Szerk.: Vizkievicz András A DNS örökítő szerepét bizonyító kísérletek
|
|
|
- Elemér Papp
- 9 évvel ezelőtt
- Látták:
Átírás
1 Az öröklődés molekuláris alapjai Szerk.: Vizkievicz András A DNS örökítő szerepét bizonyító kísérletek A DNS-nek addig nem szenteltek különösebb figyelmet, amíg biológiai kísérlettel ki nem mutatták, hogy bizonyos baktériumok öröklődését megváltoztatja. Ezt a kísérletet egyáltalán nem tervezték meg előre ban Fred Griffith egy gömb alakú baktérium - Diplococcus pneumoniae - kórokozó képességét patogenitását - vizsgálta, amely egérben vérmérgezést, emberben tüdőgyulladást okoz. E baktériumnak két törzse ismert. 1. Patogén (sima felszínű, S-törzs), amelynek egyedei az immunrendszernek ellenálló poliszacharid tokot fejlesztenek maguk köré. A tokképzés örökletes, azaz az utódok is tokosak lesznek. 2. Nem patogén, amelyek nem képeznek tokot. (Ráncos felületű, R- törzs, az immunrendszer meg tudja semmisíteni) Griffith a baktériumokat egerekbe oltotta, majd a következő eredményeket kapta: 1. Egér + élő tokos baktérium = az egerek 1-2 napon belül elpusztultak. 2. Egér + élő tok nélküli baktérium = az egerek nem pusztultak el. 3. Egér + hővel elölt tokos baktérium = az egerek nem pusztultak el. 4. Egér + hővel elölt tokos + élő tok nélküli baktérium = néhány egér elpusztult! Az elpusztult állatok hulláit megvizsgálták, és meglepődve tapasztalták, hogy bennük élő tokos baktériumok voltak. Griffith tehát azt a váratlan felfedezést tette, hogy az elölt patogén sejtek az élő, nem fertőző sejtekhez keverve, azoknak kis részét patogénné tették. Fertőzővé válva az eredetileg nem fertőző sejtek, tokot fejlesztettek, amely az immunrendszerrel szemben ellenállóvá teszi azokat a sejteket, amelyeken megtalálható. Griffith tehát az örökítő anyag létét fedezte fel, amely ép maradt a fertőző sejteket ért pusztító hőhatás után, s később átkerülve a nem kórokozó sejtekbe, azokat kapszula képzésére serkentette. Ezt a folyamatot genetikai transzformációnak nevezzük. Griffith szerint a baktérium - általa nem ismert örökítőanyaga okozta a jelenséget, ugyanakkor maga nem próbálta mindenáron meghatározni ezt az anyagot. 1

2 A Griffith által felfedezett természetes baktérium transzformáció az alapja a modern mesterséges genetikai transzformációnak, melynek során egy sejtbe valamilyen idegen DNS-t juttatnak azzal a szándékkal, hogy a bekerült örökítőanyag megváltoztatja a befogadó recipiens - sejt tulajdonságait. A bejutott DNS kifejeződik, azaz az általa kódolt fehérjét gyártja le a sejt, amelynek működése átalakítja a befogadó sejt működését. Ez az eljárás az alapja a genetikai mérnökségnek, régiesen génsebészetnek. A mesterséges baktérium transzformáció során a bejutatott DNS által kódolt fehérje termeltetése a cél, mely különféle módon kinyerve felhasználhatóvá válik, mint pl. az E. coli által termelt inzulin. Az eljárás első lépése a kívánt DNS-szakasz bejuttatása a befogadó sejtbe. Ez többféleképpen történhet pl. transzdukcióval vírusok közvetítésével vagy plazmidok segítségével transzformációval, génágyúval. (A DNS- t néhány μm átmérőjű arany vagy wolframrészecskére rögzítik, majd ezeket a részecskéket nagy sebességre felgyorsítják, nagy nyomású He vagy N 2 gázzal. A részecskék eltalálják a célsejtet, áthatolnak annak a falán és bejutatják a szállított DNS-t.) Ezeket a bejuttatandó hasznos DNS-egységeket genetikai vektoroknak nevezzük. A mesterséges transzformáció során először ún. hibrid rekombináns - plazmidot hoznak létre. A hibrid plazmidot úgy állítják elő, hogy egy baktériumból kinyert természetes plazmidba beépítik a bejuttatandó DNS szakaszt. Ezt követően a befogadó sejtet arra késztetik, hogy vegye fel az elkészített hibrid plazmidot. A technológia kivitelezését a restrikciós endonukleázok felfedezése tette lehetővé. Nevüket arról kapták, hogy megakadályozzák az idegen DNS, például a baktériumot fertőző vírus DNS-ének invázióját, a molekula feldarabolásával (restrict = korlátozni). Az endonukleáz megjelölése, pedig arra utal, hogy az idegen DNS-t a molekula belsejében levő helyeken hasítják el, és nem a végén kezdik lebontani (görög: endo=belső). A restrikciós endonukleázok nem pontosan szemben vágják el a DNS két szálát, hanem kissé eltolódva, tehát egyfonalas túlnyúló szálakat, ún. ragadós végeket képeznek, ami megkönnyíti két DNS összekapcsolását rekombináns DNS-sé. A genetikai vektorok kialakításához szükség van még az ún ligáz enzimre, amely képes összekapcsolni különböző DNS-ekből származó szabad végű DNS-darabokat, létrehozva a hibrid plazmidot. Ezek után biztosítani kell, hogy a hibrid plazmidok bejussanak a baktériumokba. Egyik lehetőség, hogy a baktériumokat és a plazmidokat kalcium-klorid oldatba helyezik, majd felváltva hideg és meleg sokknak teszik ki őket. A baktériumokra kifejlesztett módszerek módosításokkal ugyan, de használhatónak bizonyultak fejlettebb élőlényekkel kapcsolatban is. Így jöttek létre a genetikailag módosított organizmusok, azaz a GMO-k (Genetically Modified Organisms). 2

3 A GMO olyan élő szervezet, amelynek genetikai anyaga a modern géntechnológia alkalmazásával olyan új genetikai kombinációkkal rendelkezik, amely a természetben nem található meg. Az GMO előállításának módja a genetikai transzformáció. Az 1. generációs GMO tápláléknövények a növény, s így a termesztők szempontjából hasznos géneket fejeznek ki, melyek ellenállóvá teszik a növényt o a szárazság, o a hideg, o a patogének (rovarok, baktériumok, gombák és vírusok), vagy a o gyomirtószereik ellen. Pl. a kukoricába olyan géneket ültettek, amelyek a gyomirtó szerek ellen ellenállóvá teszik a növényt, s ezért a kukoricaföldeken kémiailag védekezhetünk a gyomok ellen anélkül, hogy ezzel károsítanánk a kukoricát. Továbbá beültették a Bacillus thüringiensis nevű baktérium faj rovarölő génjét (Bt toxin), amely a kártevők ellen teszi rezisztensé a növényt, hiszen kártevő elpusztításához szükséges vegyszert maga a növény termeli. A 2. generációs GMO-k olyan gazdasági haszonnövények, amelyek a fogyasztók számára hasznos anyagot termelnek nagyobb mennyiségben. o Ilyenek pl. az olajsav összetételében megváltoztatott repce és szója, a ferritin (vasraktározó fehérje) tartalmú saláta, stb. A 3. generációs GMO-k bioreaktoroknak, ill. gyáraknak tekintetők, mert olyan anyagokat termeltetünk velük, amelyeket hagyományosan az ipar produkál: o kórokozók elleni vakcinákat, hormonokat (pl. inzulin), véralvadásban szerepet játszó anyagokat stb. Genetikailag módosított haszonállatok, mint pl. o o o o gyorsabban növekvő lazac, több kazein fehérjét kifejező transzgénikus tehén, különféle emberi transzgéneket a májában kifejező sertés, abból a célból, hogy a mája átültethető legyen emberbe. Argentin tudósok emberi anyatejet adó szarvasmarhát állítottak elő. Milyen előnyök származtathatnak a GMO növényekből. Csökkentik a növények, állatok, emberek, környezet növényvédőszer terhelését. Megtakaríthatók a permetezési és vegyszer-költségek. Mivel a Bt-toxint a növény termeli, a nem kártevő ízeltlábúak között sokkal kevesebb kárt okoz, mint a hagyományos, vegyszeres rovarirtás. A gyomirtás totális gyomirtószerekkel könnyen megoldható. Csökkentik az öntözővíz, talajjavító anyag és műtrágya szükségletet. Bioszintetizátor élőlények: gyógyszer-, hormon-, enzim-, és szervtermelő állatok. Ellenérvek. A rezisztens növények elszabadulása a természetben, a transzgén megszökése. A szél által terjesztett pollen a szomszédos, hagyományos állományokban is termékenyít, ez a hagyományos fajták termelőinek és különösen a biotermesztőknek gazdasági kárt okoz, mivel az ő termékeiket is GM-tartalmúnak kell megjelölni. Abban az esetben, ha a GM-növénybe antibiotikumrezisztencia-gént is beültettek, a rezisztenciáért felelős gén(ek) átkerülhetnek az emberek és állatok szervezetében élősködő baktériumokba és azok ellenállóvá válhatnak az antibiotikummal szemben, hatástalanná téve az antibiotikumok orvosi alkalmazását és számos betegség gyógyítását. Mérgezőképesség emberre és állatra, allergia kiváltása, új allergének megjelenése. A Bt toxint a növény minden sejtje termeli, mennyisége kb szerese a hasonló permetezőszer egyszeri ajánlott dózisának. A talajba szántott növényi részekben lévő toxin lassabban bomlik el, mint a hasonló permetezőszer toxinja. Alig ismerjük a genetikailag módosított szervezetek környezeti, ökológiai, egészségügyi, társadalmi hatásait és kockázatait. 3
, vagy a o gyomirtószereik ellen. Pl.")
