Fehérjék. A fehérjék szerkezeti szintjei. Elsődleges szerkezet
|
|
|
- Ádám Bognár
- 5 évvel ezelőtt
- Látták:
Átírás

1 Fehérjék A fehérjék az élő szervezet számára nélkülözhetetlen molekulák, melyeket változatos térszerkezet és funkcionalitás jellemez. De úgy is fogalmazhatnánk, hogy a fehérjék olyan kivételes tulajdonságokkal rendelkező molekulák, amelyekből mesterséges körülmények között akár, egy általunk definiált feladat ellátására alkalmas molekuláris gépezetek is építhetők. E gépek felhasználás sokrétű lehet a gyógyászatban bevethető nanomedicináktól egészen az analitikában használható receptor struktúrákig, így számos terület számára jelenthetnek ideális megoldást. Egy fehérjemolekula több száz, esetenként több ezer, alegység (aminosav) rendezett halmaza, olyan láncmolekula (lineáris polimer), amelyet a szervezet a fehérjét kódoló gén által tartalmazott nukleotidok sorrendjét alapul véve képes felépíteni. Az élő szervezetek által termelt fehérjék 20 különféle aminosavat tartalmazhatnak. Peptidek mesterségesen is szintetizálhatóak aminosavakból (pl.: szilárd hordozó gyantán végzett acilezések sorozatával). Mitől különlegesek a fehérjék? Bár az ember is képes lineáris polimereket szintetizálni, azonban több lényegi különbség van a fehérjék és a mesterséges polimerek között. A peptidek aminosav sorrendje, s ezáltal méretük és tömegük szigorúan meghatározott. A mesterséges polimerek, oldatban nem rendelkeznek meghatározott térszerkezettel, s nagyszámú konformáció között véletlenszerűen csaponganak, addig a fehérjék polipeptidlánca képes felvenni egy jól definiált kompakt térszerkezetet. A fehérjék feltekeredettsége, kompakt térszerkezete teszi lehetővé, hogy változatos funkciókat lássanak el. A fehérjék szerkezeti szintjei Fehérjéknél megfigyelhető a különböző szerkezeti szintek hierarchiája. Az alábbi szerkezeti szinteket különböztethetjük meg: elsődleges szerkezet (kovalens szerkezet): aminosav szekvenciának megfelelő szerkezeti szint másodlagos szerkezet: polipeptid lánc rendezett, szabályos szerkezeti elemei, feltekeredettsége szuper-másodlagos szerkezet domén szerkezet harmadlagos szerkezet (térszerkezet): egyetlen polipeptid lánc térbeli szerkezete negyedleges szerkezet: vannak olyan fehérjék, amelyek több polipeptidláncból állnak. Az egyes alegységek relatív elhelyezkedése jelenti ezt a szerkezeti szintet. Elsődleges szerkezet Aminosavak A fehérjék alapvető építőelemei az aminosavak. Minden aminosav azonos vázból és egy ahhoz csatlakozó egyedi oldalláncból épül fel. Az aminosavak peptidkötéssel egymáshoz kapcsolódva alakítják ki a fehérjék polipeptidláncait, amelyek általában több száz 1
 azonos, hogy a központi szénatomhoz egy karboxil- és egy aminocsoport, (egy savas és egy bázikus csoport) valamint egy hidrogénatom kapcsolódik.")

2 aminosavból állnak. A polipeptidláncok váza periodikus szerkezet, amelyet a kapcsolódó oldalláncok tesznek változatossá. Az aminosavak alapfelépítésében (egy kivétellel) azonos, hogy a központi szénatomhoz egy karboxil- és egy aminocsoport, (egy savas és egy bázikus csoport) valamint egy hidrogénatom kapcsolódik. Az egyes aminosavak az R-rel jelölt oldalláncok minőségében különböznek egymástól. Vizes közegben az aminosavak karboxilcsoportja deprotonálodik negatívan töltötté válik, míg az aminocsoport protonálódik, pozitívan töltött lesz. Az aminosavak molekulában található központi szénatom királis (négyféle különböző funkciós csoport kapcsolódik hozzá), amely azt jelenti, hogy a poláros fény síkját a molekula elforgatja. Az elforgatás iránya kétféle lehet, nagysága mindkét esetben azonos. Így kétféle izomerről beszélhetünk, amelyek abban különböznek egymástól, hogy az egyik tengelyes tükörképe a másiknak. (L izomer és D izomer). Mesterségesen szintetizált aminosavak esetén racém elegy keletkezik mely nem forgatja el a poláros fény síkját, ugyanis a kétféle izomer aránya ötven-ötven százalék. Ezzel szemben az élőlények fehérjéiben kizárólag csak az L (balra forgató) izomer fordul elő. Bár a fehérjékben húszféle aminosav fordulhat elő, ez nem azt jelenti hogy csak húszféle létezik: az R oldallánc elvileg nagyon sok különböző csoport lehet. Az élő szervezetekben is legalább hatvanféle aminosav vagy aminosavszármazék fordul elő. A húszféle különböző genetikailag kódolt aminosavon kívül mesterségesen jóval több aminosav felhasználásával is szintetizálhatók fehérjék. Az aminosavakat általában 3 betűs, vagy 1 betűs rövidítéssel jelöljük. Az aminosavak oldalláncaik tulajdonságai szerint négyféle különböző csoportba sorolhatók. Nempoláros aminosavak: Glicin (Gly, G): A legegyszerűbb aminosav, itt csak egyetlen hidrogén van az oldallánc pozícióban. Ebből fakadóan a polipeptidlánc hajlékonysága a glicinnél a legnagyobb, hiszen itt az oldallánc méretéből adódóan nehezen alakul ki sztérikus gátlás, más oldalláncok vagy a főlánc atomjaival. 2
3 Alanin (Ala, A): Kicsit nagyobb, itt az R metil (CH3) csoportot jelöl. Valin (Val, V): még nagyobb, itt már el is ágazik az oldallánc. Prolin (Pro, P): eltér az alapszerkezettől, az oldallánc visszahajlik és kialakít egy kötést az aminocsoport nitrogénjével, gyűrűs struktúra keletkezik. Ez azt eredményezi, hogy megszűnik a főlánc C-N kötése körüli szabad forgás. Ahol a fehérjékben prolin van, ott a lánc merevvé válik, ezeknél a részeknél az α-helikális szerkezet kialakulása sem lehetséges. Cisztein (Cys, C): Különleges aminosav, azon kevesek egyike, mely kéntartalmú, funkciós csoportként tartalmaz egy SH csoportot. Ha a polipeptidláncban két cisztein oldallánc egymás közelébe kerül, akkor oxidatív környezetben egy vízkilépést követően kovalens S-S kénhíd (diszulfidhíd) alakul ki. Adott polipeptidláncon belül létrejövő kénhidakat intramolekulárisnak, míg két külön polipeptidszálat összekapcsoló kénhidakat intermolekulárisaknak nevezzük. Triptofán (Trp, W): Ez az aminosav rendelkezik a legnagyobb oldallánccal, kettős gyűrűje van és a fehérjékben általában nagyon kevés fordul elő. Szintetizálása költséges. Mivel a triptofán oldallánca viszonylag nagy kiterjedésű delokalizált rendszert tartalmaz, így közeli UV-ben 280 nm körül gerjeszthető, ezáltal alkalmas a fehérjeoldatok koncentrációjának spektrofotometriás meghatározására. Poláros aminosavak: Eredő töltéssel nem rendelkeznek, de jelentős dipólmomentumuk van. Fontos szerepet játszanak a hidrogénhíd képzésben, ahol donorként, ill. akceptorként képesek részt venni. Töltött aminosavak: Hisztidin (His, H): (gyakran a polárosok csoportjába sorolják), pk értéke kb. 7, vagyis nagyon könnyen vesz fel vagy ad le protont fiziológiás körülmények között. Enzimek aktív centrumaiban gyakori, ahol protonátviteli folyamatokban vesz részt. (a pk érték azt a ph értéket jelöli, ahol a protonált és deprotonált forma egyensúlyban van) Lizin (Lys, K) és Arginin (Arg, R): Az oldalláncok végükön könnyen protonálható 3
, Glutaminsav (Glu, E): Az oldalláncok karboxilcsoportot tartalmaznak, fiziológiás körülmények között negatívan töltöttek, deprotonáltak.")
