Az inzulin receptor szignál transzdukciója
|
|
|
- Endre Fekete
- 9 évvel ezelőtt
- Látták:
Átírás
1 Az inzulin receptor szignál transzdukciója A szeminárium célja, hogy kísérleti adatok segítségével áttekintse az inzulin receptor szignál transzdukcióját, és annak néhány, a II. típusú cukorbetegség kialakulásában esetleg szerepet játszó patológiás vonatkozását. A cukorbetegség részletes tárgyalását természetesen meghagyjuk a további évek klinikai oktatása számára, de a betegség rövid áttekintése segíthet a molekuláris mechanizmusok és a klinikai megfigyelések, tapasztalatok összekapcsolásában. A diabetes mellitus egy súlyos krónikus betegség, mely a évesek 6-11 %-át, a 65 éven felülieknek pedig 20 %-át érinti szerte a világon. A cukorbetegség, és főként egyik fajtája, a II. típusú diabetes világméretű problémát jelent, köszönhetően a hosszabb élettartamnak, és annak, hogy egyre több társadalom csatlakozik az un. nyugati életmódhoz. Történelem Bár már a régi kínai irodalom is tartalmaz utalásokat egy halálos, fokozott vizeléssel járó, senyvesztő betegségre, az első egyértelműen diabeteses esetleírás az Ebers papirusztekercsben jelent meg (kb. i.e. 1500). A szó diabetes az Apamaia-i Demetrios-tól származik, a görög diabeinein szóból, melynek jelentése szifon, és arra az eszközre utal, amellyel a megnövekedett mennyiségű vizeletet felfogták. A századokkal később hozzáadott latin megjelölés mellitus pedig mézet jelent, utalván a cukorbetegek vizeletének édes ízére. A diabetes mellitus és egy másik fokozott vizeléssel járó endokrin betegség, a diabetes insipidus egyértelmű elkülönítése a XVIII. századra tehető. Ezt követően beköszöntött a kísérletezésen alapuló modern kor ban megállapították, hogy a cukorbetegek vizeletének édes íze a glukóz tartalomnak köszönhető, és hamarosan leírták a máj és a hasnyálmirigy, azon belül is a szigetsejtek szerepét a betegségben. A következő mérföldkő egy aktív hasnyálmirigykivonat készítése, és alkalmazása kutyák, majd emberek kezelésére, F. Banting, Ch. Best, JR Macleod, és JB Collip nevéhez fűződik. Az 1922-ben történt első sikeres inzulin terápiát megelőzően a gyermekkorban akutan fellépő diabetes a halálos ítélettel volt egyenértékű. Ha nem is az első fellépő diabeteses kómában, de néhány éven belül biztosan meghaltak. Az inzulin terápia bevezetésével a betegség többé nem járt gyors halállal. A hosszabb túléléssel azonban újabb komplikációk jelentkeztek, vakság, veseelégtelenség, végtag elvesztése, szívinfarktus, ill. gutaütés. A cukorbetegség így egy rövid, visszataszító és fájdalmas betegségből (ahogy Arateus jellemezte az I. sz-ban) egy olyan krónikus betegséggé alakult át, amely hosszú, visszataszító, és még fájdalmasabb. Jóllehet a korai vizsgálatok az un. fiatalkori cukorbetegségre korlátozódtak, ez csak egy része a történetnek. A XIX. sz. végefelé Lacereaux a diabetes mellitus egy másik formáját is leírta, ez idősebb, főként jómódú emberekben jelentkezett, és étrendi változtatással is javítható volt. Ezt a formát kövér diabetes -nek nevezte, megkülönböztetve a gyorsan halálos, sovány diabetes -től, amely gyerekekben és fiatalokban lépett fel. Az 1930-as években Himsworth javasolta az inzulin-szenzitív és inzulin-rezisztens diabetes elnevezéseket. A XX.század utolsó évtizedeiben leírt számos új adat olyan terápiás eljárásokat tesz lehetővé, melyek segítségével a vércukorszint a betegekben pontosabban kontrollálható, és ez kitolja a súlyos komplikációk felléptét is. A diabetes mellitust ma egy szindromaként tartják számon, ahova számos különféle betegség tartozik, és a II. típusú diabetes a szindroma leggyakoribb megjelenési formája. Egy kis klinikum A diabetes mellitushoz számos különféle betegség tartozik, melyeknek két közös jellemzője van, a legszembeszökőbben hiperglikémiaként jelentkező anyagcserezavar, és olyan késői szövődmények kifejlődésére való hajlam, mint a vaksághoz vezető retinopátia, veseelégtelenséghez vezető nefropátia, makro-, és mikroangiopátia, és neuropátia. A diabetes felléptének oka minden formában relatív vagy abszolút inzulin hiány. Osztályozás-Az I. típusú (inzulin-dependens, vagy fiatalkori forma) diabetest a hasnyálmirigy β-sejtjeinek gyakran autoimmun károsodása okozza, amelynek következménye a csökkent inzulin szekréció, és abszolút inzulin hiány. A diabeteses esetek kb %-a tartozik ide. A II. típusú diabetest genetikai és nem genetikai

