Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
|
|
|
- Éva Tóth
- 7 évvel ezelőtt
- Látták:
Átírás


1 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A
2 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Tananyag címe: PROTEIN BIOTECHNOLÓGIA Tantárgyfelelős: Dr. Tőzsér József


3 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 1. előadás A FEHÉRJÉK BIOKÉMIAI TULAJDONSÁGAI. A FEHÉRJÉK SZINTÉZISE. A PROKARIÓTA ÉS EUKARIÓTA FEHÉRJESZINTÉZIS KÖZTI KÜLÖNBSÉGEK ÁTTEKINTÉSE.
4 A fehérjéket felépítő leggyakoribb 20 aminosav Alanin Arginin Aszparagin Aszpartát Cisztein Glutamin Glutamát Glicin Hisztidin Izoleucin Leucin Lizin Metionin Fenilalanin Prolin Szerin Treonin Triptofán Tirozin Valin

5 A globuláris fehérjék szerkezete TÁMOP /1/A

6 A detergensek csoportosítása töltöttségük szerint Detergensek: Nem ionos Anionos Kationos Amfoter


7 A detergensek hatása a fehérjékre A. B. EC PM aggregátum + IC +
8 A redukálószerek hatása a fehérjék szerkezetére A. Fehérje S S Fehérje SH SH B. SDS Urea Tiourea Fehérje S S Fehérje S S SDS Urea + Tiourea β-merkaptoetanol DTT TCEP Fehérje SH SH

9 A trns szerkezete A. B. C.
10 Az aminosavak aktiválása, a trns feltöltése Met 1. Met-tRNS szintetáz + PPi Met-AMP ATP 2. Met trns Met AMP Aminoacil-tRNS
11 A prokarióta és az eukarióta mrns szerkezete A prokarióta mrns Riboszóma kötőhely Shine-Dalgarno szekvencia Shine-Dalgarno szekvencia: AGGAGGxxAUG 5 5 UTR Start kodon (AUG) Kódoló régió fmet-t kódol 3 UTR Stop kodon (UAG, UAA, UGA) 3 Az eukarióta mrns Riboszóma kötőhely 5 m7 G 5 sapka Kozak szekvencia Kozak szekvencia: Gcc 5 UTR Kódoló régió 3 UTR Start kodon Stop kodon (AUG) Met-t kódol (UAG, UAA, UGA) A G ccaugg AAAAAA 3 polia farok
12 A prokarióta és eukarióta mrns-ek közötti különbség A. DNS Promóter Operátor Gén 1 Gén 2 Gén 3 Shine-Dalgarno szekvencia Shine-Dalgarno szekvencia Shine-Dalgarno szekvencia mrns AUG AUG AUG AUG fehérje fmet Met fmet fmet B. Gén DNS 5 m 7 G AUG AUG mrns AAAAAA 3 Met Met fehérje