4 aminocsoportot tartalmaznak. Fiziológiás körülmények között - ph 7 körül - pozitívan töltöttek. Aszparaginsav (Asp, D), Glutaminsav (Glu, E): Az oldalláncok karboxilcsoportot tartalmaznak, fiziológiás körülmények között negatívan töltöttek, deprotonáltak. Polipeptidlánc Ha két aminosav egymás közelébe kerül, akkor az egyik karboxilcsoportja (ami egy savas csoport) reakcióba lép a másik bázikus aminocsoportjával. Kilép egy vízmolekula és kialakul egy úgynevezett peptidkötés és egy dipeptid jön létre, ami annyiból hasonlít az aminosavakra (az aminosavat lehetne monopeptidnek is nevezni) hogy a két végén egy aminocsoportot ( N terminális) és egy karboxilcsoportot (C terminális) tartalmaz. A dipeptid mindkét végéhez újabb aminosavak kapcsolódhatnak, s a folyamat korlátlanul folytatható, mivel a láncvégeken mindig találhatóak amino- és karboxilcsoportok. A kialakuló lineáris polipeptidlánc valójában egy lineáris polimer. A polipeptidláncok váza (polipeptidváz) az N-C-C atomok monoton ismétlődésével kialakuló periodikus szerkezet, amit a hozzá kapcsolódó oldalláncok tesznek változatossá. A peptidkötés kialakulása: A peptidkötés kialakulásakor a kötésben résztvevő szomszédos négy atomra (C, O, N, H) kiterjedő delokalizáció jön létre (szénatom egy vegyértékelektronja válik delokalizálttá). A négy szomszédos atom egy adott síkban helyezkedik el, mivel rendszerük delokalizált, így a C-N kötés részleges kettőskötésként viselkedik, körülötte rotáció nem megengedett, azaz a peptidcsoport egy merev egységként viselkedik. A peptidkötés konfigurációját tekintve transz, a két R oldallánc, a polipeptidlánc ellentétes oldalán helyezkednek el. A peptidcsoporton belül a töltések lokálisan elválnak, az egyes atomokon parciális (részleges) töltések jelennek meg. Éppen ezért a peptidcsoport viszonylag nagy elektromos dipólmomentummal rendelkezik, erősen poláros. A peptidcsoport gyakran vesz részt H-hidakban: az NH csoport H-híd donorként, míg a CO csoport akceptorként viselkedik. 4
5 A polipeptidlánc flexibilitása: A polipeptidlánc, egy hajlékony, flexibilis szerkezet. A lánc elforgatható minden C α -hoz (központi szénatom) csatlakozó két egyes kötés körül, s ezenkívül általában még az oldalláncok is tartalmaznak szabad forgást megengedő egyeskötéseket. Mindez azt eredményezi, hogy a lánc egy jól definiált kompakt térszerkezetnek megfelelően feltekerhető, viszont a fehérjék tervezésénél e tulajdonság okozza a nehézséget, hiszen a lánc flexibilátásából adódóan igen sokféleképpen feltekerhető. Így a molekula legstabilisabb alakjának tervezése igen számításigényes feladat, ami ma még távolról sem megoldott. Pl.: hányféleképpen lehet feltekerni egy polipeptidláncot? egy átlagos fehérje mérete kb. 300 aminosav (de kb. 70 és között mozog a számuk) a fehérjék polipeptidláncának (aminosavak) sorrendje szigorúan meghatározott: aminosavszekvencia most nem vesszük figyelembe, hogy az oldallácokat is lehet tekergetni, csak a fővázat tekintjük noha a peptidváz szabadon aminosavanként két kötés körül szabadon tekergethető, az oldalláncok egymással ill. a polipeptidvázzal való ütközése miatt számos szöghelyzet mégsem jöhet létre tegyük fel, hogy aminosavanként mindkét kötés körül ahol szabad forgás van, átlagosan csak két lehetséges konformáció van (erősen alulbecsülve a lehetséges kombinációk számát), vagyis aminosavanként négy lehetséges konformációval számolva a polipeptidlánc összes konformációinak száma: =4,15*10 180, ami egy gigantikus szám. Aminosavszekvencia A fehérjék elsődleges szerkezete alatt polipeptidláncuk aminosavsorrendjét (aminosavszekvenciáját) értjük. Manapság 3-4 millió fehérjének ismerjük az aminosavsorrendjét, az ember genomjában pl. kb. harmincezer fehérje van kódolva. Ezek megtalálhatók az Interneten adatbázisokban, ezeket vizsgálva nyomon lehet követni az evolúciót, látszik hogy az összes élőlény egy közös eredetre vezethető vissza, mindegyikben ugyanazok az aminosavak vannak, ill. van néhány olyan fehérje, amelynek variánsai minden élőlényben megtalálhatók. Egy adott fehérje aminosavszekvenciája a rokon élőlényekben nagyrészt megegyezik, így több élőlényt vizsgálva megkapjuk, hogy egy-egy tulajdonságért a szekvencia melyik régiója felelős. Másodlagos szerkezet A másodlagos szerkezet a polipeptidváz lokálisan rendezett szerkezeti elemeit foglalja magában. A polipeptidváz kémiailag periodikus szerkezetű, éppen ezért Amikor egy polipeptidlánc feltekeredik, az oldalláncokat nem tekintve egy periodikus térbeli szerkezet a kedvezményezett. Ilyen szabályos szerkezet például az α-hélix és a β-redő. Az adott szerkezet kialakulásának jóslására használhatjuk a Ramachardan diagrammot. Ramachardan indiai származású tudós volt, aki a központi atomhoz tartozó két egyszeres kovalens kötés körüli rotáció térszerkezeti következményeit vizsgálta. 5

6 A polipeptidlánc csak úgy tekergethető, ha az atomok nem kerülnek egymással átfedésbe, vagyis nem léphet fel sztérikus gátlás. Ramachandran azt vizsgálta, hogy ha az R oldallánc helyére egy metilcsoportot teszünk, akkor a polipeptidváz N-C α és C α -C egyes kötései körül mely szögpárértékek lesznek megengedettek. Az egyes atomokat a van der Waals sugaruknak megfelelő merev gömbökkel helyettesítette. A megengedett szögtartományokat kétdimenzióban ábrázolva kapjuk az ún. Ramachandran térképet: 6


7 A diagramon látható, hogy alapvetően három megengedett tartomány van. A bal felső sarokban található β-tartomány a polipeptidlánc nyújtott konformációit foglalja magában. Az α-tartományban lévő megengedett konformációk között találjuk az ún. α-helikális szerkezeteket, míg az L-tartomány a hajlatrégióknak felel meg. A már ismert szerkezetű fehérjékben aminosavanként az aktuális szögpárértékeket meghatározva és ábrázolva azt láthatjuk, hogy a megfigyelt értékek nagyon jó összhangban vannak a Ramachandran diagram előrejelzésével. Esetenként egy-egy pont a megengedett tartományokon kívülre esik, aminek az az oka, hogy az atomok nem merev gömbök, távoli láncrészek kedvező kölcsönhatásai miatt lokálisan az oldallánc és főlánc atomok elektronfelhői kismértékben egymásba nyomhatók. Ramachandran a számításait a CH 3 oldalcsoportot tartalmazó alanin aminosavra végezte, Nyilván a valóságban a megengedett régiók annál szűkebbek, minél nagyobb kiterjedésű az R oldallánc. Ha a megengedett régiókban egy kiválasztott szögpárértéket vesz fel több egymás utáni aminosav, akkor periodikus térbeli szerkezethez jutunk. Ezek között van néhány, amit a főváz atomjai között kialakuló kedvező kölcsönhatás-mintázatok stabilizálnak, s ezért nagyon gyakran figyelhetők meg a fehérjékben. Ilyen pl. az α-hélix, a β-szál ill. a β-hajlat. a-hélix: Az α-hélix a Ramachandran térkép α-tartományában található szögpárértékeknek megfelelő periodikus szerkezet. A fehérjékben nagyon gyakran előforduló szerkezeti elem. Az α-hélixben az aminosavak C α atomjainak a távolsága 1,5 Ǻ. Egy teljes körülfordulás 3,6 aminosavanként következik be. A hélix átmérője 6 Ǻ, menetemelkedése 5,4 Ǻ. Az α -helikális szerkezetet az teszi igen stabillá, hogy a struktúrában a főlánc atomjai úgy helyezkednek el, hogy köztük egy igen kedvező hidrogénhidas szerkezet képes kialakulni. A hidrogén hidak a hélix tengelyével párhuzamosan helyezkednek el. Egy kiszemelt peptidkötés C=O csoportja a rákövetkező negyedik peptidkötés N-H csoportjával alakít ki H- 7
 és egy akceptorra (olyan atom, mely magános elektron párt tartalmaz) van szükség.")

8 hidat. Az α-hélix keresztmetszeti képén láthatjuk, hogy az oldalláncok a hélix tengelyére merőlegesen kifelé helyezkednek el, s így nem zavarják a főlánc atomok egymással való kölcsönhatását. Megjegyzés: a hidrogén hidak kialakulásához egy donorra (olyan atom, melyhez hidrogént kapcsolódik) és egy akceptorra (olyan atom, mely magános elektron párt tartalmaz) van szükség. A hidrogénhíd kialakulásához donor és az akceptor 1,5 2 Ǻ távolsága szükséges. A hidrogénkötés annál erősebb, minél jobban megvalósul az, hogy a kötést alkotó atomok egy egyenesben helyezkednek el. Béta-szerkezet: Ha a láncszögek a 180 -hoz közel - a Ramachandran térkép béta tartományában - vannak, akkor erősen nyújtott lánckonformációk jönnek létre. Valójában a β-szálas szerkezet egy erősen nyújtott helikális szerkezetként is felfogható, amelyben egy körülfordulásra 2 aminosav esik, a szomszédos C α atomok távolsága Å. Amíg az α-hélix esetében a polipeptidlánc közeli aminosavegységei képeznek egymással hidrogénhidakat, addig egyetlen β-szálon belül nem tudnak H-hidak kialakulni a főváz atomjai között. Azonban ha több ilyen láncot párhuzamosan egymás mellé helyezünk, akkor egy láncok közötti H-hidak által stabilizált lemezes szerkezet jöhet létre. A tapasztalat azt mutatja, hogy attól függően, hogy a szomszédos β-láncok egymással megegyező vagy ellentétes irányban futnak kétféle energetikailag kitüntetett szerkezet alakulhat ki: az ún. parallel illetve anti-parallel béta-lemezek. A fehérjékben mindkettő gyakran megfigyelhető. Az ábrán látható antiparallel β-lemezek energetikailag stabilabbak, mert itt a láncokra merőlegesen kialakuló H-hidakat alkotó atomok egy egyenesbe esnek. Ha láncok parallel helyezkednek el, akkor is kialakulhatnak a láncok közötti hidrogén hidak, amelyek azonban eléggé torzítottak és ezért az előbbieknél gyengébbek. Mindkét β-lemezes szerkezetben az oldalcsoportok (R) a lemez síkja alatt és felett, míg a hidrogén hidak a lemez síkjában helyezkednek el. Ha a β-lemez egyik oldalán csupa apoláros oldalláncok találhatók, akkor két ilyen β-lemezt egymással szembe fordítva azok a hidrofób kölcsönhatás 8
9 révén könnyen összetapadnak, s ún. β-szendvics szerkezetet alkotnak. Ugyanabban a β-lemezes szerkezetben egymástól szekvenciálisan nagyon távoli láncszakaszok is részt vehetnek. Ahhoz, hogy a fehérjék globuláris szerkezete kialakulhasson, a nyújtott láncszakaszok a fehérjék felszínét elérve megtörniük és gyakran akár 180 fokos szögben visszafordulnak. Ennek tipikus szerkezeti motívuma az ún. β-hajlat. β-hajlat: A β-hajlat a Ramachandran térkép L-tartományának megfelelő polipeptidváz konformáció. Három fő típusát különböztethetjük meg, amelyekben az a közös, hogy a polipeptidlánc négy aminosavegységen belül közel 180 fokban visszafordul. Ezeket a szerkezeteket is az tünteti ki energetikailag, hogy a főváz atomjai között kialakuló H-hidak stabilizálják őket. Általában a β-hajlat egyik aminosava glicin, ami oldalláncának kis méreténél fogva biztosítja a polipeptidlánc megfelelő flexibilitását. 9



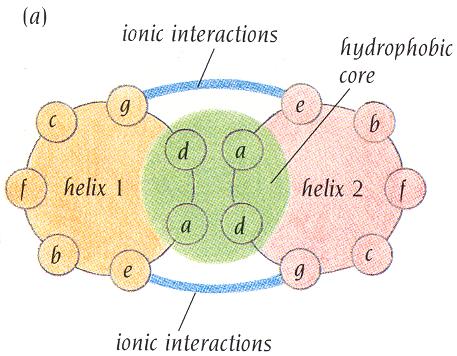