4 Az örökítő anyag meghatározását Oswald Avery végezte el 1941-ben. Avery kísérleteiben a transzformációt előidéző baktérium kivonatot előzetes kémiai tisztítási eljárásnak vetették alá. Azt találták, hogy a baktérium kivontaból poliszacharidok, (amiláz) lipidek (lipáz), proteinek (proteáz) és ribonukleinsav (RNáz) vonható ki anélkül, hogy a transzformáció mértéke lényegesen csökkenne. Azonban amikor DNS-bontó enzimekkel DNáz - kezelték a kivonatot a transzformáció elmaradt. Tehát a transzformációért felelős anyag minden kétséget kizáróan a DNS, mivel a transzformáló képességet DN-áz készítménnyel való kezelés megszüntette ben Alfred Hershey és Martha Chase baktériumokat fertőző és pusztító vírussal, ún. bakteriofágokkal végeztek vizsgálatokat. Mint minden vírus, a fágok is alapvetően két részből állnak: a központi információtároló DNS-ből, és az ezt körülvevő fehérjetokból, a kapszidból. Röviden a fertőzés úgy történik, hogy a fág hozzátapad a baktérium felületéhez, egy enzim feloldja a baktériumsejt falát, s az így keletkezett nyíláson a DNS behatol a baktériumsejtbe. A fág burka a sejten kívül marad. A bejutott fáganyag csakhamar elszaporodik a sejt anyag és energiakészletét felhasználva. A fágok érése után a már halott baktériumsejt felreped, és mintegy 1OO-5OO új, fertőzőképes fág szabadul ki belőle. 4

5 Hershey és Chase a kólibaktérium (Escherichia coli) ún. T2- fágját tanulmányozta. Ezzel a fággal olyan baktériumokat kezeltek, melyeket előzőleg radioaktív foszfort ( 32 P) tartalmazó foszforsav ill. radioaktív ként ( 35 S) tartalmazó MgSO 4 tartalmú táptalajon tartottak. A baktériumsejtből kiszabaduló érett fágok a fenti izotóppal jelölődtek, a radioaktív kén a fehérjékbe, a radioaktív foszfor a DNS-be épült be. Az így jelölt fágokkal kólisejteket fertőztek. Rövid idő után rázással a fágokat eltávolították a baktérium felületéről, majd megmérték a baktériumtömeg aktivitását. Azt tapasztalták, hogy a radioaktív foszfort tartalmazó bakteriofágok által megfertőzött baktériumok mutatták a radioaktivitást, míg a radioaktív kénnel megfertőzött baktériumok a rázást követően nem. Ebből egyértelműen következik, hogy a fág-dns nagy része bejut a baktériumsejtbe, a fehérje nagy része pedig kint marad. A baktérium szétesése után a kikerülő fágok 35 S-t nem, de 32 P-t nagy mennyiségben tartalmaztak. Miután egy sejtet egy fág fertőz meg, de ugyanakkor a sejt pusztulásakor több száz fág is keletkezhet, a vírus-dns-nek valamilyen módon sokszorozódnia kell. E problémára Matthew Meselson és Franklin Stahl ban megdönthetetlen bizonyítékokkal alátámasztott kísérlettel adták meg a választ. A kísérlet során kóli baktériumokat tenyésztettek több generáción át 15 N, tehát nehéz nitrogén-tartalmú tápoldatban. A baktérium DNS-ébe bekerült a 15 N izotóp, vagyis a sejtek nehéz DNS-t tartalmaztak. Majd ezt követően átoltották a baktériumok egy részét normál, 14 N- es tápoldatba. Egy sejtosztódás után mintát vettek, izolálták a DNS-t, és meghatározták a sűrűségét. Ugyanezt megtették a második osztódást követően is. Az első osztódás után csak közepes sűrűségű DNS-t kaptak. A második sejtosztódást követő eredmény szerint a mintában 50-50%-ban volt jelen közepes sűrűségű, és könnyű, azaz normális sűrűségű DNS. A kísérlet eredménye a DNS megkettőződésének replikációjának szemikonzervatív mechanizmusával értelmezhető. A DNS-molekulában észlelhető szabályosság a bázisok komplementaritása képezi az alapját a genetikai információ egyértelműen pontos másolásának, a replikációnak. A replikáció során az egymástól különváló két szál egy-egy újabb DNS-szál szintézisének mintájaként, templátjaként szolgál. új II. szál 5'... T-T-C-A-G-G-T-C...3' régi I. szál 3'... A-A-G-T-C-C-A-G...5' régi II. szál 5'...T-T-C-A-G-G-T-C...3' új I. szál 3'... A-A-G-T-C-C-A-G...5' 5

6 A DNS bázissorrend állandóságának alapja a DNS-szintézis szemikonzervatív jellege. Ez annyit jelent, hogy a replikáció során elváló DNS-láncok templátként szolgálnak az új láncok szintéziséhez. Az utód DNS-molekulák egyik lánca a szülői DNS-ből származik, a másik komplementer lánc újonnan szintetizálódik hozzá. (szemikonzervatív félig megmaradó - jelleg) A replikáció mechanizmusa A replikáció a DNS-szál szigorúan meghatározott részén, az ún. kezdő iniciációs - ponton indul meg, amit replikációs origónak nevezünk. 1. A másolást megelőzően a kromoszóma szakaszhoz kicsavaró fehérjék helikázok - kapcsolódnak. Kettős hélix alakban a replikciót meghatározó bázisok kevéssé hozzáférhetők, mivel a kettős hélix centrális részében helyezkednek el. A folyamatban résztvevő enzimek felszakítják a bázisok közötti H-kötéseket, aminek következtében a DNS-szakasz átmenetileg kinyílik, a DNS-szálak elválnak egymástól. E. coli-ban kb kicsavaró fehérje működik egyszerre. A kialakult egyfonalas DNS-szakaszokat stabilizálni kell. Az egyfonalas szakaszokhoz egyszálú DNS-t kötő fehérjék kapcsolódnak, amelyek megakadályozzák a kettős hélix formába való visszarendeződést. A szabaddá vált DNS-szakaszon a DNS-függő RNS polimeráz segítségével megindul a DNS-száléval komplementer, ribonukleotid egységből RNS-primer szintézise. 2. A primer RNS 3'-OH végéhez a DNS polimeráz-iii lépésenként dezoxiribonukleotidtrifoszfátokat kapcsol. A folyamat energia igényét a pirofoszfátok hidrolízise biztosítja. Az új szál tehát 5' 3' irányba nő. Ahogy az új szál hosszabbodik, a kicsavaró fehérjék elválasztják a komplementer szálakat, kialakítják a replikációs villa újabb szakaszait. Mivel a DNS két szála ellentétes polaritású úgy szintézisük is ellentétes irányba történik, hiszen a szintézis iránya mindig 5 3. Minthogy a kicsavaró fehérjék csak az egyik irányba haladnak, azaz a replikációs villa csak az egyik irányba nyílik szét, csak az egyik szál szintézise lehet folyamatos, az, amelynek növekedési iránya megegyezik a helikázok haladási irányával. Ebből következik, hogy a másik ellenkező irányba növő szál szintézise csak szakaszosan mehet végbe. (Okazaki-fragmentek) Az ellenkező irányba növekvő szálon a szintézis RNS nukleotidegység összekapcsolásáig tart. Az RNS primert a polimeráz I távolítja el, majd helyére komplementer dezoxiribonukleotidokat épít (kitölti a hézagokat). 6

7 3. Az egyes DNS-szakaszokat (fragmenteket) a ligáz kapcsolja össze úgy, hogy két nukleotidegység között foszfodiészter kötés kialakulását katalizálja. A DNS megkettőződése bonyolult, több lépésből álló folyamat, melynek lebonyolításában több enzim vesz részt. Az E. coli félóránként kettőződik, vagyis ezalatt a sejtnek újabb teljes DNS-t kell szintetizálnia. Tehát a folyamat igen gyors, a kb. 4 millió bp.-ból álló E.-coli DNS nagyjából 20 perc alatt lemásolódik. Ez azonban csak akkor történhet meg, ha a felnyílás és a másolás egy időben egyszerre több helyen indul meg. A replikáció pontossága A tulajdonságok megfelelő öröklődése szempontjából rendkívül fontos, hogy az örökítő anyagot a sejtek pontosan lemásolva adják át egymásnak. Ennek biztosítéka egyrészt a másolás pontossága, másrészt a replikáció során történt hibák kijavítása. E. coli DNS replikációja során átlagosan egyetlen hiba fordul elő 10 9 bázis beépülésekor. Az emberi genom haploid sejtenkénti DNS állomány 3 milliárd bázispárból épül fel, tehát replikációjakor átlagosan 3 hiba történik. A replikáció ilyen pontosságáért a hibajavító mechanizmusok felelősek. A replikációban bekövetkező hibák korrigálásáért a DNS polimeráz I felelős. A DNS polimeráz I felismeri a deformált szakaszt, majd a hiba mellett hasadékot létesít a DNS-szálon. A polimeráz I megfelelő nukleotidokat épít a hibásak helyébe 5' 3' irányba folyó szintézissel. Az enzim 5' 3' nukleáz aktivitással kihasítja a hibás szakaszt. A DNS ligáz az új szálat a régivel összeköti. A DNS polimeráz korrigáló működésének a hiánya emberben, pl. a xeroderma pigmentosum nevű igen súlyos betegség kialakulásához vezethet. A betegek bőre rendkívül érzékeny a napfényre ill. az UV sugárzásra. A bőr igen száraz, a szemlencse szarusodik, rendszerint több helyen bőrrák alakul ki, aminek következtében a betegek már a 30.-életévük előtt meghalnak. Az UV sugárzás a bőrben timindimereket hoz létre. A pirimidinbázisok UV fénnyel történő megvilágítás hatására egymással kovalensen összekapcsolódnak. Ezek nem illeszkednek be a kettős helixbe, megállítják a replikációt mindaddig, amíg a polimeráz I ki nem hasítja a hibás szakaszt és helyére megfelelőt nem épít. Egészséges emberben a repair mechanizmus a dimereket kihasítja a DNS-ből. A betegekben a javítórendszer hiányzik, a dimerek bennmaradnak a DNS-szálban. A hibás DNS kijavításának hiányával összefüggő, ritka betegségek mind korai halált okoznak: mint pl. a Gilford-szindróma, amely a gyermekkorban az aggkorra jellemző tünetek kialakulása jellemző. Transzkripció - átírás Az élő szervezetek bármilyen életjelensége - anyagcsere, mozgás, stb. - fehérjék működésén keresztül valósul meg. A különböző funkciókért specifikus fehérjék felelősek. Specifitásuk az aminosav sorrendjükben rejlik. Az aminosavsorrend információja a DNSmolekula bázissorrendjével hozható összefüggésbe. Eukariótákban az információt hordozó DNS a sejtmagban helyezkedik el, ugyanakkor a fehérjék szintézise a citoplazmában folyik. A DNS tehát közvetlenül nem szolgálhat mintaként (templátként) a fehérjék szintéziséhez. Ebből az következik, hogy a DNS az információt egy közvetítő molekulának adja át, amely kijutva a citoplazmába irányítja a fehérjék szintézisét. Ezt a szerepet a hírvivő, vagyis a messenger RNS (mrns) látja el. 7

8 A DNS információjának RNS-molekulára átírását transzkripciónak nevezzük. E folyamattal nemcsak az mrns szintetizálódik, hanem az összes RNS is (trns, rrns). A sejten belül az információáramlás a következő úton halad (centrális dogma): DNS transzkripció RNS transzláció fehérje tulajdonság Az információtároló DNS és az információt közvetítő RNS közötti kémiai különbség csekély: DNS dezoxiribóz timin kétszálú hélix RNS ribóz uracil egyszálú Az RNS-ek szintézisét DNS-templáton az RNS polimeráz katalizálja, melynek feladata, hogy a nukleotidtrifoszfát egységeket lépésenként összekapcsolja a mintául szolgáló DNS bázissorrendjének megfelelően. Mechanizmus 1. Kezdés (Iniciáció). Az RNS polimeráz felismeri és hozzá kötődik az átírás kezdőpontjához a promoterhez. A kapcsolódáskor DNS két szála elválik egymástól, majd az értelmes szálon megindul az átírás a bázisok párosodási szabályainak megfelelően. A = U, G = C 2. Folytatás (Elongáció). A startjeltől kezdve az RNS polimeráz leolvasva a DNS értelmes szálát, a ribonukleotidokat foszfodiészter kötéssel kapcsolja egybe. A DNS kettős hélixnek csak az egyik szála íródik át (értelmes szál), a másik soha (néma szál). Az értelmes szál DNS-szakaszonként változhat, de egy adott génre mindig állandó. A kialakuló RNS-szál átmenetileg összekapcsolódik a DNS templáttal, majd miután azonban az RNS-DNS kettős hélix labilisabb, mint a DNS-DNS kettős, a polimeráz továbbhaladása után a DNS kettős hélix újra záródik, az RNS leválik. 3. Befejezés (Termináció). A szintézis addig tart, amíg az RNS polimeráz eléri a szintézis befejezését jelentő stopjelt (szignált). A lánc vége előtt egy U-ban gazdag szekvencia található, amely az átírást követően egy hajtűhurkot palindróma - kialakítva megakasztja az enzimet, miáltal az átírás befejeződik. 8
.")
9 A prokariotáknál a transzkripció és a fehérjeszintézis csaknem szimultán folyamat, az mrns 5' vége még jóval a lánc szintézisének a befejezése előtt hozzákapcsolódik riboszómákhoz, s ezen a szakaszon már megkezdődik a fehérjeszintézis. Eukariotáknál az RNS polimeráz hatására kialakuló poliribonukleotid még nem teljes értékű végtermék, biológiai feladatának betöltése érdekében többféle átalakuláson, érésen kell keresztül mennie, mielőtt kikerül a citoplazmába. Az RNS érési folyamatai eukariótáknál (poszttranszkripciós módosítások) 1. Az mrns két végéhez különböző szerkezeti elemek kapcsolódnak. 2. Az elsődlegesen elkészült mrns-ből különböző méretű szakaszok hasadnak ki. Ez a splicing. 1. Az mrns két végéhez különböző szerkezeti elemek kapcsolódnak: védősapka, cap-képződés az 5'-végen. Poli-A farok a 3'-végen. A mrns-nek nemcsak a szintézise, hanem a lebontása is az 5'-végen kezdődik. A sejtben levő nukleázokkal szemben az ún. sapka védi az 5 véget. A cap felépítésére jellemző, hogy az utolsó bázis gyakran G, aminek 7- helyzetű N-je metilálva van (m7g), így ez extra + töltést visel, ami elektromos taszítást eredményez. Közvetlenül a transzkripció után, az RNS 3'-végéhez kb adenin nukleotidot tartalmazó poli-a farok kapcsolódik a fenti okok miatt. Az eukarióta gének néhány tulajdonsága Génnek nevezzük a DNS-molekula adott szakaszát, amely valamilyen öröklődő jelleg kialakulását megszabja egy adott polipeptidlánc kódolásán keresztül. A fejlődés különböző szintjein álló szervezetek DNS-ének információtartalma eltérő. Vírusok esetén a DNS túlnyomó része hordoz információt, sőt átfedés folytán egy-egy szakasz több információt is tartalmazhat. Prokariotákban a DNS zöme átíródik, csak elenyésző része nem. Eukariótákban azonban a DNS-nek csupán néhány %-a hordoz információt, másik nem nagy része valamilyen egyéb feladatot tölt be és tekintélyes részének jelenleg a funkcióját nem ismerjük. A haploid egyszeres kromoszómakészletű - sejtek teljes genetikai állománya a genom. Az eukarióta genom szerveződésére jellemző, hogy a genom mérete sokkal nagyobb, mint a róla átírt, érett RNS-é. Ez azt jelenti, hogy az információ a DNS-ben nem kontinuus. Emberben ma kb gén létét feltételezik, aminek mérete a teljes genom 1-2 %-a, azaz a gének kisebb nagyobb távolságra találhatók egymástól és őket át nem íródó szakaszok választják el. A genom 98 %-ának a funkciója ma még pontosan nem ismert, nagy részük a gének átíródási kifejeződési folyamatait irányítják, hiszen egy adott sejten belül a teljes génállomány töredéke fejeződik ki. 9
