A mir-146a mikrorns tumor-kialakítási szerepének és mechanizmusának vizsgálata
|
|
|
- Réka Vinczené
- 7 évvel ezelőtt
- Látták:
Átírás

1 A mir-146a mikrorns tumor-kialakítási szerepének és mechanizmusának vizsgálata Doktori dolgozat Készítette: Témavezetők: Szenthe Kálmán Dr. Minárovits János Dr. Salamon Dániel Programvezető: Prof. Dr. Erdei Anna Tanszék: ELTE-TTK, Immunológia Labormunka: Országos Epidemiológiai Központ, Mikrobiológiai Kutatócsoport, 2013.
2 Tartalomjegyzék RÖVIDÍTÉSEK... 4 BEVEZETÉS... 5 IRODALMI HÁTTÉR Epigenetikai és kapcsolódó szabályozó rendszerek DNS metiláció Hiszton módosítások Szabályozó kis RNS-ek/ RNS interferencia Polycomb- és trithorax-kötő fehérjék RNS interferencia, kis RNS-ek, mikrorns-ek A kis RNS-ek felfedezéséhez vezető út Kis RNS-ek csoportosítása és szerepük MikroRNS-ek érése és származása, működésük MikroRNSek szerepe a tumorkialakulásban A mir-146a humán mikrorns Az Epstein- Barr vírus Az EBV faktorai és szerepük a génaktivitás szabályozásában Az EBV hatása a mir-146a expresszióra CÉLKITŰZÉSEK ANYAGOK ÉS MÓDSZEREK A vizsgálatok során felhasznált sejtvonalak és tenyésztésük Kontroll PCR, szekvenálás Expressziós vizsgálatok A mir-146a éretlen alakjának kimutatása, módszertani különlegességek A mir-146a érett alakjának kimutatása DNS metilációs vizsgálat Kromatin immunprecipitáció és azt követő real-time PCR DMS in vivo footprinting EREDMÉNYEK A primer átirat térszerkezet vizsgálata és kontroll szekvenálás
3 13. mir-146a expressziós vizsgálatok DNS metilációs vizsgálat Kromatin immunprecipitáció és azt követő real-time PCR DMS in vivo footprinting EREDMÉNYEK ÉRTÉKELÉSE Epigenetikai szabályozási mechanizmusok Faktorkötések és szerepük a szabályozásban EREDMÉNYEK ÖSSZEFOGLALÁSA KÖSZÖNETNYILVÁNÍTÁS IRODALOMJEGYZÉK MELLÉKLET ANGOLNYELVŰ ÖSSZEFOGLALÓ MAGYARNYELVŰ ÖSSZEFOGLALÓ
4 RÖVIDÍTÉSEK 5mC 5-metil-citozin mrns hírvivő RNS ach3 acetil-hiszton H3 NFkB nuclear factor kappa-light-chainenhancer of activated B cells ach4 acetil-hiszton H4 PAGE poliakrilamid gélelektroforézis AGO Argonauta fehérje Piwi P-element induced wimpy testis fehérje BL Burkitt-lymphoma PLZF promyelocytic leukaemia zincfinger fehérje CD40 CD40 receptor, TNF receptor család PNK T4 polinukleotid kináz ChIP Chromatin Immunprecipitáció PU.1 PU (purine rich) box binding protein 1 CpG 5 -citozin-guanin-3 RANTES CCL5, C-C motívum ligandum 5 CXCR4 C-X-C típusú kemokin receptor 4 RIG-I retinolsav indukálta gén 1 DMS Dimetil szulfát RISC RNS indukálta csendesítő komplex EGFR epidermális növekedési faktor RITS RNSi-indukálta transzkripcionális csendesítő komplex FADD Fas-associated death domain protein RNSi RNS interferencia GAPDH glyceraldehyde-3-phosphate dehydrogenase ROCK1 Rho-associated, coiled-coil containing protein kinase 1 H3K4me2 dimetil-hiszton H3 (Lys4) SNP single nucleotide polymorphism, pontmutáció IFNγ Interferon gamma SSPE Saline-Sodium Phosphate-EDTA puffer IRAK1 Interleukin-1 receptor asszociált kináz 1 Th1 és Th2 helper (segítő) T-sejt, egyes- és kettestípusú KO egér knock down egér, bizonyos gén vagy génszakasz kitörlést hordozó egér TIC Totál bevitt kromatin LMP1 Látens membrán fehérje 1 TLR4 Toll-szerű receptor 4 LM-PCR Ligáció mediálta PCR TNF Tumor nekrózis faktor LNA Locked nucleic acid TRAF6 TNF receptor asszociált faktor 6 LPS Lipopoliszacharid, bakteriális endotoxin Treg regulátor T-sejt MeCP Metiláltcitozin-kötő fehérje trns transzfer RNS mirns mikrorns VSV vezikuláris szomatitisz vírus 4
5 BEVEZETÉS A gének aktivitásának szabályozása a sejtek felépítéséhez és működéséhez elengedhetetlen fontosságú. Ez a folyamat egy jól szabályozott, több lépéses rendszert alkot, melynek az első lépése a transzkripciós szabályozás, vagyis a génről hírvivő (messenger, mrns) RNS vagy fehérjét nem kódoló RNS molekulák átírásának, szintézisének szabályozása. A gerincesek esetében a promóterek transzkripciós aktivitását meghatározó epigenetikai szabályozómechanizmusok közé a DNS metilációt és a hiszton módosításokat soroljuk. Újabban egyes szerzők a kisméretű, szabályozó RNS-ek által modulált génaktivitást ill. kromatin szerkezetet befolyásoló egyéb folyamatokat is idetartozónak gondolják. Természetesen a reguláló folyamatok tárgyalásánál a transzkripciós faktorok és az azokhoz tartozó jelátviteli útvonalak vizsgálata és ismerete is elengedhetetlen. Az epigenetikai szabályozások olyan elsődleges, génaktivitást meghatározó folyamatok, melyek alapvetően szükségesek a sejtek megfelelő működéséhez, így megváltozásuk kóros, tumoros elváltozásokhoz vezethet. Ezen folyamatokban nagy jelentőségük van olyan géneknek, amelyek a sejt normális, alapműködéséhez szükséges komponenseit (fehérjéit) kódolják. Egy részüknek elengedhetetlen jelentősége van pl. az immunrendszer működési egységének fenntartásában, így mennyiségi megváltozásuk alapjaiban változtathatja meg a működéshez fontos folyamatok egymás utáni lépéseit. Munkám során a mir-146a humán mikrorns (mir, mirns) génexpressziós szabályozásának epigenetikai és fehérje szintű vizsgálatát kívántam elvégezni, így vizsgálva a feltételezett tumor-kialakítási szerepét és mechanizmusát. Az egészséges immunsejtekben lefolyó folyamatokban fontos szerepet tulajdonítanak neki, mennyiségi megváltozása számos kóros elváltozásban kimutatható, és ez a megváltozás a tumorkialakulási folyamatok indukáló lépése lehet. A mir-146a-n kívül számos más mikrorns tumorkialakítási szerepének vizsgálata is nagy jelentőségű napjainkban, hiszen a daganatos megbetegedések gyógyítására és diagnosztikájára az eddigi kutatások eredményei szerint jó eséllyel használhatók, hiszen a kialakulásukban is központi szerepet játszhatnak. A terápiás célú alkalmazások jelenleg még korai stádiumban vannak, hiszen mikrorns prekurzor tumorsejtekbe való célzott bejuttatása, tumorsejtekben való mikrorns mennyiség csökkentés vagy növelés és/ vagy az 5
6 immunrendszer válaszának differenciálása, irányítása nagyon komplex, bonyolult folyamatok együttes megváltoztatását jelenti, így befolyásolásukhoz a működési mechanizmusokról jelentős mennyiségű tudásra van szükségünk. 6
7 IRODALMI HÁTTÉR 1. Epigenetikai és kapcsolódó szabályozó rendszerek Az epigenetika a genom bázissorrendjén kívüli többlet információkra épülő, szabályozási formákat jelenti. Ezeknek a posztszintetikus módosításoknak feladata a genomban kódolt gének megfelelő időben és sejttípusban történő expressziós aktivitásának koordinálása. Az epigenetikus szabályozások a feladatukat a kromatin szerkezet megváltoztatásával, befolyásolásával érik el. Korábban két fő szabályozási formát különítettek el a DNS metilációt, mely a DNS citozin- guanin dinukleotidjainak (CpG) citozinját érinti, valamint a hisztonmodifikációkat, melyek a jól ismert hiszton fehérjék megfelelő pozíciójú aminosav oldalláncainak acetilációja és metilációja lehetnek. Később ez utóbbit számos módosítással egészítették ki, mint pl. a foszforilációval, ubiqitinációval és sumoilációval, valamint még további szabályozó faktorokat és mechanizmusokat is együtt tárgyalnak az epigenetikai szabályozások tárgykörével. A kis, szabályozó RNS-ek felfedezése óta kitűnt, hogy a csoportba tartozó mikrorns-eknek is jelentős szerepe lehet ezekben a folyamatokban, mind a közvetlen transzkripcionális szabályozással, mind a transzkripciós faktorok transzlációjának, tehát mennyiségének szabályozásával (ld. a 1.3. fejezet). Indokoltnak látszik egy részüknek az epigenetikai szabályozási formák közötti tárgyalása. A felsorolt szabályozási módokat emellett a polycomb és trithorax fehérjék által regulált szabályozási útvonalakkal is kiegészíthetjük, megfelelően Tsai és Baylin (2011) által leírottaknak DNS metiláció A DNS-metiláció a posztszintetikus módosítások egy fajtája, melynek feladata a meglévő genetikai információk megfelelő időben, helyen és mennyiségben történő hasznosításának biztosítása és koordinálása. A kialakulása során a citozin 5. pozícióban metilálódik, amelynek következtében 5-metil-citozinná (5 m C) alakul. A metilált citozinok döntő része az 5 -citozin-guanin-3 (CpG) palindrom dinukleotid formájában található meg (Gruenbaum és mtsai., 1981). Az 5 m C a gerincesek DNS-ében található összes citozin körülbelül 2-10%-át teszi ki (Gruenbaum és mtsai., 1981; Razin és Riggs, 1980). A genom globális metilációs mintázatának kialakításában, fenntartásában és dinamikus változtatásában nagy, DNS-(5-citozin)-metiltranszferázokat (DNMT) 7
8 tartalmazó multiprotein komplexek vesznek részt (Razin és Riggs, 1980; Ehrlich és Wang, 1981). A DNS- metilációnak számos biológiai folyamat szabályozásában van kulcsfontosságú szerepe. Így részt vesz az embriogenezis és a szöveti differenciálódás szabályozásában, a kromoszómák megfelelő szerkezetének kialakításában, az integrált virális genomok, endogén retrovírusok és transzpozonok inaktiválásában, a genomikus bevésődés (imprinting) kialakításában, valamint fontos szerepet tölt be a tumorgenezisben is. A felsorolt funkciók többségének hátterében elsősorban a DNS metilációnak a transzkripcióra kifejtett hatásai állnak, ez pedig elsősorban a gének promóter-régióiban elhelyezkedő CpG-k metilációján keresztül valósul meg (Lan és mtsai, 2010). A citozinok 5. pozícióban elhelyezkedő metil-csoportja három módon befolyásolhatja bizonyos transzkripciós faktorok bekötődését. Az egyik a direkt mód, ekkor a metiláció miatt nem képes a faktor a szabályozó régióhoz kötni (pl.: c- Myc/Myn, E2F, NFκB), a másik az indirekt mód, amikor pedig a metilált CpG-hez Metiláltcitozin-kötő fehérje (MeCP) kapcsolódik, és ez gátolja a promóter transzkripcióját. Az MeCP kötődése egyrészt közvetlenül gátolhatja a transzkripciós faktorok, illetve az RNS polimeráz bekötődését és/vagy működését, másrészt különböző korepresszor fehérjékkel léphet kapcsolatba, melyeken keresztül a lokális kromatin struktúrát olyan kompakt, transzkripcionálisan inaktív formává képes alakítani, amely megakadályozza az iniciációs komplex összeszerelődését és/vagy az aktiváló transzkripciós faktorok bekötődését, illetve működését (Ehrlich és Wang, 1981). A harmadik út a magon belüli térbeli elkülönítő hatás, amely esetben a DNS-metiltranszferáz (DNMT1) enzim a megfogott kromatin régiót transzkripciós szempontból inaktív térrészekre húzza. Mivel a CpG metilációnak bizonyítottan alapvető élettani szerepe van a normális fejlődésben, a sejtek megfelelő működésében és a genetikusan kódolt információk pontos felhasználásában, igen fontos lehet ezen folyamat és a vele kapcsolatba hozható mechanizmusok vizsgálata. Erre talán a legalkalmasabb módszer a DNS biszulfitmodifikálás, amit kísérleteim során én is felhasználtam a mir-146a promóter vizsgálatában. 8
9 1.2. Hiszton módosítások A hisztonok nagy mértékű konzervativitást mutató fehérjék, ezen tulajdonságuk oka, hogy nagyon fontos funkciót látnak el. Ez a fontos funkció pedig nem más, mint a kromatin struktúrájának dinamikus kontrollja, és ezáltal a genom potenciális információtartalmának megnövelése. A kromatin DNS-ből, hisztonokból, és nemhiszton fehérjékből áll. A kromatinszálak ismétlődő nukleoszóma egységekből épülnek fel, melyek magját négy belső hiszton fehérje (H2A, H2B, H3, H4) dimerjei adják, a hisztonmag körül kb. 150 bp hosszú dupla szálú DNS kettős hélixe helyezkedik el. A nukleoszómákat összekötő DNS szakasz kapcsolja egymás után, és ehhez a DNS-hez kapcsolódik kívülről a H1, az ötödik hiszton fehérje. A hisztonmodifikációk a hisztonok N terminális farkának elsősorban lizin aminosavait érintik (Verdone és mtsai, 2005). A lizin aminosavak ε-amino csoportja metilálódhat, acetilálódhat, ubiquitinálódhat és sumoilálódhat míg az ATP γ-foszforilcsoportja általában a hisztonok szeril-oldalláncára kerül. Az ubiquitináció a hisztonok lebontásán keresztül vezet aktivitásnövekedéshez, a sumoiláció pedig a kromatin hiszton és nem hiszton fehérjéinek reverzibilis kötésével közvetlen vagy közvetett módon képes transzkripciós gátlást okozni (Lee és Connell, 2005; Gill, 2005; Iniguez-Liuhi, 2006). A hisztonmodifikációja általában egy reverzibilis állapotot jelent, aminek kialakításában egymással ellentétes enzim-párok vesznek részt. A hisztonmodifikációk típusait, módosításaikat és a DNS kromoszómális szerkezetét a 1. ábra mutatja be. 9
10 1. ábra A hiszton oldalláncok poszt-transzlációs módosításai. A hármas hiszton 9, 14, 23, 27-es pozíciójú valamint a négyes hisztonfehérje 12 és 24-es pozíciójú lizin oldalláncai metilálódhatnak ill. acetilálódhatnak. Jelmagyarázat: a képen a megfelelő hiszton fehérje (H2A, H2B, H3, H4) aminosav szekvenciája látható (S= Szerin, K= Lizin, R= Arginin), és az azokhoz kapcsolódó módosítások (acetiláció: A; metiláció: M; ubiquitináció: U; foszforiláció: P) (Santos-Rosa és Caldas, 2005). Jelenleg még nem lezárt kérdés, hogy a hisztonmodifikációk a genom információtartalmát mennyire növelik. Két egymással szemben álló hipotézis létezik, az egyik szerint a hisztonmodifikációk hatása strukturális elven alapul, vagyis hatásuk az eu- és a heterokromatin kialakításában, így az aktivitás és inaktivitás közvetlen meghatározásában van, a másik szerint pedig létezik egy úgynevezett hiszton kód (Strahl és Allis, 2000). Ez utóbbi azt jelentené, hogy a különböző hisztonmodifikációk eltérő kombinációi eltérő hatással vannak a kromatinstruktúrára és a transzkripció hatékonyságára, valamint hogy a különböző modifikációk meghatározott mintázatokat mutatnak a kölcsönös egymásra hatásuk által (Santos-Rosa és Caldas, 2005) (2. ábra). Így a lehetséges kombinációk nagy száma miatt a hisztonmodifikációk információnövelő képessége sokkal nagyobb, mint az egyszerű strukturális hatás esetén. 10
11 2. ábra A hisztonmodifikációk kölcsönös hatása egymásra. Jelmagyarázat: a képen a megfelelő hiszton fehérje (H2A, H2B, H3, H4) aminosav szekvenciája látható (S= Szerin, K= Lizin, R= Arginin), és az azokhoz kapcsolódó módosítások (acetiláció: A; metiláció: M; ubiquitináció: U; foszforiláció: P) A modifikációk közötti pozitív hatást zöld nyíl, a negatívat pedig piros jel mutatja (Santos-Rosa és Caldas, 2005) Szabályozó kis RNS-ek/ RNS interferencia Napjainkban egyes szerzők az epigenetikai szabályozási formák közé a nemtranszlálódó kis RNS-ek által modulált folyamatoknak (RNS interferencia) a transzkripciós aktivitásra ható formáit is besorolják. Az útvonal egyelőre Saccharomyces cerevisiaeben, jórészt in vitro kísérletekkel bizonyított, de a már rendelkezésre álló információk alapján állatokban és növényekben is valószínűsítik a transzkripciónális szabályozásban való fontosságát. Az élesztőben bizonyított útvonal a későbbiekben leírt mirns éréshez nagyon hasonló útvonalat használ, nagy különbség az effektor komplex felépítésében van, melyet RNAi-indukálta transzkripcionális csendesítő komplexnek (RITS) neveznek. A kompex tartalmaz RNS és DNS kötő alegységet is, melyek segítségével a kromatinhoz vagy a készülő mrns szálhoz kötve, mind a hiszton módosításokat, mind a DNS metilációt kialakító fehérjékkel együttműködve a transzkripciós aktivitás gátlását okozva. Az RNS kötő alegység (a mikrorns-ek érésénél a 2.3. fejezetben megismert 11
12 módon) egyszálú RNS-t kötve a komplementaritás segítségével a kromatin megfelelő régiójába célozza a RITS komplexet (Verdel és mtsai, 2009). A kis kettőszálú kisrns-ek további transzkripcionális kapcsolataival összefüggésben megemlítendő, hogy egy további kis RNS fajta a mikrorns (a genom területén kódolódó nukleotid hosszúságú RNS-ek (ld fejezet)) a heterokromatin kialakulásáért is felelősek, a hisztonmodifikációt és a DNS metilációt szabályozó faktorok transzlációjának szabályozásán keresztül (Chuang és Jones, 2007) Polycomb- és trithorax-kötő fehérjék A gének aktivitásának szabályozásában a fehérjéknek, transzkripciós faktoroknak is elengedhetetlen szerepe van, mivel a lokális kromatin szerkezetet ellazult (eukromatin) vagy kompakt (heterokromatin) állapotban tarthatják. Ezeknek egy része közvetlenül a DNS-hez tud kötődni, mások pedig fehérje-fehérje kölcsönhatásokon keresztül fehérje komplexekben működve regulálnak. Ez utóbbi csoportba tartoznak a polycomb és trithorax fehérjék. A polycomb fehérjéket eredetileg a Hox gének kifejeződésének szabályozása során ismerték meg (Kiefer, 2007). Kimutatták, hogy ezek a repressziós hatású fehérjék a hármas hiszton 27-es és 9-es lizinjének metilációját segítik (H3K27me3 és H3K9me2), így inaktiválva a régiót. Nemcsak hiszton metiltranszferázokkal, hanem a DNS metiltranszferázokkal is kapcsolatba hozhatók a polycomb komplex egy komponensén (EZH2) keresztül, így kiterjesztve a repressziós hatást (Viré és mtsai., 2006). Később kimutatták, hogy az RNS interferencia (RNSi) szabályozási útvonalaival is kapcsolatba hozhatók a RISC komplex (ld. a kisrns-ek éréséről szóló 2.3. fejezet) fontos fehérjéinek (Argonaute, Dicer) az esetleges kapcsolatával (Kiefer, 2007). A trithorax fehérjék a polycomb fehérjékkel hasonló módon fejtik ki hatásukat, de épp ellenkezőleg, a transzkripciós aktivitást segítve. Kimutatták a Hox gének aktivitás fokozását (Beisel és mtsai, 2002), valamint, hogy a hiszton módosítások közül a hármas hiszton négyes lizin oldalláncának metilációját (H3K4me3) fokozzák, így kapcsolódva az eukromatikus területekhez. Ezen kívül ugyanez a kutatócsoport a trithorax fehérjék RNS interferenciával való kapcsolatát is bizonyította (Sanchez-Elsner és mtsai., 2006). 12
13 2. RNS interferencia, kis RNS-ek, mikrorns-ek 2.1. A kis RNS-ek felfedezéséhez vezető út Az első létezésükről szóló cikkek a növényi vírusokkal és transzpozonokkal foglalkozó csoportok munkájának nyomán láttak napvilágot. A petúniák chalcone szintáz génjének működésésében megfigyelhető ko-szupressziós hatást vizsgálaták (Napoli és mtsai., 1990) és kimutatták, hogy virágszín erősítés céljából az általuk plazmidon bevitt chalcone szintáz gén túlexpresszálása nemhogy a színt erősíttette volna, hanem jelentős színanyag csökkenést okozott, így fehér és kevésbé kékszínű virágok keletkeztek. Kimutatták, hogy a színhiány változó mértékben, de jórészt nem örölkődő, valamint, hogy a bevitt és az eredeti szintáz gén expressziója egyaránt lecsökkent. A jelenséget nem tudták megmagyarázni, de valamilyen metilációs mechanizmust gondoltak a háttérben. A folyamatot egy később kimutatott mechanizmus segítségével lehetett magyarázni, mely az úgynevezett RNS interferencia fogalomkörébe tartozik. Az RNS interferencia olyan folyamatok összefoglaló neve, melyekben rövid, kétszálú RNS molekulák szekvencia homológia szerint céloznak meg egyszálú RNS molekulákat és azok működését vagy gátolják, vagy degradációjukat segítik elő (ez a definíció nem pontos, hiszen a transzkripcionális géncsendesítési útvonalakat is kimutatták, ahol egyszálú DNS a célpont, az élesztő gombánál, növényeknél és állatok esetében is valószínű ez a mechanizmus (Verdel és mtsai., 2009)). Az első eredmények az sirns-ek (small interfering RNA) létét bizonyították (Hamilton és Baulcombe, 1999) kimutatva egy poszt-transzkripcionális géncsendesítési (PTGS, posttranscriptional gene silencing) folyamatot, mely növényi (a szerzők szerint gombákra is jellemző) védekező mechanizmusként működik megváltozott saját vagy virális, idegen eredetű RNS-ek ellen. A növényeknél kimutatott összefüggéseket állatokban, először Caenorhabditis elegansban (Guo és Kemphues, 1996; Fire és mtsai., 1998) vizsgálták. Guo és mtsai. (1996) C. elegansban az embrionális polaritást meghatározó PAR-1 fehérje és a hozzá kötődő és működési egységet alkotó NMY-2 fehérje működési viszonyait vizsgálták, melynek során az akkor már C.elegansban alkalmazott, de nem ismert működési mechanizmusú anti-szensz gátlás (nmy-2 antiszensz RNS-t vittek be) módszerével az NMY-2 fehérje mennyiségét csökkentve fejlődési polarizációs deffektust tapasztaltak. Később megmagyarázhatóvá vált a jelenség, hiszen a módszer során nem csak egyszálú 13
14 antiszensz RNS került be, mivel a szintézisekor kis mennyiségben a komplementer szál is jelen van, így a kétszálú RNS aktiválta az RNS interferencia útvonalát, mely az NMY-2 fehérje mrns-ének mennyiségének csökkenését okozta. Fire és Mello (1998) csoportja (akik később 2006-ban a Nobel-díjat megkapták a kísérleteikért) bizonyította, hogy az úgynevezett antiszensz gátlás sokkal hatékonyabb, ha kettősszálú RNS-t juttatnak be Kis RNS-ek csoportosítása és szerepük A szabályozó kis RNS-ek számos fajtáját kimutatták, megkülönböztetik őket származás alapján és az effektor funkciót segítő fehérje (Argonatute (AGO), vagy Piwi) komplexek alapján is (Lingel és Sattler, 2005), amelyek természetesen hatásmechanizmusbeli különbséget is jelentenek. Származás, hossz és effektor funkciók alapján számos kis RNS-t különböztetnek meg napjainkban, melyeknél a sejtben végbemenő érési folyamatok jórészt közösek. A számtalan rövid, szabályozó funkciójú kis RNS közül (sirns, piwirns, snrns, tasirns stb.) a munkám során az úgynevezett mikrorns-ekhez tartozó mir-146a-t vizsgáltam behatóbban. A mikrorns-ek a genomból származó rövid, szabályozó kis RNS-ek, melyek kódolódhatnak eukarióta genomban, valamint vírusok örökítő anyagában is, regulálva a saját és a gazda genom jelátviteli útvonalaihoz tartozó faktorok mrns-einek mennyiségét (Plasterk, 2002). A prokarióták esetében kimutattak analóg, de nem homológ mechanizmusokat, melyekben ugyancsak nem kódoló rövid RNS-eknek van szabályozó szerepe (Shabalina és Koonin, 2008) MikroRNS-ek érése és származása, működésük A mikrorns-ek speciális érési útvonalon keresztül nyerik el az effektor formájukat: pri-, pre- mir formán át különböző enzim-komplexek hasításának segítségével duplaszálú, érett mikrorns-ekké alakulnak (Plasterk, 2002). A származásuk szerint keletkezhetnek egyedi génekről, melyek csak mikrorns-eket kódolnak (ld.: mir-146), vagy származhatnak intronikus régiókból. Attól függően, hogy egy átírási egységbe hány mir tartozik, klasztereket különböztethetünk meg (Lagos- 14

15 Quintana és mtsai., 2001), pl.: mir es klaszter, melynek érett mikrorns-ei központi jelentőséget töltenek be az apoptotikus folyamatok szabályozásásban. Az állati és növényi sejtek mikrorns-ei között az érésben és a működésben is vannak különbségek, mivel az állati sejtekben a folyamat két, jól elkülöníthető részre osztható, az első szakasz a sejtmagban megy végbe, a második pedig a citoplazmában, szemben a növényi sejtek érésével, melyeknél az állati sejtek citoplazmatikus szakaszának megfelelő folyamatok a sejtmagban is lejátszódhatnak (Millar és Waterhouse, 2005). A két mikrorns út különbségét a 3. ábrán, az állati mikrorns-ek érésének pontos lépéseit pedig az 4. ábrán láthatók. 3. ábra A növényi sejtekben a sejtmagban a kétszálú, nukleotid hosszú állapotig is megtörténhet az érés, amíg az állati sejtekben ez szigorúan a citoplazmatikus fázisban történik meg. A növényi sejtek esetében a végső állapot hasítás, ez az állati sejtek esetében ritka, inkább transzlációs gátlás történik (Millar és Waterhouse, 2005). 15

