Biokémia II. Biokémiai szabályozás Szarka, András
|
|
|
- Flóra Anikó Horváth
- 8 évvel ezelőtt
- Látták:
Átírás
1 Biokémia II. Biokémiai szabályozás Szarka, András

2 Biokémia II.: Biokémiai szabályozás Szarka, András Szerzői jog 2014 Szarka András

3 Tartalom Biokémia II. - Biokémiai szabályozás... vii Előszó... viii 1. Bevezetés Biokatalizátorok enzimek Néhány szóban az enzimkinetikáról Az enzimes katalízis háttere Enzimszabályozás Allosztérikus enzimek Kooperativitás A hemoglobin szerkezete és oxigénkötése Fehérjék térszerkezetének, aktivitásának befolyásolása foszforiláció/defoszforiláció által Limitált proteolízis Inaktív zimogének és aktiválódásuk Pepszinogén Enterális proteázok zimogénjei Proteázinhibitorok Fehérjefolding, fehérjelebontás ubikvitin-proteaszóma rendszer Poszttranszlációs módosulások Fehérjefolding Hősokk-fehérjék molekuláris chaperonok Sérült fehérjék proteolízise Proteaszóma A proteaszóma felépítése A halál csókja Ubikvitináció Számos fehérje irányított lebontás révén szabályozott A génexpresszió szabályozása Génexpresszió, a gén kifejeződése Sejtdifferenciáció Azonosságok és különbségek a sejtek között A génexpresszió szabályozásának szintjei A génszabályozó fehérjék DNS-kötő részei DNS-felismerő fehérjeszerkezeti egységek Hélix-turn-hélix részletek DNS-kötő cink-ujj részletek β-redő Leucin-cipzár Hélix-loop-hélix részlet DNS-kötő fehérjék detektálása A genetikai kapcsolók munka közben A triptofán-represszor, mint egyszerű génkapcsoló A lac-operon: transzkripciós aktiválás/represszálás Az eukarióta génexpresszió szabályozása Eukarióta génregulációs fehérjék Eukarióta DNS-reguláló szekvenciák Génaktivátor fehérjék és a transzkripció A génregulációs fehérjék szinergikusan dolgoznak Eukarióta represszorok Inzulátor DNS-szekvenciák Az X-kromoszóma inaktiválása nőkben DNS-metiláció DNS-metiláció a genomi mintázatban CG-szigetek Sejtkommunikáció Az extracelluláris szignálmolekulák A szignálmolekulák hatótávolsága iii

4 Biokémia II Autokrin szabályozás Gap junction Plazmamembrán-receptorok Receptor-ioncsatornák Hét transzmembrán szegmenssel rendelkező, G-fehérje-kapcsolt receptorok Protein-kinázok, foszfoprotein-foszfatázok camp-dependens protein-kináz (protein-kináz-a) A cgmp-dependens protein-kináz (protein-kináz-g) Protein kináz-c Transzmembrán protein kinázok: növekedési faktor-receptor Foszfoprotein-foszfatázok A camp mediátorrendszer Inozitol-foszfolipid jelátviteli rendszer A guanilát-cikláz és a cgmp Nitrogén-monoxid Összetett folyamatok szabályozása Éhezés jól tápláltság A glikolízis glukoneogenezis szabályozása A glikogén felépítés lebontás szabályozása Programozott sejthalál: Apoptózis Sérült fehérjeválasz (UPR) az endoplazmás retikulumban Utószó Ábraanyag Felhasznált és ajánlott irodalom iv

5 Az ábrák listája 1.1. A sejt mint vegyipari üzem Az enzimek, mint biokatalizátorok Tipikus Michaelis Menten-kinetikát követő enzim szubsztrátkoncentráció-sebesség ábrázolása Az enzimek legfontosabb katalitikus stratégiái A lizozim katalitikus mechanizmusa Az egyik leggyakrabban előforduló prosztetikus csoport, a hem, szerkezete A sejtben folyó biokémiai reakciók összessége Feed-back gátlás. A metabolikus út egy korai szakaszán található enzimet a metabolikus út egy későbbi szakaszának terméke gátolja Többszörösen elágazó (aminosav) metabolikus út szabályozása feed-back gátlással Pozitív allosztérikus reguláció Negatív allosztérikus reguláció Egy- és több alegységes enzimek inhibitor koncentráció relatív enzimaktivitás összefüggése Két azonos alegységből álló enzim kooperatív-allosztérikus átmenete A mioglobin monomer szerkezete A hemoglobin tetramer szerkezete A hemoglobin és a mioglobin oxigéntelítési (szaturációs) görbéje A hemoglobin oxigénkötésre elszenvedett térszerkezet-változása A 2,3-biszfoszfoglicerát A hemoglobin és a 2,3-biszfoszfoglicerát kapcsolata A hemoglobin oxigéntelítési görbéje különböző ph-értéken (Bohr-effektus) Fehérje foszforiláció/defoszforiláció Szerin-/treonin- és tirozin-foszforiláció Protein kináz háromdimenziós szerkezete Kináz-kaszkád-erősítő Fehérjék aktivitásának befolyásolása foszforilációval/defoszforilációval A pepszinogén aktiválódása limitált proteolízissel Az enterolis proteázok zimogénjükből történő aktíválódása limitált proteolízissel Fehérjék transzlációjával párhuzamosan zajló folding A hsp70-család munka közben A hsp60 család szerkezeti felépítése és működése Fehérjeminőségi kontroll A proteaszóma felépítése A fehérjéhez kapcsolt ubikvitinláncok különböző szignaling jelentése A fehérjék ubikvitinációja Az ubikvitin-ligáz szabályozása Degradációs szignálok kialakulása A molekuláris biológia centrális dogmája A sejtdifferenciáció a génexpresszió megváltozása, nem a nukleotidszekvenciáé A génkifejeződés szabályozásának szintjei A DNS-kötő fehérjék kapcsolódási helye a DNS-molekula külső felszínén A nagyárok és a kisárok értelmezése a glikozidos kötések egymással bezárt szögei alapján A hélix-turn-hélix fehérjerészlet felépítése és kapcsolódása a DNS-hez A szimmetrikus DNS-regulációs szekvencia és hozzá kötődő DNS-felismerő fehérje dimer A homeodomén fehérjecsalád tagjait kódoló géneket érintő mutációk következménye (Drosophila melanogaster) A cink-ujj fehérjerészletek első csoportjának szerkezeti felépítése, illetve a klaszterképzés A leucin-cipzár fehérjerészlet szerkezete és kötődése a DNS-hez A hélix-loop-hélix fehérjerészlet szerkezete és kapcsolódása a DNS-hez Hélix-loop-hélix fehérjerészlet aktivitásának szabályozása heterodimerizációval A gél mobility shift assay (electrophoretic mobility shift assay, EMSA) működési elve A triptofán operon elemei A transzkripció lépései (1.promóter felismerése a σ-faktor által, 2.a DNS-szál felnyitása, 3. RNSszál szintézisének kezdete(iniciáció), 4. az első tíz nukleotid szintézise,5. a σ-faktor leválása a komplexről, RNS-szintézis felgyorsul (elongáció), 6. a szintézis a terminátor régióig folyik, majd a kész RNS-szál leválik a komplexről, 7. a σ-faktor ismét elfoglalja helyét az RNS-polimeráz komplexben) v

6 Biokémia II A triptofán-represszor fehérje működése A triptofán-represszor fehérje szerkezetének és DNS-kötő képességének megváltozása a két bekötődő triptofánmolekula hatására A lac-operon kettős szabályozása a lac-represszor fehérje, illetve a CAP camp-komplex által A DNS hurokképzése az enhancer- és a promóterrégiók között lehetővé teszi egymástól távoli szekvenciák egymásra hatását Az eukarióta gén szabályozási régiójának elemei Nukleoszóma-remodellezés Kovalens hisztonmódosítás A szabályozó fehérjeelemek összeszerelődése különböző funkciójú regulátorkomplexekké a DNS felületén Az enhanszoszóma felépítése Az inzulátorszekvenciák elhelyezkedése és funkciója Az X-kromoszóma inaktiválódása (kondenzálódása) a női embrionális testi sejtekben Az X-kromoszóma inaktiválódásának mechanizmusa A DNS-metiláció mechanizmusa, a fenntartó metilázok működése A genomi lenyomat hatása a génexpresszióra Az Igf2-gén metilációs lenyomata A celluláris jelátvitel szintjei Sejtfelszíni receptorok Intracelluláris magreceptorok A szignálmolekulák hatótávolsága Az extracelluláris jelre adott válasz természete, reakcióideje Az acetilkolin-nikotin receptorának izom-altípusa, az alegységek (a), az alegységek transzmembránrégiói(b), illetve az acetilkolin-kötőhelyek felülnézetből(c) A G-fehérje mint jelátviteli kapcsoló A G-fehérjék szerkezeti felépítése A G-fehérjék működés közben A camp-dependens protein-kináz felépítése A camp-dependens protein-kináz regulációja A protein kináz C felépítése és aktiválódása Transzmembrán tirozin-kináz (receptor) A camp-mediátorrendszer működés közben A camp-dependens protein kináz (PKA) szerepe a génszintű szabályozásban A foszfolipáz-c aktiválása és működése Az IP3-jelátviteli útvonal A foszfofruktokináz allosztérikus regulátorai A glikolízis/glukogeogenezis hormonális szabályozása: a foszforuktokináz II foszforilációval/defoszforilációval történő szabályozása, a camp szint hatása a glikolízis, glukoneogenezis szabályozására A glikogénlebontás (felépítés) szabályozása izom- és májsejtekben A nekrótikus és az apoptotikus sejthalál közötti morfológiai és folyamatbeli különbségek A halálligandon halálreceptoron keresztül bonyolódó külsődleges (extrinszik), illetve a mitokondrium külső membrán sérülésén, a citokrom c felszabadulásán keresztül zajló, belsődleges (intrinszik) apoptotikus útvonalak Az endoplazmás retikulum stresszt és a következményes sérült fehérjeválaszt kiváltó különböző fiziológiás, patológiás, illetve kísérleti tényezők A sérült fehérjeválasz folyamatai Az endoplazmás retikulum kiváltotta UPR, illetve az UPR elbukását követő endoplazmás retikulum eredetű apoptózis folyamata vi