 1. Az mrns két végéhez különböző szerkezeti elemek kapcsolódnak. 2.")
10 2. Az mrns hasítása - splicing. Sőt ezen túlmenően a gének tartalmaznak intron (intragén) szakaszokat, aminek átírási terméke az érett messengerből kivágódik, és tartalmaz exonokat, amelyek a gének kifejeződő szakaszai. Az exonokat egymástól nukleotidpárból álló intron szakaszok választják el. A kétféle DNS-szakasz aránya a genomban változatos, az intron mennyisége gyakran többszöröse az exonénak. A gén átírása folyamatos, a keletkező elsődleges átiratból egymást követő lépésekben hasadnak ki az üzenetet nem tartalmazó intron szakaszok és a szabadon maradt végek egymással összeillesztődnek. A folyamatot splicingnak nevezik. A vágó-illesztő folyamat mechanizmusának rendkívül pontosnak kell lennie, egy nukleotidot sem tévedhet, mert teljesen megváltozna az információ tartalom. A DNS-ről tehát transzkripcióval egy igen rövid életidejű elsődleges átirat keletkezik, amelyet régebben heterogén nukleáris RNS-nek, röviden hnrns-nek neveztek. Ezt követően felkerül a cap és a poli-a farok. Majd ezek után következik a splicing, azaz az intronok kihasítása. A splicing mechanizmusáért RNS-ből és egy sor fehérjéből álló kis komplex gömböcskék, ún. kis sejtmag ribonukleoproteinek - small nuklear ribonucleoprotein, röviden snrnp felelősek. Az snrnp ejtése: sznörp, amely a hupikék törpikék néven ismert rajzfilmfigurák -angolul smurfs - nevére rímelő lefordíthatatlan szójáték. A genetikai információ realizálódása - transzláció: a fehérjék bioszintézise Ha a DNS az élet tervrajza, akkor ehhez a fehérjék a tégla és a habarcs. De valójában ők jelentik a sejt vagy az élő szervezet összeszerelésekor az állványt és a szerszámokat, sőt ők azok a munkások, akik a kivitelezést végzik. A genetikai információt a DNS-molekula hordozza. Az információ továbbításáért az mrns-molekulák felelősek. A kérdés az, hogy a DNS-ben foglalt egydimenziós információ - nukleotid sorrend - a különféle RNS-molekulák aktív részvételével hogyan határozza meg a specifikus fehérjemolekulák aminosav sorrendjét, keletkezését. A választ a genetikai kód adja meg, amely egy olyan szabályrendszer, kódolási eljárás (nem anyagi egység), ami szerint a nukleinsavakban szereplő szöveg lefordítható a fehérjékben szereplő szövegre. Ennek a szabályrendszernek a feltárása a biológiai kutatások körében az egyik legnagyobb intellektuális teljesítmény volt. A kodonszótár megfejtése (emelt szintű érettségi feladat) A kodonszótár megfejtéséért Robert Holley, Har Ghobind Khorana és Marshall Nierenberg 1968-ban kapott Nobel-díjat. Tervszerű kísérletekkel igazolták a kapcsolatot az mrns bázishármasok és a fehérjék aminosav sorrendje között. 10

11 Nierenberg kísérletsorozatának első részében olyan mesterséges mrns-t szintetizáltak, amely csupa uracilbázist tartalmazott (poli-u). Ennek segítségével a riboszómákon egy kizárólag fenilalanin aminosavakból álló lánc jött létre. Ez a kodonszótár első megfejtett szava: UUU > Phe (fenilalanin). A kutatók ezután különböző koncentrációjú nukleotidelegyeket alkalmaztak, és figyelték a termékként kapott polipeptidben az aminosavak arányát. Ha például az elegyben csak uracilt (U) és citozint (C) tartalmazó nukleotidok vannak 2:1 arányban, ezek gyakorisága p (U) = 2/3 és q (C) = 1/3. Az egyes bázishármasok keletkezésének valószínűsége föltételezve, hogy kapcsolódásuk véletlenszerű az elemi valószínűségek szorzata lesz. Így például az UUC bázishármas ebben az elegyben x(uuc) = p(u) 2.q(C) = (2/3) 2. (1/3) = 4/27 valószínűséggel keletkezik, s mivel a létrejövő fehérjeláncban éppen ekkora volt a fenilalanin relatív gyakorisága, megállapítható, hogy ez a bázishármas is a fenilalanint kódolja. A genetikai kód A nukleinsavak ABC-je 4 betűs, a fehérjéké 20. Egyetlen nukleotid tehát nem lehet jel kodon -, mert így csupán 4 aminosav beépülését lehetne szabályozni. Ha két-két nukleotid alkotna egy-egy jelet, akkor is csak 4 2 = 16-féle kodon alakulhatna ki. A 3 nukleotidból álló egységek már 4 3 = 64-féle kombinációt hoznak létre, ami bőségesen elegendő a 20-féle aminosav kódolásához. A genetikai információ egységeit, tehát DNS- vagy RNS-nukleotid hármasok ún. tripletek alkotják. Ezeket kodonoknak nevezzük. A genetikai kód jellemzői 1. Degenerált, azaz egy aminosavat több triplet határoz meg, hiszen 20 aminosav beépülésére 64 kodon áll rendelkezésre. A leggyakoribb aminosavakat 4-6 kodon is jelöli, pl. az Arg-nak, Ser-nek, Leu-nak 6, a ritkábban előfordulókat viszont csak 1-2, ilyen pl. a Try és a Met. Az AUG-kodon a transzláció start jele (egyben a formil-metionin (21. aminosav) kodonja). Három kodon jelzi a fehérjeszintézis végét, ezeket a tripleteket stop-kodonnak nevezzük (ezek nem kódolnak aminosavakat). A stop jeleknek nincsen, a start jelnek viszont kettő trns-ük is van, így a 64-féle kodonhoz csak 62-féle trns tartozik általában. 2. Univerzális, mivel a genetikai kód a szervezetek fejlettségi szintjétől függetlenül egységes, minden élőlényre igaz (kivéve a mitokondriális DNS-t). 3. Átfedés- és kihagyásmentes, azaz egyetlen nukleotid sem lehet tagja két szomszédos kodonnak. 11
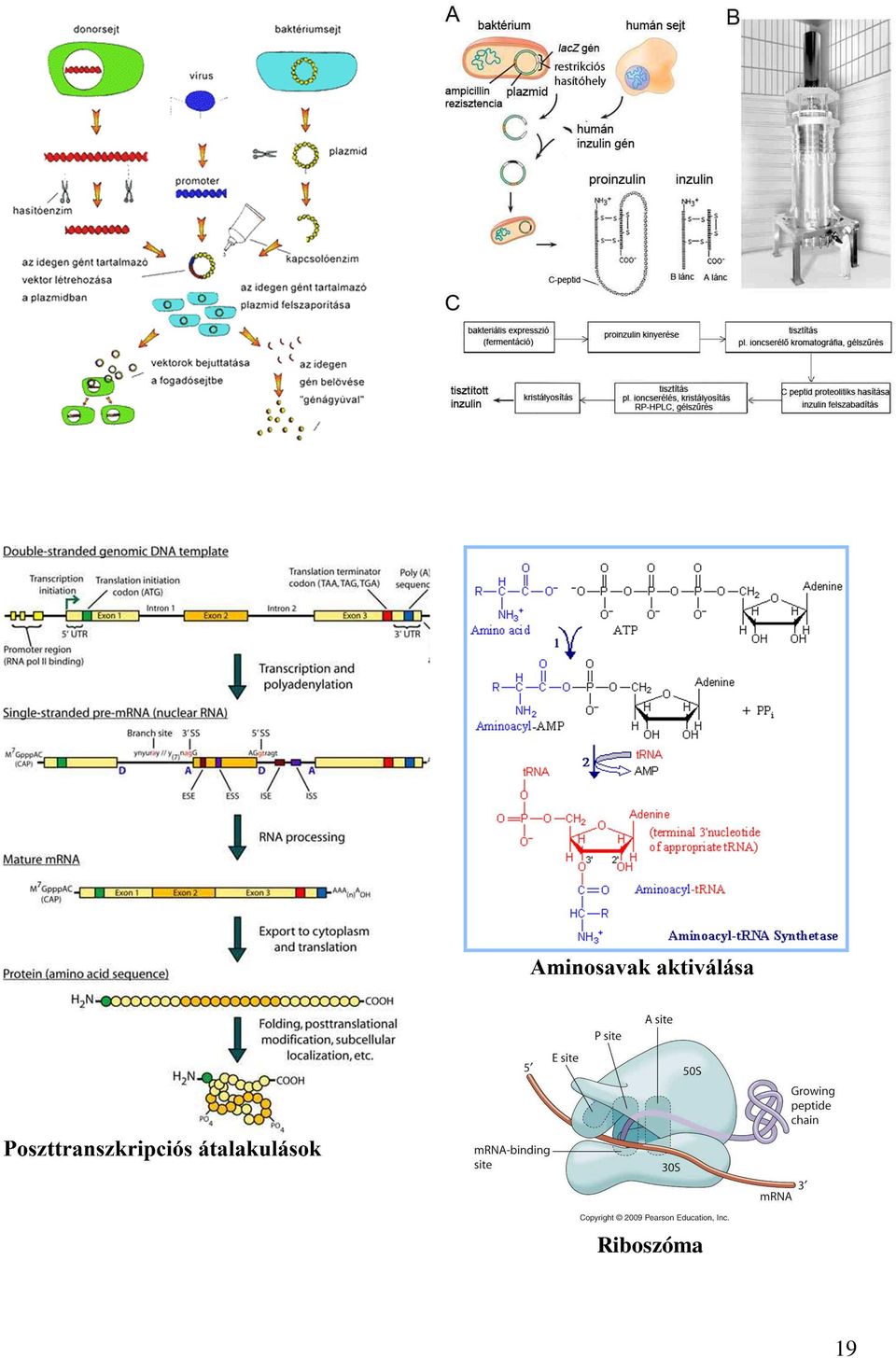
 és citozint (C) tartalmazó nukleotidok vannak 2:1 arányban, ezek gyakorisága p (U) = 2/3 és q (C) = 1/3.")