10 Szupermásodlagos szerkezet Az ismert fehérjeszerkezetekben megfigyelhető, hogy a másodlagos szerkezeti elemek bizonyos kombinációi különösen gyakran fordulnak elő. Ezeket a gyakori szerkezeti motívumokat szupermásodlagos szerkezeti elemeknek nevezzük. pl.: antiparallel béta-szálak, β α β motívum, béta-hordó,... Ilyen motívumokból félét ismerünk. A szupermásodlagos szerkezeti elemek közé tartoznak a helikális kötegek is. Ezekben 2, 3 vagy 4 α-hélix fut közel párhuzamosan, enyhén egymás köré fonódva. A hélixek egymás felé forduló oldalain apoláros oldalláncokat találunk amelyek közötti hidrofób kölcsönhatás stabilizálja a helikális kötegeket. A hélixek kölcsönhatásának eredményeként egy ún. hidrofób mag jön létre. A hélixek kifelé néző oldalán ugyanakkor általában poláros oldalláncok helyezkednek el (amfipatikus hélixek: egyik oldaluk poláros, a másik apoláros). Mivel az α-hélix két teljes körülfordulására éppen 7 aminosav esik, ezért ezekben a helikális kötegeket formáló hélixekben az apoláros aminosavak hetes ismétlődése figyelhető meg, miáltal az aminosavszekvenciából is általában könnyen felismerhetők. (Minden 7. pozíció mellett a közbenső 4. pozicióban is általában hidrofób aminosavat találunk.) A helikális kötegek kialakulása nemcsak egy fehérjén belül mehet végbe, de viszonylag gyakran megfigyelhető a fehérjék közötti kölcsönhatások esetében is, amikor a kölcsönható partnerek 1-2 helikális szegmenssel vesz részt egy közös, a két fehérjét összetartó helikális köteg létrehozásában. 10

11 11

12 Doménszerkezet A tapasztalatok azt mutatják, hogy a kb. 150 aminosavnál nagyobb fehérjék szerkezete több, egymástól többé-kevésbé független szerkezeti egységre, ún. doménekre bontható. A doméneket háromféleképpen is osztályokba sorolhatjuk, s megkülönböztethetünk: szerkezeti doméneket folding doméneket funkcionális doméneket. Egy adott domén akár mindhárom kritériumnak is megfelelhet. Szerkezeti domén: amikor egy feltekeredett polipeptidlánc (fehérjemolekula) több, kvázi független szerkezeti egységre bontható, akkor ezen egységeket szerkezeti doméneknek nevezzük. A szerkezeti domének tipikus mérete aminosav. Ha elvágjuk a doméneket összekötő pőlipeptidlánc-szakaszt (pl. egy alkalmasan választott proteolitikus enzimmel), akkor a szerkezeti domének megtartják natív térszerkezetüket. Folytonos szerkezeti doménről beszélünk akkor, ha a polipeptidlánc egy folytonos szakasza alakít ki egy domént. Ha a polipeptidlánc több szakasza vesz részt egy domén kialakításában, akkor nemfolytonos doménről beszélünk. Folding (térszerkezet kialakulás, feltekeredés) domén: a térszerkezet kialakulás szempontjából képez különálló egységet, azaz ha elrontjuk (denaturáljuk) egy folding domén szerkezetét (pl. hővel), és utána újra megfelelő körülményeket teremtünk a számára, akkor ismét képes feltekeredni, s kialakítja a struktúráját. Egy fehérje hőindukálta kitekeredési folyamatát pásztázó mikrokalorimetria segítségével tanulmányozva viszonylag egyszerűen meghatározhatjuk, hogy egy fehérje hány folding doménből áll. A legtöbb esetben hacsak nincsenek erős kölcsönhatások a domének között egy folytonos szerkezeti domén egyben folding doménként is funkcionál. Egy polipeptidlánc lehetséges konformációinak száma exponenciálisan nő az alkotó aminosavak számával. Egy nagyobb fehérje esetén a natív szerkezet megtalálása lényegesen lerövidíthető, ha a polipeptidlánc nem egy egységékét tekeredik fel, hanem olyan kisebb részekre bontható, amelyek önállóan képesek megtalálni natív térszerkezetüket. Funkcionális domén: Nagyon sok esetben az egyes doménekhez külön funkciók köthetők, pl.: az egyik domén köt valamilyen koenzimet, pl. az ATP-t (ATP-kötő domén), másik meg egy adott katalitikus funkciót végez (pl. proteáz domén). Vannak olyan fehérjecsaládok, pl. a véralvadás fehérjéi, melyek a doméneket, mint építőelemeket használják. Ez egy nagy evolúciós vívmány, hiszen egy új fehérje felépítésénél nem a nulláról kell elindulni, hanem már meglévő működő elemekből lehet új fehérjét létrehozni. 12
13 A véralvadásban részt vevő fehérjék doménszerkezete. 13

14 Harmadlagos szerkezet (térszerkezet) A harmadlagos szerkezet egyetlen polipeptidlánc térbeli szerkezete. Ma már közel 30 ezer fehérje térszerkezetét ismerjük atomi precizitással. A rendelkezésre álló adatok alapján a vízoldékony fehérjék térszerkezetének alapvető jellemzői: tömör térkitöltés poláros felszín, hidrofób mag H-híd képzési lehetőségek maximálisan kihasználtak az egyes aminosavkonformációk alapvetően a megengedett Ramachandran régióknak megfelelőek A fehérjék térszerkezete nagyon szoros térkitöltésű, jól definiált szerkezet. Ez a szerkezet általában tömörebb, mint egy átlagos molekulakristály szerkezete. A fehérje belsejében nincsenek üregek, vagy ha vannak, azt víz tölti ki, s ennek a víznek is valamilyen funkcionális szerepe van. Azért ilyen tömör szerkezetű a fehérje, hogy az aminosavakat alkotó atomok, oldalláncok közötti kölcsönhatások maximálisan kihasználásra kerüljenek. A (vízoldékony) fehérjék belsejében helyezkednek el az egymással hidrofób és van der Waals kölcsönhatásban álló apoláros oldalláncok, a külső részen pedig a vízzel kedvező kölcsönhatások kialakítására képes töltött és poláros oldalláncok. Minthogy az aminosavak közötti peptidkötés poláros csoportokat tartalmaz, ezért a polipeptidlánc feltekeredése során elkerülhetetlen, hogy a poláros csoportok egy része a fehérje belsejébe kerüljön. Ezek a peptidcsoportok azonban szinte kivétel nélkül H-hidakat képeznek egymással, s ezáltal polaritásuk leárnyékolódik. Nagyon ritka, hogy poláros rész szabadon fordulna elő a fehérje belsejében. Néha a felszínen is előfordulhatnak apoláros régiók, de ennek is oka van, például másik fehérjével való kölcsönhatás miatt fontosak. A mioglobin oxigénkötő fehérje harmadlagos szerkezete:

15 Fontos megjegyezni, hogy az apoláros lipidkörnyezetben elhelyezkedő membránfehérjék esetében mások a szerkezeti követelmények. Itt általában az apoláros lipidkörnyezettel érintkező felületi régiókbal találjuk a hidrofób oldalláncokat, míg a poláros és töltött csoportok a fehérje belsejében helyezkednek el. FEHÉRJESZERKEZET MEGHATÁROZÁS A fehérjék a legbonyolultabb molekulák, amiket ismerünk. Ezért is csábító kihívás térszerkezetük meghatározása. Pontosabban ilyen volt egykoron, mára szinte rutinfeladattá vált. Manapság több mint harmincezer fehérje szerkezetét ismerjük atomi precizitással, de ezek között sok rokon fehérje is található, amelyek különböző élőlényekben látnak el azonos funkciót. Összehasonlításképpen talán elég arra gondolnunk, hogy egy emberi sejtben kb. húszezer különféle fehérje működik. Az összes felderített szerkezet a brookhaven-i fehérje adatbankban megtalálható. Az interneten keresztül ( ) bárki kedvére gyönyörködhet bennük. A fehérjék szerkezetétnek meghatározása alapvetően háromféle kísérleti módszerrel lehetséges, ezek a röntgendiffrakció, a magmágneses rezonancia (NMR) spektroszkópia és az elektronmikroszkópia. Röntgendiffrakció Az első fehérjék térszerkezetét az 1960-as években határozták meg. Kendrew és Perutz röntgendiffrakciós vizsgálatai 7 évnyi erőfeszítés eredményeként vezettek el a hemoglobin atomi szerkezetének leírásához. Egyetlen fehérjemolekula elektronfelhője csak parányi mértékben téríti el a röntgensugarakat. Ahhoz, hogy mérhető röntgenszórást tapasztaljunk, sok ezer milliárdnyi fehérjemolekulából rendezett mintát kell előállítanunk. Általában ez azt jelenti, hogy a fehérjét kristályosítani kell. A fehérjék kristályosítása egyáltalán nem triviális feladat, hiszen miért is kellene a bonyolult alakú fehérjemolekuláknak képesnek lenniük a kristályképződésre. A tapasztalat mégis azt mutatja, hogy megfelelő körülmények között a legtöbb fehérje kristályosítható. A megfelelő körülmények megtalálása azonban esetenként akár több évnyi erőfeszítést is igényelhet. 2. ábra A mioglobin fehérje kristályának röntgenszórási képe.

16 Korábban egy röntgendiffrakciós szerkezetanalízishez a milliméteres mérettartományt megközelítő kristályok előállítására volt szükség, amit aztán akár több hónapnyi adatgyűjtés majd szerkezetszámítás és finomítás követett. Ma már a sok nagyságrenddel intenzívebb szinkrotronos sugárforrásokat használva mikrométeres kristályok is vizsgálhatók, s kedvező esetben néhány nanoszekundumnyi idő alatt elegendő adat regisztrálható. Egy fehérjekristályról készült röntgenszórási kép valójában szabályosan elhelyezkedő sötétebb és világosabb foltok összessége (2. ábra), ahol a foltok intenzitása hordozza a fehérjeszerkezetre (pontosabban elektronsűrűségre) vonatkozó információt. Felmerülhet a kérdés, hogy egy kristályba rendeződött fehérjemolekula szerkezete vajon ugyanolyan-e, mint oldott állapotban. Számos megfigyelés utal arra, hogy igen. Egy tipikus fehérjekristály térfogatának ugyanis közel 70%-a víz. A fehérje molekulák között viszonylag kicsik a kontaktusok, az intermolekuláris térben tágas vízzel telt csatornák találhatók. A legtöbb enzimfehérje kristályos állapotban megtartja katalitikus tulajdonságait: a szubsztrát molekulák a vízzel telt csatornákon keresztül képesek a kristály belsejében lévő fehérjékhez diffundálni, majd azokhoz kötődve lejátszódik a fehérje által katalizált folyamat. NM R spektroszkópia A röntgendiffrakciós szerkezetanalízis lépései. Az NMR spektroszkópiás szerkezetvizsgálat szerencsésen kiegészíti a röntgendiffrakciós analízist, mert nem igényel rendezett mintát, s természetes, vizes környezetükben képes tanulmányozni a fehérjéket. Hátránya viszont, hogy ezzel a módszerrel nagyobb méretű - jelenleg 300 aminosvnál hosszabb - fehérjék nem tanulmányozhatók. Az NMR spektroszkópiás módszerrel meghatározott szerkezetek gyakorlatilag megegyeznek a röntgendiffrakciós analízis eredményével, bizonyítva azt, hogy kristályos állapotban a fehérjék nem szenvednek el számottevő torzulást. A páratlan nukleonszámú atommagoknak saját belső mágneses momentuma (spinje) van, ami egy adott külső mágneses térben, azzal megegyező vagy ellentétes irányban helyezkedhet el. A két állapot között energiakülönbség van. Rádiófrekvenciás jelekkel át lehet billenteni a spineket egyik állapotból a másikba. Az, hogy egy kiszemelt pl. proton mekkora energiával billenthető át nemcsak a külső mágneses tértől nagyságától függ, hanem a környező molekula szerkezettől is, ugyanis a közeli atommagok és elektornok befolyásolják a lokális mágneses teret. Ezt használja ki az NMR. Különböző rádiófrekvenciás jelsorozatokkal megpróbálják átbillenteni a protonokat. Abból, hogy az egyes protonok milyen frekvencián billenthetők át következtetni lehet lokális környezetükre, s ezáltal a térszerkezetre.