2 tényezők kombinációja okozza, melyek inzulin rezisztenciához és relatív inzulin hiányhoz, tehát az inzulin rezisztencia miatt megnőtt igényhez képest inzulinhiányos állapothoz vezetnek. A pontos genetikai háttér nem ismert, de intenzív vizsgálatok tárgya. A nem genetikai tényezők közé tartozik az idősebb életkor, a magas kalóriabevitel, a túlsúly, a centrális típusú elhízás, az ülő életmód, és az alacsony születési súly. Az összes diabeteses eset %-a tartozik ide. Egyéb specifikus típusba sorolnak számos, az inzulin szekréciót vagy inzulin hatást érintő genetikai eltérést, ill. számos egyéb betegséget, amelyek másodlagosan inzulin rezisztenciához és/vagy inzulin szekréció defektushoz vezetnek. Az összes eseteknek csak 1-2 %-a sorolható ide. Terhességi diabetes esetén a diabetes a terhesség alatt lép fel először, az összes terhesség 3-5 %-ában figyelhető meg. Rendszerint szülés után normalizálódik, de az érintett nők kb. 50 %-ában évekkel később II. típusú diabetes alakul ki. A II. típusú diabetes természetrajzáról-a betegség leggyakrabban a 40 év felettieket érinti, több, mint 50 %-a 55 év felett lép fel. A betegség azonban alattomosan kezdődik, és gyakran sokáig diagnosztizálatlan marad, mert a tüneteket vagy nem ismerik fel, vagy jelentéktelennek ítélik. A II. típusú diabetest genetikai és környezeti tényezők együttesen okozzák. Egypetéjű ikrekben a konkordancia majdnem 100 %, családi halmozódást mutat (1 cukorbeteg szülő 2-szeresre, 2 cukorbeteg szülő 4-szeresre növeli az utódban való kialakulás valószínűségét), az etnikailag sokszínű populációkban előfordulása a genetikai háttérnek megfelelő, és rendkívül gyakori egyes beltenyésztett populációkban. Jóllehet a betegség iránti hajlamot egyértelműen genetikusnak tartják, a pontos gének még leírásra várnak. A betegség kialakulásában a környezeti tényezők szerepe is fontos, míg az Arizonában rezervátumban élő Pima indiánok 50 %-a cukorbeteg, a betegség szinte nem ismert a Mexikóban élő Pima indiánok között, akik viszont önfenntartásukért mezőgazdasággal foglalkoznak. Hasonlóan az életmód szerepét tükrözi az is, hogy Naurun 40 % a betegség elterjedtsége, de a bennszülött lakosság körében a betegség mindaddíg ismeretlen volt, amíg a guano kitermelés hatására át nem tértek a halászatról és vadászatról az ülő életmódra, motorozásra, magas zsírtartalmú ételekre, és el is híztak. A II. típusú cukorbetegeknél gyakran látunk elhízást, és különösen annak centrális, vagy hasra lokalizálódó formáját, de az összes elhízottaknak csak kis %-ában lép fel a betegség. A II. típusú diabetes mellitus kialakulása. A betegség kialakulásának háttere az inzulin rezisztencia, vagyis a szervezet inzulin hatásokra való csökkent érzékenysége, amely az emberekben sok évvel az első tünetek fellépte előtt már kimutatható. Ezt az állapotot az elhízás valószínűleg tovább rontja. A hasnyálmirigy β-sejtjei kompenzációként megnövelik az alap-, és étkezés után szekretált inzulin mennyiségét. Egyszercsak (nem tudni pontosan, mikor és miért) a β-sejtek nem képesek ezt a kompenzációt tovább folytatni, így glukóz intolerancia alkul ki. Évente a glukóz intoleranciát mutató betegek kb %-a progrediál diabetes mellitusba, amely egyre súlyosbodik, ahogy az inzulin rezisztencia tovább fokozódik. Késői stádiumban a β- sejtek működése teljesen elégtelenné válik, és a betegnek nagy mennyiségű exogén inzulinra lehet szüksége. Az inzulin receptor szignál transzdukciója Inzulin receptor Az inzulin receptor transzmembrán glikoprotein, amely diszulfid hidakkal kovalensen összekapcsolt két α és két β alegységből áll. Az inzulin az extracelluláris α alegységhez kötődik, a β alegység egy rövid extracelluláris részből, egy transzmembrán doménből, és egy intracelluláris részből áll, amely az inzulin által regulált tirozin kináz aktivitásért felelős. Inzulin távollétében az α alegység gátolja a β alegység tirozin kináz aktivitását. Az inzulin α alegységhez való kötődése olyan konformációváltozást okoz, amelynek hatására a β alegység felszabadul a gátlás alól. 2

3 Ekkor a receptor autofoszforilálódik, melynek során az egyik β alegység tirozin oldalláncokat foszforilál a szomszédos β alegységben. Ez az autofoszforiláció szabályozza a C-terminális régió tirozin kináz aktivitását, és időben megelőzi a további szubsztrátok tirozin-foszforilációját. A tirozinok mellett a receptorban szerin oldalláncok is foszforilálódhatnak, pl. camp dependens protein kináz, vagy protein kináz C hatására. A tirozin foszforilációval ellentétben a szerin foszforiláció gátolja a receptor tirozin kináz aktivitását. A II típusú cukorbetegségben megfigyelt csökkent inzulin receptor tirozin kináz aktivitás hátterében leírtak olyan mechanizmusokat, amelyekben a receptor szerin foszforilációja játszik szerepet. Inzulin antagonista hormonok, így pl. az adrenalin is gátolhatják az inzulin hatás kifejlődését az inzulin receptor szerin foszforilációján keresztül. Az inzulin receptor mutációja ritka, és a leírt fenotípusok súlyos inzulin rezisztenciához és korán kifejlődő cukorbetegséghez vezetnek. Ezeket a mutációkat az egyéb típusú diabetes mellitushoz sorolják, és mindeddig