12 Ha egy vagy két nukleotid kiesik vagy beékelődik, az eltolja a leolvasási keretet, és ezzel értelmetlen aminosavsorrendet hoz létre, ami teljesen működésképtelen fehérjét eredményez. (frame shift mutáció) A transzláció mechanizmusa Az mrns kodonjai közvetlenül semmilyen módon nem hozhatók kapcsolatba az őket kódoló aminosavakkal, mivel szerkezetük nem komplementer. Ezért az aminosavaknak először egy olyan közvetítő molekulához kell kapcsolódni, amely valamilyen módon kapcsolódni képes az mrns-el. A közvetítő molekula a trns amelynek feladata tehát kettős: Az adott aminosavval összekapcsolódik, s mivel szerkezete komplementer a megfelelő kodonnal, megfelelteti egymásnak az aminosavat és az aktuális kodont. A polipeptidlánc szintézise E-t igényel, ezért előzőleg az aminosavakat magasabb energiaállapotba kell hozni, azaz aktiválni kell. Az aktiválás ATP segítségével történik, majd a befektetett energia konzerválását a trns és az aminosav közötti nagyenergiájú kötés biztosítja. Aminosav aktiválása, a trns feltöltése 1. aminosav + ATP = aminosav-amp + PP 2. Az aktivált aminosav a megfelelő trns 3'-végéhez kötődik. aminosav-amp + trns = aminosav-trns + AMP Az aminosavak kapcsolódását a trns-ekhez specifikus enzimek ún. aminoaciltrns szintetázok katalizálják. Minden aminosavat külön enzim kapcsol trns molekulához, így legalább 20-féle szintetáz létezik. Lásd ábragyűjtemény. A trns szerkezete A trns a ribonukleinsavak között a legkisebb molekula, mindössze nukleotidegységből épül fel. A fehérjéket alkotó 20 aminosav mindegyikének legalább egy, némelyiknek azonban több trns felel meg. A trns-nek specifikus háromdimenziós szerkezete van, amely biztosítja a maximális bázispárosodást, a molekula maximális stabilitását. A molekula térbeli szerkezete lóherelevélre emlékeztet. A molekulának 4 specifikus része van. 1. Az aminosav megkötésére szolgáló, CCA szekvenciájú 3'-vég. 2. A mrns kodon felismerését biztosító, ún. antikodon rész. 3. Az aktiváló enzimmel való kapcsolódást biztosító enzimfelismerő hely. 4. Riboszóma kapcsolódási hely. A második hurok az ún. antikodon amely lehetővé teszi, hogy az adott trns-molekula, amely adott aminosavat szállít az mrns molekula megfelelő részéhez, a neki megfelelő kodonhoz kapcsolódjon. 12

13 Az mrns kodon és a trns antikodon komplementaritása a biztosítéka annak, hogy adott DNS-bázisszekvenciáról mindig azonos aminosav-szekvencia keletkezzék. A riboszómák A fehérje szintézis helyszínei, rrns-ekből és fehérjékből épülnek fel. Prokarióta sejtekben ezernyi, eukarióta sejtekben sok ezernyi található a citoplazmában. Működésük, hogy az mrns nukleotid sorrendjében foglalt információt fordítják le aminosavsorrendre. Szerkezetük egységes az egész élővilágban, egy nagyobb és egy kisebb alegységből állnak. A nagy alegység köti meg a trns-eket a transzláció során. A nagy alegység felületén három eltérő trns-kötőhely van: E (exit), P (peptidil-trns) és A (aminosav-trns). A két alegység a fehérjeszintézis kezdetén kapcsolódik össze, majd a transzláció végén ismét disszociálnak. A transzláció lépései prokariotákban A polipeptidlánc keletkezésének 3 fő szakasza van. 1. Iniciáció azaz láncindítás. 2. Elongáció azaz lánchosszabbítás. 3. Termináció azaz lánclezárás. Iniciáció A polipeptidlánc szintézise, csakúgy, mint a nukleinsavaké, egy irányba halad. A transzláció iránya, az mrns szintézisének irányával egyezően 5' 3'. Az mrns 5'-vége kevéssel kialakulása után már kapcsolódhat riboszómákhoz és megkezdődhet a polipeptidlánc szintézise. Az iniciáció lépései. Az mrns hozzákötődik a riboszóma kis alegységéhez. Az mrns megkötését a kis alegységben található rrns végzi. Az mrns 5 végének szekvenciája komplementer az rrns-ével. A nagy alegység összekapcsolódik a kicsivel. A start kodon AUG - a P ablakba kerül és hozzá kapcsolódik az első aminosavat szállító -formilmetionin- trns. A metionin később lehasad a polipeptidláncról. A folyamat energia igényét GTP hidrolízise fedezi. Elongáció. Lényege, hogy a kodon felismerése révén a megfelelő aminoacil-trns-ek egymást követően a riboszómához kapcsolódnak, az aminosavak között kialakul a peptidkötés, a kialakult peptidszakasz továbbítódik a riboszóma odébb mozdul - az újabb peptidkötés kialakítása érdekében. 13
, P (peptidil-trns) és A (aminosav-trns).")
14 A következő trns a riboszómán az A-helyhez kötődik. A riboszóma A-helyére kerülő újonnan érkezett trns aminosava összekapcsolódik a fmet-trns aminosav részével. A keletkezett dipeptidil-trns az A-helyen kötődik, míg az aminosavtól megfosztott trns egyelőre még a P-helyen marad. A P-helyen feleslegessé váló trns árkerül az E-helyre, majd leválik a riboszómáról. Egy-egy aminosav kapcsolódása után a keletkező trns az A-helyről a P-helyre kerül (transzlokáció), hogy ez által a következő aminoacil-trns kötődhessen az A-helyhez. A polipeptidlánc az N-terminális felöl nő a C-terminális felé. Termináció Ha a riboszóma A ablaka elérkezik a stop kodonhoz, a transzláció befejeződik. A terminációnak az az oka, hogy a három stop kodonhoz nem kapcsolódik trns, mivel a kodonoknak megfelelő komplementer antikodont tartalmazó trns nincs. A stop-jel felismerését követően a P-helyen levő utolsó trns és az aminosav közötti kötés felszakad, a polipeptidlánc leválik a riboszómáról. A három stop-szignál felismerésében riboszómális fehérjék közreműködnek. Végül riboszóma alegységek disszociálnak. Baktériumokban a fehérje szintézis sebessége rendkívül gyors, másodpercenként akár 30 aminosav is beépülhet. Az intenzív transzlációnak több oka van: az mrns DNS-ről történő transzkripciója alatt, még a nem teljesen kész mrns-en már megindul a transzláció. Ha a riboszóma bizonyos számú kodont már leolvasott, az mrns 5'-végéhez újabb riboszóma kapcsolódhat. Így gyakran egy mrns-en egyszerre jó néhány riboszóma dolgozik, és ún. poliriboszóma alakul ki, ami lehetővé teszi, hogy egy időben egy mrns-ről egyszerre több polipeptidlánc íródjék át. Lac-operon Génműködés szabályzása a prokariótákban A baktériumok tápanyagok levesében úszva élnek. Sokféle különböző cukor fordulhat elő a környezetükben. Ezek felvételéhez és lebontásához szükséges enzimek egyidejű termelése sok energiába kerül. A baktériumok érzékelni képesek a rendelkezésre álló cukor típusát és csak akkor termelik a felvételhez és lebontáshoz szükséges enzimeket, ha szükség van azokra (enzimindukció). A génműködés szabályozását leíró modell az operon. Az operon általi génműködés szabályozás a sejtek gyors és hatékony alkalmazkodását teszi lehetővé az adott környezeti feltételekhez. A laktóz-operon röviden lacoperon - volt az elsőként leírt génregulációs mechanizmus, aminek felfedezésért két francia kutató, a párizsi Pasteur Intézetben dolgozó Francois Jacob és Jacques Monod Nobel Díjat kaptak 1965-ben. 14
, hogy ez által a következő aminoacil-trns kötődhessen az A-helyhez.")
15 Az operon a génműködés-szabályozás egysége. Megkülönböztetünk a DNS-molekulában strukturális géneket, amelyek fehérjék aminosavsorrendjét kódolják, átíródnak és továbbá régió szakaszokat, amelyekről nincs átíródás nem kódolnak fehérjét -, ezek a struktúrgének átíródását lehetővé tevő egységek. A lac operon a következő elemekből áll: három strukturális (szerkezeti) gén. o Az E. coli a laktózt a b-galaktozidáz enzimmel bontja le, melyet lac Z gén kódol. o A laktóz felvételéhez szükséges a permeáz enzim is, melyet a lac Y strukturgén kódol. o Egy harmadik, a laktóz hasznosításához közvetlenül nem fontos gén a transzacetiláz (lac A,) is ugyanebbe a csoportba tartozik (piroszőlősav bontásához kell). A transz-acetiláz pontos funkciója nem ismert, eltávolítása nem gátolja számottevően az operon működését. Promóter (lac P), az RNS polimeráz kapcsolódási helye, operátor régio (Lac O), egy olyan DNS szakasz, ahol egy gátló (represszor, regulátor) fehérje képes megkötődni, megakadályozva a fenti strukturgének átírását, abban az esetben, ha nincs laktóz a tápanyagok között. Nem tartozik a lac operonhoz, - de alapvetően szükséges annak működéséhez,- az ún. lac I gén. A laci gén kódolja a represszor fehérjét, amely az operátorhoz kapcsolódva blokkolja a strukturális gének átíródását. A lac Z,Y,A gének átírása promóteren kezdődik, ahol az RNS polimeráz megkötődik a DNS-en. Tehát röviden mégegyszer a lac operon áll: promoterből (indító régió) (lac P), operátorból (operátor régió) (lac O), és lac Z, A és Y strukturgénekből áll. Az operon működésének szabályozásában részt vesz a represszor (regulátor), a lac I gén terméke a gátló fehérje és fontos a sejt környezetének laktóz tartalma. Laktóz hiányában a represszorfehérje (lac I terméke) az operátorhoz kapcsolódik és megakadályozza a strukturgének átírását. A környezet laktóz tartalmát a represszor fehérje érzékeli. A laktóz jelenlétében a represszor megköti a laktózt, konformációja megváltozik, levállik az operátorról és nem képes tovább akadályozni az RNS polimerázt a strukturgének átírásában. Eredményül átíródnak a strukturgének, fokozódik a laktóz felvétele, lebontása. 15


16 A triptofán operon (trp-operon) A lac-operon egy lebontó anyagcsereutat szabályoz, a laktóz kötődése a represszorhoz serkenti a strukturgének átírását. A trp-operon egy felépítő utat, a triptofán aminosav szintézisét befolyásolja. A Trp kötődése a represszorhoz gátolja a génátírást. A trp-operon egy promoter és egy operátor régióból egy ún. leader-szekvenciából és öt strukturgénből áll. Tehát, ha nincs trp, akkor történhet transzkripció, így a trp-szintézis megindul; ha van trp a rendszerben, akkor trp-represszor fehérje trp-nal kapcsolódva aktiválódik, megtörténik az operátor régióhoz való kötődés, és a transzkripció gátolt lesz. A trp-operonban nemcsak az átírás kezdetét (iniciáció) lehet szabályozni, hanem a végét is (termináció). Ez utóbbi folyamatot attenuációnak nevezzük, és a leader-szekvencia révén megy végbe. (bonyolult) Stresszfehérjék Forrás: Csermely Péter A stresszfehérjék legfontosabb szerepe, a hibás, nem megfelelő harmadlagos szerkezettel rendelkező fehérjék helyretekerése. A stresszfehérjék sejtjeink igen nagy mennyiségben jelen lévő, létfontosságú alkotóelemei. A stresszfehérjék segítsége nélkülözhetetlen abban, hogy a hibás szerkezetű fehérjék megtalálhassák a rájuk jellemző, helyes szerkezetet. A stresszfehérjék szerepe kettős: a hosszabb polipeptidláncok megfelelő harmadlagos szerkezetének kialakítása, a káros stresszhatásoknak kitett, károsodott fehérjék szerkezetének helyreállítása. 1. A kisméretű fehérjék betekeredése kedvező körülmények között másodpercek alatt végbemegy. Sajnos az ennél nagyobb fehérjék igen gyakran tekeredési csapdák áldozatai lesznek. Ebben az esetben a félkész fehérje egy átmeneti állapotban stabilizálódik, a hidrofób aminosavak a fehérje felszínén maradnak. Sajnos a hidrofób felszínek az ilyen félkész fehérjéket egymással való összetapadásra, aggregációra teszik hajlamossá. A fehérje-aggregátumok felszaporodása a legtöbb neurodegeneratív betegség, pl. az Alzheimer-kór, a Parkinson-kór, a prionbetegségek egyik fontos oka. A stresszfehérjéket a többi fehérjét terelgető, szerkezetük kialakításában segítő hatásuk miatt dajkafehérjéknek is nevezik. 16