16 4. ábra Az állati sejtekben történő mikrorns érés pontos lépései, a folyamatra ható faktorok jelölésével. A részleges komplementaritás transzlációs gátláshoz vezet, a ritkább teljes komplementaritás ezzel szemben degradációt eredményez. Az állati és emberi sejtek esetében az RNS polimeráz II (pol II) egy elsődleges, vagy más néven primer (pri-mir) transzkriptet ír át, mely a termodinamikai összefüggéseknek megfelelő, részleges kettőszálú térszerkezetet vesz fel. Ennek hatására hajtű régiók alakulnak ki, melyekről mikrorns-ek képződhetnek. A képződő duplaszálú pre-mikrorns alakot egy Drosha nevű RNase III típusú enzim vágja le a primer formáról. Az enzim általi levágás és felismerés szigorúan meghatározott szabályok alapján történik, melyeknek vizsgálata napjainkban sem fejeződött be. A következő lépésben az állati sejtek esetében a sejtmagból a citoplazmába kerül a hurkos, kétszálú pre-mikrorns egy membránfehérje, az Exportin-5 segítségével. A növényi sejtekben ezzel szemben ez még a sejtmagi folyamat része. A citoplazmába került duplaszálú, hurkos forma ezután hasításon megy keresztül, melyet egy RNase III típusú enzim, a Dicer végez el. A kétszálú, nukleotid hosszúságú termék a következő lépésben egy helikáz enzim segítségével egyszálúvá alakul, így beépülve egy fehérje komplexbe, a RISC-be (mely az RNS indukálta csendesítő komplex (RNA induced silencing complex) kifejezésből származik), és mrns-ek repressziójában, vagy az állati sejtek esetében ritkán, mrns hasításban vesz részt (Pontes és Pikaard, 2008). A kis RNS-ek főbb rendszerezési módja a RISC komplex egyik komponense alapján történik, mely alapvetően meghatározza a bekötődött kis RNS és a komplex effektor funkcióját, vagyis a represszió, hasítás vagy esetleg transzkripciós-aktivitás szabályozást. Az első leírt ilyen fehérje az Argonaute (AGO) fehérje volt, melynek a 16
17 szakirodalomban mára már számos fajtája kimutatott (AGO1-7), valamint más, AGO típusú fehérjét is leírtak pl. Piwi (Benfey, 1999). 3. MikroRNSek szerepe a tumorkialakulásban Korábbi kutatások kimutatták (összefoglalva: Calin és mtsai., 2006; Szenthe és mtsai, 2013a), hogy különböző kóros elváltozásokban a folyamatokra jellemző mikrorns mintázat figyelhető meg, melyet az egészséges sejtekhez, szövethez képest az adott mikrorns csökkent vagy megemelkedett expressziós mennyisége jellemez. Diagnosztikai alkalmazás céljából számos kutatócsoport kísérleteket végzett/ végez a mikrorns-ek mennyiségi meghatározására különböző kóros folyamatok esetében. Bizonyos mir-ek felhasználhatók diagnosztikai markernek, melynek feltétele, hogy a kiválasztott mir vagy mir-ek jellemzően az adott elváltozásban mutassanak be jelentősen megnövekedett vagy lecsökkent expressziót. Az 1. táblázat a jelenlegi kutatások eredményét tartalmazza, összefoglalva a B- sejtes limfoid tumorok példáján az egészséges szövettől expressziós szinten jelentősen eltérő mikrorns-eket. Az adatok a különböző kutatócsoportok munkája alapján jelentősen szórnak, részben a kiindulási minták származása miatt (sejtvonal vagy klinikai eredetű minták), részben a nem megfelelő pontossággal publikált eredmények (pl. Sandhu és mtsai., 2011), akiknél a hivatkozások nem a releváns magyarázatra mutatnak), összefüggések miatt, mely nem segíti a korrekt összefoglalás elkészítését. A jelentős mennyiségű adat ettől függetlenül lehetővé teszi, hogy mikrorns-eket vagy mikrorns csoportokat jelöljünk meg, amelyek a folyamatok ismeretében később jó eséllyel terápiás, vagy prognosztikai céllal felhasználhatók lesznek. Az első táblázatban megfigyelhetjük, hogy előfordul olyan B-sejtes limfóma (a diffúz B-sejtes és a krónikus limfoid leukémia), mely esetében a kutatócsoportok ugyan ahhoz a csoporthoz tartozó mikrorns-t a megnövekedett és a lecsökkent mennyiségű mikrorns-eknél is kiugró jelentőségűnek jelölték, ez részben az eltérő mintaeredettel (sejtvonal vagy klinikai eredetű minták) magyarázható (pl. Gibcus és mtsai, 2009 vs. Di Lisio és mtsai., 2012; a Krónikus limfotikus leukémia (CLL) esetében a mir klaszter mikrorns-ei). A másik esetben a diffúz B-sejtes limfóma jelentős mikrornsei között mindkét csoportban szerepel a let-7 család egy tagja (tumor szupresszor hatású 17
18 mir-ek), valamint a mir-143 és mir-145 ugyancsak tumorszupresszor hatású mir-ek is. Ezt az eredményt nem lehet az elérhető adatok alapján megmagyarázni, mivel mindhárom csoport (Lawrie és mtsai, 2009; Di Lisio és mtsai., 2012; Roehle és mtsai, 2008) klinikai mintákkal dolgozott. A B-sejt eredetű limfómák esetében megfigyelhető expressziós változások összefoglalásaként a teljesség igénye nélkül kimondhatjuk, hogy a jellemző a mir-155 (B-sejt fejlődési és túlélési szerep), a mir-181 (proliferációs szabályozás), a mir es klaszter (proliferációt, angiogenezist serkentik, anti-apoptotikus hatásúak), valamint a let-7 családba (tumor szupresszor hatású) tartozó mir-ek megváltozott expressziója. A jelentős mértékben lecsökkent mir-ek között az általam vizsgált mikrorns, a mir-146a (részletes bemutatás: ld.: 4. fejezet) két esetben szerepel, a krónikus limfoid leukémiánál (Di Lisio és mtsai, 2012) és a Burkitt limfómánál (Robertus és mtsai, 2010), mely a későbbiekben (4. és 5. fejezet) tárgyalt összefüggések segítségével magyarázható. Az EBV-t tartalmazó Burkitt limfóma esetekben a mir-146a mennyiségének csökkenése az LMP1 (látens membrán protein 1) által aktivált NFkB útvonal aktiválását eredményezi, melynek fontos szerepét igazolták a B-sejtek immortalizációjának kialakulásában (Kaye és mtsai, 1993). Az LMP1 az EBV által, a megfelelő látencia állapotban (III. látencia típus) termelt membrán fehérje (CD40 homológ, tumor nekrózis faktor (TNF) receptor család). Az LMP1 jelentős mértékben fokozza számos adhéziós fehérje, aktivációs marker és apoptózist gátló fehérje expresszióját (Bornkamm és Hammerschmidt, 2001) az említett útvonal segítségével. A kutatások eredményeként elmondhatjuk, hogy noha a sejtvonalakon és klinikai mintákon végzett vizsgálatoknak sikerült jellemző expressziós mintázatokat meghatározni, gyakran ugyanazok a mir-ek mennyisége változik a különböző kórfolyamatokban, hiszen ezek a mir-ek jelentős reguláló szerepet látnak el az apoptotikus és anti-apoptotikus jelek, valamint a sejtek közötti kommunikációs és sejten belüli jelátvivő útvonalak szabályozásában. A mikrorns-ek a fent felsorolt okok miatt valószínűleg a közeljövőben nem lesznek alkalmasak diagnosztikai markernek, hiszen a tumor-specificitás hiánya miatt nagy felbontású módszerek szükségesek a vizsgálatukhoz, melyek jelenleg nagyon költségesek. 18
19 1. táblázat: Potenciálisan tumor markernek és terápiás céllal alkalmazható mirns-ek a B-sejtes daganatok esetében (összefoglaló táblázat). B-sejtes limfóma Diffúz nagy B- sejtes limfóma (DLBCL) Krónikus limfoid leukémia/kis limfocitás limfóma (CLL/SLL) Megnövekedett mennyiségű mikrornsek mir-19b (Gibcus és mtsai., 2009) mir-17-5p (Roehle és mtsai., 2008) mir-143 (Lawrie és mtsai., 2009) Szerepe, feladata, kapcsolatai Csökkent mennyiségű mikrornsek Szerepe, feladata, kapcsolatai a mir klaszter onkogén tagja, proliferációt, angiogenezist segítő hatású, az apoptotikus útvonalakat gátolja a Bim és Pten faktorok képződésének gátlásával. proliferációt, angiogenezist segítő hatású, az apoptotikus útvonalakat gátolja a Bim és Pten faktorok képződésének gátlásával. a mir-145 és a mir-143 tumor szupresszor szerepű mikrornsek, a Ras aktiváció a mennyiségbeli csökkenésükhöz vezet emberi és rágcsáló sejtekben onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést eredményez mir-145 (Roehle és mtsai., 2008) mir-150 (Roehle és mtsai., 2008) mir-139 (Roehle és mtsai., 2008) a mir-145 és a mir-143 tumor szupresszor szerepű mikrornsek, a Ras aktiváció a mennyiségbeli csökkenésükhöz vezet emberi és rágcsáló sejtekben a c-myb transzkripciós faktor expresszióját szabályozza és fontos szerepet tölt be a B-sejt fejlődés folyamatában (Xiao és mtsai., 2007) Rho-kinase 2 gátlásával a metasztázis és a májrák fejlődését szabályozza mir-145 (Lawrie és mtsai., 2009) mir-149 (Roehle és mtsai., 2008) számos tumoros sejtben apoptozist indukál az Akt1 és E2F1 gátlásával mir-155 (Rai és mtsai., 2008) és (Di Lisio és mtsai., 2012) mir-320 (Roehle és mtsai., 2008) tranfekciója eredményeképpen szignifikánsan csökkent az ERK1/2 mennyisége mir-29b (Di Lisio és mtsai., 2012) p53 fehérje mennyiségét szabályozza (Park és mtsai., 2009) let-7e (Roehle és mtsai., 2008) let7 család tagja, tumor szupresszor hatású mir-146a (Di Lisio és mtsai., 2012) negatívan szabályozza az IRAK1 és TRAF6 faktorokat, melyek központi szerepűek a Toll-like, valamit citokin receptorhoz kapcsolt jelátviteli útvonalak működésében (ld.: szöveg) mir-365 (Di Lisio és mtsai., 2012) az IL-6 receptor közvetlen inhibítora mir-30b (Di Lisio és mtsai., 2012) B-sejt fejlődés gátlása (Fischer és mtsai., 2011) let-7f (Di Lisio és mtsai., 2012) let7 család tagja, tumor szupresszor hatású mir-9 (Di Lisio és mtsai., 2012) MYC/MYCN által aktivált mikrorns, E-cadherin mennyiségét és tumor metasztázist szabályoz mir-34b (Di Lisio és mtsai., 2012) tumor szuppresszor mikrorns, a promótere gyakran metilált a tumorokban pl. melanoma mir-197 (Di Lisio és mtsai., 2012) a FUS1 tumor szuppresszor gént gátolja (Du és mtsai., 2009) let-7 család (a,d,e,f,g) (Di Lisio és mtsai., 2012) let7 család tagja, tumor szupresszor hatású mir-206 (Di Lisio és mtsai., 2012) a Smad3 gátlásával a HDAC4, TGF-β-általi aktiválását gátolja, így mir-100 (Di Lisio és mtsai., 2012) az IGF2 faktor mennyiségét csökkenti emlő karcinómában növelve a hiszton acetilációt, és fokozva a gén aktivitást (Braggio és mtsai., 2012) mir-370 (Di Lisio és mtsai., 2012) a TRAF4 gátló mikrornse, mennyisége csökken a sztromális bélhám tumorokban (gastrointestinal stromal tumors) mir-125a (Di Lisio és mtsai., 2012) feltételezések szerint az NFkB negatív regulátorának a TNFAIP3-nek a közvetlen gátló mikrornse mir-19a (Gibcus és mtsai., 2009) proliferációt, angiogenezist segítő hatásúak, az apoptotikus útvonalakat gátolják a Bim és Pten faktorok képződésének gátlásával *** mir-126, * (Di Lisio és mtsai., 2012) fokozott expresszióját figyelhetjük meg az akut mielómás leukémia esetek egy részében transzlokáció eredménye képpen (Li és mtsai., 2008) mir-20a (Gibcus és mtsai., 2009) mir-143 (Di Lisio és mtsai., 2012) a mir-145 és a mir-143 tumor szupresszor szerepű mikrornsek, a Ras aktiváció a mennyiségbeli csökkenésükhöz vezet emberi és rágcsáló sejtekben mir-92 (Gibcus és mtsai., 2009) mir-146a (Di Lisio és mtsai., 2012) negatívan szabályozza az IRAK1 és TRAF6 faktorokat, melyek központi szerepűek a Toll-like, valamit citokin receptorhoz kapcsolt jelátviteli útvonalak működésében (ld.: szöveg) mir-483 (Di Lisio és mtsai., 2012) a középbél, máj és emlő karcinómákban fokozott expressziója figyelhető meg mir-15b (Calin és mtsai., 2002) az E2F1 transzkripciós faktor által növekszik a mennyiségük, a proliferációt fokozzák mir-485 (Di Lisio és mtsai., 2012) a Nuclear factor (NF)-YB feltétellezett regulátora mir-16 (Calin és mtsai., 2002) mir-17-5p (Di Lisio és mtsai., 2012) mir-20a,b (Di Lisio és mtsai., 2012) mir-182 (Di Lisio és mtsai., 2012) mir-223 (Di Lisio és mtsai., 2012) proliferációt, angiogenezist segítő hatásúak, az apoptotikus útvonalakat gátolják a Bim és Pten faktorok képződésének gátlásával. túlzott mennyisége metasztázist eredményez in vivo körülmények között sejt proliferációt gátol az IGF gátlásával mir-34a (Di Lisio és mtsai., 2012) mir-365 (Di Lisio és mtsai., 2012) mir-451 (Di Lisio és mtsai., 2012) mir-7 (Di Lisio és mtsai., 2012) tumorszupresszor hatás az IL-6 receptor közvetlen inhibítora eritrocita fejlődést szabályoz tumor növekedés és metasztázis gátló hatású, májkarcinóma esetekben a phosphoinositide 3-kinase/Akt útvonal a feltételezett célpontja
20 Nodális marginális zóna B-sejtes limfóma mir-150 (Di Lisio és mtsai., 2012) mir-221 (Arribas és mtsai., 2012) a c-myb transzkripciós faktor expresszióját szabályozza és fontos szerepet tölt be a B-sejt fejlődés folyamatában (Xiao és mtsai., 2007) fontos szerepe van az angiogenezisben és a Vegfc/Flt4 jelátviteli útvonalban, a Cdkn1b és a Pik3r1 fehérjék gátlásával sejtosztódást indukál mir-370 (Haller és mtsai., 2010) a TRAF4 gátló mikrornse, mennyisége csökken a sztromális bélhám tumorokban (gastrointestinal stromal tumors) mir-223 (Arribas és mtsai., 2012) let-7 család (Di Lisio és mtsai., 2012) let-7f (Arribas és mtsai., 2012) sejt proliferációt gátol az IGF gátlásával let7 család tagja, tumor szupresszor hatású Splenicus marginális zóna limfóma Extranodális marginális zóna B- sejtes limfóma (MALT) Follikuláris limfóma (FCL) mir-144/451 család (Di Lisio és mtsai., 2012) eritrocita fejlődést szabályoz mir-141 (Di Lisio és mtsai., 2012) a mir-141 és a mir-200a gátolja a p38α fehérjét és modulálja az oxidatív stresszre adott választ mir-200c (Di Lisio és mtsai., 2012) A mir-200 család gátolja a metasztázis inicializációját, közvetlen célpontja az E-cadherin transzkripcionális gátlóinak (ZEB1 és ZEB2) (Park és mtsai., 2008) mir-210 (Di Lisio és mtsai., 2012) apoptózis indukáló faktor, mitokondrium asszociált, valószínűleg az AIFM3 fehérje a közvetlen célpontja mir-200 klaszter (Di Lisio és mtsai., 2012) A mir-200 család gátolja a metasztázis inicializációját, közvetlen mir-126* (Di Lisio és mtsai., 2012) fokozott expresszióját figyelhetjük meg az akut mielómás leukémia célpontja az E-cadherin transzkripcionális gátlóinak (ZEB1 és ZEB2) esetek egy részében transzlokáció eredménye képpen (Li és mtsai., (Park és mtsai., 2008) 2008) mir-429 (Di Lisio és mtsai., 2012) a c-myc fehérje mrns-e a feltételezett célpontja mir-141 (Di Lisio és mtsai., 2012) a mir-141 és a mir-200a gátolja a p38α fehérjét és modulálja az oxidatív stresszre adott választ mir-138 (Di Lisio és mtsai., 2012) p53-at negatvan szabályoz mir-320 (Roehle és mtsai., 2008) tranfekciója eredményeképpen szignifikánsan csökkent a ERK1/2 mennyisége mir-9 (Di Lisio és mtsai., 2012), 9* (Roehle és NFkB regulátor, MYC/MYCN által aktivált mikrorns, E-cadherin mir-149 (Roehle és mtsai., 2008) számos tumoros sejtben apoptozist indukál az Akt1 és E2F1 gátlásával mtsai., 2008) mennyiségét és tumor metasztázist szabályoz, PRDM1/BLIMP1 mennyiségét csökkenti mir-155 (Roehle és mtsai., 2008) onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést eredményez mir-139 (Roehle és mtsai., 2008) Rho-kinase 2 gátlásával a metasztázis és a májrák fejlődését szabályozza mir-210 (Roehle és mtsai., 2008) apoptózis indukáló faktor, mitokondrium asszociált, valószínűleg az AIFM3 fehérje a közvetlen célpontja mir-301 (Roehle és mtsai., 2008) NFkB-n keresztül proliferációt szabályoz mir-143 (Lawrie és mtsai., 2009) a mir-145 és a mir-143 tumor szupresszor szerepű mikrornsek, a mir-145 (Lawrie és mtsai., 2009) Ras aktiváció a mennyiségbeli csökkenésükhöz vezet emberi és rágcsáló sejtekben mir-221 (Lawrie és mtsai., 2009) fontos szerepe van az angiogenezisben és a Vegfc/Flt4 jelátviteli útvonalban, a Cdkn1b és a Pik3r1 fehérjék gátlásával sejtosztódást indukál let-7b (Lawrie és mtsai., 2009) let7 család tagja, tumor szupresszor hatású let-7i (Lawrie és mtsai., 2009) mir-494 (Arribas és mtsai., 2012) a PTEN faktor mrns-ének mennyiségét szabályozza 20
21 Köpenysejtes (Mantle sejtes) limfóma Burkitt limfóma (BL) Primer központi idegrendszeri limfóma (PCNSL) Limfoplazmocitás limfóma (Waldenstrom makroglobulinémia, WM) mir-183 (Di Lisio és mtsai., 2012) az EGR1 tumorszupresszor gén feltételezett regulátora mir-126* (Di Lisio és mtsai., 2010) fokozott expresszióját figyelhetjük meg az akut mielómás leukémia esetek egy részében transzlokáció eredménye képpen (Li és mtsai., 2008) mir-200c (Di Lisio és mtsai., 2012) A mir-200 család gátolja a metasztázis inicializációját, közvetlen mir-29a, b, c (Zhao és mtsai., 2010) p53-at regulál (Park és mtsai., 2009) célpontja az E-cadherin transzkripcionális gátlóinak (ZEB1 és ZEB2) (Park és mtsai., 2008) mir-182 (Di Lisio és mtsai., 2010) túlzott mennyisége metasztázist eredményez in vivo körülmények között mir-150 (Zhao és mtsai., 2010) a c-myb transzkripciós faktor expresszióját szabályozza és fontos szerepet tölt be a B-sejt fejlődés folyamatában (Xiao és mtsai., 2007) mir-181 (Di Lisio és mtsai., 2010) proliferációt szabályoz, túlzott expressziója apoptótikus hatású mir-15a, b (Zhao és mtsai., 2010) az E2F1 transzkripciós faktor által növekszik a mennyiségük, a proliferációt fokozzák mir-155 (Zhao és mtsai., 2010) onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést eredményez mir-210 (Zhao és mtsai., 2010) apoptózis indukáló faktor, mitokondrium asszociált, valószínűleg az AIFM3 fehérje a közvetlen célpontja mir-17-3p (Di Lisio és mtsai., 2012) proliferációt, angiogenezist segítő hatásúak, az apoptotikus let-7 család (a,d,e,f,g) (Di Lisio és mtsai., 2012) let7 család tagja, tumor szupresszor hatású mir-17-5p (Onnis és mtsai., 2010) útvonalakat gátolják a Bim és Pten faktorok képződésének gátlásával. mir-29 család (a,b,c) (Di Lisio és mtsai., 2012) p53-at regulál (Park és mtsai., 2009) mir-18a (Di Lisio és mtsai., 2012) mir-451 (Di Lisio és mtsai., 2012) eritrocita fejlődést szabályoz mir-19a (Gibcus és mtsai., 2009) és (Di Lisio és mtsai., 2012) mir-146a (Robertus és mtsai., 2010) negatívan szabályozza az IRAK1 és TRAF6 faktorokat, melyek központi szerepűek a Toll-like, valamit citokin receptorhoz kapcsolt jelátviteli útvonalak működésében (ld.: szöveg) mir-19b (Gibcus és mtsai., 2009) és (Di Lisio és mtsai., 2012) mir-150 (Di Lisio és mtsai., 2012) a c-myb transzkripciós faktor expresszióját szabályozza és fontos szerepet tölt be a B-sejt folyamatában (Xiao és mtsai., 2007) mir-20a (Onnis és mtsai., 2010) mir-155 (Robertus és mtsai., 2010) onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést mir-92 (Di Lisio és mtsai., 2012) eredményez mir-17-5p (Fischer és mtsai., 2011) proliferációt, angiogenezist segítő hatásúak, az apoptotikus útvonalakat gátolják a Bim és Pten faktorok képződésének gátlásával. A Myc fehérjéhez kapcsolódó jelátviteli útvonalhoz tartoznak (Fischer és mtsai., 2011) mir-9 (Fischer és mtsai., 2011) MYC/MYCN által aktivált mikrorns, E-cadherin mennyiségét és tumor metasztázist szabályoz mir-30b/c (Fischer és mtsai., 2011) B-sejt fejlődés gátlása (Fischer és mtsai., 2011) mir-155 (Fischer és mtsai., 2011) onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést eredményez mir-145 (Fischer és mtsai., 2011) mir-20a (Fischer és mtsai., 2011) mir-214 (Fischer és mtsai., 2011) az ATF4 faktor a célpontja a mir-145 és a mir-143 tumor szupresszor szerepű mikrornsek, a Ras aktiváció a mennyiségbeli csökkenésükhöz vezet emberi és rágcsáló sejtekben mir-29a, b (Hodge és mtsai., 2011) p53-at regulál (Park és mtsai., 2009) mir-9 (Roccaro és mtsai., 2009), (Hodge és mtsai., 2011) és (Braggio és mtsai., 2012) MYC/MYCN által aktivált mikrorns, E-cadherin mennyiségét és tumor metasztázist szabályoz let-7a, f, g (Hodge és mtsai., 2011) let7 család tagja, tumor szupresszor hatású mir-152 (Hodge és mtsai., 2011) tumor szupresszor funkciójú mikrorns mir-21 (Hodge és mtsai., 2011) tumorgenezist regulál olyan faktorok szabályozásával, mint a bcl-2 mir-182 (Hodge és mtsai., 2011) túlzott mennyisége metasztázist eredményez in vivo körülmények között mir-125b (Hodge és mtsai., 2011) sejtosztódást gátol a Mad1 gátlásával és a SAC faktor átmeneti aktiválásával mir-15a (Braggio és mtsai., 2012) tumor szupresszor funkciójú mikrorns, a deléciója a Bcl-2-höz kapcsolható anti-apoptotikus útvonal gátlásához vezet (Braggio és mtsai., 2012) mir-181a (Hodge és mtsai., 2011) proliferációt szabályoz, túlzott expressziója apoptótikus hatású mir-16 (Braggio és mtsai., 2012) tumor szupresszor funkciójú mikrorns, a deléciója a Bcl-2-höz kapcsolható anti-apoptotikus útvonal gátlásához vezet (Braggio és mtsai., 2012) mir-155 (Braggio és mtsai., 2012) és (Roccaro és mtsai., 2009) mir-206 (Braggio és mtsai., 2012) mir-223 (Hodge és mtsai., 2011) onkogén mikrorns, sejt szaporodást szabályoz, a sejt túlélést eredményez a MAPK/ERK, PI3/AKT és NF-kB jelátviteli útvonalak gátlásával (Braggio és mtsai., 2012) a Smad3 gátlásával a HDAC4, TGF-β-általi aktiválását gátolja, így növelve a hiszton acetilációt, és fokozva a gén aktivitást (Braggio és mtsai., 2012) sejt proliferációt gátol az IGF gátlásával *** Gibcus és mtsai (2009) sejtvonalakt használtak a vizsgálataikban a Di Lisio és mtsai-val (2012) szemben, akik klinikai mintákat használtak. 21
22 A mikrorns mintázatok vizsgálatának másik oka a terápiás céllal felhasználható mikrornsek keresése. A megbetegedésekre jellemző expressziós mintázatokból következik, hogy gyakran több mikrorns mennyiségét kell befolyásolni (megnövelni vagy csökkenteni) a sikeres kezelés érdekében, valamint hogy önmagában nem lesz alkalmazható egyik kezelés sem. A terápiás kezelések egy lehetséges fajtája nem a korábban leírt, tumorsejtek mikrorns mintázatának befolyásolása, hanem az immunrendszer sejtjeinek stimulálása a mikrorns profiljuk változtatásával olyan irányba, mely a tumorszövet eliminációjához vezet. Az ilyen kezelések nagyon jelentős tudásanyagot kívánnak, hiszen egyszerre több helyen változtatjuk meg egy komplex rendszer működését. Természetesen itt is beláthatjuk, hogy mint az terápiák többségében, ebben az esetben sem egyedi kezelés, hanem kezelések kombinációja hozhatja meg a várt eredményt. A terápiás megoldásokat az is árnyalja, hogy a daganatok kialakulása sok esetben a már megismert esetekhez hasonlóan, de személyre szabottan történik, így a terápiának is legalábbis részben- személyre szabottnak kell lennie. 4. A mir-146a humán mikrorns Az epigenetikai szabályozási formák fontos szerepet töltenek be a gének megfelelő időben és helyen (sejttípusban) való kifejezésében és megváltozásuk bizonyítottan számos kóros folyamat hátterében áll. Munkám során egy, a humán genomban kódolt mikrorns epigenetikai szabályozását vizsgáltam. A mikrorns-ek a korábban leírtaknak megfelelően gének intronikus területein klaszterekben, vagy mint a mir-146a esetében is, a génről készülő a jelenleg ismert törvényszerűségek szerint az egyetlen lehetséges transzkript termékeként kódolódnak. Azt, hogy egy génről lehetséges-e mikrorns átíródása, termodinamikai összefüggések alapján, az mrns térszerkezeti vizsgálatát végző programok segítségével határozhatjuk meg. A mir-146a esetében a Jazdzewski és mtsai. által 2009-ben TargetRank software segítségével kimutatott térszerkezeti összefüggések szerint a mir-146a primer RNS átirata az 5. ábrán látható térszerkezeti formát veszi fel.
23 5. ábra mir-146a térszerkezete a termodinamikai összefüggések alapján (TargetRank software, (Jazdzewski és mtsai, 2009)). Létezik egy mikrorns, a mir-146b (10. kromoszóma), melynek szekvenciája csak néhány bázisban különbözik a 3 -felöli végén a mir-146a-tól (5. kromoszóma). A megkülönböztetésük a kis különbség miatt érett formában nehéz, de bizonyos módszerekkel (pl. Northern blot és microarray) megoldható. A szakirodalomban a mir- 146a működését vizsgálták behatóbban. A munkám során a két variáns elkülönítése a promóter aktivitás meghatározása esetében szükséges volt, hiszen a genom más területén kódolódnak, így a mennyiségbeli elkülönítésük fontos expressziós szinten az epigenetikai módosítások értékeléséhez. A mir-146b mikrorns által szabályozott folyamatok jelenleg még nincsenek rendesen tisztázva, sokszor átfednek a mir-146a eredményeivel, ami talán a nem megfelelő elkülönítésükből adódik. A mir-146a-nak számos sejttípusban központi jelentőséget nyilvánítanak, hiszen két nagyon fontos jelátviteli útvonalban (Toll-like és TNF receptor-hoz kapcsolt) a jelközvetítő faktorok mennyiségének szabályozását végzi. A mir-146a jelentőségét a 2. táblázatban láthatjuk összefoglalva, Rusca és Monticelli (2011) nyomán, kiegészítve. 23
24 2. táblázat A mir-146a funkciói és szabályozott faktorok egérben, emberben, valamint vírus fertőzött sejtekben (Rusca és Monticelli (2011) nyomán, kiegészítve. Expresszió/ funkció Szabályozott fehérje (a faktor mrns-e) Referencia Egér szöveti izolátumokban jelentős mértékű expresszió Th1 sejtekben, Th2 sejtekben pedig csökkent Monticelli és mtsai, 2005 hiányában csökkent Treg működés, IFNγ válasz csökkenését okozza STAT1 Lu és mtsai., 2010 az LPS indukálta IRAK1 mennyiségének csökkentése újszülötteknél, bakteriális hámszöveti károsító hatás elleni védelemben szerep IRAK1 Chassin és mtsai., 2010 Ember monocitákban csökkent erősségű TLR4 jelátvitel IRAK1, TRAF6 Taganov és mtsai., 2006 aktiváció indukálta sejthalál és IL-2 expresszió a Jurkat T-sejt vonalban, naív CD4+ és CD8+ T limfocitákban FADD Curtale és mtsai., 2013 monociták endotoxin (LPS) toleranciájában fontosság in vitro, TLR4- en keresztül IRAK1, TRAF6 Nahid és mtsai., 2009 tüdőepitélium sejtvonalban gátolja a gyulladási immunválaszt Perry és mtsai., 2008 megakariociták fejlődésének szabályozása CXCR4 Labbaye és mtsai., 2008 MDA-MB231 emlődaganat-sejtvonalban hatására csökkent migrációs és inváziós képessége IRAK1, TRAF6 Bhaumik és mtsai, 2008 transzfekció hatására sejtproliferáció fokozása méhnyakrák Wang és mtsai, 2008 tumor szupresszor szerep a hormonrezisztens prosztatadaganatban ROCK1 Lin és mtsai, 2008 hasnyálmirigy daganatban csökkenti az inváziós aktivitást EGFR, IRAK1 Li és mtsai., 2010 a pre-mir-146a-ban levő SNP hajlamosságot jelez a He és mtsai, 2005 pajzsmirigyrákra, expressziója fokozódik Vírus Egér makrofág sejtvonalban vizsgálva, VSV indukálja a mir-146a IRAK1, IRAK2, Hou és mtsai., 2009 expresszióját RIG-I és NFkB dependens módon TRAF6 EBV által kódolt LMP1 indukálja az expresszióját NFkB és esetleg AP-1 útján IRAK1, TRAF6 Cameron és mtsai., 2008 A mir-146a vizsgálatáról a szakirodalomban nagy mennyiségű adatot találhatunk, ezek alapján a különböző sejttípusokban betöltött szerepét és lehetséges jelátviteli szabályozásbeli fontosságát értékelhetjük. Fontos megjegyezni, hogy az emberi rendszerekben való vizsgálata sejtvonalakban történt, így fenntartásokkal lehet 24
25 csak az így megismert eredményeket az egészséges vagy a kóros szervezetben lejátszódó folyamatok megértésére alkalmazni. A korábbi vizsgálatok egér modellben kimutatták, hogy jelentős szerepe van a Th1 és Th2-es immunválasz polaritásának kialakulásában, hiszen nagy mennyiségben expresszálódik a Th1 típusú sejtekben, amíg a Th2-esekben nem (Monticelli és mtsai, 2005). További kutatások igazolták, hogy a helper T-sejteken kívül a Treg (regulátor) sejtekben is fontos szerepe van, hiszen a STAT1 transzkripciós faktor gátlásával segíti a Treg sejtek működését. A mir-146a KO egerekben az IFNγ válasz csökken, ami autoimmun betegséget okozhat (Lu és mtsai., 2010). Egy másik kutatócsoport újszülött egerekben vizsgálta a hámszöveti bakteriális fertőzésekben való szerepét. Kimutatták, hogy az LPS által kiváltott IRAK1 gátlása a mir-146a segítségével jelentős fontosságú az immuntolerancia kialakításában (Chassin és mtsai., 2010). Humán rendszerben jórészt sejtvonalban végzett vizsgálatok eredményeit összegezhetjük. A működés megértéséhez az in vitro megközelítésű, sejtvonalakban végzett kísérletek a könnyű kezelhetőség miatt megfelelőek, de a kórfolyamatokban lejátszódó folyamatokra ezek eredményeiből csak fenntartásokkal következtethetünk. A mir-146a-ról He és mtsai ben kimutatták, hogy a mir-221 és mir-222 mellett a pajzsmirigyrákos esetekben fokozódik az expressziója, diognasztikai markernek javasolták és jelezték, hogy a pre- mir-146a-ban levő SNP-k összefüggést mutatnak a daganatra való hajlammal. Taganov és mtsai. (2006) sejtvonalakban vizsgálva kimutatták, hogy a mir-146a hatására a monocitákban csökkent erősségű TLR4 jelátvitel figyelhető meg és ez a hatás az IRAK1 és a TRAF6 faktorok mennyiségének csökkenésével kapcsolható össze, melyek mrns-én megtalálható a mir-146a kötőhelye. A TLR4 jelátvitelének gátlása fontosnak bizonyult a monociták endotoxin toleranciájában is, melyeket in vitro kísérletekkel igazoltak Nahid és mtsai. (2009). A kísérletek során THP-1 sejteket LPS-sel indukáltak, és a TLR4-en lefutó jelátvitel válaszát vizsgálták mir-146a jelenlétében és hiányában (mir-146a inhibítort (komplementer oligót) transzfektáltak és az így kialakuló kétszálú, rövid RNS-t a rendszer, mint idegen RNS-t az RNS interferencia mechanizmusainak segítségével lebontja), igazolva, hogy jelenlétében a TNFα termelése csökken, így segítve az endotoxin (LPS) tolerancia kialakulását. Curtale és mtsai. (2013) Jurkat T-sejtvonalban, valamint naív CD4+ és CD8+ sejtekben kimutatták, hogy az NFkB és c-ets kötőhelyeken keresztül indukált mir-146a expressziója fontos az aktiváció indukálta 25
26 sejthalál szabályozásában, valószínűleg a FADD receptorhoz kapcsolódó útvonal gátlásával, valamint az AP-1 és IL-2 expresszió csökkentésével. Sejtvonalakban végzett kísérletek segítségével kimutatták, hogy a tüdőepitheliumban gátolja a gyulladási immunválasz kialakulását, valamint az IL-1ß gátolja az expresszióját az IL-8 és RANTES faktorok indukálásával (Perry és mtsai., 2008). A mir-146a-nak fontos szerepet tulajdonítanak még a megakariociták fejlődésében is, mivel sejtvonalakban a PLZF faktor mennyiségének növekedése in vitro csökkentette az expresszióját, miközben a CXCR4 mennyiségét növelte (Labbaye és mtsai., 2008). Daganatos sejtvonalakban számos csoport vizsgálta a szabályozásbeli jelentőségét, így MDA-MB231, emlődaganat sejtvonalban kimutatták, hogy az IRAK1 és TRAF6 faktorok szabályozásának segítségével csökkenti a migrációs és inváziós aktivitást (Bhaumik és mtsai., 2008). Méhnyakrák sejtvonalba transzfektálva sejtproliferáció fokozását érték el (Wang és mtsai., 2008), hormonrezisztens prosztatadaganatos sejtekben tumorszupresszor szerepet tulajdonítanak neki, hszen csökkent expressziós szintet mutat az egészséges szövethez képest, amíg a ROCK1 aktivitása jelentősen megnő (Lin és mtsai., 2008). További kísérletek kimutatták, hogy hasnyálmirigydaganat sejtvonalakban a fokozott expressziója csökkenti az inváziós aktivitást, valószínűleg az EGFR- IRAK1 útvonal gátlásával, míg a mennyiségi csökkenése ellentétes folyamatot indít el (Li és mtsai., 2010). Virális folyamatokban való jelentőségét a VSV (vezikuláris sztomatitisz vírus) és az EBV esetében végzett vizsgálatok mutatják. Hou és mtsai. (2009) egérmakrofág sejtvonalban kimutatták, hogy a VSV indukálja a mir-146a expresszióját RIG-I és NFkB dependens módon, ennek viszont a VSV replikáció csökkenése az eredménye. A szerzők valószínűsítik, hogy a folyamatban az IRAK1, TRAF6, valamint az IRAK2 mrns-e a mir-146a célmolekulája. Az EBV hatásáról az 5.2. fejezetben olvashatunk részletesebben, de összefoglalva kimondhatjuk, hogy EBV által kódolt LMP1 indukálja az expresszióját NFkB és esetleg AP-1 útján. Az LMP1 funkcionális TNF receptor homológ, az útvonalban negatív visszacsatolást figyelhetünk meg az NFkB- mir-146a- TRAF6 és IRAK1 vonalon, ahol az IRAK1 és a TRAF6 serkenti az NFkB-n keresztül a mir-146a expresszióját, mely negatívan visszahat az útvonalra, hiszen a TRAF6 és az IRAK1-et gátolja (Cameron és mtsai., 2008). 26
27 A fennt részletezett eredmények indokolják, hogy a mir-146a jelenlétét és hiányát befolyásoló folyamatokat (epigenetikai és jelátviteli) a szerepének tisztázása céljából fontos vizsgálni, hiszen ha tudjuk, milyen módon befolyásolható a mennyisége, akkor talán a terápiás és diagnosztikai célból való felhasználásához is közelebb kerülünk. 5. Az Epstein- Barr vírus Az Epstein-Barr vírus (EBV; humán herpesvírus 4) a Herpesviridae víruscsalád Gammaherpesvirinae alcsaládjának Lymphocryptovirus genusába tartozó kettősszálú DNS vírus. Mintegy 172Kb hosszúságú genomjának teljes szekvenciája ismert. Az emberi populáció több mint 90 %-a felnőtt koráig átesik az EBV fertőzésen. A gyermekkori fertőzések általában tünetmentesek, míg a serdülő- és a felnőttkori fertőzések gyakran mononucleosis infectiosa (MI) formájában jelentkeznek. Az EBVvel történő fertőződés orális kontaktus révén, illetve valószínűleg szexuális úton valósulhat meg, és elsősorban a B-sejteket és bizonyos típusú hámsejteket érint. A fertőződés a CR2/CD21 sejtfelszíni molekula és a virális glikoproteid gp350/220 kapcsolódásán keresztül jön létre. Fertőzést követően az emberi szervezetben a vírus látens formában egész életen át perzisztál, miközben mind látens, mind lítikus formában képes szaporodni. A vírus általában nem okoz betegséget, néhány kórkép kialakulásához azonban hozzájárulhat. Így valószínűleg központi szerepet játszik a Burkitt-lymphoma (BL), nasopharyngealis carcinoma (NPC), Hodgkin-kór (HD), továbbá az immunkárosodott betegek egyes lymphoproliferatív megbetegedéseinek pathogenezisében Az EBV faktorai és szerepük a génaktivitás szabályozásában Az EBV in vitro immortalizálja a fertőzött humán B-sejteket. A vírus látens ciklusa során majdnem 100 génje közül különböző kombinációkban mindössze a hat EBV nukleáris antigén (EBNA-1, -2, -3A, -3B, -3C, -LP), a három látens membrán protein (LMP1, -2A, -2B), az EBER RNS-ek, illetve a BamHI A régió RNS-ei expresszálódhatnak. A látens vírusfertőzés során az expressziós mintázat és az ennek hátterében álló eltérő promóter használat alapján három (I, II, III) látencia alaptípust 27
28 szoktak megkülönböztetni. A vírus látens ciklusai során számos olyan faktort expresszál, melyek több, az immortalizáció folyamatában kulcsfontosságú celluláris promótert aktiválhatnak különböző jelátviteli útvonalak aktivitásának szabályozásával. Egy ilyen faktor a II. és III. látencia típus során termelt LMP1. Az LMP1 fehérje hat transzmembrán doménből, valamint egy rövid N- terminális és egy hosszú C-terminális citoplazmatikus doménből áll (Kieff, 1996; Liebowitz és mtsai, 1986). Az LMP1 fehérje fontos szerepet játszik a B-sejtek immortalizációjában (Kaye és mtsai, 1993), valamint gátolja az epitheliális sejtek differenciálódását (Dawson és mtsai, 1990). B-sejtekben az LMP1 fehérje jelentős mértékben fokozza számos adhéziós fehérje, aktivációs marker és apoptózist gátló fehérje expresszióját (Bornkamm és Hammerschmidt, 2001). Epitheliális sejtekben az LMP1 a mátrix metalloproteináz (Yoshizaki és mtsai, 1998), a vascularis endotheliális növekedési faktor (Murono és mtsai, 2001) és az epidermális növekedési faktor receptor (Miller és mtsai, 1997) expresszióját indukálja. Az LMP1 fehérjék transzmembrán doménjeiken keresztül egymáshoz kapcsolódva lényegében ligand kötés nélkül is állandóan aktív receptornak felelnek meg (Gires és mtsai, 1997). Így a CD40 molekulához, illetve a tumor nekrózis faktor (TNF) receptor család más tagjaihoz hasonlóan az aggregált LMP1 fehérjék C-terminális citoplazmatikus doménjeihez TNF receptor asszociált faktorok (TRAF), illetve TNF receptor asszociált death domének (TRADD) kapcsolódnak, amelyeken keresztül az LMP1 aktiválni képes az NFkB-t (Izumi és Kieff, 1997; Kaye és mtsai, 1996). Az LMP1 fehérje C-terminális részén keresztül ugyanakkor más jelátviteli utakat is képes aktiválni, így például az LMP1 a két C-terminális aktivátor régió (CTAR) között található JAK-3 kötőhely révén STAT fehérjéket aktiválhat (Gires és mtsai, 1999), míg a CTAR-2 révén indukálhatja az AP-1- et (Kieser és mtsai, 1997). 28