17 Az NMR szerkezetanalízis lépései Az NMR esetében nem kell kristályokat használni, a fehérjék természetes vizes környezetükben tanulmányozhatók, viszont a mérés és az analízis exponenciálisan bonyolódik a fehérje nagyságával. Elektronmikroszkópia A fenti módszerek alapvetően vízoldékony fehérjék szerkezetének meghatározására alkalmasak. Ugyanakkor a membránfehérjék a sejtek életének számos létfontosságú folyamatában vesznek részt (pl. fotoszintézis, jelátvitel, anyagtranszport). A membránfehérjéket természetes lipidkörnyezetükből kiszakítva elvesztik natív térszerkezetüket, általában nem kristályosíthatók, vizes közegben nem tanulmányozhatók. Jelenleg az ismert fehérjeszerkezetek kevesebb, mint 1%-át teszik ki a membránfehérjék. Bár akadt már példa a lipidkörnyezetet szimuláló detergensek jelenlétében (azokkal együtt) a membránfehérjék kristályosítására és röntgendiffrakciós szerkezetvizsgálatára, a membránfehérjéket egyszerűbbnek tűnik a membránon belül rendezni, ún. kétdimenziós kristályokat létrehozva belőlük. Az ilyen rendezett vékonyrétegek szerkezetvizsgálatára kínál megoldást az elektronmikroszkópia (EM), amelynek segítségével szerencsés esetben ugyancsak atomi precizitású szerkezeti adatokat nyerhetünk. Az elektronmikroszkópia lehetőséget nyújt nagyméretű szupramolekuláris fehérjekomplexumok (negyedleges fehérjeszerkezetek) vizsgálatára is az un. egyrészecskés szerkezetanalízis révén, amikor a komplexumról készült több száz vagy több ezer EM felvételt számítógéppel kiátlagolva végezzük el a jel/zaj viszony növelését, s jutunk el az atomi szerkezet meghatározását lehetővé tévő adatokhoz.
18 A FEHÉRJÉK DINAMIKÁJA A bemutatott fehérjeszerkezetek azt a képzetet kelthetik, mintha a fehérjék statikus objektumok lennének, az őket felépítő atomok szigorúan meghatározott pozíciókban helyezkednének el. Természetesen ez nincsen így, a fehérjék örökös mozgásban, nyüzsgésben vannak, egyes részeik különböző időskálán lejátszódó mozgásokban vesznek részt. Ezek a mozgások rendkívül széles időskálát ölelnek fel, a pikoszekundumos (10-12 s) tartománytól akár az éves (10 6 s) időtartamokig. Ezen mozgások az atomi rezgésektől és az oldalláncok gyors rotációs mozgásától, a működés során gyakran megfigyelhető relatív doménmozgásokon keresztül, a neurodegeneratív betegségek (prionbetegségek) hátterében álló lassú konformációs átrendeződésekig terjednek. Molekuláris dinamikai szimulációk A fehérje egyes részeinek kollektív, irányított mozgásai gyakran meghatározó szerepet játszanak a fehérjék működésében. A fehérjék szerkezetét fenntartó erők ismeretében a rendelkezésre álló atomi szerkezet alapján a molekuláris mozgások akár számítógéppel is szimulálhatók. Sajnos ezek a szimulációk az atomok (szabadsági fokok) rendkívül nagy száma miatt még a leggyorsabb szuperszámítógépekkel is csak néhányszor 10 ns-os időtartamra végezhetők el. A kapott eredmények alapján sokszor lehetetlen következtetni pl. az enzimműködés szempontjából meghatározó milliszekundumos időskálán lejátszódó folyamatokra. Időfelbontásos röntgendiffrakció Meglepő módon a fehérjékben lejátszódó dinamikus folyamatok tanulmányozásában éppen a röntgendiffrakciós szerkezetvizsgálat siet segítségünkre. Amint azt már említettük, a rendkívül intenzív szinkrotronos sugárforrásoknál az adatgyűjtés ma már akár néhány nanoszekundum alatt elvégezhető. Ahhoz, hogy egy kristályban lévő nagyszámú fehérjemolekula funkcionálisan releváns mozgásai ne átlagolják ki egymást, ezeket a mozgásokat valahogyan szinkronizálni kell. Egy kristályban elhelyezkedő enzim általában megőrzi katalitikus képességét, a kristályt átszövő vízzel telt csatornákon keresztül a bediffundáló szubsztrát molekulákat képes megkötni, majd átalakítani. Esetenként előállíthatók olyan un. fotoaktiválható szubsztrátanalógok, amelyek szerkezetükben hasonlítanak a természetes szubsztrátra, ezért az enzim képes megkötni őket, de a szerkezeti eltérések miatt mégsem játszódik le a katalitikus folyamat. A felesleges oldalcsoportot (vagy kémiai kötést) egy gyors és erős lézerimpulzussal lehasítva az összes enzimmolekula esetén egyszerre indíthatjuk el a katalitikus reakciót. A reakció elindítása után meghatározott időtartamonként (pl. milliszekundumonként) röntgen pillanatfelvételeket készítve jellemezhetjük a katalízis szempontjából meghatározó intramolekuláris mozgásokat. Éppen a fehérjék dinamikai vizsgálata mutatta meg, hogy ezek a belső mozgások döntően meghatározzák az enzimatikus folyamat hatékonyságát. A fehérjék azért sokkal hatékonyabbak pl. a szervetlen katalizátoroknál, mert a működésük során dinamikus tulajdonságaik miatt szerkezetük folyamatosan idomulni képes a változó követelményekhez. A szubsztrát bekötődése olyan konformációs átrendeződést idéz elő, amely lehetővé teszi az (az energiagátat lecsökkentve) az átmeneti termék erős kötését is, majd a kémiai reakció megtörténte után bekövetkező relaxációs folyamatok eredményeként képes könnyen elereszteni a végterméket, lehetővé téve az újabb katalitikus ciklus megkezdését.
.")
19 Negyedleges szerkezet A több polipeptidláncból (alegységből) felépülő fehérjék és fehérjekomplexumok esetében beszélhetünk negyedleges szerkezetről, ami alatt az egyes alegységek relatív elhelyezkedése révén létrejövő térbeli struktúrát értjük. Ilyen negyedleges szerkezetű fehérje pl. a hemoglobin (oxigén transzportban játszik szerepet). A hemoglobin négy alegységből áll, amelyekből kettő-kettő megegyező szerkezetű. Az alegységek nemkovalens kölcsönhatása révén alakul ki a negyedleges szerkezet. Hasonlóképpen egy vírus több különböző fehérjealegységből felépülő fehérjeburka negyedleges szerkezetnek minősül. Ha egy fehérjekomplexum több azonos alegységet tartalmaz, akkor azok általában szimmetrikusan helyezkednek el. Erre számos példát találunk az élő szervezetekben (4. ábra). Érdemes elgondolkodni azon, hogy miért előnyös az, ha több alegységből épül fel egy működőképes rendszer, ahelyett, hogy egyetlen óriási polipeptidláncból állna. A fehérjék (ill. génjeik) szintézise elkerülhetetlenül együtt jár hibákkal. Egy több alegységből felépülő fehérjekomplexum létrehozása esetén ha hiba történik, akkor nem kell az egészet kidobni, hanem csak azt a részt, ami nem jó. Másik előnye a moduláris felépítésnek, hogy az

20 4. ábra A fehérjék negyedleges szerkezetében megfigyelhető szimmetriák. alegységek között kölcsönhatás befolyásolásán keresztül számos szabályozási lehetőséget kínál. Lehetőséget nyújt az alegységek közötti közvetlen szubsztrátátadásra is. Kiterjedt térbeli szerkezeteket (pl. flagelláris filamentum) is sokkal könnyebb azonos alegységekből építeni, s az építőelemek koncentrációján keresztül szabályozható a szerkezet kialakulása. Valójában a fehérjékből felépülő molekuláris gépezetek önszerveződő képessége nem más, mint azok alegységeikből való spontán kialakulása.
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Hemoglobin - myoglobin. Konzultációs e-tananyag Szikla Károly
 Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
A fehérjék szerkezeti hierarchiája. Fehérje-szerkezetek! Klasszikus szerkezet-funkció paradigma. szekvencia. funkció. szerkezet! Myoglobin.
 Myoglobin Fehérje-szerkezetek! MGLSDGEWQLVLNVWGKVEADIPGGQEVLIRLFK GPETLEKFDKFKLKSEDEMKASE DLKKGATVLTALGGILKKKGEAEIKPLAQSA TKKIPVKYLEFISECIIQVLQSK PGDFGADAQGAMNKALELFRKDMASNYKELGFQG Fuxreiter Mónika! Debreceni
Myoglobin Fehérje-szerkezetek! MGLSDGEWQLVLNVWGKVEADIPGGQEVLIRLFK GPETLEKFDKFKLKSEDEMKASE DLKKGATVLTALGGILKKKGEAEIKPLAQSA TKKIPVKYLEFISECIIQVLQSK PGDFGADAQGAMNKALELFRKDMASNYKELGFQG Fuxreiter Mónika! Debreceni
Fehérjeszerkezet, és tekeredés
 Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek
 Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
FEHÉRJÉK A MÁGNESEKBEN. Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium. Alkímia Ma, Budapest,
 FEHÉRJÉK A MÁGNESEKBEN Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium Alkímia Ma, Budapest, 2013.02.28. I. FEHÉRJÉK: L-α aminosavakból felépülő lineáris polimerek α H 2 N CH COOH amino
FEHÉRJÉK A MÁGNESEKBEN Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium Alkímia Ma, Budapest, 2013.02.28. I. FEHÉRJÉK: L-α aminosavakból felépülő lineáris polimerek α H 2 N CH COOH amino
A fehérjék szerkezete és az azt meghatározó kölcsönhatások
 A fehérjék szerkezete és az azt meghatározó kölcsönhatások 1. A fehérjék szerepe az élõlényekben 2. A fehérjék szerkezetének szintjei 3. A fehérjék konformációs stabilitásáért felelõs kölcsönhatások 4.
A fehérjék szerkezete és az azt meghatározó kölcsönhatások 1. A fehérjék szerepe az élõlényekben 2. A fehérjék szerkezetének szintjei 3. A fehérjék konformációs stabilitásáért felelõs kölcsönhatások 4.