7 Biokémia II. - Biokémiai szabályozás Szarka András Budapesti Műszaki és Gazdaságtudományi Egyetem, Semmelweis Egyetem, 2014 Szarka András Typotex Kiadó, ISBN: Creative Commons NonCommercial-NoDerivs 3.0 (CC BY-NC-ND 3.0) A szerző nevének feltüntetése mellett nem kereskedelmi céllal szabadon másolható, terjeszthető, megjelentethető és előadható, de nem módosítható. Készült a TÁMOP-4.1.2/A/1-11/ számú, Konzorcium a biotechnológia aktív tanulásáért című projekt keretében. vii
 A szerző nevének feltüntetése mellett nem kereskedelmi céllal szabadon másolható, terjeszthető, megjelentethető és előadható,")
8 Előszó A Biokémia II. Biokémiai szabályozás című tankönyv másodéves biomérnök hallgatók számára készült, akik a Biokémia I. tantárgy során már elsajátították a biokémiai, molekuláris biológiai alapismereteket. Ezeket alapul véve, ezekre építkezik a tankönyv, amely kiemelten a biokémiai szabályozásokkal foglalkozik. A szabályozástechnika, legyen az biológiai vagy más természetű, mindig a legnagyobb kihívást jelentő területek egyike. A tankönyvet igyekeztünk úgy felépíteni, hogy a részelemek ismertetését követően egyszerűbb, majd egyre összetettebb problémák felé haladjunk. Reméljük, sikerült mindezt világos gondolatmenet mentén megtennünk, és legalább akkora örömet jelent a tankönyv olvasása, a sejtünk titkaiba való bevezetés, mint a tankönyv kigondolása és megírása jelentett a szerző számára. Budapest, nyár Szarka András viii

9 1. fejezet - Bevezetés A tankönyv elején érdemes tisztázni, hogy mivel is foglalkozik a biokémiai szabályozástechnika. A kérdés megválaszolásához vegyük a következő példát. Tekintsünk úgy a sejtre, mint egy vegyipari üzemre. Nézzük meg, milyen hasonlóságok és analógiák vonhatók az üzem és a sejt között: üzem fal különböző specializálódott egységek sejt sejtmembrán sejtorganellumok, kompertimentumok például: energiaellátó központ mitokondrium (kloroplaszt-növényekben) raktár gépgyártó sor végszerelde központi irányítás, információszolgáltatás, elosztás 1.1. ábra - A sejt mint vegyipari üzem vakuólum (bizonyos értelemben az ER is) riboszómák endoplazmás retikulum (ER) sejtmag 1
 raktár gépgyártó sor végszerelde központi irányítás, információszolgáltatás,")
10 Bevezetés Az üzemet nyilvánvalóan el kell látni energiával. A sejt esetében ez az energia leggyakrabban a glükózból származik. A hulladék anyagokat is el kell távolítani a sejt (üzem) zavartalan működéséhez. Ilyen hulladék a CO 2, illetve egyéb toxikus vegyületek, amelyek a sejt működése során keletkeznek. Az üzem életének szigorúan szabályozott körülmények között kell folynia: a. Szigorúan meghatározott mennyiségű gépet, terméket kell előállítania. Sem a túltermelés, sem a hiány nem megengedett. b. Az egyes gyártási lépések között sem tolerálható a köztes anyagok hiánya, feleslege. c. Az energiafelhasználás (anyagfelhasználás) optimalizálására törekszünk. Ezek együttesen biztosítják az optimális gépmennyiséget. d. Fontos az egészséges munkakörülmények biztosítása, a külső vagy belső gyártás során keletkező mérgek, szennyezőanyagok eliminálása. Mindezek biztosításához szigorúan össze kell hangolni a részlépéseket, munkafolyamatokat, az egyes alapanyagok, köztitermékek, késztermékek transzportfolyamatait. A sejt esetében ezen folyamatokkal foglalkozik a biokémiai szabályozástechnika. Tanulmányaink során: 1. megismerjük a sejt gépeit, az enzimeket (mi az enzimek szerepe, milyen fontos tulajdonságokkal bírnak) 2. a biokémiai alapfolyamatokat több szinten szabályozhatjuk (a sejt működési hatásfoka [intenzitása], valamint a sejt gépeinek száma is szabályozható) a. az enzimaktivitás szabályozásának lehetőségei: i. gátlások ii. szabályozások 1. allosztérikus enzimek 2. proteolitikus hasítás 3. foszforiláció a fehérjék születése: transzláció poszttranszlációs módosulások a fehérjék aktív állapota a fehérjék bukása ubikvitizáció lebomlások, proteaszóma b. a gépek számának változtatása az enzimmennyiségek változtatása, szabályozása i. sejtdifferenciáció és génexpresszió szabályozása ii. a génexpresszió szabályozásában résztvevő DNS-t kötő fehérjék: 1. hélix-turn-hélix 2. cink-ujj fehérje 3. leucin-cippzár 2
 optimalizálására törekszünk.")
11 Bevezetés 4. hélix-loop-hélix iii. a DNS-t felismerő fehérjeszakaszok azonosítása iv. a gének ki- és bekapcsolása 1. lac-operon 2. triptofán-represszor 3. eukarióta sejtek transzkripciós szabályozása a. transzkripciós faktorok b. promóterek c. génregulációs szekvenciák d. génregulációs fehérjék v. sejtdifferenciálódás vi. poszttranszkripciós szabályozások vii. a sejtszintű információáramlás: szignál-transzdukció 1. általános kitekintés: receptorok, ligandok 2. extracelluláris jeltovábbítás: paraszimpatikus és endokrin lehetőségek 3. sejtmag receptorok: szteroid receptorok 4. sejtfelszíni receptorok: a. ioncsatorna-kapcsolt b. G-fehérjéhez kapcsolt c. enzimkapcsolt viii. célsejt adaptáció receptor leszabályozás ix. összetett folyamatok 1. glikolízis, glikogén lebontás/felépítés, vércukorszint-szabályozás 2. apoptózis, sérült fehérjeválasz (UPR) 3

12 2. fejezet - Biokatalizátorok enzimek Az enzimek szabályozásával kapcsolatos ismeretek megértéséhez szükségünk van bizonyos termodikai alaptudásra. Nézzük a következő reakciót: papír + O 2 füst + hamu + hő + CO 2 + H 2O A papír elég, hőenergiát juttat az atmoszférába, valamint vizet és széndioxidot, viszont a füst és a hamu ezekkel a dolgokkal és a melegített levegővel sohasem képes papírrá visszaalakulni. Amikor a papír elég, kémiai energiája, mint hőenergia disszipálódik. Ezzel egy időben a papír atomjai, molekulái szétszóródnak, rendezetlenek lesznek. A termodinamika nyelvén mondva szabadenergia-veszteség, szabadenergia-csökkenés következik be. Ezt az energiát használjuk fel munkavégzésre vagy kémiai reakciók végrehajtására. Általánosan elmondhatjuk: a kémiai reakciók mindig a szabadenergia-csökkenés irányába mennek végbe. Vagyis a reakciók spontán iránya az energetikailag kedvezőbb irány. Normál körülmények között a szén energetikailag legstabilabb formája a széndioxid és a hidrogéné a víz. Az élő szervezetek mégsem válnak széndioxidköddé és a papír sem kezd el lángolni kezeink között. Ennek oka, hogy az élő szervezet és a füzet molekulái egy viszonylagos stabil állapotban találhatóak és ahhoz, hogy alacsonyabb energiaszintre kerüljenek, energiát kell befektetni. Magyarul a molekuláknak aktivációs energiára van szükségük ahhoz, hogy egy jóval stabilabb (alacsonyabb energiájú) állapotba kerüljenek (2.1. ábra). Az égő füzet (papír) esetében az aktivációs energiát egy meggyújtott gyufa jelenti. A vizes oldatban található molekulák számára az energiagáton történő átjutást a szomszédos molekulákkal történő ütközés teremti meg (amely a hőmérséklet emelkedésével egyre gyakrabban következik be) ábra - Az enzimek, mint biokatalizátorok Az energiagát átugrását az élő szervezetben az enzimek segítik. Ez az energiagát-csökkentés annyira jelentős mértékű lehet, hogy az enzimek akár (több)milliószorosára is gyorsíthatják a reakciókat. Emiatt szabályozásuk 4

13 Biokatalizátorok enzimek kiemelt fontossággal bír a sejt életében, metabolizmusában. A reakció végén maguk változatlanok maradnak. Ezért tekinthetjük őket biokatalizátoroknak. Hasonlóan bármely katalizátorhoz, kizárólag termodinamikailag lehetséges reakciókat katalizálnak, mivel csak az aktivációs energiát csökkentik, ezáltal hihetetlenül megnövelve a reakcióba lépő molekulák számát (2.1. ábra); tekintve, hogy általában fehérjemolekulák, reakcióra és/vagy szubsztrátra specifikusak; (Hasonlóan bármely katalizátorhoz) Maguk nem változnak a reakció során. A hasonló reakciókat katalizáló enzimeket egyazon funkcionális csoportba soroljuk. 1. Oxidoreduktázok pl.: etanol + NAD + acetaldehid + NADH + H + 2. Transzferázok pl.: glukóz + ATP glukóz-6 foszfát + ADP 3. Hidrolázok pl.: glukóz-6-foszfát + H 2O glukóz + P i 4. Liázok pl.: 2-foszfoglicerát foszfoenolpiruvát + H 2O 5. Izomerázok pl.: glukóz-6-foszfát fruktóz-6 foszfát 6. Ligázok pl.: glutamát + NH 3 + ATP glutamin + ADP + P i A csoporton belül minden enzim specifikus, csak egy adott reakciót katalizál viszonylag szűk szubsztrát tartományon belül (gyakran csak egyetlen meghatározott molekula [szubsztrát] átalakítására képes). Például: A hexokináznak a D-glukóz szubsztrátja, de az L-glukóz már nem szubsztrátja. A trombin a fibrinben csak egy adott arginin és a szomszédos glicin között hasít, máshol nem. Természetesen ez a specifikusság különböző mértékű lehet, előfordul, hogy egy adott reakciótípusra, vagy adott funkciós csoportra korlátozódik csak, de ismertek kizárólag egyszubsztrátos enzimek is. Az enzimek hálózatot alkotnak. Az egyik enzim terméke igen gyakran egy másik szubsztrátja és így tovább. Ennek eredménye az anyagcsere utak hálózata, az energiaellátás biztosítása és a különböző molekulák szintézise Néhány szóban az enzimkinetikáról Az enzimes reakciókkal kapcsolatos, alapvető reakciókinetikai összefüggéseket, törvényszerűségeket a következő egyszerű, egyszubsztrátos enzimes reakción szeretnénk bemutatni: Az adott idő alatt az enzim által megkötött és átalakított szubszrátmolekulák száma behatárolt. Ha növeljük a szubsztrátok számát, egy maximum értékig növekszik az átalakított szubsztrát/keletkezett termék mennyisége. Ebben a pontban az enzimünk telítődött szubsztrátjával és a reakció sebessége (V max) csak attól 5