17 2. Ha a sejtet környezeti stressz éri, a sejtben lévő fehérjék károsodnak, kitekerednek. E fehérjéknek az újratekeredéshez éppen úgy a dajkafehérjék segítségre van szükségük, mint születésük pillanatában. Innen a stresszfehérje elnevezés. Környezeti stressz szinte bármilyen hirtelen változás lehet: lényegesen melegebb (hősokk fehérjék), vagy hidegebb; a környezet lényegesen savasabb, vagy lúgosabb; túl kevés az oxigén, vagy túl sok; vízhiány. Néha azonban a stresszfehérje is tehetetlennek bizonyul. Ha a selejtes fehérje menthetetlen, akkor helyretekerése helyett a lebontása következik be. Ilyenkor a stresszfehérjék pusztulásra ítélt társaikat kitekerik egészen addig, ameddig be nem férnek a lebontó enzimek szűk bemeneti csatornájába. Human Genom Project Az emberi genom a petesejt vagy hímivarsejt teljes genetikai tartalma, amely hozzávetőleg 3 milliárd DNS bázispárból áll. A HGP elsődleges célja volt kideríteni annak a kb. 3 milliárd bázispárnak a pontos sorrendjét, amely az emberi genomot alkotja. A Humán Genom Projekt tizenhárom éven keresztül zajlott, június 26-án Clinton és Blair közös sajtóértekezleten jelentették be a program első ütemének befejezését. A HGP 9 ember DNS-ét szekvenálta le. Meglepetést okozott, hogy csak kb gén van a humán genomban, ami a teljes DNS állomány 1,5 %-át teszi ki (a korábbi becslések mindegyike fölé helyezte a számukat, de van, aki re becsülte). A gének száma mindössze kétszerese a Drosophilában meghatározottaknak. A komplexitás titka nem a gének számában, hanem a gének által meghatározott fehérjék összerakási módjában van rejtve, amiben fontos szerepet játszik az alternatív splicing mechanizmusa, amely következtében egy gén többféle fehérjét is meghatározhat. Tehát a tudománytörténetben jelentős szerepet játszó klasszikus tézis: egy gén egy enzim messzemenően nem igaz. Az adatok alapján megállapítható hogy az exonok átlagos hossza: bp, az intronoké: bp, 1 gén pedig átlagosan 7-9 exonból áll. A fehérjekódot ténylegesen hordozó exonok a teljes szekvenciának a 1,5 %-át adják. Az összes hasznos információt kb. 10 %-a hordozza a DNS állománynak, amely tartalmazza o tehát az exonokat, o a t-és rrns génjeit, o továbbá a gén kifejeződését szabályozó DNS szakaszokat (promoter régió). A szekvenciák alapján 2 embernek 0,1 %-os eltérést mutatott a szekvenciája, tehát az egyezés 99,9 %-os volt. Azaz bp közül átlagosan 1000 különbözik. A HGP következő szakaszában meg kell határozni, hogy az adott tulajdonságot hány gén, azok milyen viszonylatban határozzák meg, az adott gének pontosan melyik kromoszómán helyezkednek el. Ma már kb különböző fajú élőlény (köztük 156 eukarióta) teljes genomszekvenciája ismert, de a futó, azaz még nem teljesen befejezett genomprogramok száma megközelíti a tízezret. 17

18 További eredmények: Az emberi genom mintegy száz különböző, mikro-rns-eket kódoló, evolúciósan konzervált géncsaládot tartalmaz. A mikro-rns-ek olyan RNS molekulák, amelyek fehérjét nem kódolnak, viszont befolyásolják adott mrns-ek transzlációját azáltal, hogy az mrns molekula 3 végéhez kötődve többnyire gátolják a fehérje szintézist. Vagyis a mirns-ek a génkifejeződés poszttranszkripciós szintű negatív szabályozóinak tekinthetők. Az emberi genom mintegy felét adják a különböző transzpozonok, azaz mozgékony, helyváltoztatásra képes genetikai elemek. A genom 45 százalékát alkothatják ezek a szakaszok. Többségük azonban nem aktív, és változatlanul adódik tovább egyik nemzedékről a másikra. Az átrendeződés úgy történhet, hogy o a DNS-szakasz kihasad a genomból és egy másik helyen integrálódik. Ekkor a mobilis elem az eredeti helyéről természetesen eltűnik, vagy úgy hogy o a DNS-régió megkettőződik, az egyik kópia marad a helyén, a másik máshol integrálódik. o A transzpozíció másik fő típusa RNS közvetítésével történik a donor DNS-régió RNS-be íródik át, majd azon mint templáton reverz transzkriptáz segítségével kettős láncú DNS szintetizálódik és az beépül a cél-dns-be. o Így történik az immunglobulin-gének átrendeződése a B limfociták differenciálódása során. Az epigenetika jelentősége. Ez a tudományág azzal foglalkozik, hogy a különféle környezeti tényezők hogyan hatnak a genom működésére, a gének kifejeződésére. Az epigenetikus hatások (pl. anyai hatás, táplálék, mozgás, gyógyszerek, mérgek, lelki hatások, fény, stb.) anélkül befolyásolják a gének működését, hogy megváltoztatnák a DNS bázissorendjét. Tehát e tudományterület a genom olyan megváltozásaival foglalkozik, amelyek nem jelentik a DNS nukleotidsorrendjének megváltozását, csak a DNS másodlagos kémiai módosítását (pl. egyes nukleotidok metilációját), vagy a DNS-el szorosan kölcsönható fehérjék módosulásait. A soksejtű élőlényekben az egyedfejlődés különböző szakaszain, ilyen epigenetikus módosulások irányítják a génkifejezés megváltozásait. Kivételes esetekben öröklődhetnek is az epigenetikus módosulások. Röviden összefoglalva A DNS kutatásának fontosabb korai állomásait. Friedrich Miescher ( ) 1869-ben felfedezte a DNS-t fehérvérsejtek sejtmagjában. Fred Griffith 1928-ban felfedezte a baktérium transzformációt. Oswald Avery ( ) azonosította a transzformációért felelős, vagyis a genetikai információt hordozó anyag kémiai természetét, azonosította a DNS-t. Erwin Chargaff ( ) 1950-ben megfogalmazza a bázisok arányára vonatkozó szabályt ben Alfred Hershey ( ) és Martha Chase ( ) megdönthetetlen kísérlettel bizonyította Avery eredményét. Hershey1969-ben fiziológiai és orvostudományi Nobel-díjat kapott. James D. Watson (1928-) és Francis H. C. Crick ( ) 1953-ban leírták a DNS kettős hélix szerkezetét. Watson, Crick és Wilkins 1962-ben fiziológiai és orvostudományi Nobel-díjat kapott ban Menselson és Stahl leírták a szemi-konzervatív DNS replikációt. Marshall Nirenberg ( ) és Har Gobind Khorana ( ) 1968-ban a genetikai kód megfejtéséért megkapta a Nobel-díjat. Ábragyűjtemény Fred Griffith Oswald Avery Alfred Hershey és Martha Chase 18

19 Aminosavak aktiválása Poszttranszkripciós átalakulások Riboszóma 19
A replikáció mechanizmusa
 Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
A tananyag felépítése: A BIOLÓGIA ALAPJAI. I. Prokarióták és eukarióták. Az eukarióta sejt. Pécs Miklós: A biológia alapjai
 A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A géntechnológia genetikai alapjai (I./3.)
") Az I./2. rész (Gének és funkciójuk) rövid összefoglalója A gének a DNS információt hordozó szakaszai, melyekben a 4 betű (ATCG) néhány ezerszer, vagy százezerszer ismétlődik. A gének önálló programcsomagként
Az I./2. rész (Gének és funkciójuk) rövid összefoglalója A gének a DNS információt hordozó szakaszai, melyekben a 4 betű (ATCG) néhány ezerszer, vagy százezerszer ismétlődik. A gének önálló programcsomagként
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
3. Sejtalkotó molekulák III.
 3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
Transzláció. Leolvasás - fehérjeszintézis
 Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
DER (Felületén riboszómák találhatók) Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.
 Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.") Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
a III. kategória (11-12. évfolyam) feladatlapja
 feladatlapja") 2009/2010. tanév I. forduló a III. kategória (11-12. évfolyam) feladatlapja Versenyző neve:... évfolyama: Iskolája : Település : Felkészítő szaktanár neve:.. Megoldási útmutató A verseny feladatait nyolc
2009/2010. tanév I. forduló a III. kategória (11-12. évfolyam) feladatlapja Versenyző neve:... évfolyama: Iskolája : Település : Felkészítő szaktanár neve:.. Megoldási útmutató A verseny feladatait nyolc
 Az élő szervezetek felépítése I. Biogén elemek biomolekulák alkotóelemei a természetben előforduló elemek közül 22 fordul elő az élővilágban O; N; C; H; P; és S; - élő anyag 99%-a Biogén elemek sajátosságai:
Az élő szervezetek felépítése I. Biogén elemek biomolekulák alkotóelemei a természetben előforduló elemek közül 22 fordul elő az élővilágban O; N; C; H; P; és S; - élő anyag 99%-a Biogén elemek sajátosságai:
Nukleinsavak. Szerkezet, szintézis, funkció
 Nukleinsavak Szerkezet, szintézis, funkció Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
Nukleinsavak Szerkezet, szintézis, funkció Nukleinsavak, nukleotidok, nukleozidok 1869-ben Miescher a sejtmagból egy savas természetű, lúgban oldódó foszfortartalmú anyagot izolált, amit később, eredetére
Génszerkezet és génfunkció
 Általános és Orvosi Genetika jegyzet 4. fejezetének bővítése a bakteriális genetikával 4. fejezet Génszerkezet és génfunkció 1/ Bakteriális genetika Nem szükséges külön hangsúlyoznunk a baktériumok és
Általános és Orvosi Genetika jegyzet 4. fejezetének bővítése a bakteriális genetikával 4. fejezet Génszerkezet és génfunkció 1/ Bakteriális genetika Nem szükséges külön hangsúlyoznunk a baktériumok és
CzB 2010. Élettan: a sejt
 CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
Transzgénikus növények előállítása
 Transzgénikus növények előállítása Növényi biotechnológia Területei: A növények szaporításának új módszerei Növényi sejt és szövettenyészetek alkalmazása Mikroszaporítás Vírusmentes szaporítóanyag előállítása
Transzgénikus növények előállítása Növényi biotechnológia Területei: A növények szaporításának új módszerei Növényi sejt és szövettenyészetek alkalmazása Mikroszaporítás Vírusmentes szaporítóanyag előállítása
BIOLÓGIA VERSENY 10. osztály 2016. február 20.
 BIOLÓGIA VERSENY 10. osztály 2016. február 20. Kód Elérhető pontszám: 100 Elért pontszám: I. Definíció (2x1 = 2 pont): a) Mikroszkopikus méretű szilárd részecskék aktív bekebelezése b) Molekula, a sejt
BIOLÓGIA VERSENY 10. osztály 2016. február 20. Kód Elérhető pontszám: 100 Elért pontszám: I. Definíció (2x1 = 2 pont): a) Mikroszkopikus méretű szilárd részecskék aktív bekebelezése b) Molekula, a sejt
NUKLEINSAVAK. Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag
 NUKLEINSAVAK Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag RNS = Ribonukleinsav DNS = Dezoxi-ribonukleinsav A nukleinsavak
NUKLEINSAVAK Nukleinsav: az élő szervezetek sejtmagvában és a citoplazmában található, az átöröklésben szerepet játszó, nagy molekulájú anyag RNS = Ribonukleinsav DNS = Dezoxi-ribonukleinsav A nukleinsavak
Kevéssé fejlett, sejthártya betüremkedésekből. Citoplazmában, cirkuláris DNS, hisztonok nincsenek
 1 A sejtek felépítése Szerkesztette: Vizkievicz András A sejt az élővilág legkisebb, önálló életre képes, minden életjelenséget mutató szerveződési egysége. Minden élőlény sejtes szerveződésű, amelyek
1 A sejtek felépítése Szerkesztette: Vizkievicz András A sejt az élővilág legkisebb, önálló életre képes, minden életjelenséget mutató szerveződési egysége. Minden élőlény sejtes szerveződésű, amelyek
AZ EMBERI MIKROBIOM: AZ EGYÉN, MINT SAJÁTOS ÉLETKÖZÖSSÉG Duda Ernő
 AZ EMBERI MIKROBIOM: AZ EGYÉN, MINT SAJÁTOS ÉLETKÖZÖSSÉG Duda Ernő Az NIH, az Egyesült Államok Nemzeti Egészségügyi Hivatala (az orvosi- és biológiai kutatásokat koordináló egyik intézmény) 2007 végén
AZ EMBERI MIKROBIOM: AZ EGYÉN, MINT SAJÁTOS ÉLETKÖZÖSSÉG Duda Ernő Az NIH, az Egyesült Államok Nemzeti Egészségügyi Hivatala (az orvosi- és biológiai kutatásokat koordináló egyik intézmény) 2007 végén
Az endomembránrendszer részei.
 Az endomembránrendszer Szerkesztette: Vizkievicz András Az eukarióta sejtek prokarióta sejtektől megkülönböztető egyik alapvető sajátságuk a belső membránrendszerük. A belső membránrendszer szerkezete
Az endomembránrendszer Szerkesztette: Vizkievicz András Az eukarióta sejtek prokarióta sejtektől megkülönböztető egyik alapvető sajátságuk a belső membránrendszerük. A belső membránrendszer szerkezete
3. Kombinált, amelynek van helikális és kubikális szakasza, pl. a bakteriofágok és egyes rákkeltő RNS vírusok.
 Vírusok Szerkesztette: Vizkievicz András A XIX. sz. végén Dmitrij Ivanovszkij orosz biológus a dohány mozaikosodásának kórokozóját próbálta kimutatni. A mozaikosodás a levél foltokban jelentkező sárgulása.
Vírusok Szerkesztette: Vizkievicz András A XIX. sz. végén Dmitrij Ivanovszkij orosz biológus a dohány mozaikosodásának kórokozóját próbálta kimutatni. A mozaikosodás a levél foltokban jelentkező sárgulása.
Vírusok Szerk.: Vizkievicz András
 Vírusok Szerk.: Vizkievicz András A vírusok az élő- és az élettelen világ határán állnak. Önmagukban semmilyen életjelenséget nem mutatnak, nincs anyagcseréjük, önálló szaporodásra képtelenek. Paraziták.
Vírusok Szerk.: Vizkievicz András A vírusok az élő- és az élettelen világ határán állnak. Önmagukban semmilyen életjelenséget nem mutatnak, nincs anyagcseréjük, önálló szaporodásra képtelenek. Paraziták.