A sejtek élete. 5. Robotoló törpék és óriások Az aminosavak és fehérjék R C NH 2. C COOH 5.1. A fehérjeépítőaminosavak általános
 A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
Az anyagi rendszer fogalma, csoportosítása
 Az anyagi rendszer fogalma, csoportosítása A bemutatót összeállította: Fogarasi József, Petrik Lajos SZKI, 2011 1 1 A rendszer fogalma A körülöttünk levő anyagi világot atomok, ionok, molekulák építik
Az anyagi rendszer fogalma, csoportosítása A bemutatót összeállította: Fogarasi József, Petrik Lajos SZKI, 2011 1 1 A rendszer fogalma A körülöttünk levő anyagi világot atomok, ionok, molekulák építik
Szerkesztette: Vizkievicz András
 Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
A fehérjék hierarchikus szerkezete
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Az élő anyag szerkezeti egységei: víz, nukleinsavak, fehérjék. elrendeződés, rend, rendszer, periodikus ismétlődés
 Az élő anyag szerkezeti egységei: víz, nukleinsavak, fehérjék Agócs Gergely 2013. december 3. kedd 10:00 11:40 1. Mit értünk élő anyag alatt? Az élő szervezetet felépítő anyagok. Az anyag azonban nem csupán
Az élő anyag szerkezeti egységei: víz, nukleinsavak, fehérjék Agócs Gergely 2013. december 3. kedd 10:00 11:40 1. Mit értünk élő anyag alatt? Az élő szervezetet felépítő anyagok. Az anyag azonban nem csupán
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Kémiai kötések. Kémiai kötések. A bemutatót összeállította: Fogarasi József, Petrik Lajos SZKI, 2011
 Kémiai kötések A bemutatót összeállította: Fogarasi József, Petrik Lajos SZKI, 2011 1 Cl + Na Az ionos kötés 1. Cl + - + Na Klór: 1s 2 2s 2 2p 6 3s 2 3p 5 Kloridion: 1s2 2s2 2p6 3s2 3p6 Nátrium: 1s 2 2s
Kémiai kötések A bemutatót összeállította: Fogarasi József, Petrik Lajos SZKI, 2011 1 Cl + Na Az ionos kötés 1. Cl + - + Na Klór: 1s 2 2s 2 2p 6 3s 2 3p 5 Kloridion: 1s2 2s2 2p6 3s2 3p6 Nátrium: 1s 2 2s
A kovalens kötés polaritása
 Általános és szervetlen kémia 4. hét Kovalens kötés A kovalens kötés kialakulásakor szabad atomokból molekulák jönnek létre. A molekulák létrejötte mindig energia csökkenéssel jár. A kovalens kötés polaritása
Általános és szervetlen kémia 4. hét Kovalens kötés A kovalens kötés kialakulásakor szabad atomokból molekulák jönnek létre. A molekulák létrejötte mindig energia csökkenéssel jár. A kovalens kötés polaritása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
A négymilliárd éves nanotechnológia
 A négymilliárd éves nanotechnológia Tisztelt Hölgyeim és Uraim! Ha az önök kezében lenne egy öklömnyi szén, az enyémben pedig egy ugyanakkora gyémánt, cserélnének e velem? Kérdezhetném miért, hiszen mindkettő
A négymilliárd éves nanotechnológia Tisztelt Hölgyeim és Uraim! Ha az önök kezében lenne egy öklömnyi szén, az enyémben pedig egy ugyanakkora gyémánt, cserélnének e velem? Kérdezhetném miért, hiszen mindkettő
Kémiai kötések. Kémiai kötések kj / mol 0,8 40 kj / mol
 Kémiai kötések A természetben az anyagokat felépítő atomok nem önmagukban, hanem gyakran egymáshoz kapcsolódva léteznek. Ezeket a kötéseket összefoglaló néven kémiai kötéseknek nevezzük. Kémiai kötések
Kémiai kötések A természetben az anyagokat felépítő atomok nem önmagukban, hanem gyakran egymáshoz kapcsolódva léteznek. Ezeket a kötéseket összefoglaló néven kémiai kötéseknek nevezzük. Kémiai kötések
Peptidek és fehérjék 1. Fehérjék Fehérjetekeredés. Fehérje (protein) Fehérje (protein) Aminosavak. Aminosavak
 Fehérje (protein) Aminosavak. Aminosavak") Fehérjék Fehérjetekeredés Peptidek és fehérjék 1 peptid: rövid, peptid kötéssel összekapcsolt aminosavakból álló polimer (< ~50 aminosav) fehérje: hosszú, peptid kötéssel összekapcsolt aminosavakból álló
Fehérjék Fehérjetekeredés Peptidek és fehérjék 1 peptid: rövid, peptid kötéssel összekapcsolt aminosavakból álló polimer (< ~50 aminosav) fehérje: hosszú, peptid kötéssel összekapcsolt aminosavakból álló
Bioinformatika 2 6. előadás
 6. előadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2018.10.08. PDBj: http://www.pdbj.org/ Fehérjék 3D szerkezeti adatbázisai - PDBj 2 2018.10.08.
6. előadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2018.10.08. PDBj: http://www.pdbj.org/ Fehérjék 3D szerkezeti adatbázisai - PDBj 2 2018.10.08.
A fehérjék hierarchikus szerkezete
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
A kovalens kötés elmélete. Kovalens kötésű molekulák geometriája. Molekula geometria. Vegyértékelektronpár taszítási elmélet (VSEPR)
") 4. előadás A kovalens kötés elmélete Vegyértékelektronpár taszítási elmélet (VSEPR) az atomok kötő és nemkötő elektronpárjai úgy helyezkednek el a térben, hogy egymástól minél távolabb legyenek A központi
4. előadás A kovalens kötés elmélete Vegyértékelektronpár taszítási elmélet (VSEPR) az atomok kötő és nemkötő elektronpárjai úgy helyezkednek el a térben, hogy egymástól minél távolabb legyenek A központi
A fehérjék hierarchikus szerkezete. Szerkezeti hierarchia. A fehérjék építőkövei az aminosavak. Fehérjék felosztása
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Energiaminimum- elve
 Energiaminimum- elve Minden rendszer arra törekszi, hogy stabil állapotba kerüljön. Milyen kapcsolat van a stabil állapot, és az adott állapot energiája között? Energiaminimum elve Energiaminimum- elve
Energiaminimum- elve Minden rendszer arra törekszi, hogy stabil állapotba kerüljön. Milyen kapcsolat van a stabil állapot, és az adott állapot energiája között? Energiaminimum elve Energiaminimum- elve
Fehérjék felépítése és struktúrája. Aminosav oldalláncok. A fehérjék királis elemekből (α-l-aminosavakból) épülnek fel
 épülnek fel") Fehérjék felépítése és struktúrája Aminosav oldalláncok A fehérjék királis elemekből (α-l-aminosavakból) épülnek fel Fehérjék szerkezete Anfinsen dogmája Anfinsen dogmája (vagy: termodinamikus hipotézis)
Fehérjék felépítése és struktúrája Aminosav oldalláncok A fehérjék királis elemekből (α-l-aminosavakból) épülnek fel Fehérjék szerkezete Anfinsen dogmája Anfinsen dogmája (vagy: termodinamikus hipotézis)
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Aromás: 1, 3, 5, 6, 8, 9, 10, 11, 13, (14) Az azulén (14) szemiaromás rendszert alkot, mindkét választ (aromás, nem aromás) elfogadtuk.
 Az azulén (14) szemiaromás rendszert alkot, mindkét választ (aromás, nem aromás) elfogadtuk.") 1. feladat Aromás: 1, 3, 5, 6, 8, 9, 10, 11, 13, (14) Az azulén (14) szemiaromás rendszert alkot, mindkét választ (aromás, nem aromás) elfogadtuk. 2. feladat Etil-metil-keton (bután-2-on) Jelek hozzárendelése:
1. feladat Aromás: 1, 3, 5, 6, 8, 9, 10, 11, 13, (14) Az azulén (14) szemiaromás rendszert alkot, mindkét választ (aromás, nem aromás) elfogadtuk. 2. feladat Etil-metil-keton (bután-2-on) Jelek hozzárendelése:
Kötések kialakítása - oktett elmélet
 Kémiai kötések Az elemek és vegyületek halmazai az atomok kapcsolódásával - kémiai kötések kialakításával - jönnek létre szabad atomként csak a nemesgázatomok léteznek elsődleges kémiai kötések Kötések
Kémiai kötések Az elemek és vegyületek halmazai az atomok kapcsolódásával - kémiai kötések kialakításával - jönnek létre szabad atomként csak a nemesgázatomok léteznek elsődleges kémiai kötések Kötések
Flagellin alapú filamentáris nanoszerkezetek létrehozása
 Flagellin alapú filamentáris nanoszerkezetek létrehozása Vonderviszt Ferenc PE MÜKKI Bio-Nanorendszerek Laboratórium MTA Enzimológiai Intézete MTA MFA Bakteriális flagellumok Flagelláris filamentum: ~10
Flagellin alapú filamentáris nanoszerkezetek létrehozása Vonderviszt Ferenc PE MÜKKI Bio-Nanorendszerek Laboratórium MTA Enzimológiai Intézete MTA MFA Bakteriális flagellumok Flagelláris filamentum: ~10
Intra- és intermolekuláris reakciók összehasonlítása
 Intra- és intermolekuláris reakciók összehasonlítása Intr a- és inter molekulár is r eakciok összehasonlítása molekulán belüli reakciók molekulák közötti reakciók 5- és 6-tagú gyűrűk könnyen kialakulnak.
Intra- és intermolekuláris reakciók összehasonlítása Intr a- és inter molekulár is r eakciok összehasonlítása molekulán belüli reakciók molekulák közötti reakciók 5- és 6-tagú gyűrűk könnyen kialakulnak.
Elektronegativitás. Elektronegativitás
 Általános és szervetlen kémia 3. hét Elektronaffinitás Az az energiaváltozás, ami akkor következik be, ha 1 mól gáz halmazállapotú atomból 1 mól egyszeresen negatív töltésű anion keletkezik. Mértékegysége:
Általános és szervetlen kémia 3. hét Elektronaffinitás Az az energiaváltozás, ami akkor következik be, ha 1 mól gáz halmazállapotú atomból 1 mól egyszeresen negatív töltésű anion keletkezik. Mértékegysége:
Nukleinsavak építőkövei
 ukleinsavak Szerkezeti hierarchia ukleinsavak építőkövei Pirimidin Purin Pirimidin Purin Timin (T) Adenin (A) Adenin (A) Citozin (C) Guanin (G) DS bázisai bázis Citozin (C) Guanin (G) RS bázisai bázis
ukleinsavak Szerkezeti hierarchia ukleinsavak építőkövei Pirimidin Purin Pirimidin Purin Timin (T) Adenin (A) Adenin (A) Citozin (C) Guanin (G) DS bázisai bázis Citozin (C) Guanin (G) RS bázisai bázis
Kémiai kötés. Általános Kémia, szerkezet Dia 1 /39
 Kémiai kötés 4-1 Lewis elmélet 4-2 Kovalens kötés: bevezetés 4-3 Poláros kovalens kötés 4-4 Lewis szerkezetek 4-5 A molekulák alakja 4-6 Kötésrend, kötéstávolság 4-7 Kötésenergiák Általános Kémia, szerkezet
Kémiai kötés 4-1 Lewis elmélet 4-2 Kovalens kötés: bevezetés 4-3 Poláros kovalens kötés 4-4 Lewis szerkezetek 4-5 A molekulák alakja 4-6 Kötésrend, kötéstávolság 4-7 Kötésenergiák Általános Kémia, szerkezet
Spektroszkópiai módszerek 2.