29 5.2. Az EBV hatása a mir-146a expresszióra Az EBV által termelt fehérjék közül az LMP1 hatását mutatták ki a mir-146a expressziójának szabályozásában. Az aktiváló hatás a Toll-like receptorhoz kapcsolt NFkB jelátviteli útvonal fokozásával jelenik meg, hiszen az LMP1, mint CD40 homológ a C-terminális részén köti a TNF receptor asszociált faktorokat, így beindítva a jelátviteli útvonalat (6. ábra). A B 6. ábra A: az LMP1 C-terminális részéhez kapcsolódó jelátviteli útvonalak, Eliopoulos és Young (2001) nyomán. B: a mir-146a jelátviteli kapcsolatai (Nature Reviews, Drug Dicovery). A CTAR1 doménen keresztül indított jelátviteli útvonalon a TRAF6 és IRAK1 faktorok aktivitását fokozza, melyeknek hatása az NFkB aktiválása és ennek segítségével a mir-146a promóter expressziós aktivitásánnak fokozása (Cameron és mtsai., 2008). Ez a hatás sok esetben árnyalt, hiszen a mir-146a közvetlen célpontjai ezek a faktorok, így egy negatív visszacsatolás útján azoknak a fehérjéknek a mrnseiről induló transzlációt gátolja, melyek a mennyiségét növelik. 29
30 CÉLKITŰZÉSEK Munkám során a mir-146a mikrorns tumorkialakítási szerepének értékelése céljából a következő promóterszabályozási és irodalmi adatok jelentőségét értékelő vizsgálatokat végeztem el: (1) a mir-146a promóter aktivitásának jellemzése, a prekurzor és érett mir-146a formák kimutatása EBV pozitív sejtvonalakban, (2) a DNS metiláció szerepének és jelentőségének meghatározása a promóter szabályozásában, együtt értékelve a szakirodalmi adatokkal, (3) a mir-146a gén három különböző régióját érintő aktiváló hisztonmodifikációk szerepének meghatározása a promóter aktivitásának szabályozásában, (4) a DNS metiláció és a vizsgált hisztonmodifikációk együttes szabályozási szerepének értékelése a promóter aktivitásra, (5) az in silico módszerrel valószínűsített transzkripciós faktor kötések vizsgálata in vivo módszerrel a promóter területén, (6) az EBV szerepének értékelése a mir-146a transzkripció szabályozásában, (7) a DNS metiláció hatásának értékelése a c-myc és NFkB faktorok kötőhelyén és jelentősége a mir-146a promóter aktivitásának szabályozásában. A célul kitűzött vizsgálatok elvégzéséhez elengedhetetlen a gén vizsgált régióinak pontos szekvencia-meghatározása, így célom volt a sejtvonalankénti esetleges eltérések szekvenálás segítségével való kimutatása. A tervezett vizsgálatok segítenek a szakirodalomban megtalálható adatok és értékelésében, valamint új eredményeket adnak a mikrorns gének és a fehérje kódoló gének szabályozásának komplex mechanizmusáról. Ezek az eredmények reményeim szerint segítséget nyújtanak a mir-146a esetleges tumorkialakítási mechanizmusainak és a tumorkialakulásban betöltött szerepének megértéséhez, melyek újabb lépcsőfokokat jelenthetnek a terápiás vagy megelőző célú módszerek alkalmazásához. 30
31 ANYAGOK ÉS MÓDSZEREK 6. A vizsgálatok során felhasznált sejtvonalak és tenyésztésük A vizsgálatomban szereplő sejtvonalak típusát, érési stádiumát, EBV fertőzöttségüket, valamint az expressziós aktivitás szabályozásában fontos LMP1 jelenlétét az 2. táblázat tartalmazza. Valamennyi sejtvonalat 10% FCS-t (Fetal Calf Serum), 2mM glutamint, 50 egység/ml penicillint és 50 g/ml streptomycint tartalmazó RPMI 1640 médiumban, 5% CO 2 tartalom mellett 37 C-on szuszpenziós kultúrában tenyésztettem. 2. táblázat. A vizsgálatokban használt sejtvonalak ( a Bellan és mtsai., 2005; Piccaluga és mtsai., 2011; b Raji: EBNA-6 deléciós formáját tartalmazza az Epstein-Barr vírusnak). Sejtvonal Sejttípus és B-sejt EBV EBV látencia Referencia fejlődési állapot fertőzöttség típus (LMP1 expresszió) BL-41 Sporadikus, EBV - Lenoir és mtsai., 1985 BL-41- negatív BL; + (in vitro I (LMP1 Masucci és mtsai., 1987 E95B korai centroblaszt a fertőzött) negatív) BJA-B - Menezes és mtsai., 1975 DG-75 Ben-Bassat és mtsai., 1977 CBM1- Ral-STO Limfoblasztoid + III (LMP1 pozitív) Ernberg és mtsai., 1989; Altiok és mtsai., 1992 Rael Endémiás, EBV pozitív BL; + I (LMP1 negatív) Ernberg és mtsai., 1989; Altiok és mtsai., 1992 Akata késő germinális Takada és mtsai., 1991 Raji b központi B-sejt a Hatfull és mtsai., 1988 C II (LMP1 Cheung és mtsai., 1999 negatív) C2G6 + (MDA-MB- II (LMP1 Wakisaka és mtsai.,2004 C4A3 231 klón, EBVvel fertőzött) pozitív) II (LMP1 negatív) Wakisaka és mtsai.,
32 7. Kontroll PCR, szekvenálás A PCR reakciók tervezésénél elengedhetetlen a pontos szekvencia ismerete, a megfelelő primer kötések és specifikusságok eléréséhez. A szekvencia sejtvonalra specifikus ismerete nagyon fontos a metilációs térképek elkészítéséhez, hiszen CpG dinukleotidok eltűnhetnek, valamint ki is alakulhatnak deléciók, inszerciók és inverziók eredményeként. A szekvenálás során a mir-146a gén három régiójának bázisszintű meghatározását végeztem el, a szabályozó régiót, az első exon területét és a második exon azt a részét, amely az érett mikrorns szekvenciáját tartalmazza. 3. táblázat A PCR reakció során használt primerek a kontroll szekvenáláshoz. Az egy cellában levő primerek párosíthatók, F= Forward, R= reverse (egy F és egy R primer egy pár, esetenként az egyik tag egy másik szembe levő primerrel is párosítható, ld.: három primert tartalmazó cella). Gén régió Szabályozó régió és első exon Második exon Primer szekvenciája F1: 5 -GGCAGAGTAATGCAGTGTACCAG-3 R2: 5 -CCCTAATCCCACAGCAATCATC-3 F10: 5 -CCTCAATTCAAATCCTGGCTCTG-3 F3: 5 -GGCACCCATGGAAACACTTAC-3 R4: 5 -CCAGGCTTCCTGTCTGTACTCTC-3 F6: 5 -GGTGCTCAGGAAGATTTCTCAGT-3 R5: 5 -CCCTCTTGCAGCACGTGTCAGG-3 F8: 5 -CCTGACACGTGCTGCAAGAG-3 R7: 5 -CAGCTAAGGCCTATTTGCATATG-3 R9: 5 -GTGCCATTTACTCTGCAATAATC-3 F11: 5 -GGACAGGCCTGGACTGCAAC-3 R12: 5 -TAGCTACTTGGAACCCTGCTTAGC-3 A módszer során a PCR-rel (polimerase chain reaction) amplifikált, és tisztított DNS-ből 10µl-t, 1,5µl dh 2 O-t, 0,5µl primert (10pmol/µl) és 8µl DYEnamic TM ET dye terminator (MegaBACE TM ) reagens premixet (összesen 20µl) mértem mintánként PCR csőbe, majd PCR gépben elvégeztem a szekvenálási reakciót. A program minden esetben 95 o C 20sec egy ismétléssel és 95 o C 20sec, 58 o C 15sec, 72 o C 1 perc 30 ciklusszámmal futott. A szekvenálási termék tisztítása AutoSeq TM G-50 (Amersham 32
33 Biosciences) oszlopon, a termékhez megadott protokoll alapján vagy abs. etanolos kicsapással történt. A tisztított terméket MegaBACE TM kapilláris szekvenátorban futtattam meg, majd MegaBACE TM Sequence Analyser program segítségével értékeltem. A szekvenáláshoz használt primereket a 3. táblázat tartalmazza. 8. Expressziós vizsgálatok 8.1. A mir-146a éretlen alakjának kimutatása, módszertani különlegességek A Nuclear run-on módszere lehetőséget nyújt az újonnan készülő RNS-ek mennyiségi meghatározására. Az epigenetikai vizsgálatok eredményeinek értékelésénél elengedhetetlen a pontos promóter aktivitás ismerete, mivel ennek tudatában van jelentése a különböző aktiváló vagy inaktiváló módosításoknak. Bizonyos esetekben nem alkalmazható a hagyományos megközelítés, tehát a random vagy specifikus primerrel végzett reverz transzkripció, így a mikrorns-ek esetében sem, mivel a mikrorns-ek sajátosságából adódóan maga az elsődleges transzkriptum használódik el az érési folyamat során, ellentétben a hírvivő RNS-ekkel, amelyeknél a kiindulási génre jellemző féléletidő és/ vagy génaktivitás szerint megtalálható RNS transzkriptumról készül a fehérje termék. Ha pontos promóter aktivitást szeretnénk meghatározni a mikrorns-ek esetében egy olyan modszerre van szükségünk, mely minden érési stádium RNS mennyiségét összesítve tud eredményt adni. A Nuclear run-on módszer a legmegfelelőbb erre a feladatra különösen, hogy az állati sejtrendszerek esetében a mikrorns érés két jól meghatározható részre bontható fel, melyek térben, fizikailag a sejtmagmembrán által el vannak választva. Ennek eredményeként a módszer során a sejtmagizolálás következtében az érési folyamat megreked a pre-mikrorns állapotban és így a teljes, újonnan szintetizált RNS mennyisége az arra tervezett reverz transzkripciós rendszerrel kvantitatív módon kimutatható, meghatározható. A módszert változtatásokkal Patrone és mtsai. (2000) által közölt módon végeztem, módosításokkal. A munkafolyamat vázlatát az 7. ábra mutatja be. 33
 A módszert megelőző sejtmagtisztítás a következő protokol alapján történt: 5x10 7 db sejtet 1x PBS-sel mostam kétszer, a mosások között 4 o C, 270g-vel centrifugáltam.")
34 7. ábra A Nuclear run-on és az azt követő Real-time PCR folyamata (leírást ld. a szövegben) A módszert megelőző sejtmagtisztítás a következő protokol alapján történt: 5x10 7 db sejtet 1x PBS-sel mostam kétszer, a mosások között 4 o C, 270g-vel centrifugáltam. A sejtüledéket 4ml sejt lízis oldatban vettem fel (10mM Tris-HCl ph7,4, 3mM MgCl 2, 10mM NaCl, 150mM Sucrose és 0,5% NP-40) és jégkásán inkubáltam 5 percig. A sejtmagokat 4 o C, 170g-n való centrifugálással leültettem és sejt lízis pufferben (NP40 néküli) mostam, a cetrifugálás után a sejtmagokat 150µl Storage pufferben vettem fel és frissen (de tárolható -150 o C-on) a Nuclear run-on reakcióhoz használtam fel. A mintákhoz 150µl 2x transzkripciós puffert (200mM KCl, 20mM Tris- HCl ph8, 5mM MgCl 2, 200mM Sucrose, 20% Glicerol), 1,6µl 50mM biotin jelölt UTPt (biotin-16-utp) adtam és 30 percig 29 o C-on inkubáltam. A reakciót 6µl 250mM CaCl 2 és 60 egység (U) RNáz mentes DNaseI hozzáadásával állítottam le. RNS-t utána TRI-reagens (Sigma) segítségével tisztítottam (a gyártó ajánlása szerint) és 120µl vízben vettem fel. A tisztított RNS-t az újonnan szintetizálódott RNS izolálása érdekében biotin kötő streptavidines gyöngy segítségével tisztítottam a következő módon: 120µl mintához 120µl gyöngyöt (70µl (Dynabead, Invitrogen) gyöngyöt kétszer mostam 1x BW pufferben, végül 120µl 1xBW-ben felvéve) adtam és 37 o C 2 órán keresztül inkubáltam. A biotin jelölt RNS-ek streptavidinre való kikötődése után 15% formamidot tartalmazó 1x SSC pufferben mostam a gyöngyöt kétszer, majd egyszer 2x SSC-ben és ezután 30µl RNáz mentes vízben vettem fel és a Real-time PCR vizsgálatig -150 o C-on tároltam. 34
35 A Real-time PCR reakció során a 4. táblázatban található primereket használtam, melyek az érett mikrorns szekvenciája köré (pre-mikrorns-nek megfelelő terület) lettek tervezve, hiszen a sejtmagi érés során ez a régió, ameddig eljut az útvonal. A PCR reakció a következő körülménnyekkel futott: 95 o C 10sec egy ismétléssel és 95 o C 1sec, 63 o C 5sec, 72 o C 7sec 50 ciklusszámmal futott, a reakció során LightCycler 480 gépet (Roche) használtam, valamint LightCycler 480 DNA SYBR Green I Master kitet. Izolátumok közötti kontrollként a GAPDH (glyceraldehyde-3-phosphate dehydrogenase) háztartási génre tervezett rendszert használtam, a reakció a mir-146a esetében leírtakkal volt megegyező. 4. táblázat A Nuclear run-on módszer során használt real-time PCR primerek. mir-146a F: 5 -ctc ctc gtt gtg cta ctg tct c-3 R: 5 -gat gct ctt tct cca aga cg-3 GAPDH F: 5 - gga agg tga agg tcg gagtca-3 R: 5 - atg ggt gga atc ata ttg gaaca A mir-146a érett alakjának kimutatása Az érett mikrorns mennyiségi meghatározásához kisrns Northern blot módszert használtam. A Northern blot módszer során elektroforetikus elválasztás után blot segítségével visszük át a gélről (futtató és elválasztó közeg) a membránra az RNS molekuláinkat. Az általam használt módszer esetében a bázisonkénti elválasztás érdekében 12%-os denaturáló lineárispoliakrilamid gélt használtam. A módszert Szittya és mtsai. (2002) és Csorba és mtsai. (2007) alapján végeztem, kiegészítésekkel. A sejtekből Tri-reagens (Sigma) segítségével teljes RNS-t tisztítottam és felhasználásig -150 o C-on tároltam. A kontroll mir-146a és b RNS mintákat a (Metabion) a sejtvonal minták mellett futtattam, így kontrollálva a várt mikrorns nagyságot, valamint meghatározva a mir-146a és b forma elkülöníthetőségét. A mintákat 12% denaturáló poliakrilamid (10ml végtérfogatban: 4,5g urea, 1ml 10xTBE, 3ml akrilamid (40%-os: 38 rész akrilamid: 2 rész bisz-akrilamid), 10ml-re dh 2 O-val kiegészíteni, használat előtt 6µl APS (ammonium-perszulfát)) gélelektroforézissel 35
36 (PAGE) választottam szét méret (töltöttség) alapján. Futtatás után a gélt az urea kimosása érdekében 1xSSPE oldatban áztattam, majd Hybold-N nylon membránra blottoltam át ugynevezett nedves blottolás módszerével. A membránra UV keresztkötés módszerével kötöttem ki a blottolt mintákat. A membránt kisrns pufferben (50% formamid, 5xSSPE (1xSSPE: 0,115M NaCl, 10mM nátrium-foszfát és 1mM EDTA, ph 7,4), 5xDenhardt s oldat (1xDenhardt s: 0,02% Ficoll, 0,02% poli-vinil-pirrolidon és 0,02% BSA), 0,5% SDS és 0,02 mg/ml heringsperma DNS)) előhibridizáltattam, majd hibridizáltattam. A hibridizálás radioaktivan jelölt LNA (Locked nucleid acid) próbákkal történt (jelölési reakció: 1µl 10pmolos oligo, 1µl PNK (T4 polinukleotid kináz) puffer, 1µl [γ-32p]-datp, 1µl PNK enzim, 10µl-re dh 2 O-val kiegészíteni). Inkubálás 37 o C-on egy óra. Hibridizálás 42 o C-on 12 óra. Ezt követően a membránt 1x SSC (0.15 M NaCl, 15mM Na-citrát, ph7.2), 0,1% SDS oldattal 2x 10 percig mostam, majd a jel erősségétől függően megfelelő ideig exponáltam (fél óra- 12 óra). A radioaktív jeleket Storm 840 szkennerrel (Amersham Biosciences) detektáltam, és ImageQuant szoftverrel értékeltem. A vizsgálat során a mir-146a és b forma elkülöníthetősége érdekében a minták mellé mindkét forma szintetizált érett alakját kontrollként futtattam, valamint a jelölést a specifikusság vizsgálata céljából mind az a, mind a b formára specifikus LNA oligóval elvégeztem. A sejtek (izolátumok) közötti kontrollként a spliceoszóma (az RNS-ek elsődleges transzkriptjének érését végző kis nukleáris RNS-ekből és járulékos fehérjékből álló komplex) U6 RNS-ére tervezett próbát alkalmaztam. Az oligók szekvenciáját az 5. táblázat tartalmazza. 5. táblázat A Northern-blot módszer során használt oligók. Felhasznált LNA oligo szekvenciák Kontroll U6 próba szekvenciája Szintetizált kontroll RNS szekvenciák mir-146a: 5 -aacccatggaattcagttctca-3 mir-146b: 5 -agcctatggaattcagttctca-3 5 -gca ggg gcc atg cta atc ttc tct gta tcg-3 mir-146a: 5 -cuu uga gaa cug aau ucc aug ggu u- 3 mir-146b: 5 -cug aga acu gaa uuc cau agg cug-3 36