TRANSZPORTFOLYAMATOK 1b. Fehérjék. 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS
 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
A baktériumok genetikája
 6. előadás A baktériumok genetikája A baktériumoknak fontos szerep jut a genetikai kutatásokban Előny: Haploid genom Rövid generációs idő Olcsón és egyszerűen nagy populációhoz juthatunk A prokarióták
6. előadás A baktériumok genetikája A baktériumoknak fontos szerep jut a genetikai kutatásokban Előny: Haploid genom Rövid generációs idő Olcsón és egyszerűen nagy populációhoz juthatunk A prokarióták
TestLine - Biogén elemek, molekulák Minta feladatsor
 TestLine - iogén elemek, molekulák iogén elemek, szervetlen és szerves molekulák az élő szervezetben. gészítsd ki a mondatot! aminocsoportja kondenzáció víz ún. peptidkötés 1. 1:48 Normál fehérjék biológiai
TestLine - iogén elemek, molekulák iogén elemek, szervetlen és szerves molekulák az élő szervezetben. gészítsd ki a mondatot! aminocsoportja kondenzáció víz ún. peptidkötés 1. 1:48 Normál fehérjék biológiai
,:/ " \ OH OH OH - 6 - / \ O / H / H HO-CH, O, CH CH - OH ,\ / "CH - ~(H CH,-OH \OH. ,-\ ce/luló z 5zer.~ezere
 - 6 - o / \ \ o / \ / \ () /,-\ ce/luló z 5zer.~ezere " C=,1 -- J - 1 - - ---,:/ " - -,,\ / " - ~( / \ J,-\ ribóz: a) r.yílt 12"('.1, b) gyürus íormája ~.. ~ en;én'. fu5 héli'(ef1e~: egy menete - 7-5.
- 6 - o / \ \ o / \ / \ () /,-\ ce/luló z 5zer.~ezere " C=,1 -- J - 1 - - ---,:/ " - -,,\ / " - ~( / \ J,-\ ribóz: a) r.yílt 12"('.1, b) gyürus íormája ~.. ~ en;én'. fu5 héli'(ef1e~: egy menete - 7-5.
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Gergely Pál 2009
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
A szénhidrátok lebomlása
 A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
Sportélettan zsírok. Futónaptár.hu
 Sportélettan zsírok Futónaptár.hu A hétköznapi ember csak hallgatja azokat a sok okos étkezési tanácsokat, amiket az egészségének megóvása érdekében a kutatók kiderítettek az elmúlt 20 évben. Emlékezhetünk
Sportélettan zsírok Futónaptár.hu A hétköznapi ember csak hallgatja azokat a sok okos étkezési tanácsokat, amiket az egészségének megóvása érdekében a kutatók kiderítettek az elmúlt 20 évben. Emlékezhetünk
A transzgénikus (GM) fajták fogyasztásának élelmiszer-biztonsági kockázatai
 fajták fogyasztásának élelmiszer-biztonsági kockázatai") BIOTECHNOLÓGIA O I ROVATVEZETŐ: Dr. Heszky László akadémikus A GM-növényekkel szembeni társadalmi elutasítás legfontosabb indokait az élelmiszer-biztonsági kockázatok jelentik. A géntechnológia forradalmian
BIOTECHNOLÓGIA O I ROVATVEZETŐ: Dr. Heszky László akadémikus A GM-növényekkel szembeni társadalmi elutasítás legfontosabb indokait az élelmiszer-biztonsági kockázatok jelentik. A géntechnológia forradalmian
Az örökítőanyag. Az élőlények örökítőanyaga minden esetben nukleinsav (DNS,RNS) (1)Griffith, (2)Avery, MacLeod and McCarty (3)Hershey and Chase
 (1)Griffith, (2)Avery, MacLeod and McCarty (3)Hershey and Chase") SZTE, Orv. Biol. Int., Mol- és Sejtbiol. Gyak., VIII. Az örökítőanyag Az élőlények örökítőanyaga minden esetben nukleinsav (DNS,RNS) (1)Griffith, (2)Avery, MacLeod and McCarty (3)Hershey and Chase Ez az
SZTE, Orv. Biol. Int., Mol- és Sejtbiol. Gyak., VIII. Az örökítőanyag Az élőlények örökítőanyaga minden esetben nukleinsav (DNS,RNS) (1)Griffith, (2)Avery, MacLeod and McCarty (3)Hershey and Chase Ez az
Vírusok I: általános
 1 Mi egy vírus? VÍRUSOK-I Vírusok I: általános I. Bevezetés A vírusok sejtparaziták, ami azt jelenti, hogy (1) a sejten kívül nem képesek élettevékenységet folytatni. (2) Továbbá, a vírusok a fertőzött
1 Mi egy vírus? VÍRUSOK-I Vírusok I: általános I. Bevezetés A vírusok sejtparaziták, ami azt jelenti, hogy (1) a sejten kívül nem képesek élettevékenységet folytatni. (2) Továbbá, a vírusok a fertőzött
Szerkesztette: Vizkievicz András
 Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
A kémiai energia átalakítása a sejtekben
 A kémiai energia átalakítása a sejtekben A sejtek olyan mikroszkópikus képződmények amelyek működése egy vegyi gyárhoz hasonlítható. Tehát a sejtek mikroszkópikus vegyi gyárak. Mi mindenben hasonlítanak
A kémiai energia átalakítása a sejtekben A sejtek olyan mikroszkópikus képződmények amelyek működése egy vegyi gyárhoz hasonlítható. Tehát a sejtek mikroszkópikus vegyi gyárak. Mi mindenben hasonlítanak
A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN. Somogyi János -- Vér Ágota Első rész
 A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN Somogyi János -- Vér Ágota Első rész Már több mint 200 éve ismert, hogy szöveteink és sejtjeink zöme oxigént fogyaszt. Hosszú ideig azt hitték azonban, hogy
A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN Somogyi János -- Vér Ágota Első rész Már több mint 200 éve ismert, hogy szöveteink és sejtjeink zöme oxigént fogyaszt. Hosszú ideig azt hitték azonban, hogy
DNS, RNS, Fehérjék. makromolekulák biofizikája. Biológiai makromolekulák. A makromolekulák TÖMEG szerinti mennyisége a sejtben NAGY
 makromolekulák biofizikája DNS, RNS, Fehérjék Kellermayer Miklós Tér Méret, alak, lokális és globális szerkezet Idő Fluktuációk, szerkezetváltozások, gombolyodás Kölcsönhatások Belső és külső kölcsöhatások,
makromolekulák biofizikája DNS, RNS, Fehérjék Kellermayer Miklós Tér Méret, alak, lokális és globális szerkezet Idő Fluktuációk, szerkezetváltozások, gombolyodás Kölcsönhatások Belső és külső kölcsöhatások,
AMINOSAVAK, FEHÉRJÉK
 AMINOSAVAK, FEHÉRJÉK Az aminosavak olyan szerves vegyületek, amelyek molekulájában aminocsoport (-NH2) és karboxilcsoport (-COOH) egyaránt előfordul. Felosztás A fehérjéket feloszthatjuk aszerint, hogy
AMINOSAVAK, FEHÉRJÉK Az aminosavak olyan szerves vegyületek, amelyek molekulájában aminocsoport (-NH2) és karboxilcsoport (-COOH) egyaránt előfordul. Felosztás A fehérjéket feloszthatjuk aszerint, hogy
A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek.
 Szénhidrátok Szerkesztette: Vizkievicz András A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek. A szénhidrátok
Szénhidrátok Szerkesztette: Vizkievicz András A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek. A szénhidrátok
1. ábra: A hasnyálmirigy Langerhans-szigete
 génmanipulált mikroorganizmusokkal Az elsődleges és másodlagos anyagcseretermékek előállítása után a rekombináns fehérjék gyártásáról lesz szó. Ezek olyan fehérjék, melyeket a sejt eredeti genomja nem
génmanipulált mikroorganizmusokkal Az elsődleges és másodlagos anyagcseretermékek előállítása után a rekombináns fehérjék gyártásáról lesz szó. Ezek olyan fehérjék, melyeket a sejt eredeti genomja nem
Vizsgakövetelmények Tudjon elemezni kísérleteket a DNS örökítő szerepének bizonyítására (Griffith és Avery, Hershey és Chase kísérlete).
.") 1 Vizsgakövetelmények Tudjon elemezni kísérleteket a DNS örökítő szerepének bizonyítására (Griffith és Avery, Hershey és Chase kísérlete). Ismertessen néhány példát a genetikai technológia alkalmazására
1 Vizsgakövetelmények Tudjon elemezni kísérleteket a DNS örökítő szerepének bizonyítására (Griffith és Avery, Hershey és Chase kísérlete). Ismertessen néhány példát a genetikai technológia alkalmazására
A sejtek élete. 5. Robotoló törpék és óriások Az aminosavak és fehérjék R C NH 2. C COOH 5.1. A fehérjeépítőaminosavak általános
 A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
A szénhidrátok lebomlása
 A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
A sejtes szervezıdés elemei (sejtalkotók / sejtorganellumok)
") A sejtes szervezıdés elemei (sejtalkotók / sejtorganellumok) 1 Sejtorganellumok vizsgálata: fénymikroszkóp elektronmikroszkóp pl. scanning EMS A szupramolekuláris struktúrák további szervezıdése sejtorganellumok
A sejtes szervezıdés elemei (sejtalkotók / sejtorganellumok) 1 Sejtorganellumok vizsgálata: fénymikroszkóp elektronmikroszkóp pl. scanning EMS A szupramolekuláris struktúrák további szervezıdése sejtorganellumok
1. Bevezetés. Mi az élet, evolúció, információ és energiaáramlás, a szerveződés szintjei
 1. Bevezetés Mi az élet, evolúció, információ és energiaáramlás, a szerveződés szintjei 1.1 Mi az élet? Definíció Alkalmas legyen különbségtételre élő/élettelen közt Ne legyen túl korlátozó (más területen
1. Bevezetés Mi az élet, evolúció, információ és energiaáramlás, a szerveződés szintjei 1.1 Mi az élet? Definíció Alkalmas legyen különbségtételre élő/élettelen közt Ne legyen túl korlátozó (más területen
Leukémia (fehérvérûség)
") Leukémia (fehérvérûség) Leukémia - fehérvérûség A leukémia a rosszindulatú rákos megbetegedések azon formája, amely a vérképzõ sejtekbõl indul ki. A leukémia a csontvelõben lévõ éretlen és érettebb vérképzõ
Leukémia (fehérvérûség) Leukémia - fehérvérûség A leukémia a rosszindulatú rákos megbetegedések azon formája, amely a vérképzõ sejtekbõl indul ki. A leukémia a csontvelõben lévõ éretlen és érettebb vérképzõ
Géntechnológia és fehérjemérnökség
 Géntechnológia és fehérjemérnökség Szerkesztette: Nyitray László Alexa Anita (12. és 13. fejezet) Fodor Krisztián (3. és 9. fejezet) Garai Ágnes (4. és 5. fejezet) Glatz Gábor (6. és 7. fejezet) Radnai
Géntechnológia és fehérjemérnökség Szerkesztette: Nyitray László Alexa Anita (12. és 13. fejezet) Fodor Krisztián (3. és 9. fejezet) Garai Ágnes (4. és 5. fejezet) Glatz Gábor (6. és 7. fejezet) Radnai
A felvétel és a leadás közötti átalakító folyamatok összességét intermedier - köztes anyagcserének nevezzük.
 1 Az anyagcsere Szerk.: Vizkievicz András Általános bevezető Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az élő sejtek nyílt anyagi rendszerek, azaz környezetükkel állandó
1 Az anyagcsere Szerk.: Vizkievicz András Általános bevezető Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az élő sejtek nyílt anyagi rendszerek, azaz környezetükkel állandó
Fejezet a Gulyás Méhészet által összeállított Méhészeti tudástár mézfogyasztóknak (2015) ismeretanyagból. A méz. összetétele és élettani hatása
 ismeretanyagból. A méz. összetétele és élettani hatása") A méz összetétele és élettani hatása A méz a növények nektárjából a méhek által előállított termék. A nektár a növények kiválasztási folyamatai során keletkezik, híg cukortartalmú oldat, amely a méheket
A méz összetétele és élettani hatása A méz a növények nektárjából a méhek által előállított termék. A nektár a növények kiválasztási folyamatai során keletkezik, híg cukortartalmú oldat, amely a méheket
Rekombináns Géntechnológia
 Rekombináns Géntechnológia Tartalom: 1 1. Biotechnológia, géntechnológia, társadalom 2. Genetikai rekombináció 3. Génbevitel tenyésztett sejtekbe 4. Genetikailag módosított szervezetek (GMO-k) 4a. Transzgénikus
Rekombináns Géntechnológia Tartalom: 1 1. Biotechnológia, géntechnológia, társadalom 2. Genetikai rekombináció 3. Génbevitel tenyésztett sejtekbe 4. Genetikailag módosított szervezetek (GMO-k) 4a. Transzgénikus
A prokarióták. A biológia tudománya az élők világát két alapvető birodalomra osztja: a prokariótákra és az eukariótákra.