egyetlen inzulin receptort érintő mutáció sem mutatott összefüggést a II. típusú cukorbetegség felléptével. Az inzulin receptor tirozin kináz foszforilációs célpontjai Miután aktiválódott, az inzulin receptor számos szubsztrátot foszforilál, köztük az inzulin receptor szubsztrát fehérjecsalád tagjait (IRS-1, IRS-2, IRS-3...). Az IRS fehérjék kötődnek az inzulin receptor foszfotirozin oldalláncaihoz, és miután a receptor őket is foszforilálta, további SH2 (Src homology) domént tartalmazó szignál transzdukciós fehérjéket dokkolnak, így pl. a foszfatidilinozitol 3 kinázt (PI3K), SHP2-t (protein tirozin foszfatáz), Grb2-t (adapter molekula). A különböző IRS fehérjék szöveti eloszlása eltérő, az IRS-1 és IRS-2 mindenhol megtalálható, az IRS-3 főként a zsírsejtekben, az IRS-4 pedig főként a vesében és az agyban. Az IRS fehérjék körében ismert néhány genetikai polimorfizmus, de inkább az jellemző, hogy nem egy, hanem több polimorfizmus együttes megléte fordul elő halmozottan a II. típusú diabeteses populációkban. Az IRS fehérjék foszforilációját követő molekuláris események Az inzulin különféle biológiai hatásai, mint pl. az anyagcserehatások, vagy a növekedésre gyakorolt hatás valószínűleg a szignál transzdukciónak azon a szintjén válnak szét, amikor az SH2 adapter fehérjék SH2 doménjükön keresztül a foszforilált tirozin oldalláncokhoz kötődnek. A Grb-2 egy SH2 adapter protein, amely inzulin hatáskor IRS fehérjéhez kötődik, és ez a MAP kináz kaszkád aktivációját indítja be. Ez a kaszkád az inzulin növekedést fokozó hatásában játszik szerepet, és egyéb növekedési faktor receptorok aktivációjakor is működésbe lép. Az inzulin metabolikus hatásaihoz vezető szignál transzdukciós útnak kiemelkedően fontos eleme a foszfatidilinozitol-3-kináz (PI3K). A PI3K egy heterodimer, egy regulátor alegységből (p85), és egy katalitikus alegységből (p110) áll. A regulátor alegység kötődik az IRS fehérjéhez, és a katalitikus alegység a sejtmembránban lévő foszfatidilinozitolokat foszforilálja az inozitol gyűrű 3. pozíciójában. A foszfatidilinozitol 3,4,5-triszfoszfát aktiválja a foszfatidilinozitol-3-kináz dependens kináz 1-et (PDK-1), ami a továbbiakban számos enzim aktivációjához vezet, közöttük a protein kináz B, a protein kináz C ξ, és γ formája, az mtor (mammalian target of Rapamycin), és a p70s6 kináz. A protein kináz B egy Ser/Thr kináz, és foszforilációs szubsztrátjai közé tartozik a glikogén szintáz kináz-3(gsk-3), a camp response element binding protein (crebp), és bizonyos transzkripciós faktorok. Az mtor a PI3K fehérjecsalád tagja, amely fehérjék Ser oldalláncán foszforilál, és a p70s6 kináz közvetítésével fokozza a transzlációt. Ezek a kinázok közvetítik az inzulin 3

4 anyagcserehatásait, így pl. a GLUT 4 glukóztranszporter transzlokációját az intracelluláris raktárakból a plazma membránra, a glikogén szintézis és a fehérje szintézis fokozódását, és bizonyos gének (pl. a glukokináz) expressziójának indukálását. Az inzulin glukóz transzportot stimuláló hatásában valószínűleg PI3K-tól független tényezők is szerepet játszanak. Az alábbi ábra összefoglalja az inzulin hatásait. A hasnyálmirigy β-sejtjeinek diszfunkciója cukorbetegségben A β-sejtek feladata, hogy az inzulin szekrécióját a vércukorszintnek megfelelően szabályozzák. A β-sejtekben a glukózt a glukokináz foszforilálja glukóz-6-foszfáttá, és ez a lépés határozza meg a glikolízis és a piruvát keletkezés sebességét. Így a vércukorszint növekedésekor a piruvát keletkezés sebessége is nő. Más sejtekkel összehasonlítva a β-sejtekben a piruvát szokatlanul nagy hányada lép be a mitokondriumba, acetil-koa-vá alakul, és belép a citrát-körbe, így jelentős mennyiségű ATP keletkezik és lép ki a citoplazmába. A citoplazmatikus ATP/ADP hányados növekedésével záródnak az ATP-szenzitív K + csatornák, és a plazmamembrán depolarizálódik. Ekkor kinyílnak a feszültségfüggő Ca 2+ csatornák, és a citoplazmatikus Ca 2+ szint növekedése az inzulint tartalmazó szekretoros granulumok exocitózisát idézi elő. A cukorbetegség néhány ritka formájának olyan genetikai károsodás az oka, amely a β-sejtek inzulin szekréciójának szabályozását érinti valamely ponton (pl. glukokináz, transzkripciós faktorok). II. típusú cukorbetegségben a genetikai eltérés nem alapjaiban károsítja a szekréciós mechanizmust, hanem inkább a β-sejtek inzulin rezisztenciához való hosszú távú adaptációját teszi lehetetlenné. Az inzulin rezisztens egyedek jelentős részében a β-sejtek kompenzációs működésének köszönhetően 4