37 9. DNS metilációs vizsgálat A biszulfit modifikálás módszere lehetőséget nyújt arra, hogy egy ismert szekvenciában kémiai reakcióval megkülönböztethetővé tegyük a metilált és nem metilált citozinokat. A modifikálási reakció során ugyanis a nem metilált citozinok uracillá alakulnak, amíg a Na-biszulfittal nem reagáló 5-metil-citozinok citozinok maradnak. Az ezt követő PCR-reakciókban az uracilok timinként fognak megjelenni, míg az 5 m C-ok természetesen citozinok maradnak. Ebből következően, a szekvenálást követő szekvencia analízis során az ismert eredeti szekvenciával összehasonlítva megállapítható, hogy melyik citozinok voltak metiláltak, és a metiláció a tisztított DNSpopuláció hozzávetőleg mekkora hányadában fordult elő. A módszer lényegét a 8. ábrán vázoltam fel. 8. ábra A DNS-metilációs vizsgálat folyamta. T= timin, C= citozn, G= guanin, A= adenin bázisok, U= uracil, C m = metilált citozin, M13= az M13 fág szekvencia darabja, Biotin= biotin jelölés (ld. szöveg) A modifikálási reakciót kisebb módosításokkal a Frommer és mtsai. (1992) illetve Clark és mtsai. (1994) által leírt módon végeztem. 5 g DNS-t 50 l steril 37
38 desztillált vízben oldottam, majd hozzáadtam 5,5 l frissen készített 3M NaOH-t és az elegyet 37 C-on 15 percig inkubáltam. Ezután a denaturált DNS-hez 30,5 l frissen készített 10mM Hidrokinon (Sigma) oldatot és 530 l 3,6M Na-biszulfit (Sigma) oldatot (ph5.0) adtam. Az elegyet óvatosan összekevertem majd 200 l térfogatú PCR csövekbe osztottam. A modifikálási reakciót (3 perc 95 C, 57 perc 55 C, 5 ciklusszámmal) PCR gépben végeztem el. A ciklikus modifikálást követően a Na-biszulfitot tartalmazó DNS oldathoz 6 l GeneClean üveggyöngyöt (BIO 101, MP Biotech) adtam. Öt perc szobahőmérsékleten történő inkubálást követően az elegyet 5000g-vel 30 másodpercig centrifugáltam, a felülúszót eltávolítottam, majd a csapadékot 500 l GeneClean mosófolyadékkal (BIO 101, MP Biotech) mostam. Ezt a tisztítási lépést ezután még 2- szer megismételtem, majd az üveggyöngyön megkötött DNS-t 50 l steril desztillált vízben 3 percig 50 C-on eluáltam. Az eluátumhoz ezután 11 l 3M NaOH-t adtam, majd 15 percig 37 C-on inkubáltam, végül a DNS-t 66 l 3M ammónium-acetát és 720 l abs. etanol hozzáadásával -20 C-on egy éjszakán át precipitáltam. Az ezt követő 20 perces, 4 C-on, 12000g-vel történő centrifugálás után a csapadékot 90 l 70%-os etanollal mostam, majd 20 percig 12000g-vel 4 C-on centrifugáltam, végül szobahőmérsékleten szárítottam. A modifikált DNS-t 50 l steril desztillált vízben oldottam fel. A modifikálást követően a vizsgálni kívánt PCR reakciókat fészkes PCR reakciók segítségével amplifikáltam. Az amplifikációk során alkalmazott primereket az mir-146a-promóter régiók esetében az 6. táblázat tartalmazza. 6. táblázat A mir-146a promóter metilációs analíziséhez használt primerek Primer neve és sorrendje mir-146a külső Primer szekvenciája F: 5 -GTAGAGTGTTTGGTGTTTAGTAGGTG-3 R: 5 -CCATTTACTCTACAATAATCCACTATAC-3 mir-146a belső F1: 5 -Uni-GAGAAAGGTGTTTAGGAAGATTTTTTAG-3 F2: 5 -Uni-GAGAGTATAGATAGGAAGTTTGG-3 F3: 5 -Uni-GAGGTTTTGGTTGAAATTTAGTTTG-3 F4: 5 -Uni-GGAATAAAAGTATATGTAAATAGG-3 F5: 5 -Uni-GAGAAGTTGATATTGTTAGGTTGG-3 R6: 5 -Biotin-CCACTATACAAAAACCTACTACCTCTC-3 38
39 A fészkes PCR első reakciója 50 l végtérfogatban 1 l, steril desztillált vízben oldott modifikált és tisztított DNS-t, 1-1 l 0,2mM négy fajta dntp-t és mindkét szálspecifikus, 1-1 l 40pmol-os külső primert, 0,5 l Fast Start Taq polimerázt (Roche), valamint az enzimnek megfelelő 5-5 l 10x puffert és MgCl 2 -ot tartalmazott. Az amplifikálás körülményei a metilációs analízishez használt primerek esetében az alábbi volt: 95 C 40 s, 56 C 40 s, 72 C 70 s, harmincöt ciklus. Az első PCR reakció alatt a modifikálás során uracillá átalakult metilálatlan citozinok timinként, míg a változatlanul maradt metilált citozinok citozinként amplifikálódtak. A PCR-t minden esetben 2,5 perces 95 C-os denaturálással kezdtem és az utolsó ciklus után a reakciókat még 8 percig 72 C-on tartottam. Az első PCRterméket 100-szorosára hígítottam és a második PCR reakcióhoz ebből a hígításból használtam 1 l-t. A második PCR reakciókban alkalmazott primer-párok közül az egyik primer 5 végére biotint, míg a másik primer 5 végéhez az M13 Universal primer (uni.) 17 nukleotidját (5 -fluorescein-gtaaaacgacggccagt-3 ) szintetizáltattam. Így lehetővé vált minden régió PCR termékének szekvenálása egyetlen fluoreszcensen jelölt primerrel, így a mennyiségi összehasonlítás nagyobb pontossággal volt elvégezhető. A második PCR-ek reakcióelegye mindkét szál-specifikus, belső primerből 2pmol-t tartalmazott, a többi összetevő koncentrációja megegyezett az első PCR-ekével. A PCRek hatékonyságát 5 l-nyi PCR termék 2%-os agaróz gélen (1xTAE puffer futtató közegben) történő elektroforetikus elválasztásával ellenőriztem. 10. Kromatin immunprecipitáció és azt követő real-time PCR A hiszton módosítások a DNS metilációval együtt jelentős, szigorúan szabályzott, komplex folyamatok eredményeként alakulnak ki, összefüggéseik a mai napig sok kutatás témái, új adatok megismerése fontos a helyes értékelésükhöz, valamint elengedhetetlen a promóter aktivitását szabályozó folyamatok meghatározásához, vizsgálatuk kromatin immunprecipitációval (ChIP) történik. 39
40 A kromatin immunprecipitáció módszerrel az in vivo DNS-fehérje interakciók kimutatására van lehetőségünk. Az eljárás során kémiai reakcióval kovalensen keresztkötjük az adott sejtvonalban a DNS-hez kapcsolódó fehérjéket. Majd az izolált DNS-t bp-os darabokra törjük. A vizsgálni kívánt fehérjére specifikus ellenanyaggal, és az ellenanyag Fc-részéhez kapcsolódó Protein A gyöngyökkel kicsapjuk a teljes DNS állományból a fehérjénket, és vele együtt a kovalensen keresztkötött DNS fragmentumokat. Ezek után lehetőségünk van a kinyert DNS darabok azonosítására, és így annak megállapítására, hogy a vizsgált fehérje milyen mértékben kapcsolódott in vivo egy adott szekvenciához. A ChIP módszer lényegét a 9. ábrán vázoltam fel. A Kromatin Immunprecipitációt kisebb módosításokkal a Farnham és mtsai. (2002) által közölt metodika alapján végeztem. 9. ábra Kromatin Immunprecipitáció folyamata (magyarázat: ld. szöveg) 40
41 Kiindulási sejtvonalakat felnövesztettem, hogy a kiindulási sejtszám elérje a t. A sejteket tartalmazó médium 44ml-ét 50ml-es falkon csövekbe vittem át, 1,22ml 37%-os formaldehidet (Sigma) mértem hozzá, majd 10 percig szobahőmérsékleten kevergetéssel inkubáltam. Az inkubálás során kialakultak a DNS és az ahhoz in vivo kötődő fehérjék közötti stabil kovalens kötések. A reakciót 2,38ml 2,5M-os glicinnel állítottam le, hogy az aspecifikus kötések kialakulását megakadályozzam. 5 perces 4 C 2500rpm-en történő centrifugálást követően a mintákat 5ml jéghideg PBS-ben mostam, majd centrifugáltam és a pellethez 1ml PBS-et adtam. 5 perces 4 C 4000rpm-en történő centrifugálás után 50ml-ben 1db proteáz inhibitor tablettát (Roche) tartalmazó Sejt lízis pufferből (5mM PIPES ph8.0; 85mM KCl; 0.5% NP40) 1ml-t hozzáadva szuszpendáltam fel a mintákat. Miután 5 percig jégkásán inkubáltam, majd 4000rpm-en 5 percig 4 C on centrifugáltam a sejtlizátumot, a pelletet újra 1ml sejtlízis pufferben vettem fel és megint inkubáltam 5 percig. Ezután 50ml térfogatban 1db proteáz inhibítor tablettát tartalmazó Nukleusz lízis puffer (50mM Tris-Cl ph8.1; 10mM EDTA; 1% SDS) 2ml-es térfogatában vettem fel. A mintákat 5 percig jégkásán inkubáltam, ezt követően szonikáltam. Ezután 13000rpm-en 5 percig 4 o C-on centrifugáltam a mintákat, majd a felülúszót új 1,5ml-es Eppendorf csövekbe vittem át. A szonikált lizátum 100μl-ét 6μl 5M NaCl és 2μl RNáz hozzáadása után 65 Con inkubáltam 4 órán keresztül a DNS-fehérje kapcsolatok megszüntetéséért, majd 4μl ProteinázK (20mg/ml) hozzáadása után a DNS-t tisztítottam. A kapott DNS-t 2%-os agaróz gélen megfuttatva vizsgáltam a szonikálás minőségét. Azt a lizátumot használtam a következő vizsgálataimhoz, amelyik bp nagyságúra törött. A felülúszó 100μl-éhez 900μl IP dilúciós puffert (0,01% SDS, 1,1% Triton X- 100, 1,2mM EDTA, 16,7mM Tris-HCl ph8,0, 167mM NaCl) adtam, majd 20μl 10μg/μl-es szonikált hering sperma DNS-t, 20μl 10mg/ml-es trns-t, valamint 80μl, előzőleg elkészített, aktivált 50%-os protein A-agaróz gyöngyöt mértem hozzá. (Az 50%-os protein A-agaróz gyöngyöt liofilizált protein A-agaróz gyöngy (Sigma) TE pufferben (10mM Tris-HCl ph8,1mm EDTA) történő duzzasztása, és TE-vel végzett négyszeri mosása, majd a lecentrifugált gyöngy térfogatával egyenlő térfogat TE-ben való szuszpendálása után, 1/25-öd végtérfogatú szonikált hering sperma DNS, és 1/10 végtérfogatú 1mg/ml végkoncentrációjú BSA hozzáadásával nyerjük). 41
42 30 percig 4 o C-on forgó állványon inkubáltam. Ezzel a lépéssel előtisztítottam a kromatint. Ezután 14000rpm-en 4 o C-on 5 percig centrifugáltuk a mintákat, majd ezt követően a felülúszót új Eppendorfokba vittem át. A térfogatot két egyenlő részre osztottam, felülúszót elfeleztem, az egyik feléhez 2μl 1μg/μl koncentrációjú, a vizsgált hisztonmodifikáció elleni specifikus ellenanyagot adtam, a másik feléhez pedig 2μl 1μg/μl immunizálatlan nyúl széruma ellen termeltetett ellenanyagot adtam (7. táblázat), így kontrollként használtam, majd mindkét mintát vortexeltem és 4 C-on egy éjszakán keresztül inkubáltam. 7. táblázat A kromatin immunprecipitáció módszer során használt ellenanyagok Ellenanyag megnevezése Anti-acetyl-Histone H3 Anti-acetyl-Histone H4 Normal rabbit IgG (kontroll) Anti-dimethyl- Histone H3 (Lys4) Ellenanyag típusa nyúlban termeltetett, poliklonális IgG, (Upstate) nyúlban termeltetett, poliklonális IgG, (Upstate) nyúlban termeltetett, poliklonális IgG, (Santa-Cruz Biotechnology) nyúlban termeltetett, poliklonális IgG, (Upstate) Ellenanyag katalógusszáma SC A 4 o C-os egy éjszakás inkubációs idő letelte után 30μl aktivált 50%-os protein A- agaróz gyöngyöt adtam a mintákhoz, ezután 30 percig rázattam szobahőmérsékleten. Ez idő alatt a kovalens kötésekkel stabilizált kérdéses fehérje/dns-fragmentum komplexeket specifikusan felismerő ellenanyagok Fc részükkel hozzákötődtek a sejtek felszínén lévő protein A molekulákhoz, így a teljes kromatint tartalmazó mintából azok a DNS darabok, amelyekhez a vizsgált fehérje (az adott hiszton vizsgálni kívánt modifikációs alakja) in vivo kötődik, precipitálással elválaszthatóvá váltak. Ezt 10 perces 4 o C-on, 4000rpm-en történő centrifugálással tettem meg. Az kontroll minta felülúszóját, mint teljes bevitt kromatint (TIC) tettem el, mivel ez tartalmazza a teljes bevitt DNS mennyiséget, pozitív kontrollként használhattam. 42
43 A kapott üledéket (pelletet) egyszer mostam 1ml Low Salt pufferben (0.1% SDS; 1% Triton X 100; 2mM EDTA; 20mM Tris-Cl ph8; 150mM NaCl), egyszer High Salt pufferben (0.1% SDS; 1% Triton X 100; 2mM EDTA; 20mM Tris-Cl ph8; 500mM NaCl), és kétszer mostam LiCl detergent pufferben (10mM Tris-Cl ph9.0; 1mM EDTA; 250mM LiCl; 1% IGEPAL; 1% deoxikólsav). A mosások között 10 percig inkubáltam a mintákat, majd 4000 rpm-en 1 percig centrifugáltam. A mosásokat követően inkubációk nélkül kétszer mostam 1ml TE pufferben. A mintákat 250 l Elúciós pufferben (0,1M NaHCO 3 ; 1%SDS) 15 percig szobahőmérsékleten forgattam. Eközben az ellenanyag/fehérje/dns komplexek leváltak a gyöngyről. A mintákat 4000rpm-en 10 percig centrifugáltam, majd a felülúszókat új Eppendorf csövekbe vittem át. Ezt még egyszer megismételtem, így a végtérfogat 500μl lett. A leírt folyamatok során kapott 500μl térfogatú oldatokhoz és a TIC-hez (pozitív kontroll) is 20 l NaCl-ot mértem. A fehérje és a DNS molekulák között létrehozott kovalens kötéseket 4-5 órás 65 o C-on történő inkubálással szüntettem meg. Ezután a lizátum minőség-vizsgálatánál megismert módszerrel DNS-t tisztítottam. A kinyert DNS-t 30μl steril desztillált vízben vettem fel. A DNS-t real-time PCR segítségével vizsgáltam. A reakciót a régió három területére terveztem: promóter régió, az iniciációs pont és első exon területe, valamint az első intron területére. Mindhárom primerszett esetében a reakció a következő volt: 95 o C 10sec egy ismétléssel és 95 o C 5sec, 62 o C 5sec, 72 o C 8sec, 50 ciklusszámmal futott. A vizsgálat során használt primereket a 8. táblázat tartalmazza. A vizsgált régiókat és a primer pozíciókat a Melléklet fejezetben megtalálható szekvencia tartalmazza a megértést segítendő. 8. táblázat A kromatin immunprecipitációt követő real-time PCR során használt primerek. mir-146a szabályozó régió F: 5'-GTGACTACATCTGCCTGGAAGC-3' R: 5'-CAAACTGGAAAGAACGTACAAGG-3' mir-146a iniciációs régió és első exon mir-146a első intron F: 5'-CATATGCAAATAGGCCTTAGCTG-3' R: 5'-TGTCTCCTGTCCATCCTGTCC-3' F: 5'-GTGACTTGGAGATATTTCCAGTGC-3' R: 5'-CAGTCTACAACTCCAGGTTTAAAGC-3' 43
44 11. DMS in vivo footprinting A promóter aktivitást meghatározó transzkripciós faktorok vizsgálata elengedhetetlen az epigenetikai eredmények értékeléséhez, hiszen a transzkripciós szabályozást a két rendszer komplex működése teszi lehetővé, egymástól nem elválaszthatók, külön csak fenntartásokkal értékelhetők. A DNS-fehérje kapcsolatok vizsgálatára a DMS in vivo footprinting módszert használtam. A vizsgálni kívánt faktorkötéseket és a promóteren megtalálható kötőhelyeiket a Melléklet című fejezetben levő szekvencia tartalmazza a megértést segítendő. Az Eredmények fejezetben a kiválasztásuk oka részletesen van tárgyalva. A módszert a már korábban leírt módon (Mueller és Wold, 1989, Garrity és Wold, 1992) kisebb módosításokkal (Niller és mtsai., 1995, Szenthe és mtsai, 2013c) végeztem el. A vizsgálat menetét az 10. ábrán vázoltam fel. 10. ábra DMS in vivo footprinting folyamata. A DMS kezelés hatására a fehérjék kötődésétől függően bizonyos pozíciókban gyengébben (M) máshol erősebben (M) metilálódnak a guaninok. A piperidin kezelés hatására a DNS a metilált guaninoknál törik. Az így keletkezett heterogén DNS populáció ismeretlen szabad végeit ligáció mediálta PCR (LM-PCR) segítségével amplifikáljuk, majd poliakrilamid gélelektroforézis segítségével vizualizáljuk. Sötétkék folt: fehérje, G: guanin, M: metiláció, piros vízszintes vonal: primer, vízszintes zöld vonal: DNS szál, Függőleges zöld vonal: DNS band poliakrilamid gélen, radioaktivan jelölve. Zöld nyilak: DMS kezelés által érintett bázisok (guaninok) (a nyíl vastagsága a kezelés fehérjekötéstől függő gyakoriságát jelöli). 44
45 Minden egyes vizsgálathoz 10 7 sejtet 1500g-vel 10 percig centrifugáltam, 1xPBS-el mostam majd a sejteket 1ml 1xPBS-ben szuszpendáltam. Ezután a sejtszuszpenzióhoz 5 l 99%-os dimetil szulfátot (DMS) adtam és óvatos keverés után szobahőmérsékleten 1 percig inkubáltam, minden egyéb időszakban jégkásán tartottam. A reakciót 5ml DMS stop oldat I (1xPBS-ben oldott 1%-os BSA, 100 M β- merkapto-etanol) hozzáadásával állítottam le. A sejteket ezután egyszer a fenti DMSstop oldattal, majd kétszer 1xPBS-el mostam, végül 1ml 1xPBS-ben vettük fel és a sejtekből ún. footprintelt DNS-t izoláltam. A sejtszuszpenzió 1 ml-éhez 10 ml extrakciós puffert (10mM Tris-HCl (ph8.0), 0.1M EDTA (ph8.0), 20 g/ml RNAáz, 0.5% SDS) adtam és 1 órán át 37 o C-on inkubáltam. Az elegyhez ezután 100 g/ml végkoncentrációban Proteináz K-t (Roche) adtam, és 3 órán át 50 o C-on inkubáltam. Az elegyet ezután szobahőmérsékletűre hűtöttem, és azonos térfogatú 0.5M Tris-HCl (ph8.0)-val telített fenolt mértem hozzá, majd 10 perces óvatos keverést követően 15 percig szobahőmérsékleten 5000g-vel centrifugáltam. A felső vizes fázist ezután egy másik csőbe átpipettáztam, és a fenti fenolos tisztítást még kétszer megismételtem. A felső vizes fázis tisztítását a fenolos tisztításhoz hasonló módon ezután azonos térfogatú kloroformmal is elvégeztem. Ezt követően a vizes fázishoz 0.2 térfogat 10M ammónium acetátot és 2 térfogat abszolút etanolt adtam, óvatosan összekevertem, majd 5 percig szobahőmérsékleten 5000g-vel centrifugáltam. Az így kapott DNS csapadékot ezután 70%-os etanollal 2x mostam, majd szobahőmérsékleten szárítottam. Végül steril desztillált vízben úgy oldottam fel, hogy a DNS végkoncentrációja 1 g/ l legyen. A DNS oldat tisztaságát és koncentrációját a 260 és 280nm-es hullámhosszúságon mért abszorbancia segítségével állapítottam meg. Ezzel egyidejűleg a kontroll G-reakcióhoz a sejtek egy részéből DMS kezelés nélkül izoláltam DNS-t, melynek lényege, hogy a DNS-hez kötő fehérjék így nem lesznek jelen ennél a DNS-nél. 100 l (1 g/ l koncentrációjú) kontroll DNS-hez ezután 100 l PBS-t adtam, majd az elegyet 1 l DMS-el, rövid vortexelést követően 1 percig szobahőmérsékleten inkubáltam. A reakciót 50 l jéghideg DMS Stop oldat II (1,5M nátrium-acetát (ph7.0), 1M β-merkapto-etanol) hozzáadásával állítottam le. Ezt követően 750 l abs. etanolt adtam az elegyhez, majd 20 percig 12000g-vel 4 C-on centrifugáltam. A felülúszó eltávolítása után a csapadékhoz 250 l 0,3M nátrium- 45
46 acetátot (ph5.2) és 750 l abs. etanolt adtam és rövid vortexelést követően 20 percig 12000g-vel 4 C-on centrifugáltam. A csapadékot ezután kétszer 800 l 70%-os etanollal mostam, majd vákuum alatt szárítottam és 180 l steril desztillált vízben oldottam. 180 l DMS-sel kezelt DNS oldatokhoz ezután jégkásán 20 l 10M piperidint (Fluka) adtam és rövid vortexelést követően 30 percig 90 C-on inkubáltam. Ezután az oldatot azonnal jégkására helyezve lehűtöttem majd vákuum alatt kiszárítottam. Kétszer 50 l steril desztillált vízzel való mosás és vákuum alatti szárítást követően a metilált guaninoknál hasadt DNS-eket 200 l steril desztillált vízben oldottam fel. A reakciók hatékonyságát ezután 1%-os agaróz gélen történő elektroforetikus elválasztásával ellenőriztem. A ligáció-mediálta-pcr-eket (LM-PCR) a DMS-sel és piperidinnel kezelt DNSekből (kontroll és footprintelt ) kisebb változtatásokkal (Niller és mtsai., 1995, Szenthe és mtsai, 2013c) a már korábban leírt módon (Mueller és Wold, 1989, Garrity és Wold, 1992) végeztem. A módszer lényegét az 11. ábra mutatja be. 5. ábra A Ligáció-Mediálta-PCR (LM-PCR) folyamata ( magyarázat: ld. szöveg) 46
47 Az LM-PCR során alkalmazott primereket (Metabion) az 9. táblázat tartalmazza. Az LM-PCR első primer-hosszabbítási reakciója (94 C 5 perc, 60 C 30 perc, 72 C 10 perc) 2 g DNS-t, mindegyik primersorozat I. primeréből 0,3pmol-t, 240 M dntp-t, 1 egység Vent (exo-) DNS polimerázt (New England Biolabs) és az enzimnek működéséhez szükséges puffert (10mM KCl, 10mM (NH 4 ) 2 SO 4, 20mM Tris-HCl (ph8.8), 2mM MgSO 4, 0.1% Triton X-100) tartalmazott. Ezután az elegyhez 5 l közös linkert, 1 l T4 DNS ligázt (5 egység/ l; Boehringer Mannheim), 2 l a ligáz működéséhez szükséges puffert (660mM Tris-HCl, 50mM MgCl 2, 10mM dithioerythritol, 10mM ATP (ph7.5)) és 12 l steril desztillált vizet adtam és egy éjszakán át 4 C-on inkubáltam. Az ún. közös linkert két, egyenként 20pmol/ l koncentrációjú, komplementer oligo (5 -GCGGTGACCCGGGAGATCTGAATTC-3 és 5 -GAATTCAGATC-3 ) 250mM Tris-HCl (ph7.7)-ban történő hibridizálásával (95 C 1 perc, majd körülbelül 4 óra alatt 4 C-ra hűtés) készítettem el. Ezt követően a DNS-t 6 l 3M nátrium-acetát (ph5.2) és 145 l abs. etanol hozzáadásával -20 C-on 5 percig precipitáltam, majd 12000g-vel 25 percig 4 C-on centrifugáltam. A csapadékot ezután 200 l 70%-os etanollal mostam, 5 percig 12000g-vel 4 C-on centrifugáltam, ezután vákuum alatt szárítottam és végül steril desztillált vízben oldottam. Az ezt követő 100 l térfogatú PCR reakció (20-szor : 94 C 1 perc, 60 C 1,5 perc, 72 C 3 perc) a teljes mennyiségű ligátumot a megfelelő primersorozat II. primeréből 10pmolt, a közös linker hosszabbik primeréből szintén 10pmol-t, 240 M dntp-t, 1 egység Vent (exo-) DNS polimerázt (New England Biolabs) és az enzim működéséhez szükséges puffert (lásd feljebb) tartalmazott. Ezalatt a harmadik primert (γ- 32 P)ATP-vel radioaktívan jelöltem: a 30 l térfogatú reakció a jelölni kívánt mennyiségű primer III-at (5pmol/minta), 10 egység Polinukleotid Kinázt (Boehringer Mannheim), 5 l (5000Ci/mmol) (γ- 32 P)ATP-t, és az enzimnek megfelelő puffert (50mM Tris-HCl, 10mM MgCl 2, 0.1mM EDTA, 5mM dithiothreitol, 0.1mM spermidin, (ph8.2)) tartalmazott. 1 órás 37 C-os, majd 10 perces 70 C-os inkubálást követően a jelölt primert Micro Bio-Spin (Bio-Rad) oszlopon tisztítottam. A jelölési reakció előtt leírt PCR reakció befejezését követően 10 l Vent (exo-) DNS polimeráz pufferben (lásd feljebb) oldott, 2,5nmol dntp-t, 0,5 egység Vent (exo-) DNS polimerázt, és a jelölt és tisztított primer III-ból 5pmol-t adtunk azonnal a 47
48 reakcióelegyhez. Ezt követően a reakcióelegyet 1,5 percig 94 C-on, majd 8 ciklust (94 C 2 perc, 60 C 2 perc, 72 C 5 perc) követően 5 percig 72 C-on inkubáltam. A reakcióelegyben található PCR termékeket ezután 1-szeres 0,5M Tris-HCl (ph8.0)-val telített fenollal, majd 2-szeres kloroformmal tisztítottam. A felső vizes fázishoz ezután 0,1 térfogat 3M nátrium-acetátot és 2,5 térfogat abs. etanolt adtam, majd rövid vortexelést követően 30 percre -20 C-ra helyeztem, végül 30 percig 12000g-vel, 4 C-on centrifugáltam. A csapadékot ezután 70%-os etanollal mostam, majd vákuum alatt szárítottam, végül 20 l futtató festékben (96% formamid, 0,01% brómfenol-kék, 0,01% xylén-cianol) vettem fel. A footprint-reakciók láthatóvá tételéhez 5%-os poliakrilamid gélt használtam: 12,5ml 40%-os akrilamid oldathoz (akrilamid: biszakrilamid / 19:1) 45g urea-t és 10ml 10xTBE-t adtam, majd desztillált vízzel 100ml-re töltöttem fel. Ezután az oldatot 0,45 m pórusméretű szűrön filtráltam, majd 500 l 10% APS és 50 l TEMED hozzáadása után a megfelelő méretű üveglapok közé öntöttem. Az üveglapokat előzőleg alkohollal lemostam majd szilánnal kezeltem. A mintákat közvetlenül a gélbe való bemérés előtt 2 percig 95 C-on denaturáltam, utána rögtön jégkására tettem, majd 4 l-t a fent leírt poliakrilamid gélen 1xTBE pufferben, 1700V-al, 55 C-on elektroforézissel szétválasztottam. Ezt követően a gélt 3M Whatmann szűrőpapírra vittem át, celofánnal letakartam és vákuum-szárítóval 100 percig 70 C-on szárítottam. Az így előkészített kiszárított gélt 2 órás előhívás után Phospho-imager (Personal Molecular Imager, BIO-RAD) segítségével scanneltem és a hozzá tartozó programmal értékeltem. 9. táblázat A DMS in vivo footprintinghez használt primerek (a 9. és a 12. lett radioaktívan jelölve) mir146-7 mir146-8 mir146-9 mir mir mir CAGCGCCTGACCAGAACTTCCTC-3 5 -GCGCATCCTGCACAGAACGCTCTAG-3 5 -CTCTAGAGCGCGCAGGCCAAAGCAC-3 5 -CTGGAGAGACTCTTATTTGCTGGGG-3 5 -CTGGGGTAGAGGAAGGCAGCTAAGG-3 5 -CCACACCCTGCACGCTAACCCTCTC-3 48
49 EREDMÉNYEK Eredményeim (lásd a 12-től a 15. fejezetig) a Szenthe és mtsai (2013b) közleményben szerepelnek. Az irodalmi összefoglaló egy része a Szenthe és mtsai. (2013a) közleményben került publikálásra. E két, a doktori dolgozat alapját képező közlemény, valamint egy más témában, de hasonló metodikák felhasználásával készült publikációm (Szenthe és mtsai, 2013c) a Melléklet fejezetben találhatók meg. 12. A primer átirat térszerkezet vizsgálata és kontroll szekvenálás Azt, hogy egy génről lehetséges-e mikrorns átíródása, termodinamikai összefüggések alapján, az mrns térszerkezeti vizsgálatát végző programok segítségével határozhatjuk meg. A mir-146a esetében a Jazdzewski és mtsai által ban TargetRank software segítségével kimutatott térszerkezeti összefüggéseket az mfold programmal (University at Albany, State University of New York) sikerült igazolnom (12. ábra) és kimutatni, hogy térszerkezetileg egyetlen olyan hosszú duplaszálú régió van a mir-146a génről átíródó elsődleges transzkripten (primer átirat), melyről premikrorns levágódása (a jelenlegi tudásunk szerint) lehetséges. 49
50 B 6. ábra A mir-146a térszerkezete a termodinamikai összefüggések alapján, A.: TargetRank software (Jazdzewski és mtsai, 2008); B.: mfold software eredménye, primer átirat térszerkezeti elrendeződése, a piros szín a legstabilabb régiókat jelöli, tehát ezeknek a régióknak a kialakulása a legvalószínűbb. 13. mir-146a expressziós vizsgálatok A vizsgálataim során EBV pozitív és negatív sejtvonakat használtam. A teljes, frissen expresszált mikrorns mennyiségét kimutató Nuclear run-on módszer (13. ábra) segítségével nem tudtam kimutatni mir-146a átiratot az EBV pozitív Burkitt limfóma (BL) sejtvonalak közül a Rael és az Akata sejtvonalban, valamint a C2G6, C4A3 tejmirigy epitélium karcinóma eredetű sejtvonalakban. A nazofaringeális karcinóma eredetű C666-1 sejtvonal csak kismértékű expressziót mutatott. Erős aktivitást figyelhetünk meg ezzel szemben az EBV pozitív limfoblasztoid CBM1 sejtvonal és az ugyancsak EBV pozitív Raji (BL) sejtvonal esetében. Mindkét sejtvonal LMP1-et expresszál, így a fokozott mir-146a expresszió magyarázható az LMP1 által 50
51 Relatív expresszió (%) aktivált NFkB faktor expressziót fokozó hatásával. Az EBV és LMP1 hiánya ellenére a sporadikus BL sejtvonalakban (BL41, DG75, BJAB) megfigyelhető mir-146a expresszió, a sejtvonalak közül a BJAB volt a legerősebb. A BL41 és az utólag, in vitro EBV-vel transzfektált klónja a BL41-E95B esetében kimondhatjuk, hogy az EBV jelenléte LMP1 expresszió nélkül nem serkenti a mir-146a expressziót. 13. ábra A teljes mir-146a mikrorns mennyisége a vizsgált sejtvonalakban. Az érett mikrorns mennyiségét kis RNS Northern blot segítségével vizsgáltam radioaktívan jelölt LNA próba jelöléssel, akrilamid gélben való elválasztás után (14. ábra). Az eredmények azt mutatják, hogy a teljes, frissen szintetizált RNS mennyiségét kimutató Nuclear run-on módszer eredményeivel teljes mértékben megegyeztek a kapott eredmények, kivéve egy sejtvonalat (C666-1), melynél nagyon kismértékű aktivitást mutatott a teljes, frissen szintetizált RNS mennyisége. 51

