekeredés
 Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
DNS-szekvencia meghatározás
 DNS-szekvencia meghatározás Gilbert 1980 (1958) Sanger 3-1 A DNS-polimerázok jellemzői 5'-3' polimeráz aktivitás 5'-3' exonukleáz 3'-5' exonukleáz aktivitás Az új szál szintéziséhez kell: templát DNS primer
DNS-szekvencia meghatározás Gilbert 1980 (1958) Sanger 3-1 A DNS-polimerázok jellemzői 5'-3' polimeráz aktivitás 5'-3' exonukleáz 3'-5' exonukleáz aktivitás Az új szál szintéziséhez kell: templát DNS primer
Human genome project
 Human genome project Pataki Bálint Ármin 2017.03.14. Pataki Bálint Ármin Human genome project 2017.03.14. 1 / 14 Agenda 1 Biológiai bevezető 2 A human genome project lefolyása 3 Alkalmazások, kitekintés
Human genome project Pataki Bálint Ármin 2017.03.14. Pataki Bálint Ármin Human genome project 2017.03.14. 1 / 14 Agenda 1 Biológiai bevezető 2 A human genome project lefolyása 3 Alkalmazások, kitekintés
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Az örökítőanyag. I. A tulajdonságokat meghatározó örökítőanyag/információ átadható/ transzformálható.(fred Griffith kísérlet, 1928.
 Az örökítőanyag I. A tulajdonságokat meghatározó örökítőanyag/információ átadható/ transzformálható.(fred Griffith kísérlet, 1928.) Kísérlet: Tüdőgyulladást okozó pneumococcus baktériumok vizsgálata. S
Az örökítőanyag I. A tulajdonságokat meghatározó örökítőanyag/információ átadható/ transzformálható.(fred Griffith kísérlet, 1928.) Kísérlet: Tüdőgyulladást okozó pneumococcus baktériumok vizsgálata. S
Energiatermelés a sejtekben, katabolizmus. Az energiaközvetítő molekula: ATP
 Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Johann Gregor Mendel Az olmüci (Olomouc) és bécsi egyetem diákja Brünni ágostonrendi apát (nem szovjet tudós) Tudatos és nagyon alapos kutat
 és bécsi egyetem diákja Brünni ágostonrendi apát (nem szovjet tudós) Tudatos és nagyon alapos kutat") 10.2.2010 genmisk1 1 Áttekintés Mendel és a mendeli törvények Mendel előtt és körül A genetika törvényeinek újbóli felfedezése és a kromoszómák Watson és Crick a molekuláris biológoa központi dogmája 10.2.2010
10.2.2010 genmisk1 1 Áttekintés Mendel és a mendeli törvények Mendel előtt és körül A genetika törvényeinek újbóli felfedezése és a kromoszómák Watson és Crick a molekuláris biológoa központi dogmája 10.2.2010
Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete
 Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete Polipeptidek térszerkezete Tipikus (rendezett) konformerek em tipikus (rendezetlen) konformerek Periodikus vagy homokonformerek Aperiodikus
Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete Polipeptidek térszerkezete Tipikus (rendezett) konformerek em tipikus (rendezetlen) konformerek Periodikus vagy homokonformerek Aperiodikus
Tartalom. A citoszkeleton meghatározása. Citoszkeleton. Mozgás a biológiában A CITOSZKELETÁLIS RENDSZER 12/9/2016
 Tartalom A CITOSZKELETÁLIS RENDSZER Nyitrai Miklós, 2016 november 29. 1. Mi a citoszkeleton? 2. Polimerizáció, polimerizációs egyensúly 3. Filamentumok osztályozása 4. Motorfehérjék A citoszkeleton meghatározása
Tartalom A CITOSZKELETÁLIS RENDSZER Nyitrai Miklós, 2016 november 29. 1. Mi a citoszkeleton? 2. Polimerizáció, polimerizációs egyensúly 3. Filamentumok osztályozása 4. Motorfehérjék A citoszkeleton meghatározása
A CITOSZKELETÁLIS RENDSZER FUTÓ KINGA
 A CITOSZKELETÁLIS RENDSZER FUTÓ KINGA 2013.10.09. CITOSZKELETON - DEFINÍCIÓ Fehérjékből felépülő, a sejt vázát alkotó intracelluláris rendszer. Eukarióta és prokarióta sejtekben egyaránt megtalálható.
A CITOSZKELETÁLIS RENDSZER FUTÓ KINGA 2013.10.09. CITOSZKELETON - DEFINÍCIÓ Fehérjékből felépülő, a sejt vázát alkotó intracelluláris rendszer. Eukarióta és prokarióta sejtekben egyaránt megtalálható.
sejt működés jovo.notebook March 13, 2018
 1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
Molekuláris biológiai alapok
 Biokémiai és Molekuláris Biológiai Intézet Molekuláris biológiai alapok Sarang Zsolt Dimenziók a biológiában Fehérjék (kb. 50 ezer különböző fehérje a szervezetben 21 féle aminosavból épül fel) Élő szervezetek
Biokémiai és Molekuláris Biológiai Intézet Molekuláris biológiai alapok Sarang Zsolt Dimenziók a biológiában Fehérjék (kb. 50 ezer különböző fehérje a szervezetben 21 féle aminosavból épül fel) Élő szervezetek
A molekuláris biológia eszközei
 A molekuláris biológia eszközei I. Nukleinsavak az élő szervezetekben Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje DNS feladata: információ tárolása és a transzkripció
A molekuláris biológia eszközei I. Nukleinsavak az élő szervezetekben Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje DNS feladata: információ tárolása és a transzkripció
A piruvát-dehidrogenáz komplex. Csala Miklós
 A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
5. Előadás Nukleinsavak kimutatása, szekvenálás
 5. Előadás ukleinsavak kimutatása, szekvenálás A nukleinsav kimutatás etidiumbromid 3,8-diamino-5-etil-6-fenil-fenantrédiumbromid λ g =254-366 nm λ e =590 nm 2 2 + C25 Br - X + C3 C3 C3 C (C3)2 + (C2)3
5. Előadás ukleinsavak kimutatása, szekvenálás A nukleinsav kimutatás etidiumbromid 3,8-diamino-5-etil-6-fenil-fenantrédiumbromid λ g =254-366 nm λ e =590 nm 2 2 + C25 Br - X + C3 C3 C3 C (C3)2 + (C2)3
A géntechnológia genetikai alapjai (I./3.)