5 cukorbetegség talán soha ki se alakul. Ilyenkor a kompenzatorikus inzulin hiperszekréció hátterében a β-sejtek össztömegének megnövekedése, és a β-sejtek glukóz metabolizmusában részt vevő kulcsenzimek expressziójának megváltozása játszik szerepet, de a folyamat pontos szabályozása nem ismert. Jóllehet számos β-sejtfunkciót érintő mutációról ismert, hogy növelik a II. típusú cukorbetegség kialakulásának kockázatát, a betegség fellépésének molekuláris háttere még nem világos. Megbeszélésre javasolt témák és kísérletek 1, Röviden ismételjék át azokat az inzulin hatásokat, melyek fontosak a vércukorszint szabályozása szempontjából: a, glukóz felvétel fokozódása az izomszövetben és a zsírszövetben, b, a csökkenő zsírsav felhasználás hatására bekövetkező fokozott glukóz felhasználás a szövetekben, c, a máj csökkent glukóz termelése 2, A különböző IRS fehérjék szerepét az inzulin szignál transzdukciójában és a cukorbetegség kialakulásában knock out egerekben vizsgálták, javasolt az alábbi kísérleti eredmények áttekintése (az ábrák a mellékletben találhatók): Araki E, et al. Alternative pathway of insulin signalling in mice with targeted disruption of the IRS-1 gene. Nature 1994, 372: , Figs 1f, 2-4. Befolyásolja-e az IRS-1 gén az egerek növekedését? Inzulin rezisztensek-e az IRS-1 knock out egerek? IRS-1 hiányában megtörténik-e az inzulin receptor és a PI3K foszforilációja? Hogyan lehetséges, hogy az IRS-1 hiányos egerek nem cukorbetegek? Withers DJ, et al. Disruption of IRS-2 causes type 2 diabetes in mice. Nature 1998, 391: , Figs 1e, 2-4. Háttérinformáció: az euglikémiás hiperinzulinémiás clamp módszert az inzulin rezisztencia kimutatására és mennyiségi jellemzésére használják, lényege, hogy exogén inzulin alkalmazása mellett azt a mennyiségű glukózt mérik meg, aminek beadása ahhoz szükséges, hogy a vércukorszint a mesterséges hiperinzulinémia mellett se csökkenjen. Független módszerrel mérik meg a máj által közben leadott glukóz mennyiségét, hiszen a szervek glukóz felhasználása mellett ez is befolyásolja a szervezet összes glukóz igényét, és eredményként már a korrigált összeget adják meg. Az IRS-2 hiánya befolyásolja-e a növekedést? Milyen különbségek és hasonlóságok találhatók az IRS-1 és az IRS-2 hiányos egerek szénhidrát anyagcseréjében? Inzulin hatásra foszforilálódik-e a PI3K? Milyen eltérés található az IRS-1 és az IRS-2 hiányos egerek hasnyálmirigyében? Liu SCH, et al. Insulin receptor substrate 3 is not essential for growth or glucose homeostasis. J Biol Chem 1999, 274: , Figs 2-6, Table 1. Az előző cikkekben leírtakhoz hasonló módszerekkel IRS-3 knock out egerekben nem találnak eltérést sem a növekedési ütem, sem a szénhidrát homeosztázis tekintetében. 3, A protein kináz B szerepe a glikogén szintáz kináz-3 inzulin hatásra bekövetkező inaktivációjában: Cross DAE, et al. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378: , Figs 1, 2, 3a. 5

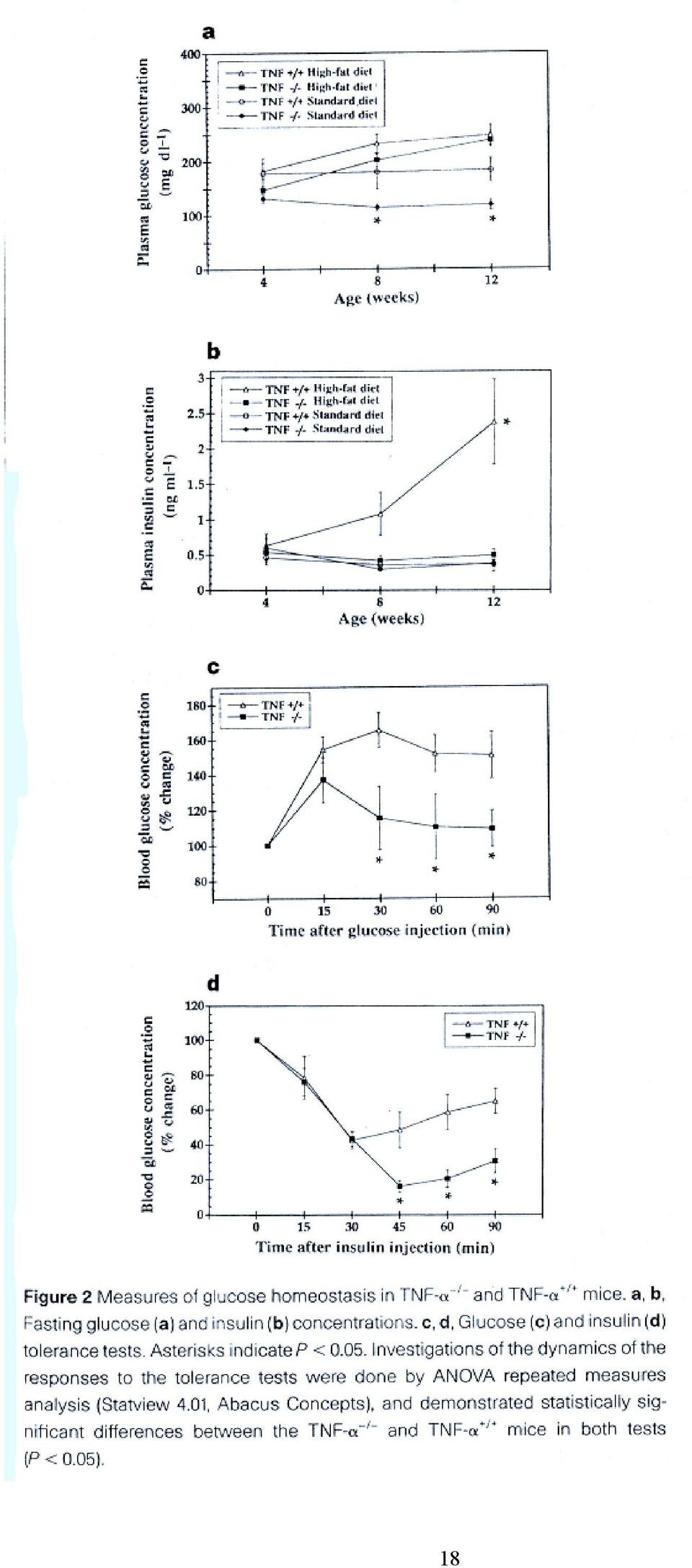
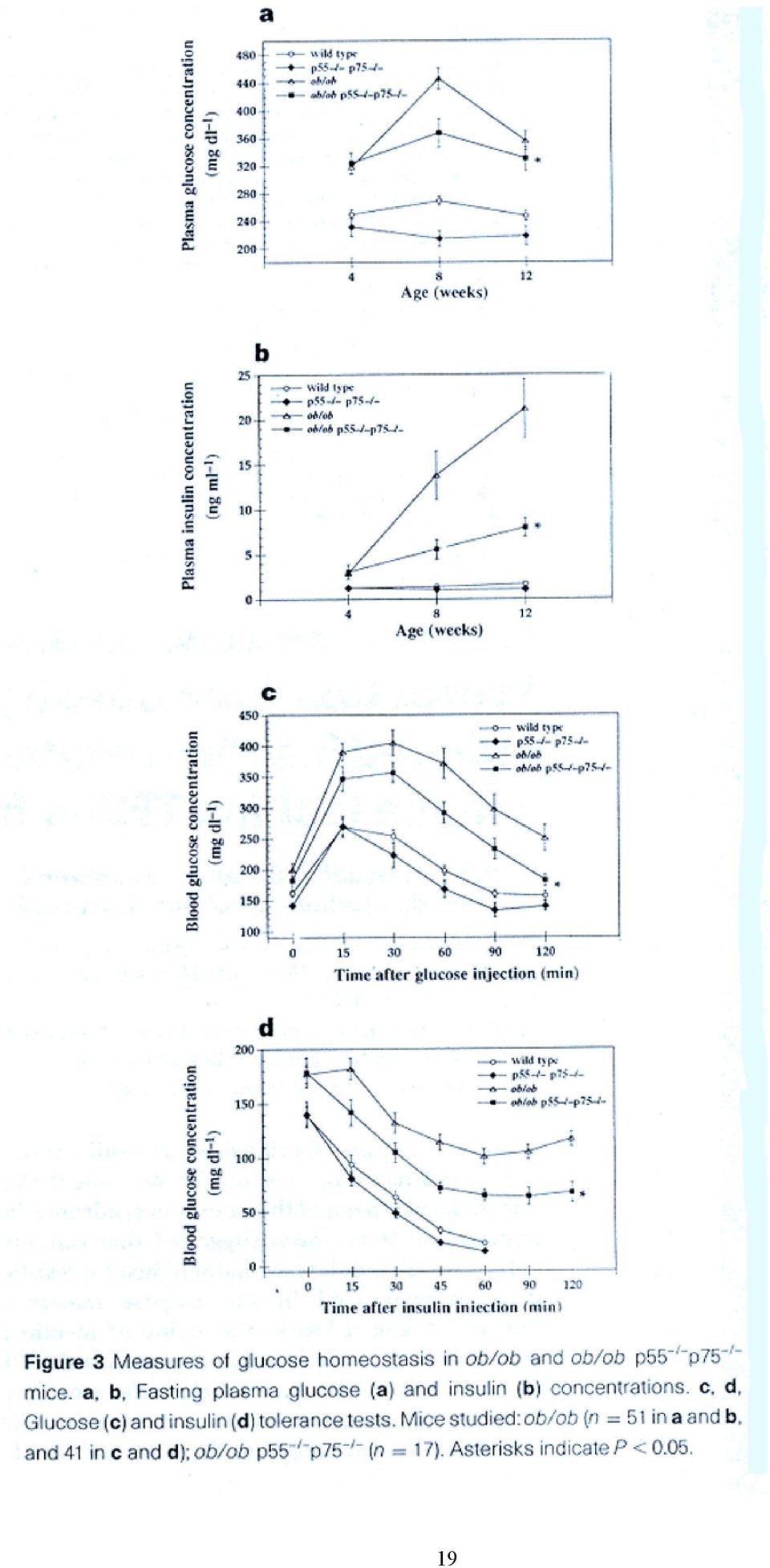
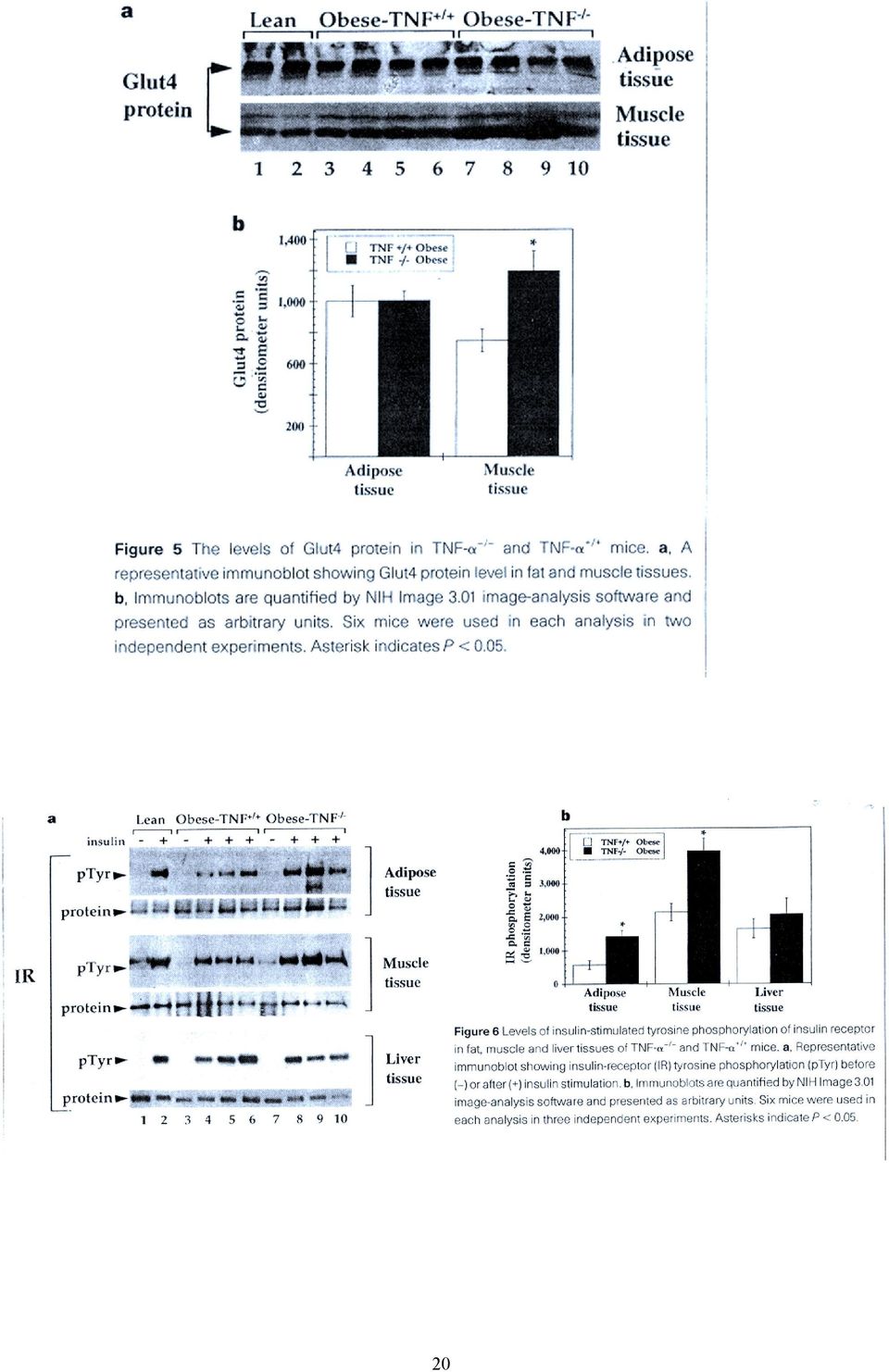
6 Háttérinformáció: A glikogén szintáz defoszforilált állapotban aktív, Ser-foszforilációval inaktiválódik. Inzulin gátolja az enzim foszforilációjában szerepet játszó kinázokat, a camp dependens protein kinázt, és a glikogén szintáz kináz-3-t (GSK-3). Ugyanekkor PI3K-dependens módon aktiválja a foszfoprotein foszfatáz-1 glikogén részecskéknél lokalizálódó poolját. Az inzulin hatására bekövetkező GSK-3 inaktiváció összefügg-e a MAP kináz kaszkáddal, vagy a PI3K-zal? Mi a protein kináz B szerepe? 4, Az elhízás és az inzulin rezisztencia közötti molekuláris kapcsolat pontosan nem ismert, de több mechanizmust is leírtak az utóbbi években (lsd. összefoglaló cikket), ezek közül példaként javasolt az alábbi cikk áttekintése: Uysal KT, et al. Protection from obesity-induced insulin resistance in mice lacking TNF-α function. Nature 1997, 389: Milyen modelleket használtak a cikkben a kövérség és az inzulin rezisztencia közötti kapcsolat kimutatására? A TNF-α hiánya hogyan befolyásolja ezt a kapcsolatot? 6

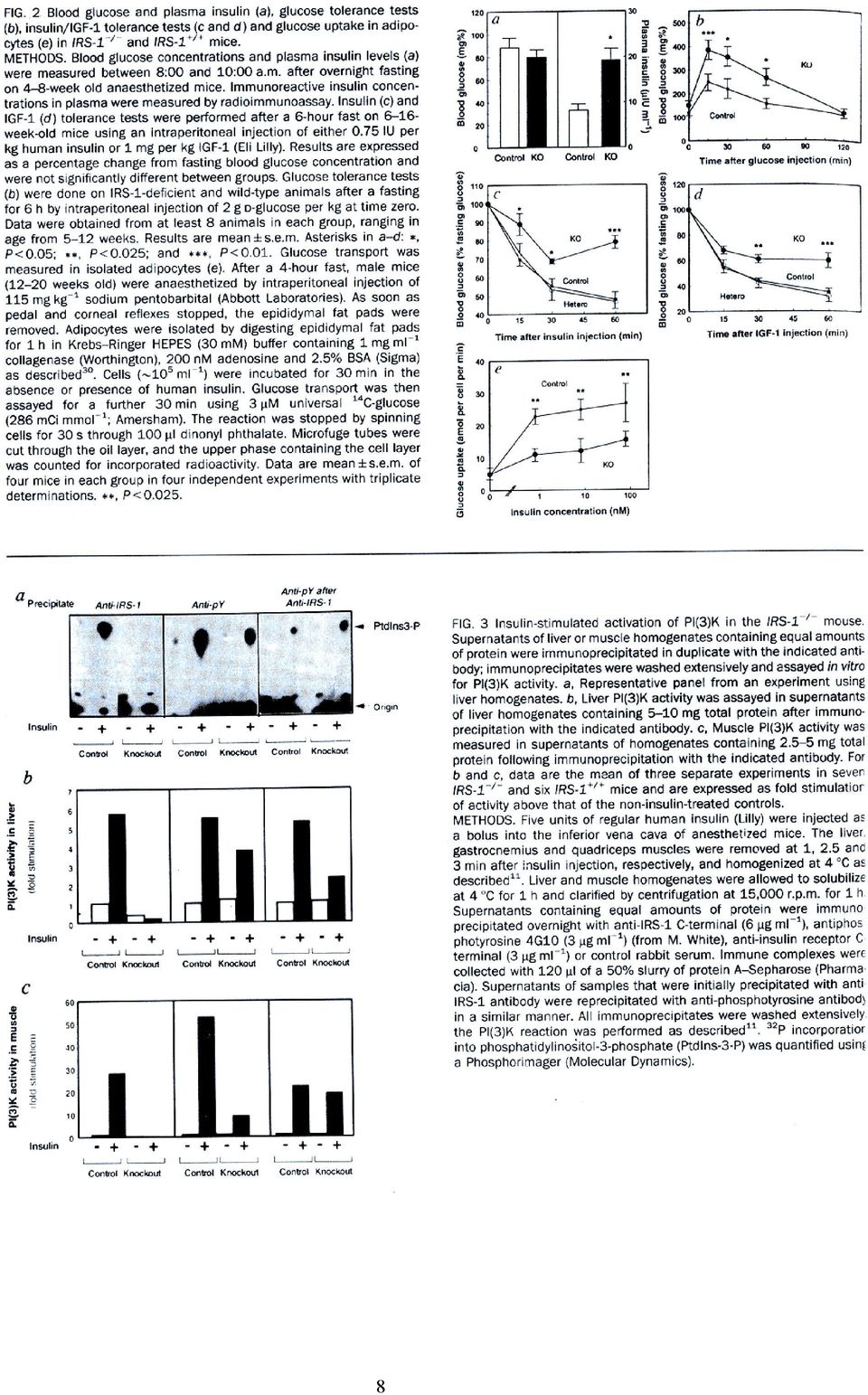
7 Ábramelléklet Araki E, et al. Alternative pathway of insulin signalling in mice with targeted disruption of the IRS-1 gene. Nature 1994, 372: , Figs 1f, 2-4. Figure 1f. Growth curves of IRS-1 + / + (control), heterozygous (Hetero) and homozygous IRS-1 - / - mutant (knock out, KO) mice. Data are from two litters and are representative of 60 animals. Values are mean ±SEM of three IRS-1 + / +, four IRS-1 + / -, and four IRS-1 - / - animals. 7
, heterozygous (Hetero) and homozygous IRS-1 - / - mutant (knock out, KO)")
8 8
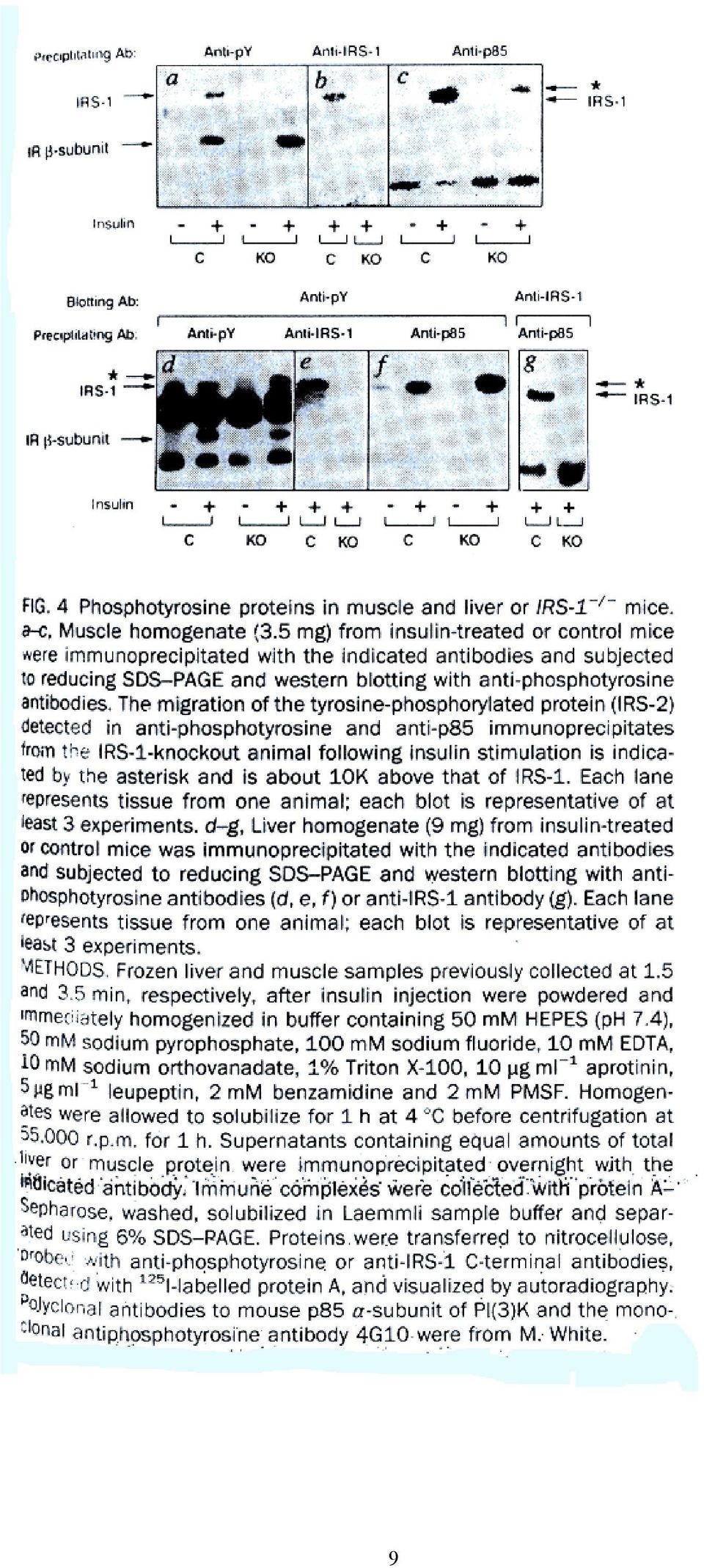
9 9
10 Withers DJ, et al. Disruption of IRS-2 causes type 2 diabetes in mice. Nature 1998, 391: , Figs 1e, 2-4. Figure 1f. Growth curves of wild type (WT), heterozygous (IRS-2 + / - ) and homozygous IRS-2 - / - mutant (IRS-2 - / - ) mice. Data are from six litters with a total of at least 15 animals per genotype. Representative of 60 animals. Values are mean ±SEM. 10
 mice.")