52 A B 14. ábra Az érett mir-146a mennyisége Northern blot módszer eredményeként kapott futtatási képen. A: a mir-146a-val végzett jelölés; B: belső kontrollként U6 RNS-re specifikus próbával végzett hibridizáció. A nyilak az érett mir-146a-t, valamint az U6 kontroll RNS-t mutatják. Minták: 1: BL41; 2: BL41-E95B; 3: BJAB; 4: DG75; 5: CBM1-Ral-STO; 6: RAEL; 7: Akata; 8: Raji; 9: C666.1; 10: C2G6; 11: C4A3; 12: szintetizált kontroll mir-146a RNS; 13: szintetizált kontroll mir-146b RNS. 52
53 14. DNS metilációs vizsgálat A vizsgálatok során a felhasznált B-sejt és epitheliális sejtvonalak esetében a mir-146a gén promóter-régióján levő, valamint kettő, az első exonon levő CpG dinukleotid metilációs vizsgálatát végeztem el. Az expressziós aktivitást nem mutató sejtvonalak közül erősen metiláltnak adódott a mir-146a promóter régió (15. ábra) a Rael és az Akata sejtvonalak esetében, amíg az ugyancsak nem expresszáló C2G6 és C4A3 sejtvonalaknál részleges metiláció volt csak megfigyelhető. A jelentős aktivitást mutató CBM1 limfoblasztoid sejtvonal esetében teljesen metilálatlannak adódott a régió. A sporadikus BL sejtvonalak (BL41, BL41-E95B, BJAB, DG75) esetében az expressziós aktivitásnak megfelelően metilálatlan promóter régiót figyelhetünk meg. A Raji, erősen expresszáló endémiás BL sejtvonal esetében a metilálatlan promóter mellé erős expressziós aktivitás párosul. Az egyetlen sejtvonal, melynek promóter-metiláltsága nem mutatott összefüggést az aktivitással a C666-1 nazofaringeális karcinóma eredetű, EBV pozitív, de LMP1 negatív sejtvonal volt, melynek metilálatlan promótere inaktívnak adódott. Ezek alapján az adatok alapján kimondhatjuk, hogy egy sejtvoonal kivételével jó összefüggés figyelhető meg a mir-146a promóter metilációja és a promóter aktivitás között. Bizonyos sejtvonalak esetében (sporadikus BL) a metilálatlan régió ellenére csak enyhe expressziós aktivitást figyelhetünk meg, ez más szabályozó folyamatok (pl. LMP1 aktivációs szerepe) fontosságát jelzi. 53

54 15. ábra CpG dinukleotidok metilációjának mértéke a biszulfit kezelés és azt követő direkt szekvenálás eredményeként B-sejt és epitheliális sejtvonalakban. A citozin metilációját az adott pozícióban levő pontok jelölik a következő módon: vonal, 0 %; egy pont, 0 25 %; két pont, %; három pont, %; négy pont, %. A szürke hátterű, 455bp hosszú régió a promóteren elhelyezkedő CpG sziget területét jelzi (-631- tól -176-ig bázisig (a nulla pozíció az iniciációs pontot jelzi)). Sárga jel: NFkB kötőhely, piros jel: c-myc kötőhely. 54
55 15. Kromatin immunprecipitáció és azt követő real-time PCR A CpG metilációhoz hasonlóan a hisztonmodifikációk a gének szabályozó régiójában hatnak a kromatin szerkezetre, így befolyásolva azok aktivitását. Kromatin immunprecipitációt és azt követő real-time PCR-t használtam, hogy meghatározzam a mir-146a gén aktivitásának a hármas hiszton acetilációjával (ach3), a négyes hisztonfehérje acetilációjával (ach4), valamint a hármas hiszton négyesszámú lizin oldalláncának kétszeres metilációjával (H3K4me2) való összefüggését. Az összefüggéseket a gén három régiójában vizsgáltam: a szabályozó régióban (promóter; 16. ábra), az iniciációs pont körül és az első exon területén (17. ábra), valamint az első intron területén (18. ábra). 16. ábra ach3, ach4 és H3K4me2 mértéke a mir-146a gén szabályozó (promóter) régiójában. 55
56 17. ábra ach3, ach4 és H3K4me2 mértéke a mir-146a gén iniciációs pontjának környékén, valamint az első exon területén. 18. ábra ach3, ach4 és H3K4me2 mértéke a mir-146a gén első intronjának területén. 56
57 Az eredmények kimutatták, hogy a CpG metiláció szempontjából nagymértékben metilált Rael és Akata sejtvonalak promótere mindhárom vizsgálati génrégióban hisztonmodifikációk szempontjából jelentősen metilálatlannak és acetilálatlannak adódott. Az aktiváló hisztonmodifikációk közepes mértéket mutattak az ugyancsak közepes mértékben CpG metilált promóterű C2G6 és C4A3 sejtvonalak esetében. Ehhez hasonlóan az inaktív promóterű C666-1 nagyon alacsony mértékben mutatott aktiváló modifikációkat a CpG metilálatlanság ellenére. Az expressziós aktivitást mutató limfoid sejtek (BL41, BL41-E95B, BJAB, DG75, CBM1, Raji) közepes vagy emelkedett mértékben tartalmaztak a szabályozó régióban H3K4me2 és ach3 modifikációkat, miközben a négyes hisztonfehérje acetilációja csak közepes mértékben volt kimutatható. Az adatok alapján kijelenthetjük, hogy a mir-146a promóter esetében az általam vizsgált hisztonmodifikációk közül a H3K4me2 és a hármas hiszton acetilációja, mely jelentős összefüggést mutatott a sejtvonalakban az expressziós aktivitással, a négyes hiszton acetilációja kevésbé, kivéve a Raji sejtvonalat, melynél jelentős mértékű ach4- et figyelhetünk meg a szabályozó régióban, mely jól korrelál az erős expressziós szinttel. Kijelenthetjük, hogy az expressziósan inaktív sejtvonalak szignifikánsan kisebb mértékben tartalmaztak aktiváló modifikációkat, mint az aktívak, az általam vizsgált mindhárom régióban. A jelerősség jelzi az adott modifikáció százalékos arányát a teljes kromatinhoz képest, így az eredmények alapján a promóter aktivitás és a hisztonmodifikációk mértékének összefüggését az szabályozó régióban, valamint az iniciációs ponttól távol, az első intron területén kapott adatok tükrözik a legjobban, mindhárom általam vizsgált aktiváló modifikáció esetében. 16. DMS in vivo footprinting A vizsgálatok során három sejtvonalat választottam ki, melyek jól képviselik az epigenetikai vizsgálataimban használt sejtvonalakat. A Rael-t, mint EBV pozitív, de mir-146a-t nem expresszáló, inaktív promóterrel rendelkező, a CBM1-et, mint EBV pozitív, limfoblasztoid, erősen expresszáló, valamint a C666-1-et, mint epitheliális, ugyancsak EBV pozitív, de nem expresszáló sejtvonalat. A feltételezett és vizsgálni kívánt faktorkötéseket a szakirodalom alapján (Cameron és mtsai, 2008), valamint a TFSEARCH ( software segítségével 57