14 Biokatalizátorok enzimek függ, hogy milyen gyorsan képes az enzim a szubsztrátot átalakítani. A maximális sebességet az enzim koncentrációjával osztva megkapjuk az enzim átviteli számát, mely az angolszász irodalomban turnover number néven ismeretes. Ennek értéke egy és tízezer között szokott lenni, általában ezer körüli. Az enzimműködés kvantitatív jellemzése elengedhetetlen az enzimek megismerése során, illetve széleskörű, irányított ipari felhasználásukhoz. Tekintsük ismételten a következő egyszubsztrátos reakciót: A reakcióval kapcsolatban a következő egyszerűsítéseket tesszük: Feltételezzük, hogy az enzim (E) és a termék (P) között annyira ritkán játszódik le enzim-szubsztrát komplexet (ES) eredményező reakció, hogy ezt el is hanyagolhatjuk. Ekkor az EP komplexet nem szükséges megjelenítenünk és a reakció előrehaladását, azaz a sebességét a következőképp fejezhetjük ki: V = kcat [ES] Ebben az esetben [ES] az enzim-szubsztrát komplex koncentrációja, k cat, az átviteli szám, ami megadja, hogy egy enzimmolekula egy másodperc alatt hány szubsztrátmolekulát alakít át. Az ES komplex koncentrációja az enzim és szubsztrát molekulák összekeverését követően hirtelen nő, míg el nem éri az egyensúlyi állapotot (steady state). Ebben az állandósult állapotban koncentrációja jó közelítéssel állandónak vehető: ES fogyás = ES képződés k -1[ES] + k cat [ES] = k 1 [E][S] A szabad enzim koncentrációja: [E] = [E 0] - [ES] Behelyettesítve: Vezessük be a Km Michaelis-konstanst: Ekkor Illetve visszaírva: V = k cat [ES] Megkapjuk a híres Michaelis Menten-egyenletet: Ha növeljük a szubsztrát mennyiségét, egy maximum értékig növekszik az átalakított szubsztrát/keletkezett termék mennyisége. Ebben a pontban az enzimünk telítődött szubsztrátjával (tehát minden enzimmolekula szubsztrátot köt) és a reakció sebessége V max. Ezt a sebességet abban a pontban érjük el, ahol: V = V max = k cat [E 0] 6

15 Biokatalizátorok enzimek Ez alapján a Michaelis Menten-egyenlet általános formája a következő: Az egyenletet grafikusan ábrázolva egy hiperbolát kapunk. A grafikus ábrázolás segítségével meghatározhatóak az enzim kinetikai paraméterei a v max és a K m. A K m értéke egyszerűen megkapható, mint a v max/2 sebességhez tartozó szubsztrátkoncentráció (2.2. ábra) ábra - Tipikus Michaelis Menten-kinetikát követő enzim szubsztrátkoncentrációsebesség ábrázolása Az enzimes katalízis háttere Az enzimek különösen nagymértékben gyorsítják meg a reakciókat. Ez számos különböző tényezőnek tudható be. Lássuk, melyek ezek: 1. Az enzimek a katalitikus hely környezetében megnövelik a lokális szubsztrátkoncentrációt (azáltal, hogy megkötik őket), valamint a reakció megkívánta helyes orientációban tartják a megfelelő atomokat. 2. Talán a legfontosabb, hogy a kötési energia (szubsztrát kötés) hozzájárul a közvetlen katalízishez. A szubsztrátmolekuláknak számtalan különböző geometriájú és elektron-eloszlású köztes állapoton kell keresztülmenniük, mielőtt a reakció végét jelentő formába jutnak. A legstabilabb átmeneti állapot felvételéhez szükséges szabadenergia-mennyiség az aktivációs energia. Igazából ez határozza meg a reakció sebességét. Az enzimek jóval nagyobb affinitással bírnak az átmeneti állapotú szubsztrát, mint a stabil forma irányába. Ez a szoros kötés nagymértékben csökkenti az aktivációs energiát, így jelentősen meggyorsítja a reakciót. Az enzimek tehát többféle generális stratégiát vetnek be a katalízis során. A legfontosabbak: a. az enzim megköti és pontosan orientálja egymáshoz a szubsztrátokat b. a szubsztrát megkötésével az enzim átrendezi annak elektroneloszlását, részlegesen + és - részeket eredményezve 7

16 Biokatalizátorok enzimek c. az enzim megfeszíti a megkötött szubsztrátmolekulát, ezzel az átmeneti állapot felé tolva 2.3. ábra - Az enzimek legfontosabb katalitikus stratégiái Katalitikus antitestek Az orientációs hatás jelentőségét segítenek megérteni az ún. katalitikus antitestek. Ezek, az enzimhez hasonló térszerkezetük révén, stabilizálják az átmeneti állapotú molekulákat, így meggyorsítva az alapreakciót, igaz, nem az enzimhez hasonló mértékben (kb szeresére). Természetesen az enzimek más úton-módon is hozzájárulnak a reakció minél gyorsabb végbemeneteléhez. Pl. pontosan pozícionált atomokat tartalmaznak, amelyek képesek a szubsztrát (átmeneti állapotú molekula) elektronszerkezetét deformálni. A következőkben a lizozim segítségével nézzünk meg egy konkrét példát az enzimes katalízisre. A lizozim tulajdonképpen egy természetes antibiotikum. Megtalálható a tojásfehérjében, a nyálban, a könnyben, illetve egyéb szekrétumokban. A bakteriális sejtfal poliszacharidjait bontja, egészen pontosan az 1-4-es glikozidos kötéseket az N-acetil-muraminsav és az N-acetilglukózamin között. Ennek hatására a baktériumok nem képesek ellenállni a turgornyomásnak és szétdurrannak. Az enzimet még Fleming fedezte fel és nevezte el 1922-ben. Kisméretű (14602 Da), könnyen izolálható. Így nem csoda, hogy ez volt az első enzim, amelynek röntgendiffrakciós szerkezete ismertté vált ábra - A lizozim katalitikus mechanizmusa A lizozim által katalizált reakció, poliszacharidláncok hidrolízise, egy energetikailag kedvező, tehát szabadenergia-csökkenéssel járó reakció. Az enzim távollétében azonban vígan elvannak ezek a poliszacharid láncok még vizes közegben (oldatban) is, bármilyen észrevehető bomlás, hidrolízis nélkül. Ez érthető, hisz a korábban említett energiagát jelen esetben is érezteti hatását. A két cukormolekulát összekötő kötés kizárólag abban az esetben hasad fel a hozzá ütköző vízmolekula hatására, ha a poliszacharidlánc adott szerkezeti torzuláson megy keresztül, felveszi az átmeneti állapotot, amelyben a kötés körüli atomok geometriája és elektroneloszlása megváltozik. Ez random módon csak igen ritkán következik be, azonban jelentősen megváltozik a helyzet, amennyiben az oldat lizozimet is tartalmaz. A lizozim aktív helye, tekintve, hogy szubsztrátja egy polimer, egy hosszanti árok, amely egyszerre hat egymáshoz kapcsolt cukrot tud megkötni. A lizozim sajátságos módon tartja a szubsztrátját, úgy, hogy kifordítsa a kötésben részt vevő cukrok egyikét, kitörve a stabilis konformációjából. A felhasítandó kötést úgy pozícionálja, hogy közel kerüljön az aktív hely két savas karakterű aminosavához (egy glutamáthoz és egy aszpartáthoz) (2.5. ábra). Az enzim az enzim- 8

17 Biokatalizátorok enzimek szubsztrát komplexben a bal oldali cukrot kifeszített, egyenes konformációban tartja, miközben a 35-ös glutamáttal mint savval támadja a cukor-cukor kötést, protont donálva a jobb oldali cukornak, illetve ellensúlyként az 52-es aszpartáttal támadja a C1-szénatomon (2.5. ábra). Ennek következtében kialakul egy kovalans kötés az 52-es aszpartát és a cukormolekula C1-szénatomja között. Mindeközben a 35-ös glutamát egy vízmolekulát (pirossal jelölve) polarizál, hogy annak oxigénatomja megtámadhassa a C1-szénatomot, így kiváltva az 52-es aszpartátot. A vízmolekula (pirossal jelölve) reakciója befejezvén a hidrolízist, visszaállítja az enzim eredeti állapotát, miközben létrejön az enzim-termék komplex (2.5. ábra). Az enzimek működésükhöz nem fehérje és nem aminosav részeket is igényelnek. Ezek egy része kovalensen kapcsolódik az anya fehérjelánchoz. Ezeket nevezzük prosztetikus csoportoknak. A fehérjeláncokhoz kapcsolódó nem aminosav alkotók lehetnek fémionok vagy szerves molekulák (koenzimek, lipidek, cukrok) (2.6. ábra) ábra - Az egyik leggyakrabban előforduló prosztetikus csoport, a hem, szerkezete Az enzimek megfelelő működésének alapja a szubsztráthoz való megfelelő hozzáférés. Ha egy reakció diffúziólimitálttá válik és/vagy az egyik enzimreakció terméke a másik kiindulási anyaga, multienzimkomplexek jönnek létre. Ez hatékonyabb működést (magasabb szubsztátkoncentrációt) eredményez. Magasabb szubsztátkoncentráció kompartmentalizáció révén is biztosítható. 9
 polarizál, hogy annak oxigénatomja megtámadhassa a C1-szénatomot, így kiváltva az 52-es aszpartátot.")