11 11
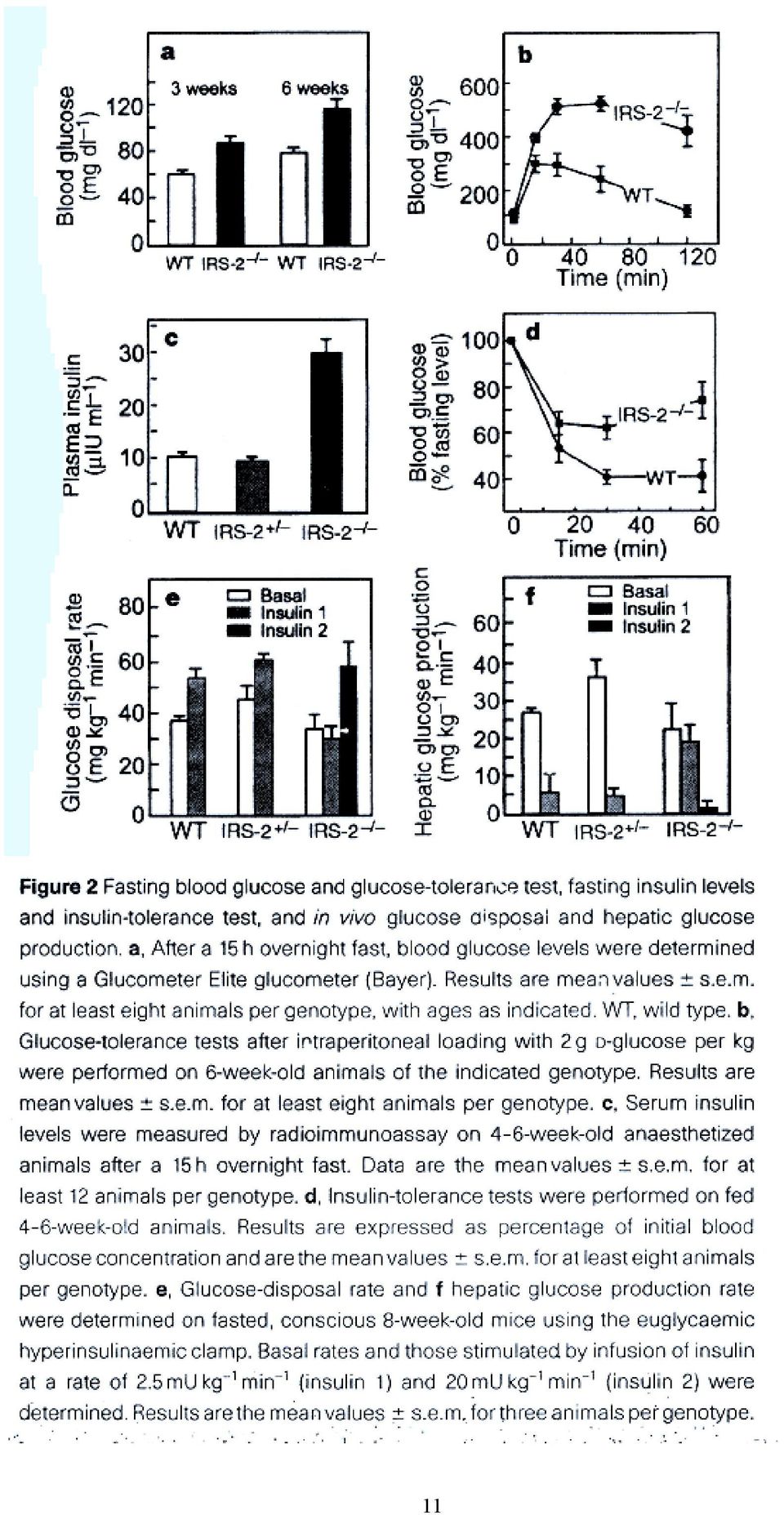
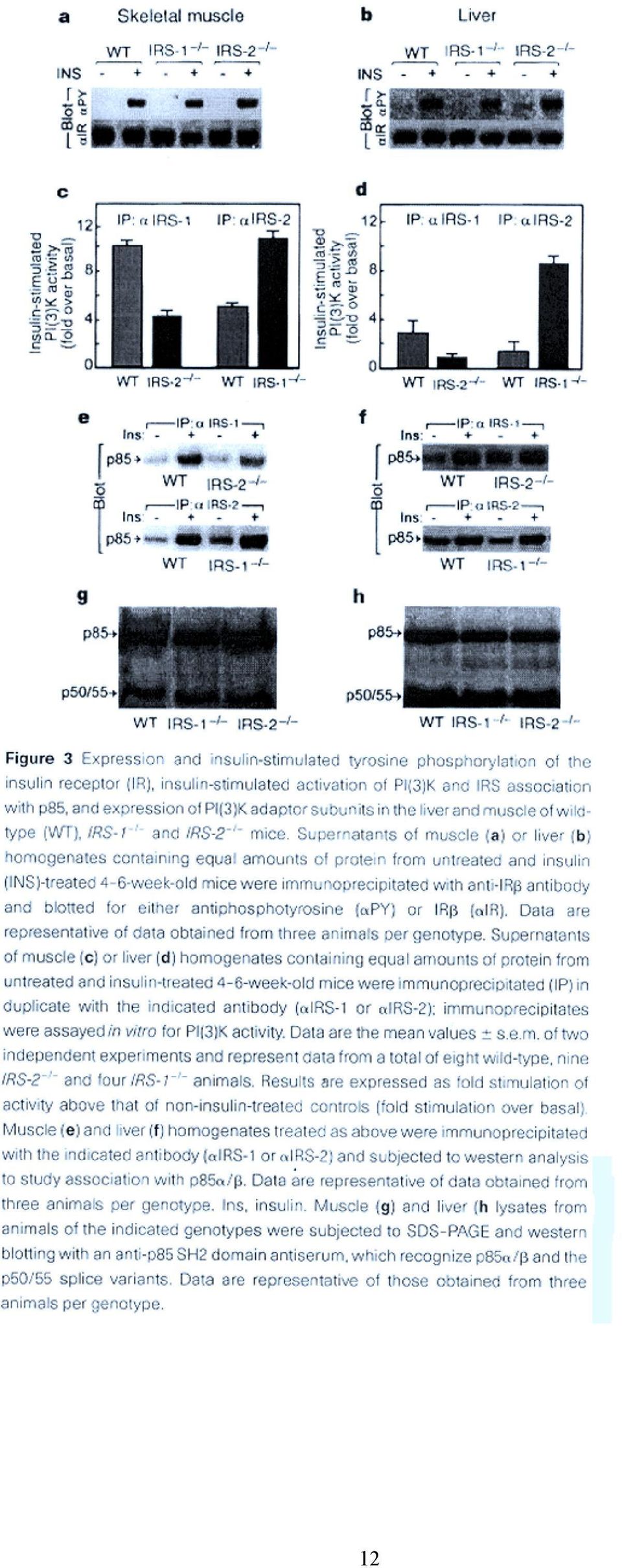
12 12
13 Figure 4. Quantification of β-cell mass by point-counting morphometric analysis on sections from the same mice analyzed blind for the genotype. Mean total pancreatic weights (g) ± SEM for each genotype were : wild type (WT) 178 ± 19, IRS-2 - / ± 12, and IRS-1 - / ± 9 (n=4). 13
 ± SEM for each genotype were : wild")
14 Cross DAE, et al. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378: , Figs 1, 2, 3a. 14

15 15
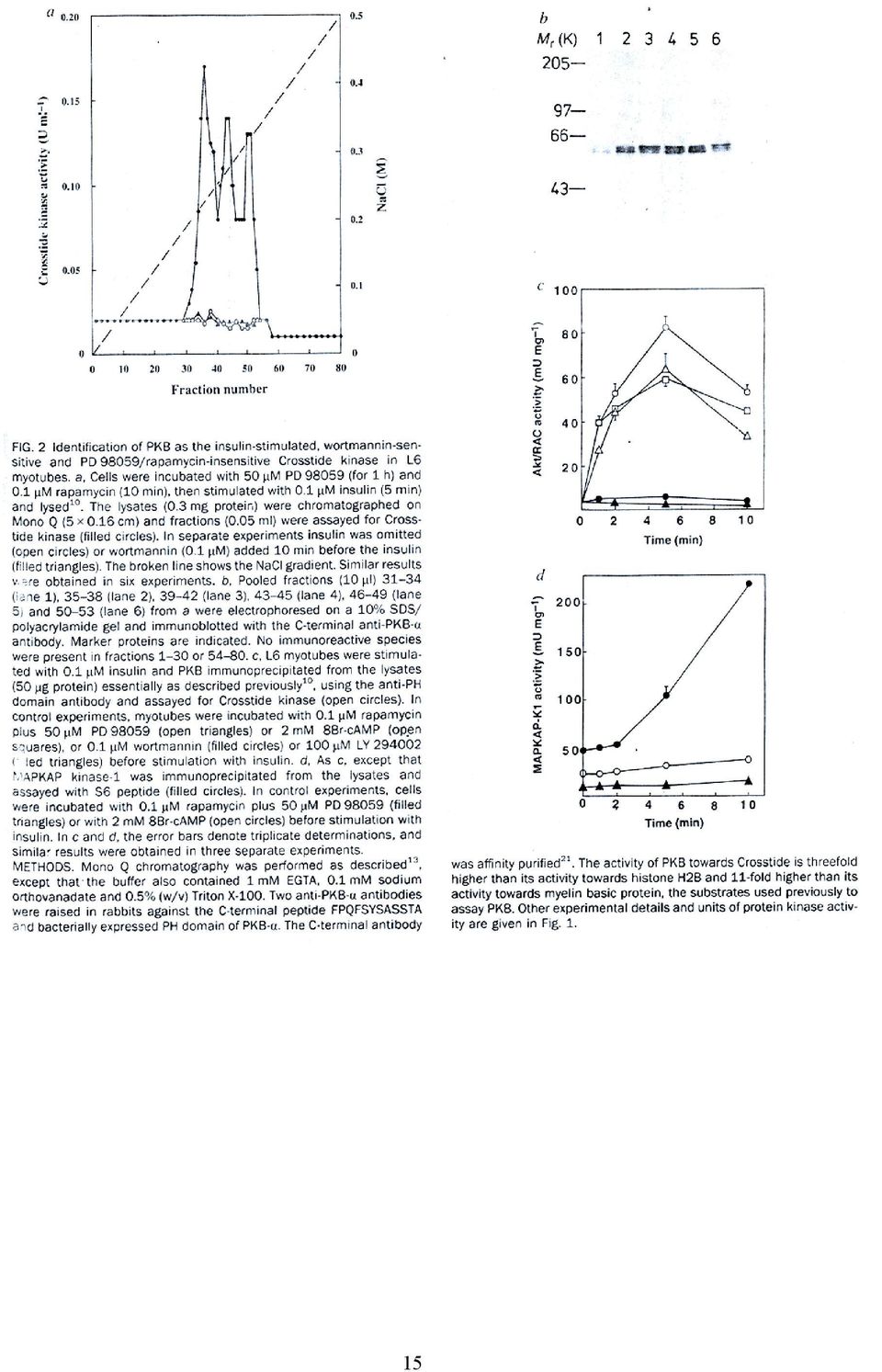
16 Figure 3a. GSK-3 is inactivated by PKB from insulin-stimulated L6 myotubes. Cells were stimulated for 5 min with 0.1 µm insulin, and PKB immunoprecipitated from 100 µg of cell lysate and used to inactivate GSK-3 isoforms. Black bars show GSK-3 activity measured after incubation with PKB as a percentage of the activity obtained in control incubations where PKB was omitted. In the absence of PKB, GSK-3 activity was stable throughout the experiment. The white bars show the activity after reactivation of GSK-3 with PP2A 1. No inactivation of GSK-3 occurred if insulin was omitted, or if wortmannin (0.1µM) was added 10 min before the insulin or if the anti- PKB antibody was incubated with peptide immunogen (0.5 mm) before immunoprecipitation. The results (± SEM) are for three experiments (each carried out in triplicate). 16

17 Uysal KT, et al. Protection from obesity-induced insulin resistance in mice lacking TNF-α function. Nature 1997, 389:

18 18
19 19
20 20
Az inzulin receptor szignál transzdukciója
 Az inzulin receptor szignál transzdukciója A szeminárium célja, hogy kísérleti adatok segítségével áttekintse az inzulin szekrécióját, az inzulin receptor szignál transzdukcióját, hatásait, és ezeknek
Az inzulin receptor szignál transzdukciója A szeminárium célja, hogy kísérleti adatok segítségével áttekintse az inzulin szekrécióját, az inzulin receptor szignál transzdukcióját, hatásait, és ezeknek
Receptor Tyrosine-Kinases
 Receptor Tyrosine-Kinases MAPkinase pathway PI3Kinase Protein Kinase B pathway PI3K/PK-B pathway Phosphatidyl-inositol-bisphosphate...(PI(4,5)P 2...) Phosphatidyl-inositol-3-kinase (PI3K) Protein kinase
Receptor Tyrosine-Kinases MAPkinase pathway PI3Kinase Protein Kinase B pathway PI3K/PK-B pathway Phosphatidyl-inositol-bisphosphate...(PI(4,5)P 2...) Phosphatidyl-inositol-3-kinase (PI3K) Protein kinase
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
A szövetek tápanyagellátásának hormonális szabályozása
 A szövetek tápanyagellátásának hormonális szabályozása Periódikus táplálékfelvétel Sejtek folyamatos tápanyagellátása (glükóz, szabad zsírsavak stb.) Tápanyag raktározás Tápanyag mobilizálás Vér glükóz
A szövetek tápanyagellátásának hormonális szabályozása Periódikus táplálékfelvétel Sejtek folyamatos tápanyagellátása (glükóz, szabad zsírsavak stb.) Tápanyag raktározás Tápanyag mobilizálás Vér glükóz
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
mi a cukorbetegség? DR. TSCHÜRTZ NÁNDOR, DR. HIDVÉGI TIBOR
 mi a cukorbetegség? DR. TSCHÜRTZ NÁNDOR, DR. HIDVÉGI TIBOR az OkTaTÓaNyag a magyar DiabeTes Társaság vezetôsége megbízásából, a sanofi TámOgaTásával készült készítette a magyar DiabeTes Társaság edukációs
mi a cukorbetegség? DR. TSCHÜRTZ NÁNDOR, DR. HIDVÉGI TIBOR az OkTaTÓaNyag a magyar DiabeTes Társaság vezetôsége megbízásából, a sanofi TámOgaTásával készült készítette a magyar DiabeTes Társaság edukációs
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Diabetes mellitus. Rácz Olivér Miskolci Egyetem Egészségügyi Kar. Diabetes mellitus
 Diabetes mellitus Rácz Olivér Miskolci Egyetem Egészségügyi Kar 2009 diamisk.ppt 1 Diabetes mellitus Glukóz homeosztázis, inzulin, inzulin antagonisták Definíció és a probléma méretei A diabetes tünetei
Diabetes mellitus Rácz Olivér Miskolci Egyetem Egészségügyi Kar 2009 diamisk.ppt 1 Diabetes mellitus Glukóz homeosztázis, inzulin, inzulin antagonisták Definíció és a probléma méretei A diabetes tünetei
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Dr.Varga Annamária belgyógyász, diabetológus
 Dr.Varga Annamária belgyógyász, diabetológus A szénhidrát-, a zsír-, a fehérje és a nukleinsav-anyagcsere komplex zavara Oka: a hasnyálmirigyben termelődő - inzulin hiánya - a szervezet inzulinnal szembeni
Dr.Varga Annamária belgyógyász, diabetológus A szénhidrát-, a zsír-, a fehérje és a nukleinsav-anyagcsere komplex zavara Oka: a hasnyálmirigyben termelődő - inzulin hiánya - a szervezet inzulinnal szembeni
Jelátviteli útvonalak 2
 Jelátviteli útvonalak 2 Információ metabolizmus Szignál transzdukció GPCR: PLC és foszfoinozitid kaszkád Szignál (pl. adrenalin) + receptor (pl. 1 -adrenerg) G q foszfolipáz-c (PLC) IP 3 (hidrofil) + DAG
Jelátviteli útvonalak 2 Információ metabolizmus Szignál transzdukció GPCR: PLC és foszfoinozitid kaszkád Szignál (pl. adrenalin) + receptor (pl. 1 -adrenerg) G q foszfolipáz-c (PLC) IP 3 (hidrofil) + DAG
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
3. Főbb Jelutak. 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3.
 Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
A cukorbetegség eiológiája és patogenezise
 A cukorbetegség eiológiája és patogenezise Dr. Pánczél Pál med. habil. egyetemi docens Semmelweis Egyetem III.sz. Belgyógyászati Klinika A normális pancreas és a Langerhans szigetek lobules intralobular
A cukorbetegség eiológiája és patogenezise Dr. Pánczél Pál med. habil. egyetemi docens Semmelweis Egyetem III.sz. Belgyógyászati Klinika A normális pancreas és a Langerhans szigetek lobules intralobular
A diabetes mellitus laboratóriumi diagnosztikája
 A diabetes mellitus laboratóriumi diagnosztikája Laborvizsgálatok célja diabetes mellitusban 1. Diagnózis 2. Monitorozás 3. Metabolikus komplikációk kimutatása és követése Laboratóriumi tesztek a diabetes
A diabetes mellitus laboratóriumi diagnosztikája Laborvizsgálatok célja diabetes mellitusban 1. Diagnózis 2. Monitorozás 3. Metabolikus komplikációk kimutatása és követése Laboratóriumi tesztek a diabetes
Jelátviteli útvonalak 1
 Jelátviteli útvonalak 1 Információ metabolizmus Szignál transzdukció 1 Jelátviteli séma Mi lehet a jel? Hormonok Növekedési faktorok Fejlődési szignálok Neurotranszmitterek Antigének Sejtfelszíni glikoproteinek