") Az I./2. rész (Gének és funkciójuk) rövid összefoglalója A gének a DNS információt hordozó szakaszai, melyekben a 4 betű (ATCG) néhány ezerszer, vagy százezerszer ismétlődik. A gének önálló programcsomagként
Az I./2. rész (Gének és funkciójuk) rövid összefoglalója A gének a DNS információt hordozó szakaszai, melyekben a 4 betű (ATCG) néhány ezerszer, vagy százezerszer ismétlődik. A gének önálló programcsomagként
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A citológia és a genetika társtudománya Citogenetika A kromoszómák eredetét, szerkezetét, genetikai funkcióját,
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A citológia és a genetika társtudománya Citogenetika A kromoszómák eredetét, szerkezetét, genetikai funkcióját,
Szerves Kémiai Problémamegoldó Verseny
 Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Genetika. Tartárgyi adatlap: tantárgy adatai
 Genetika Előadás a I. éves Génsebészet szakos hallgatók számára Tartárgyi adatlap: tantárgy adatai 2.1. Tantárgy címe Genetika 2.2. Előadás felelőse Dr. Mara Gyöngyvér, docens 2.3. Egyéb oktatási tevékenységek
Genetika Előadás a I. éves Génsebészet szakos hallgatók számára Tartárgyi adatlap: tantárgy adatai 2.1. Tantárgy címe Genetika 2.2. Előadás felelőse Dr. Mara Gyöngyvér, docens 2.3. Egyéb oktatási tevékenységek
Az enzimműködés termodinamikai és szerkezeti alapjai
 2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE
 ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE") Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
Elválasztástechnikai és bioinformatikai kutatások. Dr. Harangi János DE, TTK, Biokémiai Tanszék
 Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
A felépítő és lebontó folyamatok. Biológiai alapismeretek
 A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
Gyógyszerrezisztenciát okozó fehérjék vizsgálata
 Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
BIOMOLEKULÁK KÉMIÁJA. Novák-Nyitrai-Hazai
 BIOMOLEKULÁK KÉMIÁJA Novák-Nyitrai-Hazai A tankönyv elsısorban szerves kémiai szempontok alapján tárgyalja az élı szervezetek felépítésében és mőködésében kulcsfontosságú szerves vegyületeket. A tárgyalás-
BIOMOLEKULÁK KÉMIÁJA Novák-Nyitrai-Hazai A tankönyv elsısorban szerves kémiai szempontok alapján tárgyalja az élı szervezetek felépítésében és mőködésében kulcsfontosságú szerves vegyületeket. A tárgyalás-
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
A CITOSZKELETÁLIS RENDSZER (Nyitrai Miklós, )
") A CITOSZKELETÁLIS RENDSZER (Nyitrai Miklós, 2010.11.30.) 1. Mi a citoszkeleton? 2. Polimerizá, polimerizás egyensúly 3. ilamentumok osztályozása 4. Motorfehérjék Citoszkeleton Eukariota sejtek dinamikus
A CITOSZKELETÁLIS RENDSZER (Nyitrai Miklós, 2010.11.30.) 1. Mi a citoszkeleton? 2. Polimerizá, polimerizás egyensúly 3. ilamentumok osztályozása 4. Motorfehérjék Citoszkeleton Eukariota sejtek dinamikus
A metabolizmus energetikája
 A metabolizmus energetikája Dr. Bódis Emőke 2015. október 7. JJ9 Miért tanulunk bonyolult termodinamikát? Miért tanulunk bonyolult termodinamikát? Mert a biokémiai rendszerek anyag- és energiaáramlásának
A metabolizmus energetikája Dr. Bódis Emőke 2015. október 7. JJ9 Miért tanulunk bonyolult termodinamikát? Miért tanulunk bonyolult termodinamikát? Mert a biokémiai rendszerek anyag- és energiaáramlásának
A BIOTECHNOLÓGIA TERMÉSZETTUDOMÁNYI ALAPJAI
 A BIOTECHNOLÓGIA TERMÉSZETTUDOMÁNYI ALAPJAI Műszaki menedzser MSc hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: március 06?, április 10?, május 02?. dr. Pécs Miklós egyetemi docens
A BIOTECHNOLÓGIA TERMÉSZETTUDOMÁNYI ALAPJAI Műszaki menedzser MSc hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: március 06?, április 10?, május 02?. dr. Pécs Miklós egyetemi docens
10. Előadás. Heterociklusos vegyületek.
 0. Előadás eterociklusos vegyületek. ETECIKLUSS VEGYÜLETEK Felosztás:. telített telítetlen. heteroatomok száma 3. gyűrűk száma. heteroatomok milyensége (,, S, P, As, Si) oxa- S tia- aza- I. Monociklusos,
0. Előadás eterociklusos vegyületek. ETECIKLUSS VEGYÜLETEK Felosztás:. telített telítetlen. heteroatomok száma 3. gyűrűk száma. heteroatomok milyensége (,, S, P, As, Si) oxa- S tia- aza- I. Monociklusos,
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció 5.
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció 5.