18 3. fejezet - Enzimszabályozás A korábbiakban láttuk, hogy egy-egy enzim több milliószorosára, akár milliárdszorosára képes az általa katalizált reakció sebességét megnövelni. Ha figyelembe vesszük, hogy egy élő sejt az enzimek ezreit tartalmazza a kis citoszolban vagy más kis térfogatú kompartimentumban, továbbá, hogy ezek egy összetett metabolikus hálózatot alkotnak, melyben igen gyakran az egyik enzim terméke a másik szubsztrátja, továbbá, hogy ez a hálózat az elágazási pontok tömkelegét tartalmazza, beláthatjuk, hogy mindezek következtében pontos, precíz szabályozásra van szükség a problémamentes anyagcsere, sejtműködés lebonyolításához. Az anyagcsere összetettségéről és a szabályozási feladat által képviselt kihívásról képet kaphatunk, ha a 3.1. ábrára tekintünk, melyen néhány unatkozó biokémikus összefoglalta a sejtben folyó biokémiai reakciókat ábra - A sejtben folyó biokémiai reakciók összessége A sejt életfolyamatainak szabályozása tehát igen jelentős mértékben az azokat katalizáló enzimek mennyiségének, illetve aktivitásának szabályozására vezethető vissza. A szabályozás több szinten valósulhat meg: 1. Befolyásolható az enzimmolekulák száma ez majdnem kizárólagosan a génexpresszió szabályozása révén történik. 2. A sejt specializált kompertimentumokba telepíti a folyamatokat (enzimeket), itt a szubsztrátkoncentrációt a citoszoltól eltérően alakíthatja. 3. Célzott fehérjelebontással az enzim (fehérjék) féléletidejét befolyásolhatja. 4. A leggyorsabb és legáltalánosabb módszer a reakció sebességének befolyásolására az enzimaktivitás közvetlen és reverzibilis befolyásolása speciális molekulák segítségével, amelyek végső soron a fehérje térszerkezetének változásán keresztül fejtik ki hatásukat. A szabályozási szintek tárgyalását a legáltalánosabb 4. pontban ismertetett esettel, a már elkészült fehérjemolekulák aktivitásának befolyásolásával kezdjük. Kulcsmondatunk a következő lesz: A fehérjék térszerkezete és funkciója szinonim fogalom. 10

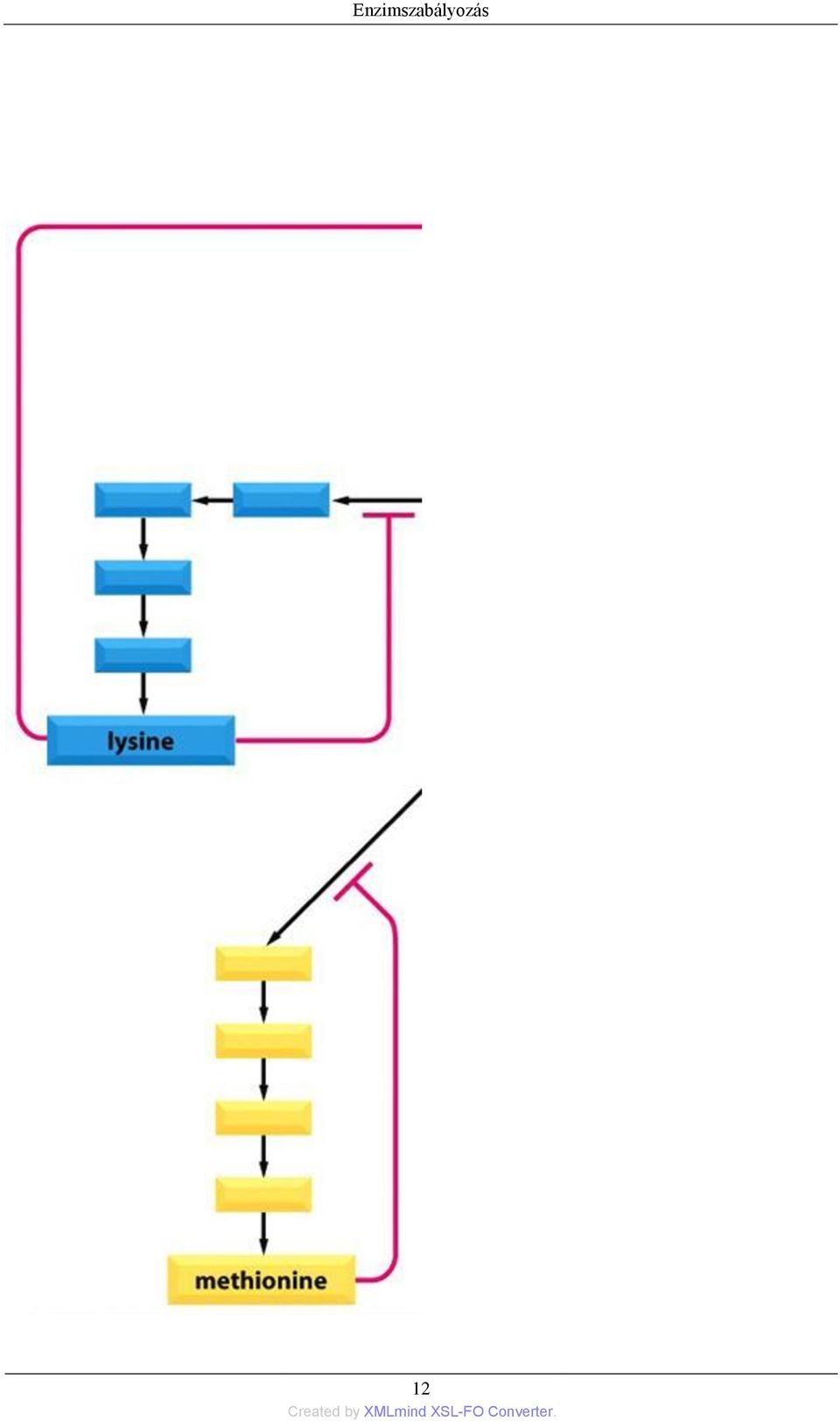
19 Enzimszabályozás Ez a mondat igen jól összefoglalja a fejezet lényegét, amennyiben befolyásoljuk, megváltoztatjuk egy fehérje térszerkezetét, akkor, az maga után vonja annak funkció- és aktivitásváltozását is! Az ilyen jellegű szabályozás igen gyakran feed-back gátlás formájában valósul meg. Feed-back gátlás során az enzimet, amely a metabolikus út egy korai szakaszán található, egy a metabolikus út későbbi szakaszának terméke gátolja. Így amikor a termék elkezd felgyülemleni, az gátolja az első enzimet, limitálva (megakadályozva), hogy további szubsztrátmolekulák lépjenek be a metabolikus útvonalba (3.2. ábra) ábra - Feed-back gátlás. A metabolikus út egy korai szakaszán található enzimet a metabolikus út egy későbbi szakaszának terméke gátolja Amikor az útvonalak elágaznak vagy egymást keresztezik, akkor többszörös szabályozópontok alakulnak ki, amelyeket a következő végtermékek kontrollálnak, mindegyik saját szintézisét befolyásolva (3.3. ábra) ábra - Többszörösen elágazó (aminosav) metabolikus út szabályozása feed-back gátlással 11
, hogy további szubsztrátmolekulák lépjenek be a metabolikus útvonalba (3.2. ábra). 3.2. ábra - Feed-back gátlás.")
20 Enzimszabályozás 12
21 Enzimszabályozás A feed-back inhibíció azonnal működésbe lép és gyorsan megszűnik, amint a termék szintje csökken. Amennyiben ez a típusú szabályozás csökkenti az enzim aktivitását, negatív szabályozásról beszélünk (3.2. ábra). Amennyiben a szabályozómolekula növeli az enzim aktivitását, pozitív szabályozásról beszélünk. Ebben az esetben a metabolikus útvonal egyik ágának terméke egy másik metabolikus útvonal enzimét aktiválja. Pl. az ADP feldúsulása aktiválja a cukor lebontásában résztvevő enzimeket, hogy az ADP ATP-vé alakulását elősegítse. Gyakran előfordul, hogy az anyagcsereút elő anyaga aktiválja a belépést katalizáló enzimet. Egymással ellentétes folyamatok esetén egy bizonyos molekula az egyik út számára allosztérikus aktivátorként, míg a visszafelé folyó út enzime számára allosztérikus inhibitorként viselkedik, így biztosítva az ellentétes utak koordinálását Allosztérikus enzimek A szabályozás elnevezése igen találó magában hordozza, annak lényegét: Allos= másik Steros=szilárd vagy háromdimenziós (térbeli) Egy szubsztráttól különböző molekula az enzim egy regulációs helyéhez kötődik (amely hely nem azonos az aktív hellyel), ezáltal befolyásolva annak aktivitását. A szabályozómolekula alakja gyakran teljesen eltérő a szubsztráthoz képest. Tehát az aktív helyhez kötődik a szubsztrát, a regulátormolekula pedig a regulációs helyhez. A két kitüntetett helynek kommunikálnia kell, hogy a regulátormolekula bekötése befolyásolhassa az aktív helyet. Ez a kommunikáció a fehérje térszerkezet-változásán keresztül valósul meg. A bekötődő szabályozómolekula konformáció változást, ez pedig aktivitásváltozást eredményez. Úgy gondoljuk, hogy a fehérjék jelentős része allosztérikus: enzimek, receptorok, szerkezeti fehérjék, motorfehérjék. A ligandok azt a konformációt stabilizálják, amelyikhez a legerősebben kötődnek, így kellően magas ligandkoncentrációnál a fehérje ezen konformációja lesz bekapcsolva. Az alaposabb megértéshez vegyük a következő példát. Adott egy fehérje, amelynek két kötőhelye van elkülönülten. Az egyikkel a szubsztrátját, a glukózt, a másikkal egy regulátormolekulát, X-et köt. Ha a glukóz bekötődése megváltoztatja az X-kötőhely alakját, akkor a két kötőhely kapcsolt. Ha mindkét ligand ugyanazon fehérje konformációt részesíti előnyben, akkor bármelyik bekötése növeli a fehérje másik irányába mutatott affinitását. Tehát ha a zárt forma köti legjobban a glukózt és ez a zárt forma szintén kedvezőbb az X-kötőhely számára, hogy X-et kössön, akkor a fehérje szorosabban köti a glukózt, ha X jelen van, mintsem ha nincs. A szabályozás ebben az esetben pozitív szabályozás (3.4. ábra) ábra - Pozitív allosztérikus reguláció 13
22 Enzimszabályozás Következésképpen a kötőhelyek összekötöttsége negatívan befolyásolja a kötődést, ha a két különböző molekula a különböző konformációkhoz szeret jobban kötődni. Ebben az esetben az első ligand bekötődése csökkenti a második (másik) ligand bekötődésének valószínűségét (3.5. ábra) ábra - Negatív allosztérikus reguláció A kötöttség mértéke mennyiségileg oda-visszaható. Ha a glukóznak nagy hatása van X kötődésére, akkor X-nek is nagy hatása van a glukóz bekötődésére. Mivel X-molekula nem a katalitikus (aktív) helyhez kötődik, nem szükségszerű, hogy bármilyen kémiai kapcsolat legyen közte és a glukóz vagy bármelyik másik ligand között, amely az aktív helyhez kötődik. Ahogy láttuk az X-molekula egyszerűen képes bekapcsolni (+ reguláció) vagy kikapcsolni (- reguláció) az enzimet. Ezáltal az allosztérikus fehérjék általános kapcsolóként működnek, amelyek megteremtik a lehetőséget, hogy egy molekula egy másik molekula sorsát meghatározza a sejten belül. 14
23 Enzimszabályozás Kooperativitás Egy alegységes enzim feed-back inhibíciója során 90% aktivitásról 10%-ra szabályozódik vissza, ha az inhibitor koncentráció 100-szorosára nő. Ez nem ad lehetőséget éles szabályozásra. A legtöbb ligand által be- és kikapcsolható enzim több egymással megegyező enzimatikus alegységből áll. Ebben az elrendeződésben, ha egy ligand beköt az egyik alegységen levő kötőhelyre, kivált egy allosztérikus változást az alegységen belül, amely később átterjedhet a szomszédos alegységekre, elősegítve őket, hogy ugyanezt a ligandot megkössék. Ennek eredményeképp egy kooperatív allosztérikus átalakulás jön létre (3.6. ábrán kék vonallal jelölve), lehetővé téve, hogy a sejtben bekövetkező kisarányú ligand koncentráció-változás teljesen aktiválja, vagy teljesen inaktiválja az egész apparátust (enzimet, fehérjét) ábra - Egy- és több alegységes enzimek inhibitor koncentráció relatív enzimaktivitás összefüggése Az első ligand bekötése nehezebb, mivel ennek során egy energetikailag kedvező kapcsolat bomlik fel az alegységek között, viszont a második ligand bekötődése jóval könnyebb, mivel ez visszaállítja a szimmetrikus molekula monomer-monomer kapcsolatát (és ezzel egyidejűleg teljesen inaktiválja az enzimet) (3.7. ábra) ábra - Két azonos alegységből álló enzim kooperatív-allosztérikus átmenete 15
24 Enzimszabályozás 16