Jelátviteli útvonalak 1 Információ metabolizmus Szignál transzdukció 1 Jelátviteli séma Mi lehet a jel? Hormonok Növekedési faktorok Fejlődési szignálok Neurotranszmitterek Antigének Sejtfelszíni glikoproteinek
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
A metabolikus szindróma genetikai háttere. Kappelmayer János, Balogh István (www.kbmpi.hu)
") A metabolikus szindróma genetikai háttere Kappelmayer János, Balogh István (www.kbmpi.hu) Definíció WHO, 1999 EGIR, 1999 ATP III, 2001 Ha három vagy több komponens jelen van a betegben: Vérnyomás: > 135/85
A metabolikus szindróma genetikai háttere Kappelmayer János, Balogh István (www.kbmpi.hu) Definíció WHO, 1999 EGIR, 1999 ATP III, 2001 Ha három vagy több komponens jelen van a betegben: Vérnyomás: > 135/85
MIKROBIOM ÉS ELHÍZÁS HEINZ GYAKY 2018 BUDAPEST
 MIKROBIOM ÉS ELHÍZÁS HEINZ GYAKY 2018 BUDAPEST HUMÁN MIKROBIOM 10 billió baktérium alkotja a 1,5-2 kg súlyú humán mikrobiomot. Elsősorban az emberi bélben található A bél által szabályozott bélflóra a
MIKROBIOM ÉS ELHÍZÁS HEINZ GYAKY 2018 BUDAPEST HUMÁN MIKROBIOM 10 billió baktérium alkotja a 1,5-2 kg súlyú humán mikrobiomot. Elsősorban az emberi bélben található A bél által szabályozott bélflóra a
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterkézés megfeleltetése az Euróai Unió új társadalmi kihívásainak a écsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMO-4.1.2-08/1/A-2009-0011 Az orvosi biotechnológiai
Az orvosi biotechnológiai mesterkézés megfeleltetése az Euróai Unió új társadalmi kihívásainak a écsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMO-4.1.2-08/1/A-2009-0011 Az orvosi biotechnológiai
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
B-sejtek szerepe az RA patológiás folyamataiban
 B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Endokrinológia. Közös jellemzők: nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások. váladékuk a hormon
 Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
TÁPLÁLKOZÁSI ZAVAROK, ELHÍZÁS. 9.10.2009 nutrmisk.ppt 1
 TÁPLÁLKOZÁSI ZAVAROK, ELHÍZÁS Rácz Olivér Miskolci Egyetem Egészségügyi Kar 9.10.2009 nutrmisk.ppt 1 GLOBALIZÁCIÓ Alultápláltság, malnutríció - a világ 1/3-a Elhízás a világ másik 1/3-a (Globezitás?) A
TÁPLÁLKOZÁSI ZAVAROK, ELHÍZÁS Rácz Olivér Miskolci Egyetem Egészségügyi Kar 9.10.2009 nutrmisk.ppt 1 GLOBALIZÁCIÓ Alultápláltság, malnutríció - a világ 1/3-a Elhízás a világ másik 1/3-a (Globezitás?) A
MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje A szénhidrátok a szervezet számára fontos, alapvető tápanyagok. Az emberi szervezetben
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje A szénhidrátok a szervezet számára fontos, alapvető tápanyagok. Az emberi szervezetben
Új terápiás lehetőségek (receptorok) a kardiológiában
 a kardiológiában") Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
A KOLESZTERIN SZERKEZETE. (koleszterin v. koleszterol)
") 19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Cukorbetegek hypertoniájának korszerű kezelése. Dr. Balogh Sándor OALI Főigazgató főorvos Budapest
 Cukorbetegek hypertoniájának korszerű kezelése Dr. Balogh Sándor OALI Főigazgató főorvos Budapest Hypertonia diabetesben 1-es típusú diabetes 2-es típusú diabetes Nephropathia diabetica albuminuria (intermittáló
Cukorbetegek hypertoniájának korszerű kezelése Dr. Balogh Sándor OALI Főigazgató főorvos Budapest Hypertonia diabetesben 1-es típusú diabetes 2-es típusú diabetes Nephropathia diabetica albuminuria (intermittáló
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL
 Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Vércukorszint szabályozás
 Vércukorszint szabályozás Raktározás: Szénhidrátok: glikogén formájában (máj, izom) Zsírok: zsírsejtek zsírszövet Fehérje: bőr alatti lazarostos kötőszövet Szénhidrát metabolizmus Szénhidrátok a bélben
Vércukorszint szabályozás Raktározás: Szénhidrátok: glikogén formájában (máj, izom) Zsírok: zsírsejtek zsírszövet Fehérje: bőr alatti lazarostos kötőszövet Szénhidrát metabolizmus Szénhidrátok a bélben
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Supplementary Table 1. Cystometric parameters in sham-operated wild type and Trpv4 -/- rats during saline infusion and
 WT sham Trpv4 -/- sham Saline 10µM GSK1016709A P value Saline 10µM GSK1016709A P value Number 10 10 8 8 Intercontractile interval (sec) 143 (102 155) 98.4 (71.4 148) 0.01 96 (92 121) 109 (95 123) 0.3 Voided
WT sham Trpv4 -/- sham Saline 10µM GSK1016709A P value Saline 10µM GSK1016709A P value Number 10 10 8 8 Intercontractile interval (sec) 143 (102 155) 98.4 (71.4 148) 0.01 96 (92 121) 109 (95 123) 0.3 Voided
Premium Health Concepts A módszer tudományos alapjai
 Premium Health Concepts A módszer tudományos alapjai A testtömeg szabályozása nagyon bonyolult, egymásra, és saját koncentrációjukra is ható hormonok valamint az idegrendszer hatásainak összessége. Számos
Premium Health Concepts A módszer tudományos alapjai A testtömeg szabályozása nagyon bonyolult, egymásra, és saját koncentrációjukra is ható hormonok valamint az idegrendszer hatásainak összessége. Számos
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Receptorok, szignáltranszdukció jelátviteli mechanizmusok
 Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Kutatási beszámoló ( )
") Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
A pszichológiai tényezők szerepe az elhízásban
 A pszichológiai tényezők szerepe az elhízásban Tisljár-Szabó Eszter eszter.szabo@sph.unideb.hu Magatartástudományi Intézet Mi az elhízás? Sokféleképpen definiálják: a. Populáció átlagsúlyhoz képest: >20-40%:
A pszichológiai tényezők szerepe az elhízásban Tisljár-Szabó Eszter eszter.szabo@sph.unideb.hu Magatartástudományi Intézet Mi az elhízás? Sokféleképpen definiálják: a. Populáció átlagsúlyhoz képest: >20-40%:
A neuroendokrin jelátviteli rendszer
 A neuroendokrin jelátviteli rendszer Hipotalamusz Hipofízis Pajzsmirigy Mellékpajzsmirigy Zsírszövet Mellékvese Hasnyálmirigy Vese Petefészek Here Hormon felszabadulási kaszkád Félelem Fertőzés Vérzés
A neuroendokrin jelátviteli rendszer Hipotalamusz Hipofízis Pajzsmirigy Mellékpajzsmirigy Zsírszövet Mellékvese Hasnyálmirigy Vese Petefészek Here Hormon felszabadulási kaszkád Félelem Fertőzés Vérzés
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
A piruvát-dehidrogenáz komplex. Csala Miklós