58 határoztam meg, a DMS in vivo footprinting módszer azon tulajdonságát figyelembe véve, hogy olyan faktorok kötései vizsgálhatók vele, melyeknek kötőhelye guanin bázist tartalmaz. A munkám során a mir-146a promóter aktivitását feltételezhetőleg befolyásoló NFkB, c-myc, c-ets és PU.1 faktorok kötődését vizsgáltam. A DMS in vivo módszer eredményeként kapott futtatási képen (19. ábra) a kontroll és DMS-sel in vivo kezelt minta guanin bázisainak egymáshoz képesti futási mintázatbeli különbségét kell figyelni. 19. ábra Faktor kötések a mir-146a promóter területén DMS in vivo módszerrel vizsgálva. Minták: 1: Rael kontroll, 2: Rael in vivo DMS kezelt, 3: CBM1 kontroll, 4: CBM1 in vivo DMS kezelt, 5: C666-1 kontroll, 6: C666-1 in vivo DMS kezelt. A bázis pozíciót mutató számok a mellékletben megtalálható szekvencián feltüntetett számokat jelentik. 58
59 Az eredmények azt mutatják, hogy az NFkB, c-ets és c-myc területén, vagy ahhoz közvetlen közel a kontrollhoz képest a CBM1 és C666-1 sejtvonalban láthatunk változást, amely fehérje kötődést valószínűsít. A Rael sejtvonalban egyik kötőhely területén sem figyelhetünk meg mintázatbeli változást, így kijelenthetjük, hogy az inaktív promóterű Rael sejtvonal aktiváló transzkripciósfaktor-kötéseket nem tartalmazott. A PU.1 faktor egyik sejtvonal esetében sem mutatott kötést. A mir-146a-t erősen expresszáló CBM1 és az inaktív promóterű C666-1 esetében nem lehet az általam vizsgált faktorok kötésében különbséget kimutatni, így a promóter-aktivitásbeli különbséget vagy az epigenetikai szabályozások eredményezik, vagy a DMS in vivo footprinting módszer segítségével nem vizsgálható faktorok eltérő kötődése okozhatja, esetleg mindkét mechanizmus együttesen. 59
60 EREDMÉNYEK ÉRTÉKELÉSE Az eukromatikus régiók növényi és állati sejtekben is transzkripcionálisan aktív géneket tartalmazó területeket jeleznek (Van Driel és mtsai, 2004; Wu és mtsai, 2011b). A fehérjét kódoló gének transzkripcionális aktivitását epigenetikai szabályozási mechanizmusok befolyásolják. A fehérje kódoló génekhez hasonlóan, az önálló promóterrel rendelkező mikrorns gének esetében is hasonló szabályozási folyamatokat tételezhetünk fel. A nem önálló promóterrel rendelkező, kódoló (de intronikus) területeken elhelyezkedő mikrorns-eknél a T-sejtek esetében kimutatták, hogy a fehérjét kódoló génre ható kromatin módosítások a kis RNS mennyiségét is befolyásolják (Barsik és mtsai, 2009). A mikrorns-eket kódoló gének epigenetikai szabályozásáról nagyon kevés adatot találhatunk a szakirodalomban (pl. Liang és mtsai, 2009; Mazar és mtsai, 2011) annak ellenére, hogy mennyiségi megváltozásuk jelentős folyamatok (kórfolyamatok) hátterében állhat. A mir-146a humán mikrorns tumor-kialakítási szerepének és mechanizmusának megértéséhez elengedhetetlen azon folyamatok ismerete, melyek befolyásolják a gén aktivitását. Elsődlegesek, a gén expresszióját befolyásoló epigenetikai és fehérje (transzkripciós faktorok) szintű szabályozó folyamatok, melyek megváltozása jelezheti a kóros folyamatok kialakulását. Az egészséges immunsejtekben zajló folyamatokban fontos szerepet tulajdonítanak a mir-146a-nak, mennyiségi megváltozása számos kóros elváltozásban kimutatható, ez nem csak a tumorkialakulási folyamatok következménye, hanem indukáló lépése is lehet. 17. Epigenetikai szabályozási mechanizmusok A vizsgálataimban B-sejtvonalakat és epiteliális tumor eredetű sejtvonalakat használtam. A kapott eredmények nem alkalmazhatók közvetlenül a szervezetben lejátszódó tumorképződési mechanizmusok, folyamatok bizonyítására, de mint modellrendszer könnyen használhatók a gén expressziós aktivitását befolyásoló alapvető folyamatok tanulmányozására és így segíthetik az alap törvényszerűségek megismerését. Az expressziós vizsgálatokat két céllal végeztem, egyrészt a promóter aktivitás meghatározása (az epigenetikai vizsgálatok eredményével együtt tárgyalandó), 60
61 másrészt a mikrorns-ek promóter aktivitást jellemző formájának (pri-, pre- vagy érett mirns forma mennyisége korrelál-e a promóter aktivitással) meghatározása céljából. A szakirodalomban megtalálható néhány cikkben (pl. Liang és mtsai, 2009; Mazar és mtsai, 2011), melyek a mikrorns gének epigenetikai szabályozásáról szólnak minden esetben az érett forma mennyiségével jellemezik a promóter aktivitást. Ez, a kisrnsérés szabályozásának bonyolultsága miatt kérdéseket vet fel. A Northern blot (érett forma) és Nuclear run-on ( pre formáig összesítve kimutató) módszerek segítségével kapott eredmények alapján kimondhatjuk, hogy a mir-146a esetében az érett, valamint a primer és pre forma összesített eredményei között nem sikerült jelentős mennyiségi különbségeket kimutatni (az erősen expresszáló Raji és BJAB sejtvonal között csak dominancia különbség figyelhető meg) az általam vizsgált tizenegy sejtvonal egyikében sem, így kijelenthetjük, hogy az érett mikrorns mennyiségéből a mir-146a esetében következtetni lehet a promóter aktivitásra. Ezek az eredmények természetesen nem azt jelentik, hogy más gének esetében nem lehetnek eltérések, így a Nuclear run-on módszer használata javasolt a mikrorns gének promóter aktivitásának jellemzéséhez. Az epigenetikai vizsgálatok eredményei alapján kimondhatjuk, hogy az aktív mir-146a promóterű sejtvonalak esetében a régió DNS metilációs szempontból metilálatlan volt, valamint közepes vagy jelentős mértékben kapcsolódott az általam vizsgált aktiváló hisztonmodifikációkhoz (H3K4me2, ach3, ach4). Ezzel szemben a nem aktív mir-146a promóterű sejtvonalakban a metilált vagy részlegesen metilált szabályozó régió területén nem, vagy csak kis mértékben volt kimutatható az aktiváló hisztonmodifikációk jelenléte. Kivételként megemlíthetjük, hogy a nem aktív promóterű sejtvonalak közül egyedül a nazofaringeális karcinóma eredetű C666-1 sejtvonalban tapasztaltam a szabályozó régióban metilálatlan CpG dinukleotidokat, viszont az inaktív promóternek megfelelően az aktiváló hisztonmodifikációk nem voltak kimutathatók a régióban. Ezek az összefüggések jelzik, hogy a DNS metiláció hiánya önmagában nem elégséges a mir-146a promóter bekapcsolásához. A hisztonmodifikációs vizsgálatok kimutatták, hogy az aktiváló modifikációk teljesen hiányoztak az erősen DNS metilált promóterű BL sejtvonalakban (Rael és Akata). Közepes vagy kis mértékben voltak megtalálhatók a nem expresszáló epitheliális sejtvonalakban. Nem minden sejtvonal esetében figyelhettünk meg közvetlen összefüggést a mir-146a expresszió és az általam vizsgált aktiváló modifikációk között, ez jelzi a korábban említettekkel összefüggésben, hogy más 61
62 faktorok szabályozó hatása is fontos lehet a promóter aktivitásának szabályozásában. A H3K4me2 és ach3 modifikációk a promóterrégió és első intron területén nagy mértékben jelen voltak a mir-146a-t expresszáló sejtvonalak nagy részénél, míg az ach4 modifikáció nem volt kimutatható számottevő mértékben. Ezek az eredmények azt jelzik, hogy a mir-146a gén esetében a szabályozó régió és az iniciációs ponttól kódoló irányban elhelyezkedő területek ugyancsak fontos szerepet játszhatnak a hisztonmodifikációkon keresztüli aktivitás-szabályozásban, de modifikációtól függő mértékben. Az EBV-t hordozó B-sejtvonalakban a fertőzött sejt érési állapotától függő módon expresszálódnak a virális gének (Takács és mtsai, 2010), ezt úgynevezett látencia állapottól függő expressziós mintázatnak nevezzük. Az első látenciában a vírus egyetlen sejtmagi fehérjéje expresszálódik (EBNA1), melynek szerepe a vírus episzóma kromatinhoz való horgonyzásában van, valamint az EBER nem transzlálódó RNS-ek vannak még jelen. A hármas látencia állapotban ezzel szemben mind a hat EBNA fehérje, valamint a három látens membrán fehérje (LMP1, LMP2A és LMP2B) és az EBER nem transzlálódó RNS-ek is expresszálódnak. A termelődő faktorok közül az LMP1 onkoprotein a mir-146a expressziót indukálja, az NFkB útvonalon keresztül (Motsch és mtsai, 2007; Cameron és mtsai, 2008), ennek a folyamatnak az eredménye valószínűleg, hogy a CBM1 és Raji hármas látencia típusú sejtvonalakban erőteljes, míg az LMP1 negatív egyes látencia típusú Rael és Akata sejtvonalakban egyáltalán nem tapasztaltunk mir-146a expressziót. Az EBV vagy az LMP1 hiányában közepes expressziós szintet tapasztaltunk a BL41 és DG75 sejtvonalakban, míg az ugyancsak EBV negatív BJAB sejtvonal nagyon erőteljes expressziót mutatott. Ezek az összefüggések az LMP1 hiányában nem magyarázhatók az LMP1 aktiváló hatásával főleg, hogy a promóter egyik nagyon fontos szabályozó faktorának, az NFkB-nek az egyik alegysége, a RelB nagyon kis mennyiségben (de transzfekcióval fokozható módon) van jelen a BJAB sejtvonalban (Pai és mtsai, 2002). Ezeket az összefüggéseket még árnyalja, hogy a mir-146a által negatívan szabályzott TRAF6 és IRAK1 a Tolllike receptorhoz kapcsolt NFkB jelátviteli útvonal része, így a kialakuló negatív visszacsatoláson (Taganov és mtsai, 2006) keresztül egy komplex szabályozási metódus képzelhető el. A BJAB és a Raji sejtvonal erőteljes mir-146a expressziója jelzi, hogy az NFkB-n kívül más faktor pozitív hatása is szükséges a mir-146a promóter aktiválásához, hiszen az esetleges NFkB-n keresztüli, LMP1-től független fokozó hatás 62
63 a BJAB alacsony RelB szintje miatt nem tudna érvényesülni. A kérdéses aktivátor faktor hatása lehet az EBV negatív BL sejtvonalak (BJAB, BL41 és DG75) erős vagy közepes mértékű mir-146a expressziója. Ennek az aktivátor mechanizmusnak a vizsgálatához még további kutatások szükségesek (a transzkripciós faktorok általi szabályozás folyamatának részleges vizsgálatát ld. 18. fejezet), de kiegészítésként elmondhatjuk, hogy más kutatócsoportok feltételezik, hogy az LMP1-en kívül más EBV által expresszált fehérjék is pozitívan hatnak a promóterre, ilyen faktor pl. az EBNA2 (Rosato és mtsai, 2012), mely a hármas látencia állapotban termelődik. Az EBV esetleges hatását a promóterre a korábban részletezett összefüggéseken kívül az EBV-t nem tartalmazó BL41 és az in vitro EBV-vel transzfektált klónja, a BL41-E95B sejtvonalak eredményeivel is jellemezhetjük. A BL41-E95B alacsonyabb mir-146a expressziót mutatott, mint az EBV negatív párja, a BL41. A BL41-E95B esetében az LMP1 nem expresszálódik (Masucci és mtsai., 1987), de a fentiekben jelzett, még nem ismert faktor által sem aktiválódik a promóter, így az aktivitást szabályozó, a DNS metiláción kívüli, további jelentős inaktiváló mechanizmusok hatása is feltételezhető. Az epigenetikai szabályozási mechanizmusok eredményeinek összefoglalásaként elmondhatjuk, hogy mind a CpG metilációnak, mind a hisztonmodifikációknak jelentős szerepe van a mir-146a promóter szabályozásában, de gazdasejttől függő mértékben. A CpG metiláció szükséges feltétele a promóter inaktiválásának, de az inaktivitást egyéb, a munkám során nem vizsgált szabályozó mechanizmusok eredményeként is megfigyelhetjük. Az aktiváláshoz nem elégséges a demetiláció, az aktváló hisztonmodifikációk jelenléte is szükséges, hozzátéve, hogy az aktiváló modifikációk eltérő mértékben járulhatnak hozzá az aktivitás szabályozásához. 18. Faktorkötések és szerepük a szabályozásban A mir-146a promóter aktivitásának az LMP1-en kívüli szabályozását végző faktor vizsgálatát in vivo footprinting módszer segítségével vizsgáltam (három, a mir- 146a promóter aktivitást befolyásoló fontosabb tulajdonságok alapján kiválasztott sejtvonalban), mely az eddig kimutatott faktorkötések (Cameron és mtsai., 2008) nagy részére releváns információt ad (a guanin bázist tartalmazó kötőhelyű faktorok 63
64 kimutatásához megfelelő). A módszer tulajdonságainak és a faktorok fontosságának figyelembevétele alapján a PU.1, c-ets, c-myc és NFkB faktorok kötődését vizsgáltam. Mivel az NFkB és c-myc faktorok szerepéről és működési mechanizmusáról található adat a szakirodalomban, valamint a mir-146a és EBV-s kapcsolatuk is részletesen vizsgált, a következőkben ezen faktorok kötési eredményeinek tárgyalására szorítkoznék. A másik két faktor (PU.1 és c-ets) eredményét a 16. fejezet tartalmazza. Korábban Cameron és munkacsoportja (2008) által in silico kimutatott fehérje kötődések, valamint ugyanezen csoport által az NFkB kötőhely területén végzett báziscsere vizsgálat kimutatta, hogy az NFkB kötődését csökkentő hatások a mir-146a expresszió csökkenését is okozzák. Vizsgálataim során in vivo módszerrel sikerült kimutatnom az NFkB kötődését két sejtvonalban, melyek közül az epitheliális sejtvonal a kötés ellenére nem expresszált mir-146a-t. A nem aktív promóterű Rael sejtvonal esetében a kötődés nem volt kimutatható a kontrollhoz képest. Az epitheliális sejtvonal a Cameron csoportja által használt epitheliális sejtvonalhoz képest az LMP1 expresszió hiányában különbözik (LMP1 pozitív sejtvonalat használtak, mely mir-146a expressziót mutatott), mely magyarázhatja, hogy az LMP1 általi aktiváció hiányában a C666-1 esetében az NFkB kötődése ellenére sem tapasztalhatunk mir-146a expressziót. A Rael, mint inaktív sejtvonal esetében semmilyen aktiváló faktorkötődést nem figyelhetünk meg, mely jól korrelál a promóter inaktivitásával. A másik faktor, melynek kötődése kimutatható volt az aktív CBM1 és inaktív C666-1 sejtvonalban a c-myc, melynek EBV-s kapcsolatát korábban részletesen vizsgálták más kutatócsoportok. Ezen adatok alapján elmondhatjuk, hogy a c-myc egy DNS metiláció érzékeny transzkripciós faktor, mely a Burkitt limfómás tumorok esetében a 8. kromoszómáról valamelyik immunglobulin gén lókuszára (2p11-12, 14q32, 22q11) transzlokálódik, így folyamatos expressziót mutat. Az állandó jelenlét a sejtproliferáció fokozódását eredményezi, ami a tumorszövet fennmaradásához, terjedéséhez vezethet. Ez a változás apotózisra érzékenyíti a sejtet, ami ellen EBV jelenlétében a sejt által termelt EBER RNS-ek és a gazdasejt fenotípusos megváltozása (állandóan osztódó, de memóriasejt érési állapotot mutató fenotípus) segítségével védekezik. Ez utóbbi mechanizmus egyébként, valamint a p53 gén mutációja, és a bcl-2, IL-10 antiapoptotikus faktorok termelődése EBV hiányában is segítheti az úgynevezett sporadikus (tehát EBV-t nem tartalmazó) BL kialakulását és fennmaradását (Kitagawa és mtsai., 2000; Nanbo és Takada, 2002, Klein, 1996; Kieff, 1996). 64
65 A korábbi adatokat kiegészítendő a kutatócsoportunk még nem közölt adatai alapján a c-myc azokban a sejtvonalakban mutatott erőteljes fehérje szintű expressziót (BL41, BL41-E95B, Akata, Rael, C666, C2G6, C4A3), melyek a mir-146a-t nem, vagy csak kis mértékben fejezték ki. Ezzel szemben az erősen expresszáló sejtvonalakban (BJAB, CBM1 és Raji) alacsony volt a c-myc expresszió (DG75 kivétel, mert gyenge c- Myc és mir-146a expressziót mutatott). Ezen összefüggések ismeretében nem magyarázható közvetlenül a mir-146a expresszió mértéke, hozzátéve, hogy a c-myc csak más fehérjékkel együtthatva tudja kifejteni a hatását (pl. Max fehérje, Prendergast és mtsai, 1991), mely tovább árnyalhatja a közvetlen transzkripcionális hatását a mir- 146a promóter aktivitására. A faktor kötések és DNS metilációs eredmények kapcsolatát tárgyalva elmondhatjuk, hogy az NFkB és a c-myc metilációérzékeny kötődéssel rendelkezik, ami a kötőhelyükön megtalálható CpG dinukleotidok metilálódásakor kötésgátlás formájában jelentkezik. Az általam vizsgált sejtvonalak esetében a C4A3 sejtvonal 25%-nál alacsonyabb, elhanyagolható mértékű metilációjától eltekintve minden sejtvonal esetében metilálatlannak adódott a c-myc kötőhely területe, így a Rael sejtvonal esetében megfigyelhető kötődéshiány talán azzal magyarázható, hogy a régió jelentős mértékű metiláltságot, valamint aktiváló hisztonmodifikációktól való mentességet mutatott, és ez a teljes inaktiváltsági állapot valószínűleg a kötőhelyen fizikailag nem teszi lehetővé a c-myc kötést, annak metilálatlansága ellenére. Az NFkB kötés a szakirodalom alapján a c-myc-hez hasonló módon metiláció érzékeny. A sejtvonalak nagy részében metilálatlannak adódott a kötőhelye, a két inaktív promóterű sejtvonal (Akata és Rael) kivételével, mellyel összefüggésben, a footprinting módszerrel vizsgált Rael sejtvonal esetében nem is volt tapasztalható NFkB kötődés a kötőhelyen. A C666-1 sejtvonal esetében megfigyelhető c-myc és NFkB kötés a régió metilálatlanságával jó összefüggést mutat, az ennek ellenére megfigyelhető promóter inaktivitás egyéb szabályozófolyamatok szerepét mutatja. A faktorkötések összefoglalásaként elmondhatjuk, hogy részletes sejtvonalankénti vizsgálat szükséges a szerepük megítéléséhez, hiszen az NFkB és c-myc kötődéseket együtt érdemes figyelembe venni, természetesen a kofaktoraik jelenlétének ismeretében. 65
66 EREDMÉNYEK ÖSSZEFOGLALÁSA A mir-146a humán mikrorns tumor-kialakítási szerepének és mechanizmusának megértéséhez a mennyiségi viszonyait befolyásoló mechanizmusok ismerete elengedhetetlen. Vizsgálataim során a következő megállapításokra jutottam: a mir-146a promóter aktivitását jobban tükrözi az újonnan képződő átirat (prekurzor RNS) mennyisége, mint az érett mir-146a alaké. A DNS metiláció hiánya önmagában nem elégséges a promóter bekapcsolásához, hiszen az inaktív promóterű sejtvonalaknál csak két esetben figyelhettünk meg teljes metilációt, a többi sejtvonalnál közepes mértékű, vagy egy esetben egyáltalán nem tartalmazott metilált CpG dinukleotidokat a szabályozó régió. Az aktív promóterű sejtvonalak minden esetben metilálatlan promótert tartalmaztak. A vizsgált hisztonmodifikációk közepes vagy jelentős mértékben voltak jelen az aktív promóterű sejtvonalak esetében, elsősorban a szabályozó régió és az első intron területén, ezzel szemben az inaktív sejtvonalaknál kismértékben vagy egyáltalán nem lehetett a jelenlétüket kimutatni. A DNS metiláció és a hisztonmodifikációk kapcsolataként elmondhatjuk, hogy az aktiváló modifikációk szinte teljesen hiányoztak a hipermetilált, inaktív mir- 146a promóterekkel jellemezhető BL sejtvonalakban, és szintjük alacsony volt az epiteliális sejtvonalakban, melyeket változó szintű promóter metiláció jellemzett. A mir-146a promóter területén in silico kimutatott faktorkötőhelyek in vivo vizsgálata eredményeként kimondhatjuk, hogy a kiválasztott sejtvonalak esetében a c-ets, c-myc és NFkB kötések a mir-146a-t expresszáló BL sejtvonalban és az expressziót nem mutató epitheliális sejtvonalban voltak kimutathatók, ezzel szemben az inaktív BL sejtvonal egyik faktor kötődését sem mutatta. 66
67 A BL sejtvonalak faktorkötése jó összefüggést mutatott a promóter aktivitással, miközben az epiteliális sejtvonal esetében más faktorok vagy modifikációk hiányának túlsúlya okozhatja a régió inaktivitását. A PU.1 faktor kötése nem volt kimutatható az általam vizsgált sejtvonalak egyikében sem. A szakirodalmi adatok alapján az aktivitás szabályozásában fontosnak vélt két faktor jelentőségéről kimondhatjuk, hogy a c-myc és az NFkB is metiláció érzékeny kötődéssel rendelkezik. A c-myc esetében az általam használt minden sejtvonal esetében metilálatlannak adódott a kötőhely (egy esetben nagyon gyenge metiláció volt megfigyelhető), így a kötődést elvileg létre tud jönni. Elképzelhető, hogy a mir-146a-t nem expresszáló BL sejtvonalaknál a promóter régió represszív kromatin szerkezete gátolhatja a c-myc kötődését. Az NFkB kötődés esetében a c-myc esetében leírottakhoz hasonló összefüggések figyelhetők meg, az inaktív promóterű Rael és Akata sejtvonalak kivételével, melyeknél erőteljes kötőhelyi metiláció gátolhatja a faktor kapcsolódását. Az összefüggések értékelésénél fontos a sejtvonal LMP1 pozitivitása, mivel az növeli az NFkB mennyiségét. Bizonyos EBV negatív sejtvonalakban az LMP1 hiányában is megfigyelhetünk erőteljes vagy közepes mértékű mir-146a expressziót, ez az egyéb aktiváló faktorok, mechanizmusok fontosságát jelzi. BJAB sejtvonal vizsgálata azt mutatja, hogy EBV negatív sejtben és alacsony NFkB szint esetén is előfordulhat erőteljes mir-146a promóter aktivitás. A C666-1 epitheliális sejtvonal analízise arra utal, hogy NFkB kötődése a mir- 146a gén szabályozó régiójához nem elegendő a mir-146a promóter aktiválásához. 67
68 KÖSZÖNETNYILVÁNÍTÁS Szeretnék köszönetet mondani témavezetőimnek Dr. Minárovits Jánosnak és Dr. Salamon Dánielnek a rengeteg segítségért, a hasznos szakmai tanácsaikért, támogatásukért, melyeknek köszönhetően elvégezhettem a dolgozatomhoz szükséges munkát. Köszönet illeti Dr. Bánáti Ferencet, tiszteletbeli témavezetőmet, akivel minden technikai kérdésemet megoszthattam és sok segítséget és lelki támogatást nyújtott a munka elvégzéséhez. Köszönöm családomnak, akik ösztökéltek a munka elvégzésére, kiemelve Bánkuti Barbarát, aki a végső forma elkészítésénél nagy segítséget nyújtott. Köszönöm Dr. Hans Helmut Nillernek, hogy lehetőségem volt a Regensburgi Egyetem Orvosi- Mikrobiológiai- és Higéniai Intézetében az in vivo footprinting kísérleteket elvégezni, valamint Lózsa Ritának, hogy a kisrns Northern blot vizsgálatot megtanította és segítette a gödöllői Mezőgazdasági Biotechnológiai Kutatóközpontban való munkámat. Köszönet illeti Dr. Szathmáry Zsuzsát, munkaadómat, aki segítette a doktori munka befejezését célirányos kutatási feladatok kijelölésével. Végül, de nem utolsó sorban köszönöm, Koroknai Anitának, Báthori Zoltánnak és Kormuta Zsuzsának a kutatócsoportban eltöltött időszakot és baráti segítségüket. 68
69 IRODALOMJEGYZÉK Altiok E, Minarovits J, Hu LF, Contreras-Brodin B, Klein G, Ernberg I. (1992). Hostcell-phenotype-dependent control of the BCR2/BWR1 promoter complex regulates the expression of Epstein-Barr virus nuclear antigens 2-6. Proc Natl. Acad Sci U S A 89, Arribas AJ, Campos-Martín Y, Gómez-Abad C, Algara P, Sánchez-Beato M, Rodriguez-Pinilla MS, Montes-Moreno S, Martinez N, Alves-Ferreira J, Piris MA, Mollejo M. (2012). Nodal marginal zone lymphoma: gene expression and mirna profiling identify diagnostic markers and potential therapeutic targets. Blood. Vol. 119, No. 3, pp. e9-e21. DOI: /blood Barski A, Jothi R, Cuddapah S, Cui K, Roh TY, Schones DE, Zhao K. (2009). Chromatin poises mirna- and protein-coding genes for expression. Genome Res. 19 (2009) Beisel C, Imhof A, Greene J, Kremmer E, Sauer F. (2002). Histone methylation by the Drosophila epigenetic transcriptional regulator Ash1. Nature, 419, , doi: /nature01126 Bellan C, Lazz S, Hummel M, Palummo N, de Santi M, Amato T, Nyagol J, Sabattini E, Lazure T, Pileri SA, Raphael M, Stein H, Tosi P and Leoncini L. (2005). Immunoglobulin gene analysis reveals 2 distinct cells of origin for EBV-positive and EBV-negative Burkitt lymphomas. Blood 106, Ben-Bassat H, Goldblum N, Mitrani S, Goldblum T, Yoffey JM, Cohen MM, Bentwich, Z, Ramot B, Klein E and Klein G. (1977). Establishment in continuous culture of a new type of lymphocyte from a "Burkitt like" malignant lymphoma (line D.G.-75). Int J Cancer 19, Benfey PN. (1999). Stem cells: A tale of two kingdoms. Current Biology 9:R171 R172 Bhaumik D, Scott GK, Schokrpur S, Patil CK, Campisi J, Benz CC. (2008). Expression of microrna-146 suppresses NF-κB activity with reduction of metastatic potential in breast cancer cells. Oncogene, vol. 27, no. 42, pp Bornkamm, G.W., Hammerschmidt, W. (2001). Molecular virology of Epstein-Barr virus. Phil. Trans. R. Soc. Lond. 356: Braggio E, Philipsborn C, Novak A, Hodge L, Ansell S, Fonseca R. (2012). Molecular pathogenesis of Waldenstrom's macroglobulinemia. Haematologica, Vol. 97, No. 9, pp DOI: /haematol Calin GA, Croce CM. (2006). MicroRNA signatures in human cancers. Nature Reviews Vol6. 69
70 Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, Rassenti L, Kipps T, Negrini M, Bullrich F, Croce CM. (2002). Frequent deletions and down-regulation of micro-rna genes mir15 and mir16 at 13q14 in chronic lymphocytic leukemia. Proceedings of the National Academy of Sciences of the United States of America, Vol. 99, No. 24, pp Cameron JE, Yin Q, Fewell C, Lacey M, McBride J, Wang X, Lin Z, Schaefer BC, Flemington EK. (2008). Epstein-Barr Virus Latent Membrane Protein 1 Induces Cellular MicroRNA mir-146a, a Modulator of Lymphocyte Signaling Pathways. Journal of Virology, p Vol. 82, No. 4, doi: /jvi Chassin C, Kocur M, Pott J, Duerr CU, Gütle D, Lotz M, Hornef MW. (2010). MiR- 146a mediates protective innate immune tolerance in the neonate intestine. Cell Host and Microbe, vol. 8, no. 4, pp , Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW, Tsang YS, Wong N, Whitney BM and Lee JC. (1999). Nasopharyngeal carcinoma cell line (C666-1) consistently harbouring Epstein-Barr virus. Int J Cancer 83, Chuang JC, Jones PA. (2007). Epigenetics and MicroRNAs. Pediatric Research, Vol. 61, No. 5, Pt 2 Clark SJ, Harrison J, Paul CL, Frommer M. (1994). High sensitivity mapping of methylated cytosines. Nucl. Acids Res. 22: Curtale G, Citarella F, Carissimi C, Goldoni M, Carucci N, Fulci V, Franceschini D, Meloni F, Barnaba V, Macino G. (2013). An emerging player in the adaptive immune response: microrna-146a is a modulator of IL-2 expression and activation-induced cell death in T lymphocytes. Blood. 115(2): doi: /blood Csorba T, Bovi A, Dalmay T, Burgyán J. (2007). The p122 subunit of tobacco mosaic virus replicase is a potent silencing suppressor and compromises both sirna and mirna mediated pathways. J. Virol. 81, Dawson CW, Rickinson AB, Young LS. (1990). Epstein-Barr virus latent membrane protein inhibits human epithelial cell differentiation. Nature 344: Di Lisio L, Gómez-López G, Sánchez-Beato M, Gómez-Abad C, Rodríguez ME, Villuendas R, Ferreira BI, Carro A, Rico D, Mollejo M, Martínez MA, Menárguez J, Díaz-Alderete A, Gil J, Cigudosa JC, Pisano DG, Piris MA, Martínez N. (2010). Mantle cell lymphoma: transcriptional regulation by micrornas. Leukemia, Vol. 24, No. 7, pp DOI: /leu Di Lisio L, Sánchez-Beato M, Gómez-López G, Rodríguez ME, Montes-Moreno S, Mollejo M, Menárguez J, Martínez MA, Alves FJ, Pisano DG, Piris MA, Martínez N. (2012). MicroRNA signatures in B-cell lymphomas. Blood Cancer Journal, Vol. 2, No. 2, pp. e57. DOI: /bcj Du L, Schageman JJ, Subauste MC, Saber B, Hammond SM, Prudkin L, Wistuba II, Ji L, Roth JA, Minna JD, Pertsemlidis A. (2009). mir-93, mir-98, and mir-197 regulate expression of tumor suppressor gene FUS1. Molecular Cancer Research, Vol. 7, No. 8, pp DOI: / MCR
71 Ehrlich M, Wang RYH. (1981). 5-methylcytosine in eukaryotic DNA. Science 212: Eliopoulos AG, Young LS. (2001). LMP1 structure and signal transduction. Semin Cancer Biol Dec;11(6): Ernberg I, Falk K, Minarovits J, Busson P, Tursz T, Masucci MG, Klein G. (1989). The role of methylation in the phenotype dependent modulation of Epstein Barr nuclear antigen 2 and latent membrane protein genes in cells latently infected with Epstein Barr virus. J Gen Virol 70, Farnham P. (2002). Chromatin Immunprecipitation (ChIPs) protocol (Farnham Lab). Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. (1998). Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, vol 391. Fischer L, Hummel M, Korfel A, Lenze D, Joehrens K, Thiel E. (2011). Differential micro-rna expression in primary CNS and nodal diffuse large B-cell lymphomas. Neuro Oncol. (10): doi: /neuonc/nor107. Frommer M, McDonald LE, Millar DS, Collis CM, Watt F, Grigg GW, Molloy PL, Paul CL. (1992). A genomic sequencing protocol that yields a positive display of 5- methylcytosine residues in individual DNA strands. Proc. Natl. Acad. Sci. USA 89: Garrity PA, Wold BJ. (1992). Effects of different DNA polymerases in ligationmediated PCR: Enhanced genomic sequencing and in vivo footprinting. Proc. Natl. Acad. Sci. USA 89: Gibcus JH, Tan LP, Harms G, Schakel RN, de Jong D, Blokzijl T, Möller P, Poppema S, Kroesen BJ, van den Berg A. (2009). Hodgkin lymphoma cell lines are characterized by a specific mirna expression profile. Neoplasia, Vol. 11, No. 2, pp Gill G. (2005). Something about SUMO inhibits transcription. Cur. Op. in Gen. & Dev. 15: Gires O, Kohlhuber F, Kilger E, Baumann M, Kieser A, Kaiser C, Zeidler R, Scheffer B, Ueffing M, Hammerschmidt W. (1999). Latent membrane protein 1 of Epstein- Barr virus interacts with JAK3 and activates STAT proteins. EMBO J. 18: Gires O, Zimber-Strobl U, Gonnella R, Ueffing M, Marschall G, Zeidler R, Pich D, Hammerschmidt W. (1997). Latent membrane protein 1 of Epstein-Barr virus mimics a constitutively active receptor molecule. EMBO J. 16: Gruenbaum Y, Stein R, Cedar H, Razin A. (1981). Methylation of CpG sequences in eukaryotic DNA. FEBS Lett. 124: Guo S, Kemphues KJ. (1996). A non-muscle myosin required for embrionic polarity in Caenorhabditis elegans. Nature, Vol382 71
72 Haller F, von Heydebreck A, Zhang JD, Gunawan B, Langer C, Ramadori G, Wiemann S, Sahin O. (2010). Localization- and mutation-dependent microrna (mirna) expression signatures in gastrointestinal stromal tumours (GISTs), with a cluster of co-expressed mirnas located at 14q Journal of pathology and bacteriology, Vol. 220, No. 1, pp DOI: /path Hamilton AJ, Baulcombe DC. (1999). A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science, Vol Hatfull G, Bankier AT, Barrell BG, Farrell PJ. (1988). Sequence analysis of Raji Epstein-Barr virus DNA. Virology 164 (1988) He H, Jazdzewski K, Li W, Liyanarachchi S, Nagy R, Volinia S, Calin GA, Liu CG, Franssila K, Suster S, Kloos RT, Croce CM, de la Chapelle A. (2005). The role of microrna genes in papillary thyroid carcinoma. Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no. 52, pp Hodge LS, Elsawa SF, Grote DM, Price-Troska TL, Asmann YW, Fonseca R, Gertz MA, Witzig TE, Novak AJ, Ansell SM. (2011). MicroRNA expression in tumor cells from Waldenstrom's macroglobulinemia reflects both their normal and malignant cell counterparts. Blood Cancer Journal, Vol. 1, No. 6, 2011, pp. e24. DOI: /bcj Hou J, Wang P, Lin L, Liu X, Ma F, An H, Wang Z, Cao X. (2009). MicroRNA-146a Feedback Inhibits RIG-I-Dependent Type I IFN Production in Macrophages by Targeting TRAF6, IRAK1, and IRAK2. J Immunol, 183: , doi: /jimmunol Iniguez-Lluhi JA. (2006). For a healthy histone code, a little SUMO in the tail keeps the acetyl away. ACS Chem Biol. 1(4): Izumi KM, Kieff ED. (1997). The Epstein-Barr virus oncogene product latent membrane protein 1 engages the tumor necrosis factor receptor-associated death domain protein to mediate B lymphocyte growth transformation and activate NFkappaB. Proc. Natl. Acad. Sci. USA 94: Jazdzewski K, Liyanarachchi S, Swierniak M, Pachucki J, Ringel MD, Jarzab B, de la Chapelle A. (2009). Polymorphic mature micrornas from passenger strand of premir-146a contribute to thyroid cancer. Proc Natl Acad Sci USA, 106(5): doi: /pnas Kaye KM, Devergne O, Harada JN, Izumi KM, Yalamanchili R, Kieff E, Mosialos G. (1996). Tumor necrosis factor receptor associated factor 2 is a mediator of NFkappa B activation by latent infection membrane protein 1, the Epstein-Barr virus transforming protein. Proc. Natl. Acad. Sci. USA 93: Kaye, K.M., Izumi, K.M., Kieff, E. (1993). Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA 90: Kiefer JC. (2007). Epigenetics in Development. Developmental Dynamics, 236:
73 Kieff E. (1996). Epstein-Barr virus and its replication. Fields BN, Knipe DM, Howley PM (Editors) Virology, 3rd ed. Lippincott-Raven, Philadelphia: Kieser A, Kilger E, Gires O, Ueffing M, Kolch W, Hammerschmidt W. (1997). Epstein-Barr virus latent membrane protein-1 triggers AP-1 activity via the c-jun N-terminal kinase cascade. EMBO J. 16: Kitagawa N, Goto M, Kurozumi K, Maruo S, Fukayama M, Naoe T, Yasukawa M, Hino K, Suzuki T, Todo S, Takada K. (2000). Epstein-Barr virus-encoded poly(a)(-) RNA supports Burkitt s lymphoma growth through interleukin-10 induction. EMBO J. 15;19(24): Klein G. (1996). EBV-B cell interactions: Immortalization, rescue from apoptosis, tumorigenicity (A short review). Acta. Microbiol. Immunol. Hung. 43: Labbaye C, Spinello I, Quaranta MT, Pelosi E, Pasquini L, Petrucci E, Biffoni M, Nuzzolo ER, Billi M, Foà R, Brunetti E, Grignani F, Testa U, Peschle C. (2008). A three-step pathway comprising PLZF/miR-146a/CXCR4 controls megakaryopoiesis. Nature Cell Biology, vol 10, no 7, Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T. (2001). Identification of Novel Genes Coding for Small Expressed RNAs. Science 294, 853 DOI: /science Lan J, Hua S, He X, Zhang Y. (2010). DNA methyltransferases and methyl-binding proteins of mammals. Acta Biochim Biophys Sin (Shanghai). 42(4): Lawrie CH, Chi J, Taylor S, Tramonti D, Ballabio E, Palazzo S, Saunders NJ, Pezzella F, Boultwood J, Wainscoat JS, Hatton CS. (2009). Expression of micrornas in diffuse large B cell lymphoma is associated with immunophenotype, survival and transformation from follicular lymphoma. Journal of Cellular and Molecular Medicine, Vol. 13, No. 7, pp DOI: /j x. Lee KM, O'Connell MJ. (2006). A new SUMO ligase in the DNA damage response. DNA Repair 5 (2006) Lenoir GM, Vuillaume M and Bonnardel C. (1985). The use of lymphomatous and lymphoblastoid cell lines in the study of Burkitt's lymphoma. IARC Sci Publ 60, Li Y, Vandenboom TG 2nd, Wang Z, Kong D, Ali S, Philip PA, Sarkar FH. (2010). mir-146a Suppresses Invasion of Pancreatic Cancer Cells. Cancer Reasearch, 70(4), DOI: / CAN Li Z, Lu J, Sun M, Mi S, Zhang H, Luo RT, Chen P, Wang Y, Yan M, Qian Z, Neilly MB, Jin J, Zhang Y, Bohlander SK, Zhang DE, Larson RA, Le Beau MM, Thirman MJ, Golub TR, Rowley JD, Chen J. (2008). Distinct microrna expression profiles in acute myeloid leukemia with common translocations. Proceedings of the National Academy of Sciences of the United States of America, Vol. 105, No. 40, pp DOI: /pnas Liang R, Bates DJ, Wang E. (2009). Epigenetic Control of MicroRNA Expression and Aging. Current Genomics 10,
74 Liebowitz D, Wang D, Kieff E. (1986). Orientation and patching of the latent infection membrane protein encoded by Epstein-Barr virus. J. Virol. 58: Lin SL, Chiang A, Chang D, Ying SY. (2008). Loss of mir- 146a function in hormone-refractory prostate cancer. RNA, vol. 14, no. 3, pp , Lingel A, Sattler M. (2005). Novel modes of protein RNA recognition in the RNAi pathway. Current Opinion in Structural Biology. 15: , DOI /j.sbi Lu LF, Boldin MP, Chaudhry A, Lin LL, Taganov KD, Hanada T, Yoshimura A, Baltimore D, Rudensky AY. (2010). Function of mir-146a in controlling treg cellmediated regulation of Th1 responses. Cell, vol. 142, no. 6, pp , Masucci MG, Torsteindottir S, Colombani J, Brautbar C, Klein E, Klein G. (1987). Down-regulation of class I HLA antigens and of the Epstein-Barr virus-encoded latent membrane protein in Burkitt lymphoma lines. Proc Natl Acad Sci U S A; 84(13): Mazar J, Khaitan D, DeBlasio D, Zhong C, Govindarajan SS, Kopanathi S, Zhang S, Ray A, Perera RJ. (2011). Epigenetic Regulation of MicroRNA Genes and the Role of mir-34b in Cell Invasion and Motility in Human Melanoma. Pos One. Volume 6, Issue 9, e24922 Menezes J, Leibold W, Klein G, Clements G. (1975). Establishment and characterization of an Epstein-Barr virus (EBC)-negative lymphoblastoid B cell line (BJA-B) from an exceptional, EBV-genome-negative African Burkitt's lymphoma. Biomedicine 22, Millar AA, Waterhouse PM. (2005). Plant and animal micrornas: similarities and differences. Funct Integr Genomics 5: DOI /s Miller WE, Mosialos G, Kieff E, Raab-Traub N. (1997). Epstein-Barr virus LMP1 induction of the EGFR is mediated through a TRAF signaling pathway distinct from NF-κB activation. J. Virol. 71: Monticelli S, Ansel KM, Xiao C, Socci ND, Krichevsky AM, Thai TH, Rajewsky N, Marks DS, Sander C, Rajewsky K, Rao A, Kosik KS. (2005). MicroRNA profiling of the murine hematopoietic system. Genome Biology, 6:R71 (doi: /gb r71) Motsch N, Pfuhl T, Mrazek J, Barth S, Grässer FA. (2007). Epstein-Barr virusencoded latent membrane protein 1 (LMP1) induces the expression of the cellular microrna mir-146a. RNA Biology 4(3): Mueller PR, Wold B. (1989). In vivo footprinting of a muscle specific enhancer by ligation mediated PCR. Science 246: Murono S, Inoue H, Tanabe T, Joab I, Yoshizaki T, Furukawa M, Pagano JS. (2001). Induction of cycloxygenase-2 by Epstein-Barr virus latent membrane protein 1 is involved in vascular endothelial growth factor production in nasopharyngeal carcinoma cells. Proc. Natl. Acad. Sci. USA 98:
75 Nahid MA, Pauley KM, Satoh M, Chan EK. (2009). mir-146a Is Critical for Endotoxin-induced Tolerance. The Journal of Biological Chemistry Vol. 284, no. 50, pp Nanbo A, Takada K. (2002). The role of Epstein Barr virus-encoded small RNAs (EBERs) in oncogenesis. Rev. Med. Virol. 12: Napoli C, Lemieux C, Jorgensen R. (1990). Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans. The Plant Cell, Vol. 2, , April 1990 Niller HH, Glaser G, Knüchel R, Wolf H. (1995). Nucleoprotein complexes and DNA 5 -ends at orip of Epstein-Barr virus. J. Biol. Chem. 270: Onnis A, De Falco G, Antonicelli G, Onorati M, Bellan C, Sherman O, Sayed S, Leoncini L. (2010). Alteration of micrornas regulated by c-myc in Burkitt Lymphoma. PLoS ONE, Vol. 5, No. 9, DOI: /journal.pone Pai S, O'Sullivan BJ, Cooper L, Thomas R, Khanna R. (2002). RelB nuclear translocation mediated by C-terminal activator regions of Epstein-Barr virusencoded latent membrane protein 1 and its effect on antigen-presenting function in B cells. J. Virol. 76(4): Park SM, Gaur AB, Lengyel E, Peter ME. (2008). The mir-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Development, Vol. 22, No. 7, pp DOI: /gad Park SY, Lee JH, Ha M, Nam JW, Kim VN. (2009). mir-29 mirnas activate p53 by targeting p85 alpha and CDC42. Nature structural and molecular biology, Vol. 16, No. 1, pp DOI: /nsmb Patrone G, Puppo F, Cusano R, Scaranari M, Ceccherini I, Puliti A, Ravazzolo R. (2000). Nuclear run-on assay using biotin labeling, magnetic bead capture and analysis by fluorescence-based RT-PCR. Biotechniques. 29(5):1012-4, Perry MM, Moschos SA, Williams AE, Shepherd NJ, Larner-Svensson HM, Lindsay MA. (2008). Rapid changes in microrna-146a expression negatively regulate the interleukin-1β induced inflammatory response in human lung alveolar epithelial cells. J Immunol April 15; 180(8): Piccaluga PP, De Falco G, Kustagi M, Gazzola A, Agostinelli C, Tripodo C, Leucci E, Onnis A, Astolfi A, Sapienza MR, Bellan C, Lazzi S, Tumwine L, Mawanda M, Ogwang M, Calbi V, Formica S, Califano A, Pileri SA and Leoncini L. (2011). Gene expression analysis uncovers similarity and differences among Burkitt lymphoma subtypes. Blood 117, Plasterk RH. (2002). RNA Silencing: The Genome s Immune System. Science Vol 296 Pontes O, Pikaard CS. (2008). sirna and mirna processing: new functions for Cajal bodies. Current Opinion in Genetics & Development 18:1 7, DOI /j.gde
76 Prendergast GC, Lawe D, Ziff EB. (1991). Association of Myn, the murin homolog of max, with c-myc stimulates methylation-sensitive DNA binding and ras cotransformation. Cell 65: Rai D, Karanti S, Jung I, Dahia PL, Aguiar RC. (2008). Coordinated expression of microrna-155 and predicted target genes in diffuse large B-cell lymphoma. Cancer Genetics and Cytogenetics, Vol. 181, No. 1, pp DOI: /j.cancergencyto Razin A, Riggs D. (1980). DNA methylation and gene function. Science 210: Robertus JL, Kluiver J, Weggemans C, Harms G, Reijmers RM, Swart Y, Kok K, Rosati S, Schuuring E, van Imhoff G, Pals ST, Kluin P, van den Berg A. (2010). MiRNA profiling in B non-hodgkin lymphoma: a MYC-related mirna profile characterizes Burkitt lymphoma. British journal of haematology, Vol. 149, No. 6, pp DOI: /j x. Roccaro AM, Sacco A, Chen C, Runnels J, Leleu X, Azab F, Azab AK, Jia X, Ngo HT, Melhem MR, Burwick N, Varticovski L, Novina CD, Rollins BJ, Anderson KC, Ghobrial IM. (2009). microrna expression in the biology, prognosis, and therapy of Waldenström macroglobulinemia. Blood, Vol. 113, No. 18, pp DOI: /blood Roehle A, Hoefig KP, Repsilber D, Thorns C, Ziepert M, Wesche KO, Thiere M, Loeffler M, Klapper W, Pfreundschuh M, Matolcsy A, Bernd HW, Reiniger L, Merz H, Feller AC. (2008). MicroRNA signatures characterize diffuse large B-cell lymphomas and follicular lymphomas. British journal of haematology, Vol. 142, No. 5, pp DOI: /j x. Rosato P, Anastasiadou E, Garg N, Lenze D, Boccellato F, Vincenti S, Severa M, Coccia EM, Bigi R, Cirone M, Ferretti E, Campese AF, Hummel M, Frati L, Presutti C, Faggioni A, Trivedi P. (2012). Differential regulation of mir-21 and mir-146a by Epstein-Barr virus-encoded EBNA2. Leukemia 26(11): doi: /leu Rusca, N. and Monticelli, S. (2011). MiR-146a in immunity and disease. SAGE- Hindawi Access to Research Molecular Biology International. Vol 2011, Article ID , doi: /2011/ Sanchez-Elsner T, Gou D, Kremmer E, Sauer F. (2006). Noncoding RNAs of trithorax response elements recruit Drosophila Ash1 to Ultrabithorax. Science, 311(5764): Sandhu SK, Croce CM, Garzon R. (2011). Micro-RNA Expression and Function in Lymphomas. Advances in Hematology, Vol Article ID , 12 pages doi: /2011/ Santos-Rosa H, Caldas C. (2005). Chromatin modifier enzymes, the histone code and cancer. Eur. J. of Cancer. 41, Shabalina SA, Koonin EV. (2008). Origins and evolution of eukaryotic RNA interference. Trends Ecol Evol. 23(10): doi: /j.tree
77 Strahl BD, Allis CD. (2000). The language of covalent histone modifications. Nature 403: Szenthe K, Koroknai A, Banati F, Bathori Z, Helmut Niller H, Wolf H, Nagy N, Klein E, Minarovits J, Salamon D. (2013c). The role of DNA hypomethylation, histone acetylation and in vivo protein-dna binding in Epstein-Barr virus-induced CD23 upregulation. Biochem Biophys Res Commun. 435(1):8-15. doi: /j.bbrc Szenthe K, Koroknai A, Banati F, Bathori Z, Lozsa R, Burgyan J, Wolf H, Salamon D, Nagy K, Niller HH, Minarovits J. (2013b). The 5' regulatory sequences of active mir-146a promoters are hypomethylated and associated with euchromatic histone modification marks in B lymphoid cells. Biochem Biophys Res Commun. 433(4): Szenthe K, Nagy K, Buzas K, Niller HH, Minarovits J. (2013a). MicroRNAs as targets and tools in B-cell lymphoma therapy. Journal of Cancer Therapy 4: DOI: /jct A057 Szittya G, Molnár A, Silhavy D, Hornyik C, Burgyán J. (2002). Short defective interfering RNAs of tombusviruses are not targeted but trigger posttranscriptional gene silencing against their helper virus. Plant Cell, 14(2): Taganov KD, Boldin MP, Chang KJ, Baltimore D. (2006). NF-kappaB-dependent induction of microrna mir-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A., 15;103(33): Takacs M, Banati F, Koroknai A, Segesdi J, Salamon D, Wolf H, Niller HH, Minarovits J. (2010). Epigenetic regulation of latent Epstein-Barr virus promoters. Biochim. Biophys. Acta 1799 (2010) Takada K, Horinouchi K, Ono Y, Aya T, Osato T, Takahashi M, Hayasaka S. (1991). An Epstein-Barr virus-producer line Akata: establishment of the cell line and analysis of viral DNA. Virus Genes 5 (1991) Tsai HC, Baylin SB. (2011). Cancer epigenetics: linking basic biology to clinical medicine. Cell Research 21: Van Driel R, Fransz P. (2004). Nuclear architecture and genome functioning in plants and animals: what can we learn from both? Exp. Cell. Res. 296 (2004) Verdel A, Vavasseur A, Le Gorrec M, Touat-Todeschini L. (2009). Common themes in sirna-mediated epigenetic silencing pathways. Int. J. Dev. Biol. 53: doi: /ijdb av Verdone L, Caserta M and Di Mauro E. (2005). Role of histone acetylation in the control of gene expression. Biochem Cell Biol 83, Viré E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, Morey L, Van Eynde A, Bernard D, Vanderwinden JM, Bollen M, Esteller M, Di Croce L, de Launoit Y, Fuks F. (2006). The Polycomb group protein EZH2 directly controls DNA methylation. Nature, 439 (7078):
78 Wakisaka N, Kondo S, Yoshizaki T, Murono S, Furukawa M, Pagano JS. (2004). Epstein-Barr virus latent membrane protein 1 induces synthesis of hypoxiainducible factor 1 alpha. Mol. Cell. Biol. 24 (2004) Wang X, Tang S, Le SY, Lu R, Rader JS, Meyers C, Zheng ZM. (2008). Aberrant expression of oncogenic and tumor-suppressive micrornas in cervical cancer is required for cancer cell growth. PLoS ONE, vol. 3, no. 7, article e2557. Wu Y, Kikuchi S, Yan H, Zhang W, Rosenbaum H, Iniquez AL, Jiang J. (2011b) Euchromatic subdomains in rice centromers are associated with genes and transcription. Plant Cell 23 (2011b) Xiao C, Calado DP, Galler G, Thai TH, Patterson HC, Wang J, Rajewsky N, Bender TP, Rajewsky K. (2007). MiR-150 controls B cell differentiation by targeting the transcription factor c-myb. Cell, Vol. 131, No. 1, pp Yoshizaki T, Sato H, Furukawa M, Pagano JS. (1998). The expression of matrix metalloproteinase 9 is enhanced by Epstein-Barr virus latent membrane protein 1. Proc. Natl. Acad. Sci. USA 95: Zhao JJ, Lin J, Lwin T, Yang H, Guo J, Kong W, Dessureault S, Moscinski LC, Rezania D, Dalton WS, Sotomayor E, Tao J, Cheng JQ. (2010). MicroRNA expression profile and identification of mir-29 as a prognostic marker and pathogenetic factor by targeting CDK6 in mantle cell lymphoma. Blood, Vol. 115, No. 13, pp DOI: /blood
 Jelmagyarázat: Oct-1 kötőhely NFκB HSF2 PU.1 c-ets STAT RBPJκ cacgtg c-myc kötőhely Dőlt, szürke betűs területen: feltételezett c-myc kötőhely (Cameron és mtsai.")
79 MELLÉKLET mir-146a szekvencia LOCUS: NT_023133, bp, DNA, linear (a ChIP módszerhez felhasznált primerek kötőhelyeivel és a footprinting módszer esetében vizsgált in silico kimutatott faktorok kötőhelyei) Jelmagyarázat: Oct-1 kötőhely NFκB HSF2 PU.1 c-ets STAT RBPJκ cacgtg c-myc kötőhely Dőlt, szürke betűs területen: feltételezett c-myc kötőhely (Cameron és mtsai., 2008) Hosszan aláhúzott régió: exonok, közte kihagyással intron, Piros betűvel a hisztonmodifikációs vizsgálatban felhasznált primerek szekvenciája (ld.: 15. fejezet) 79