25 Enzimszabályozás Jóval élesebb hatás érhető el, ha a ligand egy nagyobb szerkezethez, mondjuk egy 12 polipeptid láncból álló enzimhez kötődik. A szubsztrátétól eltérő szerkezetű allosztérikus effektor hatását heterotróp hatásnak nevezzük. Több azonos alegységből álló fehérje (enzim) esetén figyelték meg az allosztéria egy sajátos változatát, a homotróp hatást. Ebben az esetben egyetlen molekula, enzimek esetén maga a szubsztrát, képes betölteni az allosztérikus ligand szerepét. Az élesebb hatás alapja ez esetben is az lesz, hogy az első ligand (szubsztrát) bekötődése nehezebb, majd a bekötéssel együtt járó konformáció változás hatására a második, harmadik, negyedik szubsztráté már sokkal könnyebb lesz. A pozitív homotróp hatás érvényesülése esetén a hiperbolikus telítési görbe torzul: szigmoid lesz. Kis szubsztrát koncentráció esetén kicsi a kötődés valószínűsége, a szubsztrát koncentráció növekedtével (és a konformáció változással) a kötődési esély megnő. Ennek eredménye lesz a szubsztrát koncentráció emelkedésével meredeken emelkedő telítési görbe. A kooperativitás egyértelmű előnye tehát a szűkebb ligand/szubsztrát koncentráció-tartományban bekövetkező éles aktivitásváltozás. Ez pedig igen komoly regulációs előnyt jelent. A kooperativitás és az allosztérikus szabályozás bemutatására a legjobb példát a hemoglobin oxigénkötése adja. Így a következőkben ezzel részletesen is foglalkozni fogunk A hemoglobin szerkezete és oxigénkötése A hemoglobin szerkezetét és oxigénkötésének sajátságait, szabályozását egy másik oxigén kötésére képes molekula, a mioglobin szerkezetével, oxigénkötési sajátságaival együtt tárgyaljuk. Tesszük ezt azért, mert a párhuzamos tárgyalási mód igen fontos biokémiai szabályszerűségek megvilágítására ad módot. A két molekula igen hasonló szerkezettel rendelkezik, mindkettő globin fehérjeláncból és a hozzá kovalensen kapcsolódó hem prosztetikus csoportból áll. Igen fontos különbség azonban, hogy míg a mioglobin monomer, addig a hemoglobin tetramer (3.8. és 3.9. ábra) ábra - A mioglobin monomer szerkezete 3.9. ábra - A hemoglobin tetramer szerkezete 17
26 Enzimszabályozás Mielőtt a két oxigénkötésre képes molekula szerkezetének mélységeibe merülnénk, ejtsünk néhány szót az oxigéntárolás/szállítás szerepéről. Anyagcserénk oxigén nélkül elképzelhetetlen lenne, ez könnyen belátható, ha csak a terminális oxidáció irdatlan oxigénigényére gondolunk. A hemoglobin vitathatatlan szerepét az oxigénszállításban aláhúzza a megfigyelés, mely szerint a hemoglobinmentes vér oxigénkoncentrációja 5 ml oxigén/liter, mindez hemoglobinnal 250 ml oxigén/liter. A hemoglobin, mint minden hemoprotein, vasat (Fe[II]) és az azt koordináló protoporfirin-ix vázat tartalmaz. Ez a váz adja a hemoproteinek jellegzetes színét. A vas koordinációs kötései közül 4 a protoporfirin gyűrű N- atomjaival jön létre, 2 további kötés a hem síkjának két oldalán. Egy a globinlánc egy (proximális) hisztidinjével jön létre, ebben az irányban kiemelkedik a dezoxi-hemoglobinban a porfinváz síkjából a vasatom. A másik oldalon található az oxigén kötőhely. Itt érdemes megjegyeznünk, hogy a szén-monoxid igen veszélyes, mivel 300-szor jobban kötődik a hemoglobinhoz, mint az oxigén. Fontos tudnunk, hogy kizárólag a ferro(ii)- hemoglobin köt oxigént, a ferri (III) vagy más néven methemoglobin nem képes oxigént kötni. Ha végignézzük a hemoproteinek funkcióit, megállapíthatjuk, hogy az oxigénszállítás, kötés (Hb, Mb) Fe (II) állapotban valósul meg, azonban az elektrontranszfer-reakciókban résztvevő hemoproteinek (pl. citokrómok) esetében fontos szerepet kap a Fe (II) Fe (III) átmenet, hasonlóan a redoxireakciókhoz (pl. citokróm P450 enzimek), ahol a Fe (II) megköti az oxigént majd redukálja, miközben Fe (II) Fe (III) átmenet következik be. A rövid kitérőt követően térjünk vissza a két molekula térszerkezetéhez. A hemoglobint tehát 4 polipeptidlánc alkotja, melyek mindegyikéhez egy-egy hem tartozik. Ezekhez egy-egy oxigénmolekula kötődhet. A felnőtt hemoglobin A-t (adult) két alfa és két béta lánc alkotja (3.9. ábra). A magzati hemoglobin F (foetalis) két alfa és két gamma láncból áll. A láncokat ionos és apoláris kötések tartják össze. Ha egy pillantást vetünk a mioglobin és a hemoglobin oxigéntelítési (szaturációs) görbéjére (3.10. ábra) megállapíthatjuk, hogy azok jelentős különbséget mutatnak, míg a mioglobin klasszikus hiperbolikus telítési görbével jellemezhető, addig a hemoglobin telítési görbéje szigmoid ábra - A hemoglobin és a mioglobin oxigéntelítési (szaturációs) görbéje 18
27 Enzimszabályozás Az ábrát figyelmesebben tanulmányozva megállapíthatjuk, hogy a mioglobin alkalmatlan az oxigénszállításra, mivel a szövetekben nem képes leadni azt. Láthatjuk, hogy a hiperbolikus telítési görbe miatt a tüdőre jellemző oxigén parciális nyomáson telítődik (közel 100%-os szaturáció) oxigénnel, azonban a szövetekre jellemző parciális nyomáson még mindig igen magasan halad a görbe. A parciális nyomáskülönbségek csak igen csekély mértékű oxigén leadását tennék lehetővé (3.10. ábrán magenta nyilakkal jelölt). Nagyobb mennyiségű oxigén leadása csak igen csekély oxigén parciális nyomás mellett lehetséges. Ezzel függ össze a mioglobin fiziológiás szerepe: az izmokban történő oxigéntárolás. Oxigén tárolására kifejezetten kedvező ez a karakterisztika. A mioglobinnal szemben a hemoglobin szigmoid telítési görbével jellemezhető, így a tüdőben telítődik oxigénnel (közel 100%-os a szaturáció ebben az esetben is), a szövetekben tapasztalható alacsonyabb parciális nyomáson pedig leadja azt (3.10. ábrán kék nyíllal jelölve). A hemoglobin így ideális jelölt az oxigén szállítására. Mi állhat az eltérő karakterisztika hátterében? Ahogy azt korábban említettük, a fehérjék szerkezete és funkciója között szoros összefüggés van. A mioglobin monomer, így a szokásos hiperbolikus telítési görbével jellemezhető, a hemoglobin azonban tetramer, melynek láncai között érvényesül a kooperativitás, ami szigmoiddá teszi telítési görbéjét. Lássuk csak, hogyan támogatja a hemoglobin szerkezete a kooperativitás megvalósulását! Amennyiben a hemoglobin négy alfa vagy négy béta-láncból állna, hiperbolikus oxigéntelítési görbéje lenne. A deoxihemoglobinban (oxigént nem kötő forma) a vasatom 0,06 nm-rel kilóg a porfiringyűrű síkjából (3.11. ábra). Az oxigénkötés megváltoztatja a hemoglobin negyedleges szerkezetét. Az oxigén megkötésével a vasatom behúzódik a gyűrűbe, a vele kovalensen összekötött fehérjelánc szerkezete is megváltozik, ezért megváltoznak az alfa- és béta-láncok közötti H-híd-kötések és apoláris kölcsönhatások, valamint felszakad a deoxihemoglobint stabilizáló 8 ionpár ábra - A hemoglobin oxigénkötésre elszenvedett térszerkezet-változása 19
28 Enzimszabályozás A deoxihemoglobinban tehát ionpárok kötik össze a globinláncokat (Tense forma), míg az oxihemoglobinban (Relaxed forma) ezek felszakadnak. Így már érthető, hogy miért lesz szigmoid a hemoglobin oxigéntelítési görbéje, hisz a feszített deoxi-formát stabilizáló ionpárok akadályozzák az oxigén megkötését. (A deoximioglobin és az oximioglobin szerkezete hasonló.) Felmerül a kérdés, hogy mi stabilizálja a deoxihemoglobin negyedleges szerkezetét? A válasz megadásában segít a megfigyelés, mely szerint a hemoglobinoldatban nagyobb affinitással köti az oxigént, mint a vörösvértesten belül. Tehát a sejten belül valami lerontja a hemoglobin oxigén iránti affinitását. Ez a faktor egy glikolízis-mellékreakció során képződő, a vörösvértestben nagy mennyiségben termelődő kis molsúlyú anyag, a 2,3-biszfoszfoglicerát (3.12. ábra) ábra - A 2,3-biszfoszfoglicerát Nézzük meg, pontosan milyen szerepet tölt be a 2,3-biszfoszfoglicerát a hemoglobin-oxigénkötés szabályozásában! A 2,3-biszfoszfoglicerát ekvimorális arányban kötődik a deoxihemoglobinhoz, 1 tetramer hemoglobin-molekula 1 molekula 2,3-biszfoszfoglicerátot (2,3-BPG) köt. A 2,3-BPG-nek fiziológiás viszonyok között 5 negatív töltése van (3.12. ábra). A hemoglobin 2,3-BPG kötőhelyét a béta-láncok 4 His, illetve 2 Lys aminosava alakítja ki. A pozitív töltésű aminosav-oldalláncok és a negatív töltésű 2,3-BPG közötti kölcsönhatások olyan kötődést hoznak léte, amely stabilizálja a deoxi-hb negyedleges szerkezetét (3.13. ábra). 20