 A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
klorid ioncsatorna az ABC (ATP Binding Casette) fehérjecsaládba tartozik, amelyek általánosságban részt vesznek a gyógyszerek olyan alapvetı
 fehérjecsaládba tartozik, amelyek általánosságban részt vesznek a gyógyszerek olyan alapvetı") Szegedi Tudományegyetem Farmakológiai és Farmakoterápiai Intézet Igazgató: Prof. Dr. Varró András 6720 Szeged, Dóm tér 12., Tel.: (62) 545-682 Fax: (62) 545-680 e-mail: varro.andras@med.u-szeged.hu Opponensi
Szegedi Tudományegyetem Farmakológiai és Farmakoterápiai Intézet Igazgató: Prof. Dr. Varró András 6720 Szeged, Dóm tér 12., Tel.: (62) 545-682 Fax: (62) 545-680 e-mail: varro.andras@med.u-szeged.hu Opponensi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Szénhidrát anyagcsere. Kőszegi Tamás, Lakatos Ágnes PTE Laboratóriumi Medicina Intézet
 Szénhidrát anyagcsere Kőszegi Tamás, Lakatos Ágnes PTE Laboratóriumi Medicina Intézet Szénhidrát anyagcsere sommásan Izomszövet Zsírszövet Máj Homeosztázis Hormon Hatás Szerv Inzulin Glukagon Sejtek glükóz
Szénhidrát anyagcsere Kőszegi Tamás, Lakatos Ágnes PTE Laboratóriumi Medicina Intézet Szénhidrát anyagcsere sommásan Izomszövet Zsírszövet Máj Homeosztázis Hormon Hatás Szerv Inzulin Glukagon Sejtek glükóz
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
Lipidek anyagcseréje és az ateroszklerózis (érelmeszesedés)
") Lipidek anyagcseréje és az ateroszklerózis (érelmeszesedés) Rácz Olivér Miskolci Egyetem Egészségügyi Kar 22.9.2009 ateromisk.ppt 1 Az érelmeszesedés csak a XIX. évszázad második felétől orvosi probléma
Lipidek anyagcseréje és az ateroszklerózis (érelmeszesedés) Rácz Olivér Miskolci Egyetem Egészségügyi Kar 22.9.2009 ateromisk.ppt 1 Az érelmeszesedés csak a XIX. évszázad második felétől orvosi probléma
Az AT 1A -angiotenzinreceptor G-fehérjétől független jelátvitelének vizsgálata C9 sejtekben. Doktori tézisek. Dr. Szidonya László
 Az AT 1A -angiotenzinreceptor G-fehérjétől független jelátvitelének vizsgálata C9 sejtekben Doktori tézisek Dr. Szidonya László Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető:
Az AT 1A -angiotenzinreceptor G-fehérjétől független jelátvitelének vizsgálata C9 sejtekben Doktori tézisek Dr. Szidonya László Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető:
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-8/1/A-29-11 Az orvosi biotechnológiai
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-8/1/A-29-11 Az orvosi biotechnológiai
A szénhidrátok anyagcseréje. SZTE AOK Biokémiai Intézet Gyógyszerész hallgatók számára 2014.
 A szénhidrátok anyagcseréje SZTE AOK Biokémiai Intézet Gyógyszerész hallgatók számára 2014. A szénhidrátok emésztése és felszívódása Táplálkozás: növényi keményítő, szacharóz, laktóz (tej, tejtermékek)
A szénhidrátok anyagcseréje SZTE AOK Biokémiai Intézet Gyógyszerész hallgatók számára 2014. A szénhidrátok emésztése és felszívódása Táplálkozás: növényi keményítő, szacharóz, laktóz (tej, tejtermékek)
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai
 Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
A krónikus veseelégtelenség kardio-metabolikus kockázata
 A krónikus veseelégtelenség kardio-metabolikus kockázata Dr. Kiss István Fıv. Önk. Szent Imre Kórház, Belgyógyászati Mátrix Intézet, Nephrologia-Hypertonia Profil 1 2 SZÍV ÉS ÉRRENDSZERI KOCKÁZAT ÉS BETEGSÉG
A krónikus veseelégtelenség kardio-metabolikus kockázata Dr. Kiss István Fıv. Önk. Szent Imre Kórház, Belgyógyászati Mátrix Intézet, Nephrologia-Hypertonia Profil 1 2 SZÍV ÉS ÉRRENDSZERI KOCKÁZAT ÉS BETEGSÉG
Homeosztázis A szervezet folyadékterei
 Homeosztázis A szervezet folyadékterei Homeosztázis Homeosztázis: a folytonos változások mellett az organizáció állandóságát létrehozó biológiai jelenség. A belső környezet szabályozott stabilitása. Megengedett
Homeosztázis A szervezet folyadékterei Homeosztázis Homeosztázis: a folytonos változások mellett az organizáció állandóságát létrehozó biológiai jelenség. A belső környezet szabályozott stabilitása. Megengedett
Többgénes jellegek. 1. Klasszikus (poligénes) mennyiségi jellegek. 2.Szinte minden jelleg több gén irányítása alatt áll
 mennyiségi jellegek. 2.Szinte minden jelleg több gén irányítása alatt áll") Többgénes jellegek Többgénes jellegek 1. 1. Klasszikus (poligénes) mennyiségi jellegek Multifaktoriális jellegek: több gén és a környezet által meghatározott jellegek 2.Szinte minden jelleg több gén irányítása
Többgénes jellegek Többgénes jellegek 1. 1. Klasszikus (poligénes) mennyiségi jellegek Multifaktoriális jellegek: több gén és a környezet által meghatározott jellegek 2.Szinte minden jelleg több gén irányítása
16. A sejtek kommunikációja: jelátviteli folyamatok (szignál-transzdukció)
") 16. A sejtek kommunikációja: jelátviteli folyamatok (szignál-transzdukció) 2016. február 25. Lippai Mónika lippai@elte.hu Minden sejt érzékel többféle, más sejtek által kibocsájtott jelmolekulát. - A jeleket
16. A sejtek kommunikációja: jelátviteli folyamatok (szignál-transzdukció) 2016. február 25. Lippai Mónika lippai@elte.hu Minden sejt érzékel többféle, más sejtek által kibocsájtott jelmolekulát. - A jeleket
A pajzsmirigy. A pajzsmirigyhormonok
 A pajzsmirigy A pajzsmirigyhormonok a pajzsmirigy mintegy 20 g súlyú páros szerv tireociták follikulusok körül; pajzsmirigykolloid (tireoglobulin glikoprotein) tárolás a pajzsmirigy 2 aktív hormont termel:
A pajzsmirigy A pajzsmirigyhormonok a pajzsmirigy mintegy 20 g súlyú páros szerv tireociták follikulusok körül; pajzsmirigykolloid (tireoglobulin glikoprotein) tárolás a pajzsmirigy 2 aktív hormont termel:
A D-vitamin anyagcsere hatásai ECH Molnár Gergő Attila. PTE KK, II.sz. Belgyógyászati Klinika és NC. memphiscashsaver.com
 A D-vitamin anyagcsere hatásai memphiscashsaver.com Molnár Gergő Attila PTE KK, II.sz. Belgyógyászati Klinika és NC A D-vitamin képződése és sokrétű hatása http://www.insanemedicine.com/ D-vitamin és szénhidrát-anyagcsere
A D-vitamin anyagcsere hatásai memphiscashsaver.com Molnár Gergő Attila PTE KK, II.sz. Belgyógyászati Klinika és NC A D-vitamin képződése és sokrétű hatása http://www.insanemedicine.com/ D-vitamin és szénhidrát-anyagcsere
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
1-es típusú diabetes mellitus