 A prokarióták A biológia tudománya az élők világát két alapvető birodalomra osztja: a prokariótákra és az eukariótákra. Szerk.: Vizkievicz András A prokarióták legszembetűnőbb közös sajátsága, hogy sejtjükben
A prokarióták A biológia tudománya az élők világát két alapvető birodalomra osztja: a prokariótákra és az eukariótákra. Szerk.: Vizkievicz András A prokarióták legszembetűnőbb közös sajátsága, hogy sejtjükben
A proteomika új tudománya és alkalmazása a rákdiagnosztikában
 BIOTECHNOLÓGIAI FEJLESZTÉSI POLITIKA, KUTATÁSI IRÁNYOK A proteomika új tudománya és alkalmazása a rákdiagnosztikában Tárgyszavak: proteom; proteomika; rák; diagnosztika; molekuláris gyógyászat; biomarker;
BIOTECHNOLÓGIAI FEJLESZTÉSI POLITIKA, KUTATÁSI IRÁNYOK A proteomika új tudománya és alkalmazása a rákdiagnosztikában Tárgyszavak: proteom; proteomika; rák; diagnosztika; molekuláris gyógyászat; biomarker;
Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus. Szülő- és betegtájékoztató
 Bevezetés Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus Szülő- és betegtájékoztató Minden szülő azt várja, hogy gyermeke egészséges lesz. Így azután mélyen megrázza őket, ha megtudják, hogy
Bevezetés Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus Szülő- és betegtájékoztató Minden szülő azt várja, hogy gyermeke egészséges lesz. Így azután mélyen megrázza őket, ha megtudják, hogy
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Bay Péter
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
12/4/2014. Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció. 1952 Hershey & Chase 1953!!!
 Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció 1859 1865 1869 1952 Hershey & Chase 1953!!! 1879 1903 1951 1950 1944 1928 1911 1 1. DNS szerkezete Mi az örökítő anyag? Friedrich Miescher
Genetika 7-8 ea. DNS szerkezete, replikáció és a rekombináció 1859 1865 1869 1952 Hershey & Chase 1953!!! 1879 1903 1951 1950 1944 1928 1911 1 1. DNS szerkezete Mi az örökítő anyag? Friedrich Miescher
Molekuláris biológus M.Sc. Prokarióták élettana
 Molekuláris biológus M.Sc. Prokarióták élettana Bakteriális DNS replikáció. A génexpresszió szabályozása prokariótákban. Plazmidok, baktériumok transzformálása. A prokarióta genom nukleoid egyetlen cirkuláris
Molekuláris biológus M.Sc. Prokarióták élettana Bakteriális DNS replikáció. A génexpresszió szabályozása prokariótákban. Plazmidok, baktériumok transzformálása. A prokarióta genom nukleoid egyetlen cirkuláris
Géntechnológia és fehérjemérnökség
 Géntechnológia és fehérjemérnökség elektronikus-jegyzet szerzők: Az ELTE Biokémiai Tanszék Munkaközössége Alexa Anita (12. és 13. fejezet), Fodor Krisztián (3. és 9. fejezet), Garai Ágnes (4. és 5. fejezet),
Géntechnológia és fehérjemérnökség elektronikus-jegyzet szerzők: Az ELTE Biokémiai Tanszék Munkaközössége Alexa Anita (12. és 13. fejezet), Fodor Krisztián (3. és 9. fejezet), Garai Ágnes (4. és 5. fejezet),
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
7. A SEJT A SEJT 1. ÁLTALÁNOS TUDNIVALÓK
 A SEJT 1. ÁLTALÁNOS TUDNIVALÓK DIA 1 DIA 2 DIA 3 DIA 4 A sejtbiológia a biológiának az a tudományterülete, amely a sejt szerkezeti felépítésével, a különféle sejtfolyamatokkal (sejtlégzés, anyagtranszport,
A SEJT 1. ÁLTALÁNOS TUDNIVALÓK DIA 1 DIA 2 DIA 3 DIA 4 A sejtbiológia a biológiának az a tudományterülete, amely a sejt szerkezeti felépítésével, a különféle sejtfolyamatokkal (sejtlégzés, anyagtranszport,
Előadások témája: Elsősorban a DNS, a gének és genomok molekuláris biológiája. Tételsorok mindenkinek a honlapon:
 MOLEKULÁRIS BIOLÓGIA Előadások témája: Elsősorban a DNS, a gének és genomok molekuláris biológiája Előadásokra járni kötelező, de nincs névsor olvasás. Zárthelyi dolgozat nincs. Vegyész és hidrobiológus
MOLEKULÁRIS BIOLÓGIA Előadások témája: Elsősorban a DNS, a gének és genomok molekuláris biológiája Előadásokra járni kötelező, de nincs névsor olvasás. Zárthelyi dolgozat nincs. Vegyész és hidrobiológus
Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015
 2015") Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015 A kérdés 1. A sejtről általában, a szervetlen alkotórészeiről, a vízről részletesen. 2. A sejtről általában, a szervetlen alkotórészeiről,
Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015 A kérdés 1. A sejtről általában, a szervetlen alkotórészeiről, a vízről részletesen. 2. A sejtről általában, a szervetlen alkotórészeiről,
Fehérjeszerkezet, és tekeredés. Futó Kinga
 Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Makromolekulák. Fehérjetekeredé. rjetekeredés. Biopolimer. Polimerek
 Biopolimerek Makromolekulá Makromolekulák. Fehé Fehérjetekeredé rjetekeredés. Osztódó sejt magorsófonala 2011. November 16. Huber Tamá Tamás Dohány levél epidermális sejtjének aktin hálózata Bakteriofágból
Biopolimerek Makromolekulá Makromolekulák. Fehé Fehérjetekeredé rjetekeredés. Osztódó sejt magorsófonala 2011. November 16. Huber Tamá Tamás Dohány levél epidermális sejtjének aktin hálózata Bakteriofágból
ANATÓMIA FITNESS AKADÉMIA
 ANATÓMIA FITNESS AKADÉMIA sejt szövet szerv szervrendszer sejtek általános jellemzése: az élet legkisebb alaki és működési egysége minden élőlény sejtes felépítésű minden sejtre jellemző: határoló rendszer
ANATÓMIA FITNESS AKADÉMIA sejt szövet szerv szervrendszer sejtek általános jellemzése: az élet legkisebb alaki és működési egysége minden élőlény sejtes felépítésű minden sejtre jellemző: határoló rendszer
Génmódosítás: bioszféra
 bioszféra Génmódosítás: Nagy butaság volt politikusaink részérôl az alaptalan GMO-ellenesség alaptörvényben való rögzítése. A témával foglalkozó akadémikusok véleménye külföldön és Magyarországon egészen
bioszféra Génmódosítás: Nagy butaság volt politikusaink részérôl az alaptalan GMO-ellenesség alaptörvényben való rögzítése. A témával foglalkozó akadémikusok véleménye külföldön és Magyarországon egészen
BIOLÓGIA. PRÓBAÉRETTSÉGI 2004. május EMELT SZINT. 240 perc
 PRÓBAÉRETTSÉGI 2004. május BIOLÓGIA EMELT SZINT 240 perc Útmutató A feladatok megoldására 240 perc fordítható, az id leteltével a munkát be kell fejezni. A feladatok megoldási sorrendje tetsz leges. A
PRÓBAÉRETTSÉGI 2004. május BIOLÓGIA EMELT SZINT 240 perc Útmutató A feladatok megoldására 240 perc fordítható, az id leteltével a munkát be kell fejezni. A feladatok megoldási sorrendje tetsz leges. A
Génátvitel magasabb rendű állatokba elméleti megfontolások, gyakorlati eredmények és génterápiás lehetőségek
 MEZÕGAZDASÁGI BIOTECHNOLÓGIÁK Génátvitel magasabb rendű állatokba elméleti megfontolások, gyakorlati eredmények és génterápiás lehetőségek Tárgyszavak: génátvitel; transzfekció; transzgenézis; génterápia;
MEZÕGAZDASÁGI BIOTECHNOLÓGIÁK Génátvitel magasabb rendű állatokba elméleti megfontolások, gyakorlati eredmények és génterápiás lehetőségek Tárgyszavak: génátvitel; transzfekció; transzgenézis; génterápia;
CIÓ A GENETIKAI INFORMÁCI A DNS REPLIKÁCI
 A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
Ésszerű és korszerű táplálkozás élelmiszer általi fertőzésveszély és egyéb kockázatok társadalmi reagálás
 ÉLELMISZERIPARI BIOTECHNOLÓGIÁK Ésszerű és korszerű táplálkozás élelmiszer általi fertőzésveszély és egyéb kockázatok társadalmi reagálás Tárgyszavak: táplálkozás; egészség; fertőzés; mérgezés; élelmiszer-biztonság;
ÉLELMISZERIPARI BIOTECHNOLÓGIÁK Ésszerű és korszerű táplálkozás élelmiszer általi fertőzésveszély és egyéb kockázatok társadalmi reagálás Tárgyszavak: táplálkozás; egészség; fertőzés; mérgezés; élelmiszer-biztonság;
Élettan. előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45
 Élettan előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45 oktató: Dr. Tóth Attila, adjunktus ELTE TTK Biológiai Intézet, Élettani és Neurobiológiai tanszék
Élettan előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45 oktató: Dr. Tóth Attila, adjunktus ELTE TTK Biológiai Intézet, Élettani és Neurobiológiai tanszék
Eszközszükséglet: Szükséges anyagok: tojás, NaCl, ammónium-szulfát, réz-szulfát, ólom-acetát, ecetsav, sósav, nátrium-hidroxid, desztillált víz
 A kísérlet, megnevezés, célkitűzései: Fehérjék tulajdonságainak, szerkezetének vizsgálata. Környezeti változások hatásának megfigyelése a fehérjék felépítésében. Eszközszükséglet: Szükséges anyagok: tojás,
A kísérlet, megnevezés, célkitűzései: Fehérjék tulajdonságainak, szerkezetének vizsgálata. Környezeti változások hatásának megfigyelése a fehérjék felépítésében. Eszközszükséglet: Szükséges anyagok: tojás,
KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA
 ÉRETTSÉGI VIZSGA 2010. május 12. BIOLÓGIA KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA 2010. május 12. 8:00 Az írásbeli vizsga időtartama: 120 perc Pótlapok száma Tisztázati Piszkozati OKTATÁSI ÉS KULTURÁLIS MINISZTÉRIUM
ÉRETTSÉGI VIZSGA 2010. május 12. BIOLÓGIA KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA 2010. május 12. 8:00 Az írásbeli vizsga időtartama: 120 perc Pótlapok száma Tisztázati Piszkozati OKTATÁSI ÉS KULTURÁLIS MINISZTÉRIUM
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
6. Zárványtestek feldolgozása
 6. Zárványtestek feldolgozása... 1 6.1. A zárványtestek... 1 6.1.1. A zárványtestek kialakulása... 2 6.1.2. A feldolgozási technológia... 3 6.1.2.1. Sejtfeltárás... 3 6.1.2.2. Centrifugálás, tisztítás...
6. Zárványtestek feldolgozása... 1 6.1. A zárványtestek... 1 6.1.1. A zárványtestek kialakulása... 2 6.1.2. A feldolgozási technológia... 3 6.1.2.1. Sejtfeltárás... 3 6.1.2.2. Centrifugálás, tisztítás...
A koleszterin-anyagcsere szabályozása (Csala Miklós)
") A koleszterin-anyagcsere szabályozása (Csala Miklós) A koleszterin fontos építőeleme az emberi sejteknek, fontos szerepe van a biológiai membránok fluiditásának szabályozásában. E mellett hormonok és epesavak
A koleszterin-anyagcsere szabályozása (Csala Miklós) A koleszterin fontos építőeleme az emberi sejteknek, fontos szerepe van a biológiai membránok fluiditásának szabályozásában. E mellett hormonok és epesavak
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
Kereskedelmi forgalomban lévő rekombináns gyógyszerkészítmények
 Kereskedelmi forgalomban lévő rekombináns gyógyszerkészítmények Írta: Barta Zsolt Biomérnök hallgató 2007 Tartalomjegyzék 1 Rekombináns inzulin [1]... 3 2 A humán növekedési hormon rekombináns módon történő
Kereskedelmi forgalomban lévő rekombináns gyógyszerkészítmények Írta: Barta Zsolt Biomérnök hallgató 2007 Tartalomjegyzék 1 Rekombináns inzulin [1]... 3 2 A humán növekedési hormon rekombináns módon történő
Heszky László Transzgénikus növények - az emberiség diadala vagy félelme?