80 80
81 81
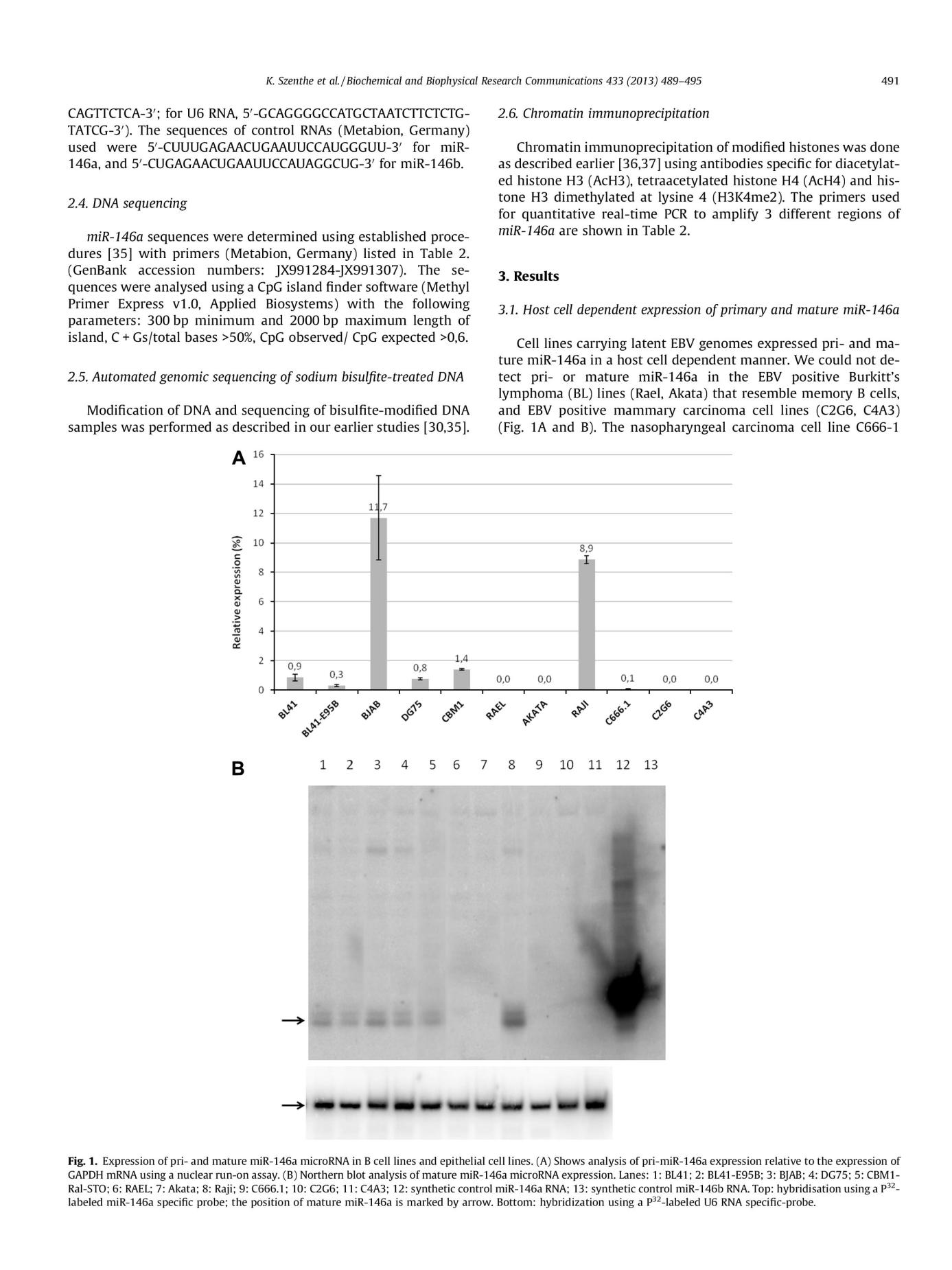
82 82
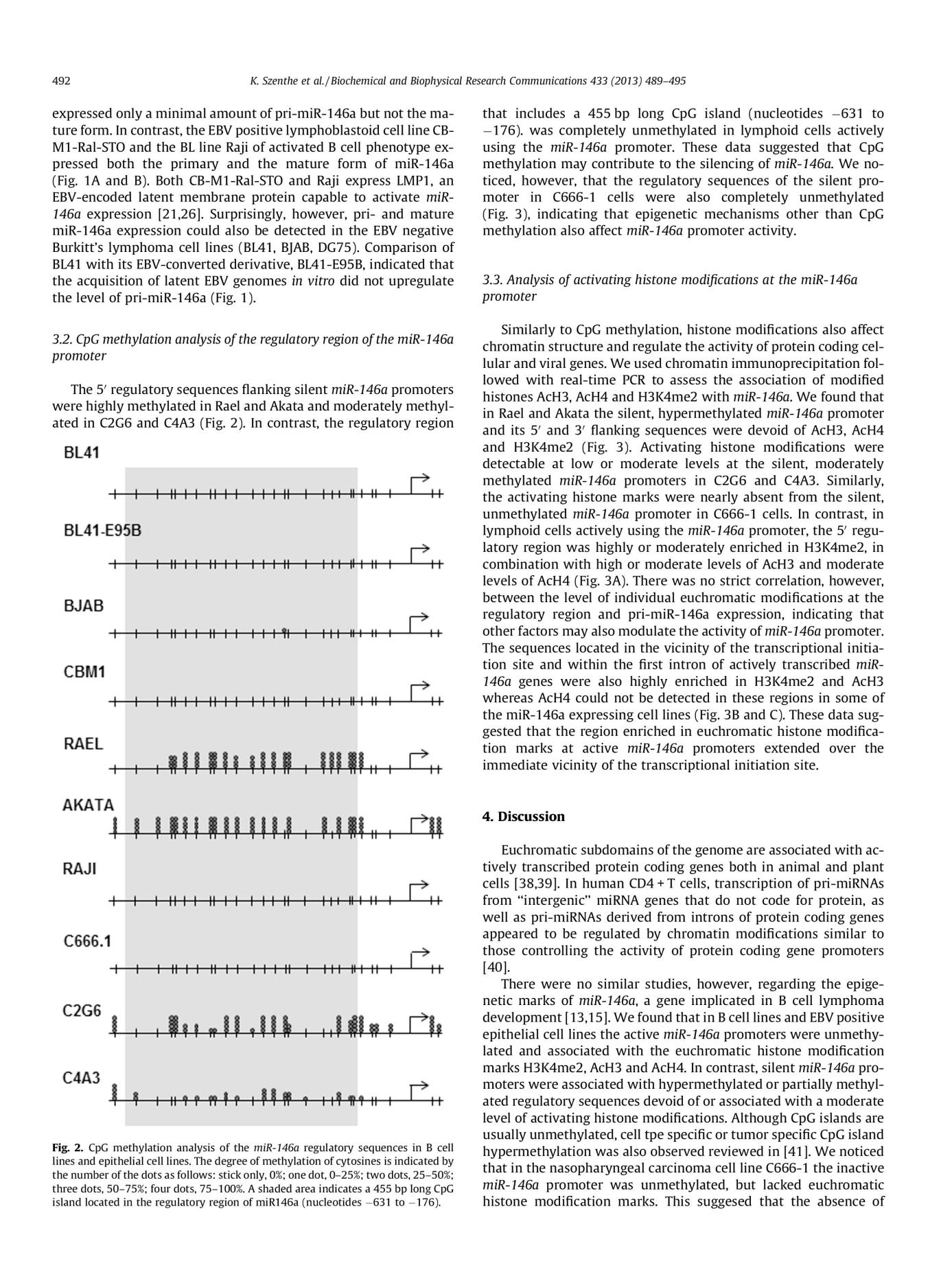
83 83
84 84