 Diabetológia nem diabetológusoknak 1-es típusú diabetes mellitus Tabák Gy. Ádám Az endokrinológia modern gyakorlata Nem csak endokrinológusoknak Budapest, 2015. március 14. 1-es típusú diabetes mellitus
Diabetológia nem diabetológusoknak 1-es típusú diabetes mellitus Tabák Gy. Ádám Az endokrinológia modern gyakorlata Nem csak endokrinológusoknak Budapest, 2015. március 14. 1-es típusú diabetes mellitus
A HIPERINZULINÉMIA SZEREPE ELHÍZÁSTÓL FÜGGETLEN POLICISZTÁS OVÁRIUM SZINDRÓMÁBAN
 A HIPERINZULINÉMIA SZEREPE ELHÍZÁSTÓL FÜGGETLEN POLICISZTÁS OVÁRIUM SZINDRÓMÁBAN Csenteri Orsolya Karola, Dr. Gődény Sándor Debreceni Egyetem Népegészségügyi Kar Megelőző Orvostani Intézet BEVEZETÉS PCOS
A HIPERINZULINÉMIA SZEREPE ELHÍZÁSTÓL FÜGGETLEN POLICISZTÁS OVÁRIUM SZINDRÓMÁBAN Csenteri Orsolya Karola, Dr. Gődény Sándor Debreceni Egyetem Népegészségügyi Kar Megelőző Orvostani Intézet BEVEZETÉS PCOS
Lipotoxicitás ellen védő hatóanyagok vizsgálata inzulinóma sejteken
 Lipotoxicitás ellen védő hatóanyagok vizsgálata inzulinóma sejteken Doktori értekezés Simon-Szabó Laura Semmelweis Egyetem Gyógyszertudományok Doktori Iskola Témavezetők: prof. Kéri György, DSc., kutatóprofesszor
Lipotoxicitás ellen védő hatóanyagok vizsgálata inzulinóma sejteken Doktori értekezés Simon-Szabó Laura Semmelweis Egyetem Gyógyszertudományok Doktori Iskola Témavezetők: prof. Kéri György, DSc., kutatóprofesszor
LIPID ANYAGCSERE (2011)
") LIPID ANYAGCSERE LIPID ANYAGCSERE (2011) 5 ELİADÁS: 1, ZSÍRK EMÉSZTÉSE, FELSZÍVÓDÁSA + LIPPRTEINEK 2, ZSÍRSAVAK XIDÁCIÓJA 3, ZSÍRSAVAK SZINTÉZISE 4, KETNTESTEK BIKÉMIÁJA, KLESZTERIN ANYAGCSERE 5, MEMBRÁN
LIPID ANYAGCSERE LIPID ANYAGCSERE (2011) 5 ELİADÁS: 1, ZSÍRK EMÉSZTÉSE, FELSZÍVÓDÁSA + LIPPRTEINEK 2, ZSÍRSAVAK XIDÁCIÓJA 3, ZSÍRSAVAK SZINTÉZISE 4, KETNTESTEK BIKÉMIÁJA, KLESZTERIN ANYAGCSERE 5, MEMBRÁN
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika
 Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Glikolízis. Csala Miklós
 Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Zsírsav szintézis. Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P. 2 i
 Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
TÁJÉKOZTATÓ lobbitevékenységről a 2006. évi XLIX. törvény 30. alapján
 TÁJÉKOZTATÓ lobbitevékenységről a 2006. évi XLIX. törvény 30. alapján Lobbista neve: VAJDA ANDRÁS Lobbiigazolvány sorszáma: 07325 A tájékoztató időszaka: 2009. III. negyedév 1. A lobbitevékenységgel érintett
TÁJÉKOZTATÓ lobbitevékenységről a 2006. évi XLIX. törvény 30. alapján Lobbista neve: VAJDA ANDRÁS Lobbiigazolvány sorszáma: 07325 A tájékoztató időszaka: 2009. III. negyedév 1. A lobbitevékenységgel érintett
1b. Fehérje transzport
 1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
8. előadás. Sejt-sejt kommunikáció és jelátvitel
 8. előadás Sejt-sejt kommunikáció és jelátvitel A sejt-sejt szignalizáció evolúciója A Saccharomyces cerevisiae (sörélesztő) élesztőnek két párosodási típusa van: a és α A különböző párosodási típusokba
8. előadás Sejt-sejt kommunikáció és jelátvitel A sejt-sejt szignalizáció evolúciója A Saccharomyces cerevisiae (sörélesztő) élesztőnek két párosodási típusa van: a és α A különböző párosodási típusokba
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
PhD értekezés tézisei DIFFERENCIÁCIÓS ÉS TÚLÉLÉSI JELÁTVITEL PATKÁNY PHEOCHROMOCYTOMA SEJTEKBEN
 PhD értekezés tézisei DIFFERENCIÁCIÓS ÉS TÚLÉLÉSI JELÁTVITEL PATKÁNY PHEOCHROMOCYTOMA SEJTEKBEN DR. PAP MARIANNA TÉMAVEZETŐ: PROF. DR. SZEBERÉNYI JÓZSEF PÉCSI TUDOMÁNYEGYETEM ÁLTALÁNOS ORVOSTUDOMÁNYI KAR
PhD értekezés tézisei DIFFERENCIÁCIÓS ÉS TÚLÉLÉSI JELÁTVITEL PATKÁNY PHEOCHROMOCYTOMA SEJTEKBEN DR. PAP MARIANNA TÉMAVEZETŐ: PROF. DR. SZEBERÉNYI JÓZSEF PÉCSI TUDOMÁNYEGYETEM ÁLTALÁNOS ORVOSTUDOMÁNYI KAR
Szerkesztette: dr Lázár Sarnyai Nóra
 Szerkesztette: dr Lázár Sarnyai Nóra A cukorbetegség (diabetes mellitus) a szénhidrátanyagcsere zavara, amely a vérben, illetve a testnedvekben megemelkedett cukorszinttel és kóros anyagcseretermékekkel
Szerkesztette: dr Lázár Sarnyai Nóra A cukorbetegség (diabetes mellitus) a szénhidrátanyagcsere zavara, amely a vérben, illetve a testnedvekben megemelkedett cukorszinttel és kóros anyagcseretermékekkel
ISMÉTLŐDŐ STRESSZ HATÁSAINAK VIZSGÁLATA ZEBRADÁNIÓN (DANIO RERIO)
") ISMÉTLŐDŐ STRESSZ HATÁSAINAK VIZSGÁLATA ZEBRADÁNIÓN (DANIO RERIO) Buza Eszter, Váradi László, Csenki Zsolt, Müller Tamás, Jeney Zsigmond, Mézes Miklós Szent István Egyetem Mezőgazdaság-és Környezettudományi
ISMÉTLŐDŐ STRESSZ HATÁSAINAK VIZSGÁLATA ZEBRADÁNIÓN (DANIO RERIO) Buza Eszter, Váradi László, Csenki Zsolt, Müller Tamás, Jeney Zsigmond, Mézes Miklós Szent István Egyetem Mezőgazdaság-és Környezettudományi
Glucagon. A-sejtekben termelődik (+ GI és CNS L sejtjei) Egyláncú peptid, MW: 3,500; aa:29. preprohormon MW: 18,000 prohormon (glycentin) MW: 12,000
 Egyláncú peptid, MW: 3,500; aa:29. preprohormon MW: 18,000 prohormon (glycentin) MW: 12,000") Glucagon A-sejtekben termelődik (+ GI és CNS L sejtjei) Egyláncú peptid, MW: 3,500; aa:29 preprohormon MW: 18,000 prohormon (glycentin) MW: 12,000 Szignalizáció: camp (Gs) Additív hatás katekolaminokkal
Glucagon A-sejtekben termelődik (+ GI és CNS L sejtjei) Egyláncú peptid, MW: 3,500; aa:29 preprohormon MW: 18,000 prohormon (glycentin) MW: 12,000 Szignalizáció: camp (Gs) Additív hatás katekolaminokkal
1. Mi jellemző a connexin fehérjékre?
 Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Biokémiai és Molekuláris Biológiai Intézet. Lipid anyagcsere. Balajthy Zoltán, Sarang Zsolt
 Biokémiai és Molekuláris Biológiai Intézet Lipid anyagcsere Balajthy Zoltán, Sarang Zsolt Anabolikus és katabolikus folyamatok a szervezetben Lipidek osztályozása Lipidek szerepe a szervezetben Lipidek
Biokémiai és Molekuláris Biológiai Intézet Lipid anyagcsere Balajthy Zoltán, Sarang Zsolt Anabolikus és katabolikus folyamatok a szervezetben Lipidek osztályozása Lipidek szerepe a szervezetben Lipidek
Molekuláris genetikai vizsgáló. módszerek az immundefektusok. diagnosztikájában
 Molekuláris genetikai vizsgáló módszerek az immundefektusok diagnosztikájában Primer immundefektusok A primer immundeficiencia ritka, veleszületett, monogénes öröklődésű immunhiányos állapot. Családi halmozódást
Molekuláris genetikai vizsgáló módszerek az immundefektusok diagnosztikájában Primer immundefektusok A primer immundeficiencia ritka, veleszületett, monogénes öröklődésű immunhiányos állapot. Családi halmozódást