 Heszky László Transzgénikus növények - az emberiség diadala vagy félelme? I. Tudományos alapok I. 1. Amit a földi élet titkáról tudunk 1. ábra Nyolc éves voltam, amikor 1953. február 28-án, egy szombat
Heszky László Transzgénikus növények - az emberiség diadala vagy félelme? I. Tudományos alapok I. 1. Amit a földi élet titkáról tudunk 1. ábra Nyolc éves voltam, amikor 1953. február 28-án, egy szombat
Zöld élelmiszerek Tibet, Hawaii, Peru, Tajvan, Ecuador, Kalifornia és Brazília
 Zöld élelmiszerek Tibet, Hawaii, Peru, Tajvan, Ecuador, Kalifornia és Brazília A természet kvintesszenciái A kvintesszencia szó Arisztotelész természeti filozófiájából indul ki és a latin quinta essentia,
Zöld élelmiszerek Tibet, Hawaii, Peru, Tajvan, Ecuador, Kalifornia és Brazília A természet kvintesszenciái A kvintesszencia szó Arisztotelész természeti filozófiájából indul ki és a latin quinta essentia,
NAP EGÉSZSÉG NAP SZÉPSÉG NAPOZÁS KOCKÁZATAI AZ UV SUGARAK ÉS AZOK HATÁSAI A BŐRRE
 ÚJ NAPOZÓ TERMÉKEK 2012 ELÉRHETö: 2012 május AZ UV SUGARAK ÉS AZOK HATÁSAI A BŐRRE NAP EGÉSZSÉG Az UV sugarak stimulálják a D vitamint termelődését, amely megköti a kálciumot és segít megerősíteni a csontjainkat.
ÚJ NAPOZÓ TERMÉKEK 2012 ELÉRHETö: 2012 május AZ UV SUGARAK ÉS AZOK HATÁSAI A BŐRRE NAP EGÉSZSÉG Az UV sugarak stimulálják a D vitamint termelődését, amely megköti a kálciumot és segít megerősíteni a csontjainkat.
1. SEJT-, ÉS SZÖVETTAN. I. A sejt
 1. SEJT-, ÉS SZÖVETTAN SZAKMAI INFORMÁCIÓTARTALOM I. A sejt A sejt cellula az élő szervezet alapvető szerkezeti és működési egysége, amely képes az önálló anyag cserefolyamatokra és a szaporodásra. Alapvetően
1. SEJT-, ÉS SZÖVETTAN SZAKMAI INFORMÁCIÓTARTALOM I. A sejt A sejt cellula az élő szervezet alapvető szerkezeti és működési egysége, amely képes az önálló anyag cserefolyamatokra és a szaporodásra. Alapvetően
Hazai méhészeti genomikai és genetikai vizsgálatok
 AKÁCKÖRÚTON Hazai méhészeti genomikai és genetikai vizsgálatok Előző cikkünkben arról írtunk, milyen új eszköztárral rendelkezünk a XXI. században a genetikai vizsgálatok területén, és mit adhat a molekuláris
AKÁCKÖRÚTON Hazai méhészeti genomikai és genetikai vizsgálatok Előző cikkünkben arról írtunk, milyen új eszköztárral rendelkezünk a XXI. században a genetikai vizsgálatok területén, és mit adhat a molekuláris
Biokatalízis, biokonverziók, biotranszformációk Rákhely, Gábor
 Biokatalízis, biokonverziók, biotranszformációk Rákhely, Gábor Biokatalízis, biokonverziók, biotranszformációk Rákhely, Gábor Publication date 2012 Szerzői jog 2012 Szegedi Tudományegyetem TÁMOP-4.1.2.A/1-11/1
Biokatalízis, biokonverziók, biotranszformációk Rákhely, Gábor Biokatalízis, biokonverziók, biotranszformációk Rákhely, Gábor Publication date 2012 Szerzői jog 2012 Szegedi Tudományegyetem TÁMOP-4.1.2.A/1-11/1
TUMORELLENES ANTIBIOTIKUMOK
 TUMORELLENES ANTIBIOTIKUMOK A rák gyógyszeres kezelése nem megoldott - néhány antibiotikum segíthet átmenetileg. Nincs igazán jó és egyértelmű terápiája, alternatívák: - sebészeti beavatkozás - besugárzás
TUMORELLENES ANTIBIOTIKUMOK A rák gyógyszeres kezelése nem megoldott - néhány antibiotikum segíthet átmenetileg. Nincs igazán jó és egyértelmű terápiája, alternatívák: - sebészeti beavatkozás - besugárzás
A biológia tudománya az élők világát két alapvető egységre ún. doménre, birodalomra - osztja: a prokariótákra és az eukariótákra.
 A prokarióták Szerkesztette: Vizkievicz András A biológia tudománya az élők világát két alapvető egységre ún. doménre, birodalomra - osztja: a prokariótákra és az eukariótákra. A prokarióták (legszembetűnőbb)
A prokarióták Szerkesztette: Vizkievicz András A biológia tudománya az élők világát két alapvető egységre ún. doménre, birodalomra - osztja: a prokariótákra és az eukariótákra. A prokarióták (legszembetűnőbb)
1. Az élő szervezetek felépítése és az életfolyamatok 17
 Élődi Pál BIOKÉMIA vomo; Akadémiai Kiadó, Budapest 1980 Tartalom Bevezetés 1. Az élő szervezetek felépítése és az életfolyamatok 17 Mi jellemző az élőre? 17. Biogén elemek 20. Biomolekulák 23. A víz 26.
Élődi Pál BIOKÉMIA vomo; Akadémiai Kiadó, Budapest 1980 Tartalom Bevezetés 1. Az élő szervezetek felépítése és az életfolyamatok 17 Mi jellemző az élőre? 17. Biogén elemek 20. Biomolekulák 23. A víz 26.
Poligénes v. kantitatív öröklődés
 1. Öröklődés komplexebb sajátosságai 2. Öröklődés molekuláris alapja Poligénes v. kantitatív öröklődés Azok a tulajdonságokat amelyek mértékegységgel nem, vagy csak nehezen mérhetők, kialakulásuk kevéssé
1. Öröklődés komplexebb sajátosságai 2. Öröklődés molekuláris alapja Poligénes v. kantitatív öröklődés Azok a tulajdonságokat amelyek mértékegységgel nem, vagy csak nehezen mérhetők, kialakulásuk kevéssé
Ipari n-hexán-frakcióban, mely 2 % C 6 -izomert tartalmazott néhány tized % pentán mellett, a benzol koncentrációját 0-5 % között, a C 2 H 5 SH-ként
 T 43524 OTKA Zárójelentés 2003-2006 Az egyre szigorodó környezetvédelmi előírások a gépjárművek káros emissziójának egyre alacsonyabb határértékeit szabják meg, melyeket a motorhajtóanyagok minőségjavításával,
T 43524 OTKA Zárójelentés 2003-2006 Az egyre szigorodó környezetvédelmi előírások a gépjárművek káros emissziójának egyre alacsonyabb határértékeit szabják meg, melyeket a motorhajtóanyagok minőségjavításával,
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak?
 A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
Sporttáplálkozás. Étrend-kiegészítők. Készítette: Honti Péter dietetikus. 2015. július
 Sporttáplálkozás Étrend-kiegészítők Készítette: Honti Péter dietetikus 2015. július Étrend-kiegészítők Élelmiszerek, amelyek a hagyományos étrend kiegészítését szolgálják, és koncentrált formában tartalmaznak
Sporttáplálkozás Étrend-kiegészítők Készítette: Honti Péter dietetikus 2015. július Étrend-kiegészítők Élelmiszerek, amelyek a hagyományos étrend kiegészítését szolgálják, és koncentrált formában tartalmaznak
Bioaktív peptidek technológiáinak fejlesztése
 Bioaktív peptidek technológiáinak fejlesztése BIOAKTÍV PEPTIDEK A kolosztrum kitűnő fehérjeforrás, melyben az esszenciális aminosavak és más organikus nitrogén-forrásként szolgáló vegyületek rendkívül
Bioaktív peptidek technológiáinak fejlesztése BIOAKTÍV PEPTIDEK A kolosztrum kitűnő fehérjeforrás, melyben az esszenciális aminosavak és más organikus nitrogén-forrásként szolgáló vegyületek rendkívül
A gidrán fajta genetikai változatosságának jellemzése mitokondriális DNS polimorfizmusokkal Kusza Szilvia Sziszkosz Nikolett Mihók Sándor,
 1 A gidrán fajta genetikai változatosságának jellemzése mitokondriális DNS polimorfizmusokkal Kusza Szilvia Sziszkosz Nikolett Mihók Sándor, (Debreceni Egyetem Állattenyésztéstani Tanszék) A bármilyen
1 A gidrán fajta genetikai változatosságának jellemzése mitokondriális DNS polimorfizmusokkal Kusza Szilvia Sziszkosz Nikolett Mihók Sándor, (Debreceni Egyetem Állattenyésztéstani Tanszék) A bármilyen
KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA
 É RETTSÉGI VIZSGA 2015. október 21. BIOLÓGIA KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA 2015. október 21. 14:00 Az írásbeli vizsga időtartama: 120 perc Pótlapok száma Tisztázati Piszkozati EMBERI ERŐFORRÁSOK MINISZTÉRIUMA
É RETTSÉGI VIZSGA 2015. október 21. BIOLÓGIA KÖZÉPSZINTŰ ÍRÁSBELI VIZSGA 2015. október 21. 14:00 Az írásbeli vizsga időtartama: 120 perc Pótlapok száma Tisztázati Piszkozati EMBERI ERŐFORRÁSOK MINISZTÉRIUMA
A basidiomycota élesztőgomba, a Filobasidium capsuligenum IFM 40078 törzse egy olyan
 A basidiomycota élesztőgomba, a Filobasidium capsuligenum IFM 40078 törzse egy olyan fehérjét (FC-1 killer toxint) választ ki a tápközegbe, amely elpusztítja az opportunista patogén Cryptococcus neoformans-t.
A basidiomycota élesztőgomba, a Filobasidium capsuligenum IFM 40078 törzse egy olyan fehérjét (FC-1 killer toxint) választ ki a tápközegbe, amely elpusztítja az opportunista patogén Cryptococcus neoformans-t.
A BIZOTTSÁG 2009/120/EK IRÁNYELVE
 2009.9.15. Az Európai Unió Hivatalos Lapja L 242/3 IRÁNYELVEK A BIZOTTSÁG 2009/120/EK IRÁNYELVE (2009. szeptember 14.) a fejlett terápiás gyógyszerkészítmények tekintetében az emberi felhasználásra szánt
2009.9.15. Az Európai Unió Hivatalos Lapja L 242/3 IRÁNYELVEK A BIZOTTSÁG 2009/120/EK IRÁNYELVE (2009. szeptember 14.) a fejlett terápiás gyógyszerkészítmények tekintetében az emberi felhasználásra szánt
Oktatáskutató és Fejlesztő Intézet TÁMOP-3.1.1-11/1-2012-0001 XXI. századi közoktatás (fejlesztés, koordináció) II. szakasz KÉMIA 4.
 II. szakasz KÉMIA 4.") Oktatáskutató és Fejlesztő Intézet TÁMOP-3.1.1-11/1-2012-0001 XXI. századi közoktatás (fejlesztés, koordináció) II. szakasz KÉMIA 4. MINTAFELADATSOR KÖZÉPSZINT 2015 Az írásbeli vizsga időtartama: 120 perc
Oktatáskutató és Fejlesztő Intézet TÁMOP-3.1.1-11/1-2012-0001 XXI. századi közoktatás (fejlesztés, koordináció) II. szakasz KÉMIA 4. MINTAFELADATSOR KÖZÉPSZINT 2015 Az írásbeli vizsga időtartama: 120 perc
Az összetett szavak orvosi és nyelvészszemmel
 Bősze Péter Laczkó Krisztina Az összetett szavak orvosi és nyelvészszemmel BEVEZETÉS A következőkben egy helyesírási kérdőíves vizsgálat eredményét ismertetjük, amely az összetett szavak köréből tartalmazott
Bősze Péter Laczkó Krisztina Az összetett szavak orvosi és nyelvészszemmel BEVEZETÉS A következőkben egy helyesírási kérdőíves vizsgálat eredményét ismertetjük, amely az összetett szavak köréből tartalmazott
Az élő szervezetek menedzserei, a hormonok
 rekkel exponálunk a munka végén) és azt utólag kivonjuk digitálisan a képekből. A zajcsökkentés dandárját mindig végezzük a raw-képek digitális előhívása során, mert ez okozza a legkevesebb jelvesztést
rekkel exponálunk a munka végén) és azt utólag kivonjuk digitálisan a képekből. A zajcsökkentés dandárját mindig végezzük a raw-képek digitális előhívása során, mert ez okozza a legkevesebb jelvesztést
Konferencia a tapasztalatok jegyében
 Konferencia a tapasztalatok jegyében 2010. november Dornbach Ildikó szakmai igazgató Új biológia, új fizika, régi beidegzések Edzőink felbecsülhetetlen értékű tevékenysége Köztársasági Érdemrendet minden
Konferencia a tapasztalatok jegyében 2010. november Dornbach Ildikó szakmai igazgató Új biológia, új fizika, régi beidegzések Edzőink felbecsülhetetlen értékű tevékenysége Köztársasági Érdemrendet minden