29 Enzimszabályozás A 2,3-BPG az oxihemoglobinhoz nem kötődik, az oxigenálódás folyamán a láncok elmozdulása miatt a BPG kötőhely megváltozik, szűk lesz a 2,3-BPG számára. Így az 2,3-BPG tulajdonképpen a hemoglobin allosztérikus regulátora ábra - A hemoglobin és a 2,3-biszfoszfoglicerát kapcsolata A 2,3-BPG koncentrációja nagyjából 4,5 mm és szabályozott; a szabályozás pontos mechanizmusa jelenleg ismeretlen. A glikolízisben résztvevő egyes enzimek szintézisének zavarai a vörösvértestekben befolyásolják a 2,3-BPG szintet. Így hexokináz-hiányos állapotokban a 2,3-BPG szint csökken, ezért az oxigéntelítési görbe balra tolódik, a hemoglobin oxigénaffinitása nő. Piruvát-kináz hiányos állapotokban a görbe jobbra tolódik, mert a 2,3-BPG koncentráció emelkedett, a hemoglobin oxigénaffinitása csökken. A krónikus hipoxia esetén a 2,3-BPG szint megemelkedik 4,5 mm-ról 8,0 mm-ra. Magaslati levegőn (4500 m) 2 nap alatt 7,0 mm-ra emelkedik. Tulajdonképpen ezt használják ki a hegymászók és ezért építenek akklimatizációs táborokat. Ilyenkor, pár nap alatt megemelkedik a vér 2,3-BPG szintje, aminek hatására a hemoglobin oxigénkötése leromlik, így nagyobb lesz a tüdő és szövetek telítettsége közötti különbség, jobbra tolódik a hemoglobin oxigéntelítési görbéje és jóval nagyobb mennyiségű oxigént tud leadni a szövetekben. Tengerszintre visszatérve a 2,3-BPG szintje viszonylag gyorsan visszaáll. A tárolt vér 2,3-BPG tartalma folyamatosan csökken, ezért oxigén affinitása nő. Transzfúzió esetén ez előnytelen, ezért a tárolt vérhez inozint adnak, amelynek hatására 2,3-BPG képződik a glikolízis során. 21
30 Enzimszabályozás A hemoglobin F kisebb mértékben köti a 2,3-BPG-t, mint a Hb A, így a Hb F oxigénaffinitása nagyobb, mint a Hb A-é. A Hb F Hb A-nál nagyobb oxigénaffinitása elősegíti az oxigén bejutását az anyai keringésből a magzati vérkeringésbe. Bohr-effektus A hemoglobin és a mioglobin között további lényeges különbség az, hogy a hemoglobin protonokat és széndioxidot is köt, illetve szállít. A ph csökkenésével a hemoglobin oxigénaffinitása csökken (ph 7,6 po 2= 40 Hgmm értéknél a visszatartott oxigén 80%, míg ph 6,8 po 2= 40 Hgmm értéknél a visszatartott oxigén 45%). A keletkezett széndioxid kb. 15%-a a hemoglobinhoz kötődik. Fokozott anyagcsere (pl. izommunka) esetén több széndioxid termelődik, ilyenkor az oxigénszükséglet is fokozódik. A laktáttermelés megemelkedik, a vér ph értéke csökken, a hemoglobin oxigéntelítési görbéje jobbra tolódik (3.14. ábra). A jelenséget Christian Bohr dán fiziológus (Niels Bohr fizikus édesapja) írta le, tiszteletére Bohr-effektusnak nevezték el. A képződött széndioxidot, illetve a protonok egy részét a hemoglobin úgy köti meg, hogy a béta-lánc N-terminális aminocsoportjával karbamátot (OOC-HN-R) képez. További sókötések jönnek létre, ami tovább csökkenti a hemoglobin oxigén affinitását. A tüdőalveolusokban a hemoglobin leadja a felvett széndioxidot, illetve protonokat és oxigént vesz fel. Mindezek eredményeképpen fokozott izommunka során a szövetekben leadható oxigén mennyisége jelentősen megnő ábra - A hemoglobin oxigéntelítési görbéje különböző ph-értéken (Bohr-effektus) 22
31 Enzimszabályozás Fehérjék térszerkezetének, aktivitásának befolyásolása foszforiláció/defoszforiláció által Míg az előző fejezetben ismertetett allosztérikus szabályozás során a regulátormolekula másodrendű kötésekkel pillanatszerű asszociáció/disszociáció során kapcsolódott a fehérjéhez és így igen gyorsan bekövetkezett a térszerkezet-/aktivitásváltozás, addig a fehérjék foszforilációja kovalens módosításról lévén szó: 1. jóval lassabb, 2. tartósabb, 3. enzim-katalizált (mind a foszforiláció, mind a defoszforiláció) folyamat (3.15. ábra). A kétféle szabályozási mód (allosztérikus, foszforiláció/defoszforilációval történő) kiegészíti egymást az eukarióta sejtekben. A foszforiláció két módon is érintheti, módosíthatja a fehérjéket: 1. Minden PO 4 csoport 2 negatív töltést hordoz. Ennek következtében jelentős szerkezeti változást generálhat: pl. pozitívan töltött aminosav részleteket vonzhat magához. A foszfátcsoportot eltávolítva természetesen visszaáll az eredeti helyzet. 2. A fehérjéhez kapcsolódott foszfátcsoport egy olyan szerkezet része is lehet, amelyet más fehérjék kötőhelye ismer fel. Nagyobb fehérjékben gyakran fordulnak elő olyan kis fehérjedomének, modulok, amelyek kötőhelyül szolgálhatnak más foszforilált fehérjék részére. Egy ilyen modul az SH2-domén, amely foszfotirozint tartalmazó peptidszakaszokhoz köt, de rajta kívül még vagy 10 hasonló fehérjerészlet ismeretes. A foszforiláció ilyeténképp fehérjeszerkezetek össze- és szétszerelésében is fontos tényező. A fehérjefoszforiláció mint szabályozási mechanizmus elterjedtségét mi sem mutatja jobban, mint az a megfigyelés, mely szerint az emlőssejtek fehérjéjének mintegy harmada található foszforilált állapotban az adott pillanatban. A PO csoport az ATP-terminális foszfátcsoportjának terhére történő felvitelét transzferázok, a protein kinázok katalizálják (3.15. ábra) ábra - Fehérje foszforiláció/defoszforiláció 23
32 Enzimszabályozás Az ATP hidrolízis miatt a reakció egyirányú. A sejtben több száz különböző protein kináz található, amelyek a fehérjék szerin-/treonin- (Ser/Thr kinázok) vagy tirozin (Tyr kinázok) oldalláncainak hidroxilcsoportjára transzferálják a foszfátcsoportot (3.16. ábra) ábra - Szerin-/treonin- és tirozin-foszforiláció A foszfátcsoport eltávolítását a foszfoprotein-foszfatázok katalizálják. Hasonlóan a kinázokhoz a sejt foszfoprotein-foszfatázok tömkelegét tartalmazza, amelyek specificitása jelentős különbséget mutat, némelyikük a fehérjék tágabb csoportját képes foszforilálni, vagy defoszforilálni, míg mások kizárólagosan csak egy fehérjével képesek ezt megtenni. A protein kinázok egy nagy enzimcsalád tagjai, amelynek tagjai egy nagyjából 290 aminosavas-katalitikus (kináz) doménnel rendelkeznek. A fehérjelánc további részei nagyobb változatosságot mutatnak az egyes családtagok között (3.17. ábra). Ezek felelősek a szubsztrát felismerésért, illetve a kinázaktivitás szabályozásáért ábra - Protein kináz háromdimenziós szerkezete 24
Hemoglobin - myoglobin. Konzultációs e-tananyag Szikla Károly
 Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Az sejt gépei az enzimek. Az enzimek ezt az aktivációs energiagátat csökkentik.
 Az sejt gépei az enzimek papír + O 2 füst + hamu + hő + CO 2 + H 2 O A kémiai reakciók mindig a szabadenergia csökkenés irányába mennek végbe. Miért nem alakul át minden anyag a számára legalacsonyabb
Az sejt gépei az enzimek papír + O 2 füst + hamu + hő + CO 2 + H 2 O A kémiai reakciók mindig a szabadenergia csökkenés irányába mennek végbe. Miért nem alakul át minden anyag a számára legalacsonyabb
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
A bioenergetika a biokémiai folyamatok során lezajló energiaváltozásokkal foglalkozik.
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
A METABOLIZMUS ENERGETIKÁJA
 A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2014.10.01. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2014.10.01. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA
 A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2013.10.02. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
A METABOLIZMUS ENERGETIKÁJA Futó Kinga 2013.10.02. Metabolizmus Metabolizmus = reakciók együttese, melyek a sejtekben lejátszódnak. Energia nyerés szempontjából vannak fototrófok ill. kemotrófok. szervesanyag
ENZIMSZINTŰ SZABÁLYOZÁS
 ENZIMEK 1833.: Sörfőzés kapcsán kezdtek el vele foglalkozni (csírázó árpa vizsgálata) valamilyen anyag katalizátorként működik (Berzelius, 1835.) 1850. körül: ez valamilyen N-tartalmú szervesanyag 1874.:
ENZIMEK 1833.: Sörfőzés kapcsán kezdtek el vele foglalkozni (csírázó árpa vizsgálata) valamilyen anyag katalizátorként működik (Berzelius, 1835.) 1850. körül: ez valamilyen N-tartalmú szervesanyag 1874.:
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
VEBI BIOMÉRÖKI MŰVELETEK KÖVETELMÉNYEK. Pécs Miklós: Vebi Biomérnöki műveletek. 1. előadás: Bevezetés és enzimkinetika
 VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
VEBI BIOMÉRÖKI MŰVELETEK
 VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Több szubsztrátos enzim-reakciókról beszélve két teljesen különbözõ rekció típust kell megismernünk.