 Spektroszkópiai módszerek 2. NMR spektroszkópia magspinek rendeződése külső mágneses tér hatására az eredő magspin nem nulla, ha a magot alkotó nukleonok közül legalább az egyik páratlan a szerves kémiában
Spektroszkópiai módszerek 2. NMR spektroszkópia magspinek rendeződése külső mágneses tér hatására az eredő magspin nem nulla, ha a magot alkotó nukleonok közül legalább az egyik páratlan a szerves kémiában
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
3. A kémiai kötés. Kémiai kölcsönhatás
 3. A kémiai kötés Kémiai kölcsönhatás ELSŐDLEGES MÁSODLAGOS OVALENS IONOS FÉMES HIDROGÉN- KÖTÉS DIPÓL- DIPÓL, ION- DIPÓL, VAN DER WAALS v. DISZPERZIÓS Kémiai kötések Na Ionos kötés Kovalens kötés Fémes
3. A kémiai kötés Kémiai kölcsönhatás ELSŐDLEGES MÁSODLAGOS OVALENS IONOS FÉMES HIDROGÉN- KÖTÉS DIPÓL- DIPÓL, ION- DIPÓL, VAN DER WAALS v. DISZPERZIÓS Kémiai kötések Na Ionos kötés Kovalens kötés Fémes
Biológiai makromolekulák szerkezete
 Biológiai makromolekulák szerkezete Biomolekuláris nemkovalens kölcsönhatások Elektrosztatikus kölcsönhatások (sóhidak: 4-6 kcal/m, dipól-dipól: ~10-1 kcal/m Diszperziós erők (~10-2 kcal/m) Hidrogén hidak
Biológiai makromolekulák szerkezete Biomolekuláris nemkovalens kölcsönhatások Elektrosztatikus kölcsönhatások (sóhidak: 4-6 kcal/m, dipól-dipól: ~10-1 kcal/m Diszperziós erők (~10-2 kcal/m) Hidrogén hidak
A fehérjék szerkezetét fenntartó kölcsönhatások
 A fehérjék szerkezetét fenntartó kölcsönhatások Bionanorendszerek A fehérjék szerkezetét fenntartó kölcsönhatások alapvetően négy nagyobb csoportba oszthatók: 1. elektrosztatikus kölcsönhatások (sóhíd:
A fehérjék szerkezetét fenntartó kölcsönhatások Bionanorendszerek A fehérjék szerkezetét fenntartó kölcsönhatások alapvetően négy nagyobb csoportba oszthatók: 1. elektrosztatikus kölcsönhatások (sóhíd:
Biopolimer 12/7/09. Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. DNS. Polimerek. Kardos Roland DNS elsődleges szerkezete
 Biopolimerek Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. Osztódó sejt magorsófonala Kardos Roland 2009.10.29. Dohány levél epidermális sejtjének aktin hálózat Bakteriofágból kiszabaduló
Biopolimerek Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. Osztódó sejt magorsófonala Kardos Roland 2009.10.29. Dohány levél epidermális sejtjének aktin hálózat Bakteriofágból kiszabaduló
Aminosavak, peptidek, fehérjék. Béres Csilla
 Aminosavak, peptidek, fehérjék Béres Csilla Aminosavak Az aminosavak (más néven aminokarbonsavak) olyan szerves vegyületek, amelyek molekulájában aminocsoport (- NH 2 ) és karboxilcsoport (-COOH) egyaránt
Aminosavak, peptidek, fehérjék Béres Csilla Aminosavak Az aminosavak (más néven aminokarbonsavak) olyan szerves vegyületek, amelyek molekulájában aminocsoport (- NH 2 ) és karboxilcsoport (-COOH) egyaránt
Az enzimműködés termodinamikai és szerkezeti alapjai
 2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
Peptidek és fehérjék szerkezetvizsgálata spektroszkópia és in silico módszerekkel
 Peptidek és fehérjék szerkezetvizsgálata spektroszkópia és in silico módszerekkel Mik a peptidek és fehérjék? L-konfigurációjú a-aminosavakból felépülő lineáris polimerek 3 betűs kód: -Thr-His-Ile-Ser-Ser-Ile-Met-Pro-Leu-Glu-
Peptidek és fehérjék szerkezetvizsgálata spektroszkópia és in silico módszerekkel Mik a peptidek és fehérjék? L-konfigurációjú a-aminosavakból felépülő lineáris polimerek 3 betűs kód: -Thr-His-Ile-Ser-Ser-Ile-Met-Pro-Leu-Glu-
Speciális fluoreszcencia spektroszkópiai módszerek
 Speciális fluoreszcencia spektroszkópiai módszerek Fluoreszcencia kioltás Fluoreszcencia Rezonancia Energia Transzfer (FRET), Lumineszcencia A molekuláknak azt a fényemisszióját, melyet a valamilyen módon
Speciális fluoreszcencia spektroszkópiai módszerek Fluoreszcencia kioltás Fluoreszcencia Rezonancia Energia Transzfer (FRET), Lumineszcencia A molekuláknak azt a fényemisszióját, melyet a valamilyen módon
Modern Fizika Labor. Fizika BSc. Értékelés: A mérés dátuma: A mérés száma és címe: 13. mérés: Molekulamodellezés PC-n. 2008. április 29.
 Modern Fizika Labor Fizika BSc A mérés dátuma: A mérés száma és címe: 13. mérés: Molekulamodellezés PC-n Értékelés: A beadás dátuma: 2008. május 6. A mérést végezte: 1/5 A mérés célja A mérés célja az
Modern Fizika Labor Fizika BSc A mérés dátuma: A mérés száma és címe: 13. mérés: Molekulamodellezés PC-n Értékelés: A beadás dátuma: 2008. május 6. A mérést végezte: 1/5 A mérés célja A mérés célja az
Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei
 Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Gazdálkodási modul Gazdaságtudományi ismeretek I Közgazdasá Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei KÖRNYEZETGAZDÁLKODÁSI
Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Gazdálkodási modul Gazdaságtudományi ismeretek I Közgazdasá Adatgyűjtés, mérési alapok, a környezetgazdálkodás fontosabb műszerei KÖRNYEZETGAZDÁLKODÁSI
AMINOSAVAK, FEHÉRJÉK
 AMINOSAVAK, FEHÉRJÉK Az aminosavak olyan szerves vegyületek, amelyek molekulájában aminocsoport (-NH2) és karboxilcsoport (-COOH) egyaránt előfordul. Felosztás A fehérjéket feloszthatjuk aszerint, hogy
AMINOSAVAK, FEHÉRJÉK Az aminosavak olyan szerves vegyületek, amelyek molekulájában aminocsoport (-NH2) és karboxilcsoport (-COOH) egyaránt előfordul. Felosztás A fehérjéket feloszthatjuk aszerint, hogy
 9. Előadás Fehérjék Előzmények Peptidkémia Analitikai kémia Protein kémia 1901 E.Fischer : Gly-Gly 1923 F. Pregl : Mikroanalitika 1952 Stein and Moore : Aminosav analizis 1932 Bergman és Zervas : Benziloxikarbonil
9. Előadás Fehérjék Előzmények Peptidkémia Analitikai kémia Protein kémia 1901 E.Fischer : Gly-Gly 1923 F. Pregl : Mikroanalitika 1952 Stein and Moore : Aminosav analizis 1932 Bergman és Zervas : Benziloxikarbonil
Bio-nanorendszerek. Vonderviszt Ferenc. Pannon Egyetem Nanotechnológia Tanszék
 Bio-nanorendszerek Vonderviszt Ferenc Pannon Egyetem Nanotechnológia Tanszék Technológia: képesség az anyag szerkezetének, az anyagot felépítő részecskék elrendeződésének befolyásolására. A technológiai
Bio-nanorendszerek Vonderviszt Ferenc Pannon Egyetem Nanotechnológia Tanszék Technológia: képesség az anyag szerkezetének, az anyagot felépítő részecskék elrendeződésének befolyásolására. A technológiai
41. ábra A NaCl rács elemi cellája
 41. ábra A NaCl rács elemi cellája Mindkét rácsra jellemző, hogy egy tetszés szerint kiválasztott pozitív vagy negatív töltésű iont ellentétes töltésű ionok vesznek körül. Különbség a közvetlen szomszédok
41. ábra A NaCl rács elemi cellája Mindkét rácsra jellemző, hogy egy tetszés szerint kiválasztott pozitív vagy negatív töltésű iont ellentétes töltésű ionok vesznek körül. Különbség a közvetlen szomszédok
ALKÍMIA MA Az anyagról mai szemmel, a régiek megszállottságával.
 ALKÍMIA MA Az anyagról mai szemmel, a régiek megszállottságával www.chem.elte.hu/pr Kvíz az előző előadáshoz Programajánlatok október 18. 16:00 ELTE Kémiai Intézet 065-ös terem Észbontogató (www.chem.elte.hu/pr)
ALKÍMIA MA Az anyagról mai szemmel, a régiek megszállottságával www.chem.elte.hu/pr Kvíz az előző előadáshoz Programajánlatok október 18. 16:00 ELTE Kémiai Intézet 065-ös terem Észbontogató (www.chem.elte.hu/pr)
Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia II. kategória 2. forduló Megoldások
 ktatási Hivatal rszágos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia II. kategória 2. forduló Megoldások I. FELADATSR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D 9.
ktatási Hivatal rszágos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia II. kategória 2. forduló Megoldások I. FELADATSR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D 9.
Fehérjeszerkezet, és tekeredés. Futó Kinga
 Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
Thomson-modell (puding-modell)
") Atommodellek Thomson-modell (puding-modell) A XX. század elejére világossá vált, hogy az atomban található elektronok ugyanazok, mint a katódsugárzás részecskéi. Magyarázatra várt azonban, hogy mi tartja
Atommodellek Thomson-modell (puding-modell) A XX. század elejére világossá vált, hogy az atomban található elektronok ugyanazok, mint a katódsugárzás részecskéi. Magyarázatra várt azonban, hogy mi tartja
AZ ANYAGI HALMAZOK ÉS A MÁSODLAGOS KÖTÉSEK. Rausch Péter kémia-környezettan
 AZ ANYAGI HALMAZOK ÉS A MÁSODLAGOS KÖTÉSEK Rausch Péter kémia-környezettan Hogy viselkedik az ember egyedül? A kémiában ritkán tudunk egyetlen részecskét vizsgálni! - az anyagi részecske tudja hogy kell
AZ ANYAGI HALMAZOK ÉS A MÁSODLAGOS KÖTÉSEK Rausch Péter kémia-környezettan Hogy viselkedik az ember egyedül? A kémiában ritkán tudunk egyetlen részecskét vizsgálni! - az anyagi részecske tudja hogy kell
Altalános Kémia BMEVESAA101 tavasz 2008
 Folyadékok és szilárd anayagok 3-1 Intermolekuláris erők, folyadékok tulajdonságai 3-2 Folyadékok gőztenziója 3-3 Szilárd anyagok néhány tulajdonsága 3-4 Fázisdiagram 3-5 Van der Waals kölcsönhatások 3-6
Folyadékok és szilárd anayagok 3-1 Intermolekuláris erők, folyadékok tulajdonságai 3-2 Folyadékok gőztenziója 3-3 Szilárd anyagok néhány tulajdonsága 3-4 Fázisdiagram 3-5 Van der Waals kölcsönhatások 3-6
3. Sejtalkotó molekulák III.
 3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
Makromolekulák. I. A -vázas polimerek szerkezete és fizikai tulajdonságai. Pekker Sándor
 Makromolekulák I. A -vázas polimerek szerkezete és fizikai tulajdonságai Pekker Sándor MTA SZFKI Telefon:392-2222/845, Fax:392-229, Email: pekker@szfki.hu SZFKI tanfolyam: www.szfki.hu/moodle/course/ a
Makromolekulák I. A -vázas polimerek szerkezete és fizikai tulajdonságai Pekker Sándor MTA SZFKI Telefon:392-2222/845, Fax:392-229, Email: pekker@szfki.hu SZFKI tanfolyam: www.szfki.hu/moodle/course/ a
Fehérjeszerkezet, fehérjetekeredés
 Fehérjeszerkezet, fehérjetekeredés A fehérjeszerkezet szintjei A fehérjetekeredés elmélete: Anfinsen kísérlet Levinthal paradoxon A feltekeredés tölcsér elmélet 2014.11.05. Aminosavak és fehérjeszerkezet
Fehérjeszerkezet, fehérjetekeredés A fehérjeszerkezet szintjei A fehérjetekeredés elmélete: Anfinsen kísérlet Levinthal paradoxon A feltekeredés tölcsér elmélet 2014.11.05. Aminosavak és fehérjeszerkezet
T I T - M T T. Hevesy György Kémiaverseny. A megyei forduló feladatlapja. 7. osztály. A versenyző jeligéje:... Megye:...