85 85

86 86

87 87

88 88
89 89
90 90
91 91
92 92

93 93

94 94
95 95

96 96
97 97

98 98
99 99
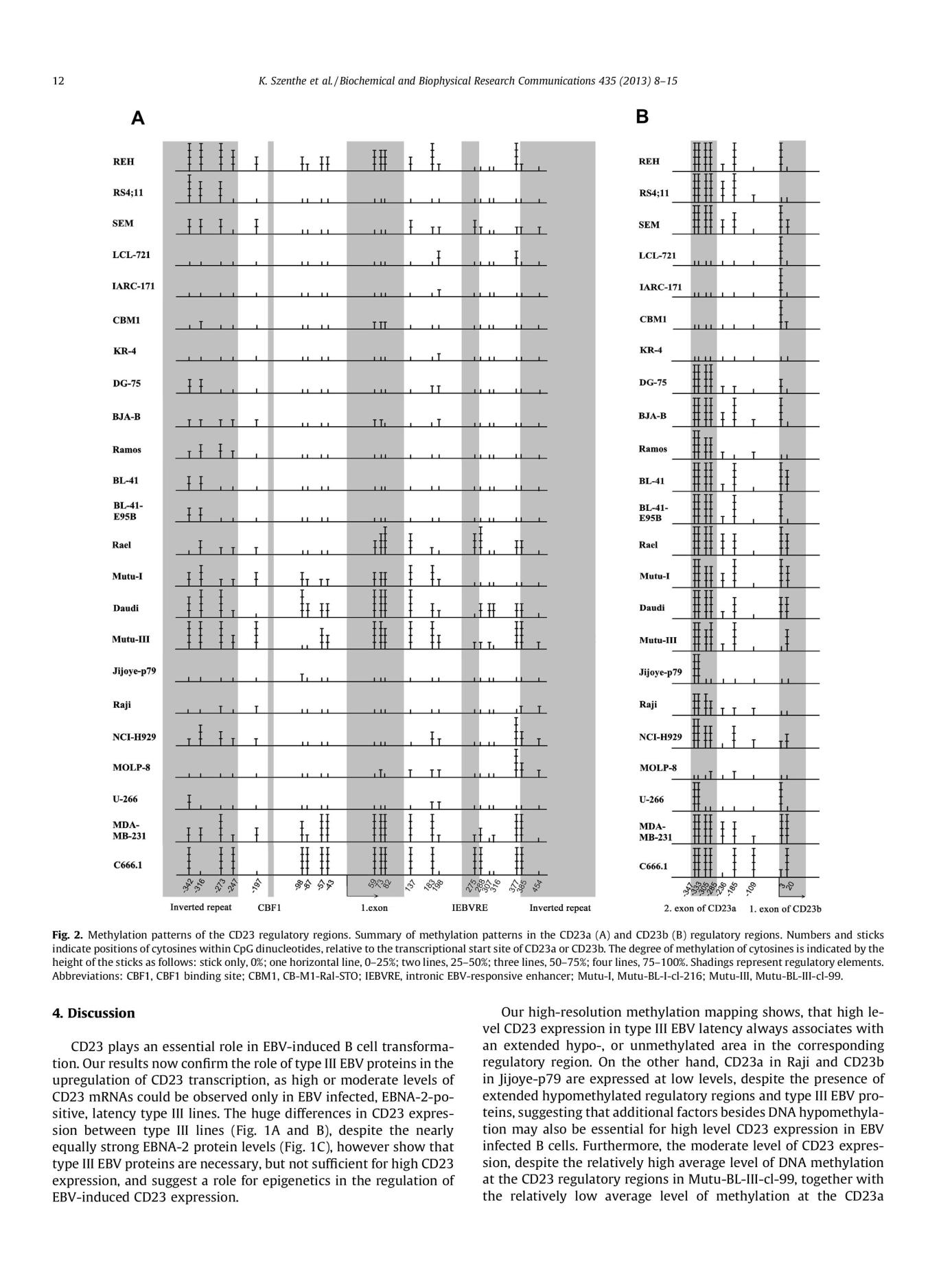
100 100
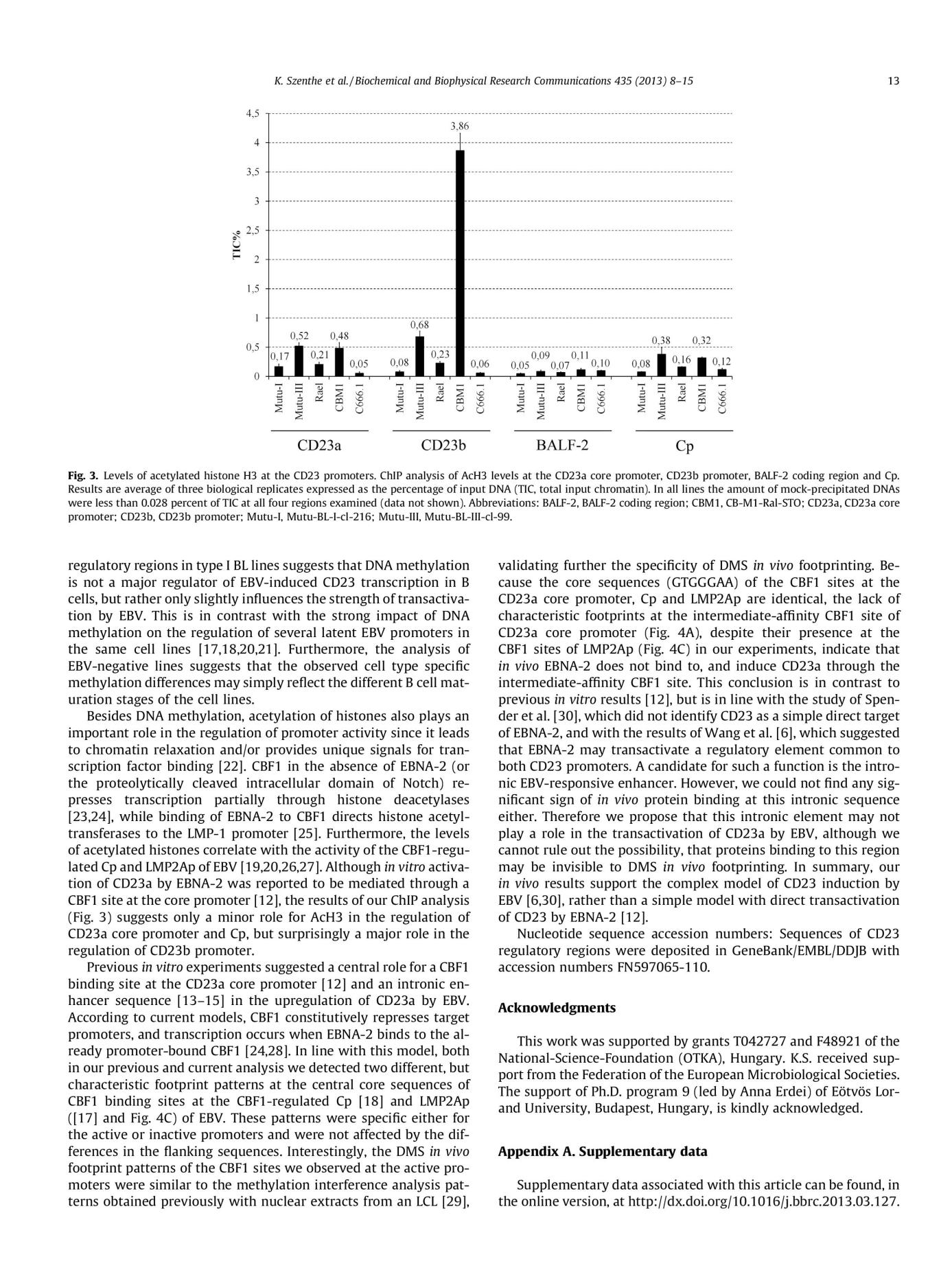
101 101

102 102
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata
 sejtek mikro RNS (mir) profiljának vizsgálata") Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
TÉMAKÖRÖK. Ősi RNS világ BEVEZETÉS. RNS-ek tradicionális szerepben
 esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában
 Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában Czimmerer Zsolt, Csenki-Bakos Zsolt, Urbányi Béla TOX 2017 Tudományos Konferencia 2017.10.12. Bükfürdő A sejtek
Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában Czimmerer Zsolt, Csenki-Bakos Zsolt, Urbányi Béla TOX 2017 Tudományos Konferencia 2017.10.12. Bükfürdő A sejtek
Hamar Péter. RNS világ. Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. www.meetthescientist.hu 1 26
 Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei)
") Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Epigenetikai Szabályozás
 Epigenetikai Szabályozás Kromatin alapegysége a nukleoszóma 1. DNS Linker DNS Nukleoszóma mag H1 DNS 10 nm 30 nm Nukleoszóma gyöngy (4x2 hiszton molekula + 146 nukleotid pár) 10 nm-es szál 30 nm-es szál
Epigenetikai Szabályozás Kromatin alapegysége a nukleoszóma 1. DNS Linker DNS Nukleoszóma mag H1 DNS 10 nm 30 nm Nukleoszóma gyöngy (4x2 hiszton molekula + 146 nukleotid pár) 10 nm-es szál 30 nm-es szál
Az X kromoszóma inaktívációja. A kromatin szerkezet befolyásolja a génexpressziót
 Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL
 Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Norvég Finanszírozási Mechanizmus által támogatott projekt HU-0115/NA/2008-3/ÖP-9 ÚJ TERÁPIÁS CÉLPONTOK AZONOSÍTÁSA GENOMIKAI MÓDSZEREKKEL KÖZÖS STRATÉGIA KIFEJLESZTÉSE MOLEKULÁRIS MÓDSZEREK ALKALMAZÁSÁVAL
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
DNS replikáció. DNS RNS Polipeptid Amino terminus. Karboxi terminus. Templát szál
 DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
- Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=>
, így próbálta leírni a sejt specializációt=>") 1 - Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=> a sejtek sorsa meg van határozva, mint egy üveggolyó,
1 - Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=> a sejtek sorsa meg van határozva, mint egy üveggolyó,
Immunológia alapjai előadás. A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és
 Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE
 Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Immunológia I. 4. előadás. Kacskovics Imre
 Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az adaptív immunválasz kialakulása. Erdei Anna Immunológiai Tanszék ELTE
 Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció 5.
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció 5.
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Epigenetikai szabályozás
 Epigenetikai szabályozás 1. A Transzkripció epigenetikai szabályozása Dia 1 A kromatin szerkezete A sejtciklus S fázisában egyetlen sejtben kb 4x1 méter hosszúságú DNS található 4x23 darabra vágva (4N
Epigenetikai szabályozás 1. A Transzkripció epigenetikai szabályozása Dia 1 A kromatin szerkezete A sejtciklus S fázisában egyetlen sejtben kb 4x1 méter hosszúságú DNS található 4x23 darabra vágva (4N
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere
 I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere Kopper László A fejezet célja, hogy megismerje a hallgató a daganatok kialakulásában szerepet
I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere Kopper László A fejezet célja, hogy megismerje a hallgató a daganatok kialakulásában szerepet
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
A preventív vakcináció lényege :
 Vakcináció Célja: antigénspecifkus immunválasz kiváltása a szervezetben A vakcina egy olyan készítmény, amely fokozza az immunitást egy adott betegséggel szemben (aktiválja az immunrendszert). A preventív
Vakcináció Célja: antigénspecifkus immunválasz kiváltása a szervezetben A vakcina egy olyan készítmény, amely fokozza az immunitást egy adott betegséggel szemben (aktiválja az immunrendszert). A preventív
Antigén, Antigén prezentáció
 Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
http://www.rimm.dote.hu Tumor immunológia
 http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
http://www.rimm.dote.hu Tumor immunológia A tumorok és az immunrendszer kapcsolatai Tumorspecifikus és tumorasszociált antigének A tumor sejteket ölő sejtek és mechanizmusok Az immunológiai felügyelet
Sejtmag, magvacska magmembrán
 Sejtmag, magvacska magmembrán Láng Orsolya Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Kompartmentalizáció Prokaryóta Cytoplazma Eukaryóta Endomembrán Kromatin Plazma membrán Eredménye
Sejtmag, magvacska magmembrán Láng Orsolya Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Kompartmentalizáció Prokaryóta Cytoplazma Eukaryóta Endomembrán Kromatin Plazma membrán Eredménye
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Daganatok kialakulásában szerepet játszó molekuláris folyamatok
 Daganatok kialakulásában szerepet játszó molekuláris folyamatok Genetikai változások Onkogének Tumorszuppresszor gének DNS hibajavító gének Telomer és telomeráz Epigenetikai változások DNS-metiláció Mikro-RNS
Daganatok kialakulásában szerepet játszó molekuláris folyamatok Genetikai változások Onkogének Tumorszuppresszor gének DNS hibajavító gének Telomer és telomeráz Epigenetikai változások DNS-metiláció Mikro-RNS
Doktori értekezés tézisei
 Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Doktori értekezés tézisei A komplement- és a Toll-szerű receptorok kifejeződése és szerepe emberi B-sejteken fiziológiás és autoimmun körülmények között - az adaptív és a természetes immunválasz kapcsolata
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
ADATBÁNYÁSZAT I. ÉS OMICS
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ADATBÁNYÁSZAT
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ADATBÁNYÁSZAT
INTRACELLULÁRIS PATOGÉNEK
 INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
INTRACELLULÁRIS PATOGÉNEK Bácsi Attila, PhD, DSc etele@med.unideb.hu Debreceni Egyetem, ÁOK Immunológiai Intézet INTRACELLULÁRIS BAKTÉRIUMOK ELLENI IMMUNVÁLASZ Példák intracelluláris baktériumokra Intracelluláris
3. Főbb Jelutak. 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3.
 Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
B-sejtek szerepe az RA patológiás folyamataiban
 B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
B-sejtek szerepe az RA patológiás folyamataiban Erdei Anna Biológiai Intézet Immunológiai Tanszék Eötvös Loránd Tudományegyetem Immunológiai Tanszék ORFI, Helia, 2015 április 17. RA kialakulása Gary S.
Az Oxidatív stressz hatása a PIBF receptor alegységek összeszerelődésére.
 Újabban világossá vált, hogy a Progesterone-induced blocking factor (PIBF) amely a progesteron számos immunológiai hatását közvetíti, nem csupán a lymphocytákban és terhességgel asszociált szövetekben,
Újabban világossá vált, hogy a Progesterone-induced blocking factor (PIBF) amely a progesteron számos immunológiai hatását közvetíti, nem csupán a lymphocytákban és terhességgel asszociált szövetekben,
TDK lehetőségek az MTA TTK Enzimológiai Intézetben
 TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
Látens Epstein-Barr vírus genomok szabályozó régióinak epigenetikai jellemzése limfoid és nazofaringeális karcinóma sejtekben.
 Biológia Doktori Iskola, Immunológia Doktori Program Eötvös Loránd Tudományegyetem, Természettudományi Kar Doktori Iskola- és Program vezetője: Prof. Erdei Anna Látens Epstein-Barr vírus genomok szabályozó
Biológia Doktori Iskola, Immunológia Doktori Program Eötvös Loránd Tudományegyetem, Természettudományi Kar Doktori Iskola- és Program vezetője: Prof. Erdei Anna Látens Epstein-Barr vírus genomok szabályozó
Összefoglalás első fejezete
 Összefoglalás Az utóbbi években a transzkriptómikai vizsgálatokban alkalmazott technológiák fejlődésének köszönhetően a genom kutatás területén jelentős előrehaladás figyelhető meg. A disszertáció első
Összefoglalás Az utóbbi években a transzkriptómikai vizsgálatokban alkalmazott technológiák fejlődésének köszönhetően a genom kutatás területén jelentős előrehaladás figyelhető meg. A disszertáció első
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A TATA-kötő fehérje asszociált faktor 3 (TAF3) p53-mal való kölcsönhatásának funkcionális vizsgálata
 p53-mal való kölcsönhatásának funkcionális vizsgálata") Ph.D. ÉRTEKEZÉS TÉZISEI A TATA-kötő fehérje asszociált faktor 3 (TAF3) p53-mal való kölcsönhatásának funkcionális vizsgálata Buzás-Bereczki Orsolya Témavezetők: Dr. Bálint Éva Dr. Boros Imre Miklós Biológia
Ph.D. ÉRTEKEZÉS TÉZISEI A TATA-kötő fehérje asszociált faktor 3 (TAF3) p53-mal való kölcsönhatásának funkcionális vizsgálata Buzás-Bereczki Orsolya Témavezetők: Dr. Bálint Éva Dr. Boros Imre Miklós Biológia
AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA. Balicza-Himer Leonóra. Doktori értekezés tézisei
 AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA Doktori értekezés tézisei Balicza-Himer Leonóra Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola / Immunológia
AZ ADENOZIN HATÁSA A PERIFÉRIÁS CD4 + T-LIMFOCITÁK AKTIVÁCIÓJÁRA Doktori értekezés tézisei Balicza-Himer Leonóra Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola / Immunológia
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Preeclampsia-asszociált extracelluláris vezikulák
 Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
Preeclampsia-asszociált extracelluláris vezikulák hatása(i) a monocita sejt működésére Kovács Árpád Ferenc 1, Láng Orsolya 1, Kőhidai László 1, Rigó János 2, Turiák Lilla 3, Fekete Nóra 1, Buzás Edit 1,
Immunológia I. 2. előadás. Kacskovics Imre (imre.kacskovics@ttk.elte.hu)
") Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
OTKA ZÁRÓJELENTÉS (F48921)
") OTKA ZÁRÓJELENTÉS (F48921) Salamon Dániel F48921 1. A korábbi munkáinkban leírt, jól karakterizált látens Epstein-Barr vírus (EBV) genomokat hordozó, I. látencia-típusú Rael és Mutu-BL-I-cl-216 Burkitt
OTKA ZÁRÓJELENTÉS (F48921) Salamon Dániel F48921 1. A korábbi munkáinkban leírt, jól karakterizált látens Epstein-Barr vírus (EBV) genomokat hordozó, I. látencia-típusú Rael és Mutu-BL-I-cl-216 Burkitt
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogénekimmunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
MUTÁCIÓK. A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik.
 MUTÁCIÓK A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik. Pontmutáció: A kromoszóma egy génjében pár nukleotidnál következik be változás.
MUTÁCIÓK A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik. Pontmutáció: A kromoszóma egy génjében pár nukleotidnál következik be változás.
Intézeti Beszámoló. Dr. Kovács Árpád Ferenc
 Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Intézeti Beszámoló Dr. Kovács Árpád Ferenc 2015.12.03 135 millió újszülött 10 millió újszülött Preeclampsia kialakulása kezdetét veszi Preeclampsia tüneteinek megjelenése Preeclampsia okozta koraszülés
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
I. A sejttől a génekig
 Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
NÖVÉNYÉLETTAN. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010
 NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Sejtfal szintézis és megnyúlás Környezeti tényezők hatása a növények növekedésére és fejlődésére Előadás áttekintése
NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Sejtfal szintézis és megnyúlás Környezeti tényezők hatása a növények növekedésére és fejlődésére Előadás áttekintése
Biológus MSc. Molekuláris biológiai alapismeretek
 Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Az immunrendszer alapjai, sejtöregedés, tumorképződés. Biológiai alapismeretek
 Az immunrendszer alapjai, sejtöregedés, tumorképződés Biológiai alapismeretek Az immunrendszer Immunis (latin szó): jelentése mentes valamitől Feladata: a szervezetbe került idegen anyagok: 1. megtalálása
Az immunrendszer alapjai, sejtöregedés, tumorképződés Biológiai alapismeretek Az immunrendszer Immunis (latin szó): jelentése mentes valamitől Feladata: a szervezetbe került idegen anyagok: 1. megtalálása
A szamóca érése során izolált Spiral és Spermidin-szintáz gén jellemzése. Kiss Erzsébet Kovács László
 A szamóca érése során izolált Spiral és Spermidin-szintáz gén jellemzése Kiss Erzsébet Kovács László Bevezetés Nagy gazdasági gi jelentıségük k miatt a gyümölcs lcsök, termések fejlıdésének mechanizmusát
A szamóca érése során izolált Spiral és Spermidin-szintáz gén jellemzése Kiss Erzsébet Kovács László Bevezetés Nagy gazdasági gi jelentıségük k miatt a gyümölcs lcsök, termések fejlıdésének mechanizmusát
Az ellenanyagok szerkezete és funkciója. Erdei Anna Immunológiai Tanszék ELTE
 Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Szervezetünk védelmének alapja: az immunológiai felismerés
 Szervezetünk védelmének alapja: az immunológiai felismerés Erdei Anna ELTE, TTK, Biológiai Intézet Immunológiai Tanszék ELTE, Pázmány-nap, 2012. Az immunrendszer fő feladata a gazdaszervezet védelme a
Szervezetünk védelmének alapja: az immunológiai felismerés Erdei Anna ELTE, TTK, Biológiai Intézet Immunológiai Tanszék ELTE, Pázmány-nap, 2012. Az immunrendszer fő feladata a gazdaszervezet védelme a
2007/11/05 Molekuláris biológia előadások - Putnoky 1-1
 1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
Mit tud a genetika. Génterápiás lehetőségek MPS-ben. Dr. Varga Norbert
 Mit tud a genetika Génterápiás lehetőségek MPS-ben Dr. Varga Norbert Oki terápia Terápiás lehetőségek MPS-ben A kiváltó okot gyógyítja meg ERT Enzimpótló kezelés Őssejt transzplantáció Genetikai beavatkozások
Mit tud a genetika Génterápiás lehetőségek MPS-ben Dr. Varga Norbert Oki terápia Terápiás lehetőségek MPS-ben A kiváltó okot gyógyítja meg ERT Enzimpótló kezelés Őssejt transzplantáció Genetikai beavatkozások
sejt működés jovo.notebook March 13, 2018
 1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN
 2016. 10. 14. KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN Kovács Árpád Ferenc 1, Pap Erna 1, Fekete Nóra 1, Rigó János 2, Buzás Edit 1,
2016. 10. 14. KERINGŐ EXTRACELLULÁRIS VEZIKULÁK ÁLTAL INDUKÁLT GÉNEXPRESSZIÓS MINTÁZAT VIZSGÁLATA TROPHOBLAST SEJTVONALBAN Kovács Árpád Ferenc 1, Pap Erna 1, Fekete Nóra 1, Rigó János 2, Buzás Edit 1,
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
DOKTORI ÉRTEKEZÉS TÉZISEI. Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben.
 DOKTORI ÉRTEKEZÉS TÉZISEI Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben Tóth Mónika Témavezető: Dr. Bálint Éva SZTE TTIK Biológia Doktori Iskola
DOKTORI ÉRTEKEZÉS TÉZISEI Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben Tóth Mónika Témavezető: Dr. Bálint Éva SZTE TTIK Biológia Doktori Iskola
Szinoviális szarkómák patogenezisében szerepet játszó szignálutak jellemzése szöveti multiblokk technika alkalmazásával
 Szinoviális szarkómák patogenezisében szerepet játszó szignálutak jellemzése szöveti multiblokk technika alkalmazásával Fónyad László 1, Moskovszky Linda 1, Szemlaky Zsuzsanna 1, Szendrői Miklós 2, Sápi
Szinoviális szarkómák patogenezisében szerepet játszó szignálutak jellemzése szöveti multiblokk technika alkalmazásával Fónyad László 1, Moskovszky Linda 1, Szemlaky Zsuzsanna 1, Szendrői Miklós 2, Sápi
Az ADA2b adaptor fehérjéket tartalmazó hiszton acetiltranszferáz komplexek szerepének vizsgálata Drosophila melanogaster-ben
 Az ADA2b adaptor fehérjéket tartalmazó hiszton acetiltranszferáz komplexek szerepének vizsgálata Drosophila melanogaster-ben DOKTORI TÉZIS Pankotai Tibor Témavezető: Dr. Boros Imre Miklós Szeged, 2007
Az ADA2b adaptor fehérjéket tartalmazó hiszton acetiltranszferáz komplexek szerepének vizsgálata Drosophila melanogaster-ben DOKTORI TÉZIS Pankotai Tibor Témavezető: Dr. Boros Imre Miklós Szeged, 2007
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a PD-L1 és PD-1 fehérjék túlélésre gyakorolt hatása
 Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
A biológia szerepe az egészségvédelemben
 A biológia szerepe az egészségvédelemben Nagy Kinga nagy.kinga@mail.bme.hu 2017.10.24 Mikróbák az ember szolgálatában (Néhány példán keresztül bemutatva) Antibiotikumok (gombák, baktériumok) Restrikciós
A biológia szerepe az egészségvédelemben Nagy Kinga nagy.kinga@mail.bme.hu 2017.10.24 Mikróbák az ember szolgálatában (Néhány példán keresztül bemutatva) Antibiotikumok (gombák, baktériumok) Restrikciós
Az ABCG2 multidrog transzporter fehérje szerkezetének és működésének vizsgálata
 Az ABCG2 multidrog transzporter fehérje szerkezetének és működésének Kutatási előzmények Az ABC transzporter membránfehérjék az ATP elhasítása (ATPáz aktivitás) révén nyerik az energiát az általuk végzett
Az ABCG2 multidrog transzporter fehérje szerkezetének és működésének Kutatási előzmények Az ABC transzporter membránfehérjék az ATP elhasítása (ATPáz aktivitás) révén nyerik az energiát az általuk végzett
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogének immunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogének immunmoduláns hatásai Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun betegségek,
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogének immunmoduláns hatásai Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun betegségek,
FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére FEHÉRJE VAKCINÁK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA III.
A genomiális medicina szép új világa
 A genomiális medicina szép új világa HALADÁS A REUMATOLÓGIA, IMMUNOLÓGIA ÉS OSTEOLÓGIA TERÜLETÉN 2012 2014 2015. ÁPRILIS 16. Falus András Semmelweis Egyetem Genetikai Sejt és Immunbiológiai Intézet A medicina
A genomiális medicina szép új világa HALADÁS A REUMATOLÓGIA, IMMUNOLÓGIA ÉS OSTEOLÓGIA TERÜLETÉN 2012 2014 2015. ÁPRILIS 16. Falus András Semmelweis Egyetem Genetikai Sejt és Immunbiológiai Intézet A medicina
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Válasz Prof. Szabó Dóra opponensi véleményére
 Válasz Prof. Szabó Dóra opponensi véleményére Köszönöm Szabó Dóra Professzor Nőnek a doktori értekezésem gondos áttanulmányozását, az újdonságok kiemelését és támogatását a doktori cím odaítéléséhez. Az
Válasz Prof. Szabó Dóra opponensi véleményére Köszönöm Szabó Dóra Professzor Nőnek a doktori értekezésem gondos áttanulmányozását, az újdonságok kiemelését és támogatását a doktori cím odaítéléséhez. Az
A rák, mint genetikai betegség
 A rák, mint genetikai betegség Diák: Ferencz Arnold-Béla la Felkész szítı tanár: József J Éva Bolyai Farkas Elméleti leti LíceumL Mi is a rák r tulajdonképpen? A rák r k egy olyan betegség g ahol sejt
A rák, mint genetikai betegség Diák: Ferencz Arnold-Béla la Felkész szítı tanár: József J Éva Bolyai Farkas Elméleti leti LíceumL Mi is a rák r tulajdonképpen? A rák r k egy olyan betegség g ahol sejt
A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet
 A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet Mi a feladata az Immunrendszernek? 1. Védelem a kórokozók ellen 2. Immuntolerancia fenntartása Mik is azok a kórokozók? Kórokozók alatt
A csodálatos Immunrendszer Lányi Árpád, DE, Immunológiai Intézet Mi a feladata az Immunrendszernek? 1. Védelem a kórokozók ellen 2. Immuntolerancia fenntartása Mik is azok a kórokozók? Kórokozók alatt
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van.
 In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van. Kneif Józsefné PTE KK Pathologiai Intézet Budapest 2017. 05. 26 Kromoszóma rendellenesség kimutatás PCR technika: izolált nukleinsavak
In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van. Kneif Józsefné PTE KK Pathologiai Intézet Budapest 2017. 05. 26 Kromoszóma rendellenesség kimutatás PCR technika: izolált nukleinsavak
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
Epigenetika kihívások az ökotoxikológiában
 Epigenetika kihívások az ökotoxikológiában Bakonyi Gábor és Szabó Borbála Szent István Egyetem, Gödöllő Állattani és Állatökológiai Tanszék V. ÖKOTOXIKOLÓGIAI KONFERENCIA, MAGYAR ÖKOTOXIKOLÓGIAI TÁRSASÁG,
Epigenetika kihívások az ökotoxikológiában Bakonyi Gábor és Szabó Borbála Szent István Egyetem, Gödöllő Állattani és Állatökológiai Tanszék V. ÖKOTOXIKOLÓGIAI KONFERENCIA, MAGYAR ÖKOTOXIKOLÓGIAI TÁRSASÁG,