 .5.Több szubsztrátos reakciók Több szubsztrátos enzim-reakciókról beszélve két teljesen különbözõ rekció típust kell megismernünk. A.) Egy enzim, ahhoz, hogy terméket képezzen, egyszerre több különbözõ
.5.Több szubsztrátos reakciók Több szubsztrátos enzim-reakciókról beszélve két teljesen különbözõ rekció típust kell megismernünk. A.) Egy enzim, ahhoz, hogy terméket képezzen, egyszerre több különbözõ
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Az enzimműködés termodinamikai és szerkezeti alapjai
 2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Bevezetés a biokémiába fogorvostan hallgatóknak
 Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
[S] v' [I] [1] Kompetitív gátlás
![[S] v' [I] [1] Kompetitív gátlás](/thumbs/93/114506146.jpg "[S] v' [I] [1] Kompetitív gátlás") 8. Szeminárium Enzimkinetika II. Jelen szeminárium során az enzimaktivitás szabályozásával foglalkozunk. Mivel a klinikai gyakorlatban használt gyógyszerhatóanyagok jelentős része enzimgátló hatással bír
8. Szeminárium Enzimkinetika II. Jelen szeminárium során az enzimaktivitás szabályozásával foglalkozunk. Mivel a klinikai gyakorlatban használt gyógyszerhatóanyagok jelentős része enzimgátló hatással bír
A biokémia alapjai. Typotex Kiadó. Wunderlich Lívius Szarka András
 A biokémia alapjai Wunderlich Lívius Szarka András Összefoglaló: A jegyzet elsősorban egészségügyi mérnök MSc. hallgatók részére íródott, de hasznos segítség lehet biomérnök és vegyészmérnök hallgatók
A biokémia alapjai Wunderlich Lívius Szarka András Összefoglaló: A jegyzet elsősorban egészségügyi mérnök MSc. hallgatók részére íródott, de hasznos segítség lehet biomérnök és vegyészmérnök hallgatók
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
ALLOSZTÉRIKUSAN SZABÁLYOZÓ METABOLITOK HATÁSA A PIRUVÁT-KINÁZ L és M IZOENZIMRE
 ALLOSZTÉRIKUSAN SZABÁLYOZÓ METABOLITOK HATÁSA A PIRUVÁT-KINÁZ L és M IZOENZIMRE A glukóz piruváttá (illetve laktáttá) történő átalakulása során (glikolízis), illetve a glukóz reszintézisben (glukoneogenezis)
ALLOSZTÉRIKUSAN SZABÁLYOZÓ METABOLITOK HATÁSA A PIRUVÁT-KINÁZ L és M IZOENZIMRE A glukóz piruváttá (illetve laktáttá) történő átalakulása során (glikolízis), illetve a glukóz reszintézisben (glukoneogenezis)
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
A metabolizmus energetikája
 A metabolizmus energetikája Dr. Bódis Emőke 2015. október 7. JJ9 Miért tanulunk bonyolult termodinamikát? Miért tanulunk bonyolult termodinamikát? Mert a biokémiai rendszerek anyag- és energiaáramlásának
A metabolizmus energetikája Dr. Bódis Emőke 2015. október 7. JJ9 Miért tanulunk bonyolult termodinamikát? Miért tanulunk bonyolult termodinamikát? Mert a biokémiai rendszerek anyag- és energiaáramlásának
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek
 Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
A gázcsere alapjai, a légzési gázok szállítása
 A gázcsere alapjai, a légzési gázok szállítása Alapfogalmak szárazföldi gerincesek: a hatékony gázcseréhez a környezet és a sejtek közötti egyszerű diffúzió nem elég - légutak kialakítása (melegítés, párásítás,
A gázcsere alapjai, a légzési gázok szállítása Alapfogalmak szárazföldi gerincesek: a hatékony gázcseréhez a környezet és a sejtek közötti egyszerű diffúzió nem elég - légutak kialakítása (melegítés, párásítás,
Elválasztástechnikai és bioinformatikai kutatások. Dr. Harangi János DE, TTK, Biokémiai Tanszék
 Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
A piruvát-dehidrogenáz komplex. Csala Miklós
 A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
Fehérjeszerkezet, és tekeredés
 Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Glikolízis. Csala Miklós
 Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Reakciókinetika és katalízis
 Reakciókinetika és katalízis 14. előadás: Enzimkatalízis 1/24 Alapfogalmak Enzim: Olyan egyszerű vagy összetett fehérjék, amelyek az élő szervezetekben végbemenő reakciók katalizátorai. Szubsztrát: A reakcióban
Reakciókinetika és katalízis 14. előadás: Enzimkatalízis 1/24 Alapfogalmak Enzim: Olyan egyszerű vagy összetett fehérjék, amelyek az élő szervezetekben végbemenő reakciók katalizátorai. Szubsztrát: A reakcióban
TRANSZPORTFOLYAMATOK A SEJTEKBEN
 16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
Kémiai reakciók sebessége
 Kémiai reakciók sebessége reakciósebesség (v) = koncentrációváltozás változáshoz szükséges idő A változás nem egyenletes!!!!!!!!!!!!!!!!!! v= ± dc dt a A + b B cc + dd. Melyik reagens koncentrációváltozását
Kémiai reakciók sebessége reakciósebesség (v) = koncentrációváltozás változáshoz szükséges idő A változás nem egyenletes!!!!!!!!!!!!!!!!!! v= ± dc dt a A + b B cc + dd. Melyik reagens koncentrációváltozását
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai
 Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói
 1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
Szerkesztette: Vizkievicz András
 Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
A felépítő és lebontó folyamatok. Biológiai alapismeretek
 A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ AMINOSAVAK ANYAGCSERÉJE 1. kulcsszó cím: Az aminosavak szerepe a szervezetben
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ AMINOSAVAK ANYAGCSERÉJE 1. kulcsszó cím: Az aminosavak szerepe a szervezetben A szénhidrátokkal és a lipidekkel ellentétben szervezetünkben nincsenek aminosavakból
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ AMINOSAVAK ANYAGCSERÉJE 1. kulcsszó cím: Az aminosavak szerepe a szervezetben A szénhidrátokkal és a lipidekkel ellentétben szervezetünkben nincsenek aminosavakból
Energiatermelés a sejtekben, katabolizmus. Az energiaközvetítő molekula: ATP
 Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Az edzés és energiaforgalom. Rácz Katalin
 Az edzés és energiaforgalom Rácz Katalin katalinracz@gmail.com Homeosztázis Az élő szervezet belső állandóságra törekszik. Homeosztázis: az élő szervezet a változó külső és belső körülményekhez való alkalmazkodó
Az edzés és energiaforgalom Rácz Katalin katalinracz@gmail.com Homeosztázis Az élő szervezet belső állandóságra törekszik. Homeosztázis: az élő szervezet a változó külső és belső körülményekhez való alkalmazkodó
Enzimaktivitás szabályozása
 2017. 03. 12. Dr. Tretter László, Dr. olev rasziir Enziaktivitás szabályozása 2017. árcius 13/16. Mit kell tudni az előadás után: 1. Reverzibilis inhibitorok kinetikai jellezői és funkcionális orvosbiológiai
2017. 03. 12. Dr. Tretter László, Dr. olev rasziir Enziaktivitás szabályozása 2017. árcius 13/16. Mit kell tudni az előadás után: 1. Reverzibilis inhibitorok kinetikai jellezői és funkcionális orvosbiológiai
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK
 POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
Fehérjék. SZTE ÁOK Biokémiai Intézet
 Fehérjék Csoportosítás Funkció alapján Szerkezetük alapján Kapcsolódó nem peptid részek alapján Szintézisük Transzkripció - sejtmag Transzláció - citoplazma Poszttranszlációs módosítások (folding) - endoplazmatikus
Fehérjék Csoportosítás Funkció alapján Szerkezetük alapján Kapcsolódó nem peptid részek alapján Szintézisük Transzkripció - sejtmag Transzláció - citoplazma Poszttranszlációs módosítások (folding) - endoplazmatikus
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL
 TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje A szénhidrátok a szervezet számára fontos, alapvető tápanyagok. Az emberi szervezetben
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA A SZÉNHIDRÁTOK ANYAGCSERÉJE 1. kulcsszó cím: A szénhidrátok anyagcseréje A szénhidrátok a szervezet számára fontos, alapvető tápanyagok. Az emberi szervezetben
Kémiai átalakulások. A kémiai reakciók körülményei. A rendszer energiaviszonyai
 Kémiai átalakulások 9. hét A kémiai reakció: kötések felbomlása, új kötések kialakulása - az atomok vegyértékelektronszerkezetében történik változás egyirányú (irreverzibilis) vagy megfordítható (reverzibilis)
Kémiai átalakulások 9. hét A kémiai reakció: kötések felbomlása, új kötések kialakulása - az atomok vegyértékelektronszerkezetében történik változás egyirányú (irreverzibilis) vagy megfordítható (reverzibilis)
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A SZÉNHIDRÁTOK 1. kulcsszó cím: SZÉNHIDRÁTOK
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A SZÉNHIDRÁTOK 1. kulcsszó cím: SZÉNHIDRÁTOK A szénhidrátok általános képlete (CH 2 O) n. A szénhidrátokat két nagy csoportra oszthatjuk:
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A SZÉNHIDRÁTOK 1. kulcsszó cím: SZÉNHIDRÁTOK A szénhidrátok általános képlete (CH 2 O) n. A szénhidrátokat két nagy csoportra oszthatjuk:
, mitokondriumban (peroxiszóma) citoplazmában
 citoplazmában") -helye: máj, zsírszövet, vese, agy, tüdő, stb. - nem a β-oxidáció megfordítása!!! β-oxidáció Zsírsav-szintézis -------------------------------------------------------------------------------------------
-helye: máj, zsírszövet, vese, agy, tüdő, stb. - nem a β-oxidáció megfordítása!!! β-oxidáció Zsírsav-szintézis -------------------------------------------------------------------------------------------
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A citoszol szolubilis fehérjéi. A citoplazma matrix (citoszol) Caspase /Kaszpáz/ 1. Enzimek. - Organellumok nélküli citoplazma
 Caspase /Kaszpáz/ 1. Enzimek. - Organellumok nélküli citoplazma") A citoplazma matrix (citoszol) A citoszol szolubilis fehérjéi 1. Enzimek - Organellumok nélküli citoplazma -A sejt fejlődéstani szempontból legősibb része (a sejthártyával együtt) Glikolízis teljes enzimrendszere