 T I T - M T T Hevesy György Kémiaverseny A megyei forduló feladatlapja 7. osztály A versenyző jeligéje:... Megye:... Elért pontszám: 1. feladat:... pont 2. feladat:... pont 3. feladat:... pont 4. feladat:...
T I T - M T T Hevesy György Kémiaverseny A megyei forduló feladatlapja 7. osztály A versenyző jeligéje:... Megye:... Elért pontszám: 1. feladat:... pont 2. feladat:... pont 3. feladat:... pont 4. feladat:...
Sillabusz orvosi kémia szemináriumokhoz 1. Kémiai kötések
 Sillabusz orvosi kémia szemináriumokhoz 1. Kémiai kötések Pécsi Tudományegyetem Általános Orvostudományi Kar 2010-2011. 1 A vegyületekben az atomokat kémiai kötésnek nevezett erők tartják össze. Az elektronok
Sillabusz orvosi kémia szemináriumokhoz 1. Kémiai kötések Pécsi Tudományegyetem Általános Orvostudományi Kar 2010-2011. 1 A vegyületekben az atomokat kémiai kötésnek nevezett erők tartják össze. Az elektronok
NMR a peptid- és fehérje-kutatásban
 NMR a peptid- és fehérje-kutatásban A PDB adatbázisban megtalálható NMR alapú fehérjeszerkezetek számának alakulása az elmúlt évek során 4000 3500 3000 2500 2000 1500 1000 500 0 1987 1988 1989 1990 1991
NMR a peptid- és fehérje-kutatásban A PDB adatbázisban megtalálható NMR alapú fehérjeszerkezetek számának alakulása az elmúlt évek során 4000 3500 3000 2500 2000 1500 1000 500 0 1987 1988 1989 1990 1991
Aminosavak általános képlete NH 2. Csoportosítás: R oldallánc szerkezete alapján: Semleges. Esszenciális aminosavak
 Aminosavak 1 Aminosavak általános képlete N 2 soportosítás: oldallánc szerkezete alapján: Apoláris Poláris Bázikus Savas Semleges Esszenciális aminosavak 2 (apoláris) Glicin Név Gly 3 Alanin Ala 3 3 Valin
Aminosavak 1 Aminosavak általános képlete N 2 soportosítás: oldallánc szerkezete alapján: Apoláris Poláris Bázikus Savas Semleges Esszenciális aminosavak 2 (apoláris) Glicin Név Gly 3 Alanin Ala 3 3 Valin
Elválasztástechnikai és bioinformatikai kutatások. Dr. Harangi János DE, TTK, Biokémiai Tanszék
 Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Összefoglalók Kémia BSc 2012/2013 I. félév
 Összefoglalók Kémia BSc 2012/2013 I. félév Készült: Eötvös Loránd Tudományegyetem Kémiai Intézet Szerves Kémiai Tanszékén 2012.12.17. Összeállította Szilvágyi Gábor PhD hallgató Tartalomjegyzék Orgován
Összefoglalók Kémia BSc 2012/2013 I. félév Készült: Eötvös Loránd Tudományegyetem Kémiai Intézet Szerves Kémiai Tanszékén 2012.12.17. Összeállította Szilvágyi Gábor PhD hallgató Tartalomjegyzék Orgován
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak
 BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
Kormeghatározás gyorsítóval
 Beadás határideje 2012. január 31. A megoldásokat a kémia tanárodnak add oda! 1. ESETTANULMÁNY 9. évfolyam Olvassa el figyelmesen az alábbi szöveget és válaszoljon a kérdésekre! Kormeghatározás gyorsítóval
Beadás határideje 2012. január 31. A megoldásokat a kémia tanárodnak add oda! 1. ESETTANULMÁNY 9. évfolyam Olvassa el figyelmesen az alábbi szöveget és válaszoljon a kérdésekre! Kormeghatározás gyorsítóval
4. FEHÉRJÉK. 2. Vázanyagok. Az izmok alkotórésze (pl.: a miozin). Inak, izületek, csontok szerves komponensei, az ún. vázfehérjék (szkleroproteinek).
. Inak, izületek, csontok szerves komponensei, az ún. vázfehérjék (szkleroproteinek).") 4. FEÉRJÉK 4.0. Bevezetés A fehérjék elsısorban α-l-aminosavakból felépülı biopolimerek. A csak α-laminosavakat tartalmazó fehérjék a proteinek. evüket a görög proteios szóból kapták, ami elsırangút jelent.
4. FEÉRJÉK 4.0. Bevezetés A fehérjék elsısorban α-l-aminosavakból felépülı biopolimerek. A csak α-laminosavakat tartalmazó fehérjék a proteinek. evüket a görög proteios szóból kapták, ami elsırangút jelent.
Kémiai kötés. Általános Kémia, szerkezet Slide 1 /39
 Kémiai kötés 12-1 Lewis elmélet 12-2 Kovalens kötés: bevezetés 12-3 Poláros kovalens kötés 12-4 Lewis szerkezetek 12-5 A molekulák alakja 12-6 Kötésrend, kötéstávolság 12-7 Kötésenergiák Általános Kémia,
Kémiai kötés 12-1 Lewis elmélet 12-2 Kovalens kötés: bevezetés 12-3 Poláros kovalens kötés 12-4 Lewis szerkezetek 12-5 A molekulák alakja 12-6 Kötésrend, kötéstávolság 12-7 Kötésenergiák Általános Kémia,
A cukrok szerkezetkémiája
 A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
Fehérjeszerkezet analízis. Fehérjeszerkezet analízis. Fehérjeszerkezet analízis. Fehérjeszerkezet analízis. Fehérjeszerkezet analízis
 Szerkezet Protein Data Bank (PDB) http://www.rcsb.org/pdb ~ 35 701 szerkezet közepes felbontás 1552 szerkezet d 1.5 Å 160 szerkezet d 1.0 Å 10 szerkezet d 0.8 Å (atomi felbontás) E globális minimum? funkció
Szerkezet Protein Data Bank (PDB) http://www.rcsb.org/pdb ~ 35 701 szerkezet közepes felbontás 1552 szerkezet d 1.5 Å 160 szerkezet d 1.0 Å 10 szerkezet d 0.8 Å (atomi felbontás) E globális minimum? funkció
FELADATMEGOLDÁS. Tesztfeladat: Válaszd ki a helyes megoldást!
 FELADATMEGOLDÁS Tesztfeladat: Válaszd ki a helyes megoldást! 1. Melyik sorozatban található jelölések fejeznek ki 4-4 g anyagot? a) 2 H 2 ; 0,25 C b) O; 4 H; 4 H 2 c) 0,25 O; 4 H; 2 H 2 ; 1/3 C d) 2 H;
FELADATMEGOLDÁS Tesztfeladat: Válaszd ki a helyes megoldást! 1. Melyik sorozatban található jelölések fejeznek ki 4-4 g anyagot? a) 2 H 2 ; 0,25 C b) O; 4 H; 4 H 2 c) 0,25 O; 4 H; 2 H 2 ; 1/3 C d) 2 H;
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
A kémiai kötés magasabb szinten
 A kémiai kötés magasabb szinten 11-1 Mit kell tudnia a kötéselméletnek? 11- Vegyérték kötés elmélet 11-3 Atompályák hibridizációja 11-4 Többszörös kovalens kötések 11-5 Molekulapálya elmélet 11-6 Delokalizált
A kémiai kötés magasabb szinten 11-1 Mit kell tudnia a kötéselméletnek? 11- Vegyérték kötés elmélet 11-3 Atompályák hibridizációja 11-4 Többszörös kovalens kötések 11-5 Molekulapálya elmélet 11-6 Delokalizált
Műszeres analitika II. (TKBE0532)
") Műszeres analitika II. (TKBE0532) 7. előadás NMR spektroszkópia Dr. Andrási Melinda Debreceni Egyetem Természettudományi és Technológiai Kar Szervetlen és Analitikai Kémiai Tanszék NMR, Nuclear Magnetic
Műszeres analitika II. (TKBE0532) 7. előadás NMR spektroszkópia Dr. Andrási Melinda Debreceni Egyetem Természettudományi és Technológiai Kar Szervetlen és Analitikai Kémiai Tanszék NMR, Nuclear Magnetic
Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete
 Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete Polipeptidek térszerkezete Tipikus (rendezett) konformerek em tipikus (rendezetlen) konformerek Periodikus vagy homokonformerek Aperiodikus
Peptid- és fehérjék másodlagos-, harmadlagos- és negyedleges szerkezete Polipeptidek térszerkezete Tipikus (rendezett) konformerek em tipikus (rendezetlen) konformerek Periodikus vagy homokonformerek Aperiodikus
Cikloalkánok és származékaik konformációja
 1 ikloalkánok és származékaik konformációja telített gyűrűs szénhidrogének legegyszerűbb képviselője a ciklopropán. Gyűrűje szabályos háromszög alakú, ennek megfelelően szénatomjai egy síkban helyezkednek
1 ikloalkánok és származékaik konformációja telített gyűrűs szénhidrogének legegyszerűbb képviselője a ciklopropán. Gyűrűje szabályos háromszög alakú, ennek megfelelően szénatomjai egy síkban helyezkednek
A BAKTERIORODOPSZIN. Péter Imre AINLHQ
 A BAKTERIORODOPSZIN Péter Imre AINLHQ BEVEZETÉS A napfény energiáját az élőlények (növények, algák) egy bonyolult folyamat, a fotoszintézis során alakítják át és tárolják. Létezik egy baktérium, a Halobacterium
A BAKTERIORODOPSZIN Péter Imre AINLHQ BEVEZETÉS A napfény energiáját az élőlények (növények, algák) egy bonyolult folyamat, a fotoszintézis során alakítják át és tárolják. Létezik egy baktérium, a Halobacterium
Kémiai reakciók mechanizmusa számítógépes szimulációval
 Kémiai reakciók mechanizmusa számítógépes szimulációval Stirling András stirling@chemres.hu Elméleti Kémiai Osztály Budapest Stirling A. (MTA Kémiai Kutatóközpont) Reakciómechanizmus szimulációból 2007.
Kémiai reakciók mechanizmusa számítógépes szimulációval Stirling András stirling@chemres.hu Elméleti Kémiai Osztály Budapest Stirling A. (MTA Kémiai Kutatóközpont) Reakciómechanizmus szimulációból 2007.
R R C X C X R R X + C H R CH CH R H + BH 2 + Eliminációs reakciók
 Eliminációs reakciók Amennyiben egy szénatomhoz távozó csoport kapcsolódik és ugyanazon a szénatomon egy (az ábrákon vel jelölt) bázis által protonként leszakítható hidrogén is található, a nukleofil szubsztitúció
Eliminációs reakciók Amennyiben egy szénatomhoz távozó csoport kapcsolódik és ugyanazon a szénatomon egy (az ábrákon vel jelölt) bázis által protonként leszakítható hidrogén is található, a nukleofil szubsztitúció
ELEMI RÉSZECSKÉK ATOMMODELLEK
 ELEMI RÉSZECSKÉK ATOMMODELLEK Az atomok felépítése Készítette: Horváthné Vlasics Zsuzsanna Mi van az atomok belsejében? DÉMOKRITOSZ (Kr.e. 460-370) az anyag nem folytonos parányi, tovább nem bontható,
ELEMI RÉSZECSKÉK ATOMMODELLEK Az atomok felépítése Készítette: Horváthné Vlasics Zsuzsanna Mi van az atomok belsejében? DÉMOKRITOSZ (Kr.e. 460-370) az anyag nem folytonos parányi, tovább nem bontható,
Kémiai kötés. Általános Kémia, szerkezet Slide 1 /39
 Kémiai kötés 4-1 Lewis elmélet 4-2 Kovalens kötés: bevezetés 4-3 Poláros kovalens kötés 4-4 Lewis szerkezetek 4-5 A molekulák alakja 4-6 Kötésrend, kötéstávolság 4-7 Kötésenergiák Általános Kémia, szerkezet
Kémiai kötés 4-1 Lewis elmélet 4-2 Kovalens kötés: bevezetés 4-3 Poláros kovalens kötés 4-4 Lewis szerkezetek 4-5 A molekulák alakja 4-6 Kötésrend, kötéstávolság 4-7 Kötésenergiák Általános Kémia, szerkezet
Atomszerkezet. Atommag protonok, neutronok + elektronok. atompályák, alhéjak, héjak, atomtörzs ---- vegyérték elektronok
 Atomszerkezet Atommag protonok, neutronok + elektronok izotópok atompályák, alhéjak, héjak, atomtörzs ---- vegyérték elektronok periódusos rendszer csoportjai Periódusos rendszer A kémiai kötés Kémiai
Atomszerkezet Atommag protonok, neutronok + elektronok izotópok atompályák, alhéjak, héjak, atomtörzs ---- vegyérték elektronok periódusos rendszer csoportjai Periódusos rendszer A kémiai kötés Kémiai
I. Atomszerkezeti ismeretek (9. Mozaik Tankönyv:10-30. oldal) 1. Részletezze az atom felépítését!
 1. Részletezze az atom felépítését!") I. Atomszerkezeti ismeretek (9. Mozaik Tankönyv:10-30. oldal) 1. Részletezze az atom felépítését! Az atom az anyagok legkisebb, kémiai módszerekkel tovább már nem bontható része. Az atomok atommagból és
I. Atomszerkezeti ismeretek (9. Mozaik Tankönyv:10-30. oldal) 1. Részletezze az atom felépítését! Az atom az anyagok legkisebb, kémiai módszerekkel tovább már nem bontható része. Az atomok atommagból és
TRANSZPORTFOLYAMATOK A SEJTEKBEN
 16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások
 Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Biomolekulák nanomechanikája A biomolekuláris rugalmasság alapjai
 Fogorvosi Anyagtan Fizikai Alapjai Biomolekulák nanomechanikája A biomolekuláris rugalmasság alapjai Mártonfalvi Zsolt Biofizikai és Sugárbiológiai Intézet Semmelweis Egyetem Budapest Biomolekulák mint
Fogorvosi Anyagtan Fizikai Alapjai Biomolekulák nanomechanikája A biomolekuláris rugalmasság alapjai Mártonfalvi Zsolt Biofizikai és Sugárbiológiai Intézet Semmelweis Egyetem Budapest Biomolekulák mint
Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET)
") Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET) Biofizika szeminárium PTE ÁOK Biofizikai Intézet Huber Tamás 2014. 02. 11-13. A gerjesztett állapotú elektron lecsengési lehetőségei Gerjesztés Fluoreszcencia
Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET) Biofizika szeminárium PTE ÁOK Biofizikai Intézet Huber Tamás 2014. 02. 11-13. A gerjesztett állapotú elektron lecsengési lehetőségei Gerjesztés Fluoreszcencia
folsav, (a pteroil-glutaminsav vagy B 10 vitamin) dihidrofolsav tetrahidrofolsav N CH 2 N H H 2 N COOH
 dihidrofolsav tetrahidrofolsav N CH 2 N H H 2 N COOH") folsav, (a pteroil-glutaminsav vagy B 10 vitamin) 2 2 2 2 pirimidin rész pirazin rész aminobenzoesav rész glutaminsav rész pteridin rész dihidrofolsav 2 2 2 2 tetrahidrofolsav 2 2 2 2 A dihidrofolát-reduktáz
folsav, (a pteroil-glutaminsav vagy B 10 vitamin) 2 2 2 2 pirimidin rész pirazin rész aminobenzoesav rész glutaminsav rész pteridin rész dihidrofolsav 2 2 2 2 tetrahidrofolsav 2 2 2 2 A dihidrofolát-reduktáz
Szemináriumi feladatok (alap) I. félév
 I. félév") Szemináriumi feladatok (alap) I. félév I. Szeminárium 1. Az alábbi szerkezet-párok közül melyek reprezentálják valamely molekula, vagy ion rezonancia-szerkezetét? Indokolja válaszát! A/ ( ) 2 ( ) 2 F/
Szemináriumi feladatok (alap) I. félév I. Szeminárium 1. Az alábbi szerkezet-párok közül melyek reprezentálják valamely molekula, vagy ion rezonancia-szerkezetét? Indokolja válaszát! A/ ( ) 2 ( ) 2 F/
Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET) Modern Biofizikai Kutatási Módszerek
 Modern Biofizikai Kutatási Módszerek") Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET) Modern Biofizikai Kutatási Módszerek 2012. 11. 08. Fotonok és molekulák ütközése Fény (foton) ütközése a molekulákkal fényszóródás abszorpció E=hν
Fluoreszcencia módszerek (Kioltás, Anizotrópia, FRET) Modern Biofizikai Kutatási Módszerek 2012. 11. 08. Fotonok és molekulák ütközése Fény (foton) ütközése a molekulákkal fényszóródás abszorpció E=hν
Vezetők elektrosztatikus térben
 Vezetők elektrosztatikus térben Vezető: a töltések szabadon elmozdulhatnak Ha a vezető belsejében a térerősség nem lenne nulla akkor áram folyna. Ha a felületen a térerősségnek lenne tangenciális (párhuzamos)
Vezetők elektrosztatikus térben Vezető: a töltések szabadon elmozdulhatnak Ha a vezető belsejében a térerősség nem lenne nulla akkor áram folyna. Ha a felületen a térerősségnek lenne tangenciális (párhuzamos)
Adatgyőjtés, mérési alapok, a környezetgazdálkodás fontosabb mőszerei
 GazdálkodásimodulGazdaságtudományismeretekIKözgazdaságtan KÖRNYEZETGAZDÁLKODÁSIMÉRNÖKIMScTERMÉSZETVÉDELMIMÉRNÖKIMSc Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Adatgyőjtés, mérési
GazdálkodásimodulGazdaságtudományismeretekIKözgazdaságtan KÖRNYEZETGAZDÁLKODÁSIMÉRNÖKIMScTERMÉSZETVÉDELMIMÉRNÖKIMSc Tudományos kutatásmódszertani, elemzési és közlési ismeretek modul Adatgyőjtés, mérési
A kémiai kötés. Kémiai kölcsönhatás
 A kémiai kötés Kémiai kölcsönhatás ELSŐDLEGES MÁSODLAGOS KOVALENS IONOS FÉMES HIDROGÉN- KÖTÉS DIPÓL- DIPÓL, ION- DIPÓL, VAN DER WAALS v. DISZPERZIÓS Ionos kötés Na Cl Ionpár képződése e - Na + Cl - Na:
A kémiai kötés Kémiai kölcsönhatás ELSŐDLEGES MÁSODLAGOS KOVALENS IONOS FÉMES HIDROGÉN- KÖTÉS DIPÓL- DIPÓL, ION- DIPÓL, VAN DER WAALS v. DISZPERZIÓS Ionos kötés Na Cl Ionpár képződése e - Na + Cl - Na:
Enzimek. Enzimek! IUBMB: szisztematikus nevek. Enzimek jellemzése! acetilkolin-észteráz! legalább 10 nagyságrend gyorsulás. szubsztrát-specificitás
 Enzimek acetilkolin-észteráz! Enzimek! [s -1 ] enzim víz carbonic anhydrase 6x10 5 10-9 karbonikus anhidráz acetylcholine esterase 2x10 4 8x10-10 acetilkolin észteráz staphylococcal nuclease 10 2 2x10-14
Enzimek acetilkolin-észteráz! Enzimek! [s -1 ] enzim víz carbonic anhydrase 6x10 5 10-9 karbonikus anhidráz acetylcholine esterase 2x10 4 8x10-10 acetilkolin észteráz staphylococcal nuclease 10 2 2x10-14
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Bioinformatika 2 5.. előad
 5.. előad adás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 03. 21. Fehérje térszerkezet t megjelenítése A fehérjék meglehetősen összetett
5.. előad adás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 03. 21. Fehérje térszerkezet t megjelenítése A fehérjék meglehetősen összetett
4.3. Az AcLysSerProValValGluGly heptapeptid Al(III)-kötő sajátságának jellemzése
-kötő sajátságának jellemzése") 4.3. Az AcLysProValValGly heptapeptid Al(III)-kötő sajátságának jellemzése Az Al(III)fehérje kölcsönhatás megismerése céljából olyan neurofilamentum peptidfragmenst vizsgáltunk, mely az oldalláncban oxigén
4.3. Az AcLysProValValGly heptapeptid Al(III)-kötő sajátságának jellemzése Az Al(III)fehérje kölcsönhatás megismerése céljából olyan neurofilamentum peptidfragmenst vizsgáltunk, mely az oldalláncban oxigén
A tananyag felépítése: A BIOLÓGIA ALAPJAI. I. Prokarióták és eukarióták. Az eukarióta sejt. Pécs Miklós: A biológia alapjai
 A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
NMR spektroszkópia a fehérje biokémiában
 NMR spektroszkópia a fehérje biokémiában Závodszky Péter Beinrohr László MTA SzBK Enzimológiai Intézet NMR spektroszkópia a fehérje biokémiában Závodszky Péter Beinrohr László MTA SzBK Enzimológiai Intézet
NMR spektroszkópia a fehérje biokémiában Závodszky Péter Beinrohr László MTA SzBK Enzimológiai Intézet NMR spektroszkópia a fehérje biokémiában Závodszky Péter Beinrohr László MTA SzBK Enzimológiai Intézet
6. változat. 3. Jelöld meg a nem molekuláris szerkezetű anyagot! A SO 2 ; Б C 6 H 12 O 6 ; В NaBr; Г CO 2.
 6. változat Az 1-től 16-ig terjedő feladatokban négy válaszlehetőség van, amelyek közül csak egy helyes. Válaszd ki a helyes választ és jelöld be a válaszlapon! 1. Jelöld meg azt a sort, amely helyesen
6. változat Az 1-től 16-ig terjedő feladatokban négy válaszlehetőség van, amelyek közül csak egy helyes. Válaszd ki a helyes választ és jelöld be a válaszlapon! 1. Jelöld meg azt a sort, amely helyesen