A citoplazma matrix (citoszol) A citoszol szolubilis fehérjéi 1. Enzimek - Organellumok nélküli citoplazma -A sejt fejlődéstani szempontból legősibb része (a sejthártyával együtt) Glikolízis teljes enzimrendszere
Gyógyszerrezisztenciát okozó fehérjék vizsgálata
 Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
BIOKÉMIA GYAKORLÓ TESZT 1. DEMO (FEHÉRJÉK, ENZIMEK, TERMODINAMIKA, SZÉNHIDRÁTOK, LIPIDEK)
") BIOKÉMIA GYAKORLÓ TESZT 1. DEMO (FEHÉRJÉK, ENZIMEK, TERMODINAMIKA, SZÉNHIDRÁTOK, LIPIDEK) 1. Keresse meg a baloldali oszlopban található fehérje szerkezeti szintekre jellemző a jobboldali oszlopban lévő
BIOKÉMIA GYAKORLÓ TESZT 1. DEMO (FEHÉRJÉK, ENZIMEK, TERMODINAMIKA, SZÉNHIDRÁTOK, LIPIDEK) 1. Keresse meg a baloldali oszlopban található fehérje szerkezeti szintekre jellemző a jobboldali oszlopban lévő
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
Szerves Kémiai Problémamegoldó Verseny
 Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Zsírsav szintézis. Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P. 2 i
 Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
A szénhidrátok lebomlása
 A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
A disszimiláció Szerk.: Vizkievicz András A disszimiláció, vagy lebontás az autotróf, ill. a heterotróf élőlényekben lényegében azonos módon zajlik. A disszimilációs - katabolikus - folyamatok mindig valamilyen
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Katalízis. Tungler Antal Emeritus professzor 2017
 Katalízis Tungler Antal Emeritus professzor 2017 Fontosabb időpontok: sósav oxidáció, Deacon process 1860 kéndioxid oxidáció 1875 ammónia oxidáció 1902 ammónia szintézis 1905-1912 metanol szintézis 1923
Katalízis Tungler Antal Emeritus professzor 2017 Fontosabb időpontok: sósav oxidáció, Deacon process 1860 kéndioxid oxidáció 1875 ammónia oxidáció 1902 ammónia szintézis 1905-1912 metanol szintézis 1923
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak
 BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Reakciókinetika. aktiválási energia. felszabaduló energia. kiindulási állapot. energia nyereség. végállapot
 Reakiókinetika aktiválási energia kiindulási állapot energia nyereség felszabaduló energia végállapot Reakiókinetika kinetika: mozgástan reakiókinetika (kémiai kinetika): - reakiók időbeli leírása - reakiómehanizmusok
Reakiókinetika aktiválási energia kiindulási állapot energia nyereség felszabaduló energia végállapot Reakiókinetika kinetika: mozgástan reakiókinetika (kémiai kinetika): - reakiók időbeli leírása - reakiómehanizmusok
Kutatási programunk fő célkitűzése, az 2 -plazmin inhibitornak ( 2. PI) és az aktivált. XIII-as faktor (FXIIIa) közötti interakció felderítése az 2
 és az aktivált. XIII-as faktor (FXIIIa) közötti interakció felderítése az 2") Kutatási programunk fő célkitűzése, az -plazmin inhibitornak ( PI) és az aktivált XIII-as faktor (FXIIIa) közötti interakció felderítése az PI N-terminális szakaszának megfelelő különböző hosszúságú peptidek
Kutatási programunk fő célkitűzése, az -plazmin inhibitornak ( PI) és az aktivált XIII-as faktor (FXIIIa) közötti interakció felderítése az PI N-terminális szakaszának megfelelő különböző hosszúságú peptidek
Mire költi a szervezet energiáját?
 Glükóz lebontás Lebontó folyamatok A szénhidrátok és zsírok lebontása során széndioxid és víz keletkezése közben energia keletkezik (a széndioxidot kilélegezzük, a vizet pedig szervezetünkben felhasználjuk).
Glükóz lebontás Lebontó folyamatok A szénhidrátok és zsírok lebontása során széndioxid és víz keletkezése közben energia keletkezik (a széndioxidot kilélegezzük, a vizet pedig szervezetünkben felhasználjuk).
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
Biofizika I 2013-2014 2014.12.02.
 ÁTTEKINTÉS AZ IZOM TÍPUSAI: SZERKEZET és FUNKCIÓ A HARÁNTCSÍKOLT IZOM SZERKEZETE MŰKÖDÉSÉNEK MOLEKULÁRIS MECHANIZMUSA IZOM MECHANIKA Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet
ÁTTEKINTÉS AZ IZOM TÍPUSAI: SZERKEZET és FUNKCIÓ A HARÁNTCSÍKOLT IZOM SZERKEZETE MŰKÖDÉSÉNEK MOLEKULÁRIS MECHANIZMUSA IZOM MECHANIKA Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet
A kémiai energia átalakítása a sejtekben
 A kémiai energia átalakítása a sejtekben A sejtek olyan mikroszkópikus képződmények amelyek működése egy vegyi gyárhoz hasonlítható. Tehát a sejtek mikroszkópikus vegyi gyárak. Mi mindenben hasonlítanak
A kémiai energia átalakítása a sejtekben A sejtek olyan mikroszkópikus képződmények amelyek működése egy vegyi gyárhoz hasonlítható. Tehát a sejtek mikroszkópikus vegyi gyárak. Mi mindenben hasonlítanak
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek.
 Szénhidrátok Szerkesztette: Vizkievicz András A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek. A szénhidrátok
Szénhidrátok Szerkesztette: Vizkievicz András A szénhidrátok az élet szempontjából rendkívül fontos, nélkülözhetetlen vegyületek. A bioszféra szerves anyagainak fő tömegét adó vegyületek. A szénhidrátok
Célkitűzés/témák Fehérje-ligandum kölcsönhatások és a kötődés termodinamikai jellemzése
 Célkitűzés/témák Fehérje-ligandum kölcsönhatások és a kötődés termodinamikai jellemzése Ferenczy György Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biokémiai folyamatok - Ligandum-fehérje kötődés
Célkitűzés/témák Fehérje-ligandum kölcsönhatások és a kötődés termodinamikai jellemzése Ferenczy György Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biokémiai folyamatok - Ligandum-fehérje kötődés
A cukrok szerkezetkémiája
 A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
Az X kromoszóma inaktívációja. A kromatin szerkezet befolyásolja a génexpressziót
 Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
1b. Fehérje transzport
 1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A citoszolikus NADH mitokondriumba jutása
 A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
Reakciókinetika. Általános Kémia, kinetika Dia: 1 /53
 Reakciókinetika 9-1 A reakciók sebessége 9-2 A reakciósebesség mérése 9-3 A koncentráció hatása: a sebességtörvény 9-4 Nulladrendű reakció 9-5 Elsőrendű reakció 9-6 Másodrendű reakció 9-7 A reakciókinetika
Reakciókinetika 9-1 A reakciók sebessége 9-2 A reakciósebesség mérése 9-3 A koncentráció hatása: a sebességtörvény 9-4 Nulladrendű reakció 9-5 Elsőrendű reakció 9-6 Másodrendű reakció 9-7 A reakciókinetika
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét. Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet
 Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Elektronegativitás. Elektronegativitás
 Általános és szervetlen kémia 3. hét Elektronaffinitás Az az energiaváltozás, ami akkor következik be, ha 1 mól gáz halmazállapotú atomból 1 mól egyszeresen negatív töltésű anion keletkezik. Mértékegysége:
Általános és szervetlen kémia 3. hét Elektronaffinitás Az az energiaváltozás, ami akkor következik be, ha 1 mól gáz halmazállapotú atomból 1 mól egyszeresen negatív töltésű anion keletkezik. Mértékegysége:
IX. Szénhidrátok - (Polihidroxi-aldehidek és ketonok)
") IX Szénhidrátok - (Polihidroxi-aldehidek és ketonok) A szénhidrátok polihidroxi-aldehidek, polihidroxi-ketonok vagy olyan vegyületek, amelyek hidrolízisekor az előbbi vegyületek keletkeznek Növényi és
IX Szénhidrátok - (Polihidroxi-aldehidek és ketonok) A szénhidrátok polihidroxi-aldehidek, polihidroxi-ketonok vagy olyan vegyületek, amelyek hidrolízisekor az előbbi vegyületek keletkeznek Növényi és
A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN. Somogyi János -- Vér Ágota Első rész
 A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN Somogyi János -- Vér Ágota Első rész Már több mint 200 éve ismert, hogy szöveteink és sejtjeink zöme oxigént fogyaszt. Hosszú ideig azt hitték azonban, hogy
A MITOKONDRIUMOK SZEREPE A SEJT MŰKÖDÉSÉBEN Somogyi János -- Vér Ágota Első rész Már több mint 200 éve ismert, hogy szöveteink és sejtjeink zöme oxigént fogyaszt. Hosszú ideig azt hitték azonban, hogy
Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei
 Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Gazdálkodási modul Gazdaságtudományi ismeretek I. Közgazdasá Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei KÖRNYEZETGAZDÁLKODÁSI
Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Gazdálkodási modul Gazdaságtudományi ismeretek I. Közgazdasá Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei KÖRNYEZETGAZDÁLKODÁSI
Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások
 Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Fémionok szerepe az élő szervezetben: a bioszervetlen kémia alapjainak megismerése
 Fémionok szerepe az élő szervezetben: a bioszervetlen kémia alapjainak megismerése Előadó: Lihi Norbert Debreceni Egyetem Szervetlen és Analitikai Kémiai Tanszék Bioszervetlen Kémiai Kutatócsoport A bioszervetlen
Fémionok szerepe az élő szervezetben: a bioszervetlen kémia alapjainak megismerése Előadó: Lihi Norbert Debreceni Egyetem Szervetlen és Analitikai Kémiai Tanszék Bioszervetlen Kémiai Kutatócsoport A bioszervetlen
Biológia 3. zh. A gyenge sav típusú molekulák mozgása a szervezetben. Gyengesav transzport. A glükuronsavval konjugált molekulákat a vese kiválasztja.
 Biológia 3. zh Az izomösszehúzódás szakaszai, molekuláris mechanizmusa, az izomösszehúzódás során milyen molekula deformálódik és hogyan? Minden izomrosthoz kapcsolódik kegy szinapszis, ez az úgynevezett
Biológia 3. zh Az izomösszehúzódás szakaszai, molekuláris mechanizmusa, az izomösszehúzódás során milyen molekula deformálódik és hogyan? Minden izomrosthoz kapcsolódik kegy szinapszis, ez az úgynevezett
sejt működés jovo.notebook March 13, 2018
 1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J