13 A riboszómák szerkezete Prokarióta riboszóma (70S) 50S nagy alegység 23S RNS 5S RNS 35 fehérje 70S riboszóma 30S kis alegység 16S RNS 21 fehérje Eukarióta riboszóma (80S) 60S nagy alegység 28S RNS 5S RNS, 5.8S RNS 49 fehérje 80S riboszóma 40S kis alegység 18S RNS 33 fehérje
14 A működő riboszóma szerkezete P hely a peptid lánctrns kötődésére Naszcens peptid lánc A hely az aminoacil-trns kötődésére 60S nagy alegység E P 5 3 A A riboszóma haladási iránya mrns E hely az üres trns távozására 40S kis alegység
15 A fehérjeszintézis iniciációja prokariótákban IF-1 IF-2 30S alegység IF-3 fmet Shine-Dalgarno szekvencia 5 5 UTR AUG 3 UTR 3
16 A fehérjeszintézis iniciációja eukariótákban 40S alegység Met Kozak szekvencia eif-2 5 m7 G AUG 5 UTR 3 UTR AAAAAA 3
17 Az iniciáció befejeződése GDP eif-2 Met 60S alegység Met GTP eif-2 5 m7 G 5 UTR AUG 40S alegység 3 UTR AAAAAA 3
18 A fehérje szintézis második lépése: az elongáció Ala GTP EF-Tu GDP EF-Tu Met Ala Met Ala GTP 5 mrns E P A 3 EF-Tu Új aminoacil-trns belépése 5 E P A 3 mrns Met Ala GDP EF-G GTP EF-G Met Ala Új peptid kötés kialakulása (Peptidil transzferáz) 5 E P A 3 mrns Transzlokáció 5 E P A 3 mrns
19 Az elongáció jellegzetességei Riboszóma haladási iránya NH 3 mrns + NH 3 Peptidlánc növekedésének iránya + NH 3
20 A fehérje szintézis harmadik lépése: a termináció RF1 5 UAG 3 mrns STOP kodon + NH 3 RF1 5 UAG 3 mrns + NH 3 COO -
21 A fehérje szintézis helye az eukariótákban Sejtmag DNS mrns Citoszol + NH 3 + NH 3 mrns + NH 3
22 A fehérje szintézis helye a prokariótákban TÁMOP /1/A
23 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 2. előadás A FEHÉRJÉK FELTEKEREDÉSE, A 3D SZERKEZET KIALAKULÁSA. CHAPERONOK. A FOLDING HIBÁI, FOLDING BETEGSÉGEK
24 Az élő rendszerekben előforduló főbb kötéstípusok: hidrogén hidak Kovalens kötés Hidrogén donor Hidrogén akceptor N H N H O H O H 3,1 Å 3,04 Å 2,88 Å 2,7 Å N O N O Erős hidrogén hidak O O H Gyenge hidrogén híd Hidrogén kötés/hidrogén híd

25 Az élő rendszerekben előforduló főbb kötéstípusok: elektrosztatikus kölcsönhatás A. R COO - 2,8 Å + H 3 N R B.

26 Az élő rendszerekben előforduló főbb kötéstípusok: hidrofób kölcsönhatás A. B. apoláros molekula apoláros molekula apoláros molekula apoláros molekula
27 A víz poláros molekula, az élő rendszerekben stabilizálja a szerkezetet A. B

28 A fehérjék feltekeredése/foldingja TÁMOP /1/A


29 A fehérjék szerkezete Másodlagos szerkezet Lokális rendezett szerkezet Elsődleges szerkezet Aminosav sorrend Harmadlagos szerkezet A polipeptid lánc térbeli szerkezete Negyedleges szerkezet Több polipeptid láncból álló fehérje szerkezete
30 A fehérjék elsődleges szerkezete: az aminosav-sorrend N-terminus + NH 3 S S S S COO - C-terminus

31 Az egyes aminosav egységeket összetartó peptid kötés TÁMOP /1/A

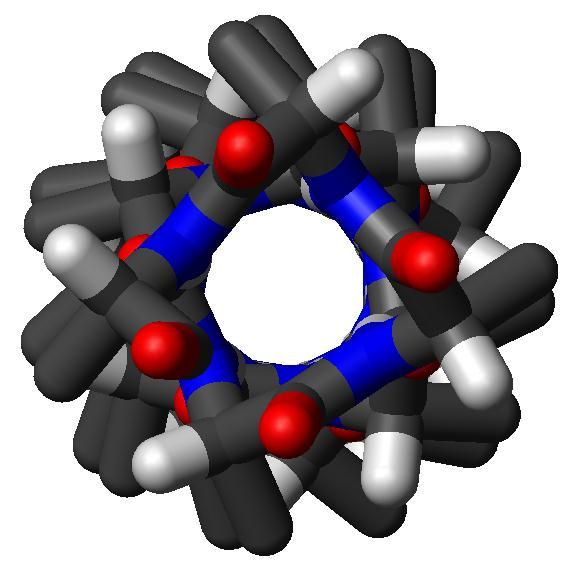
32 Az alfa hélix A. B.

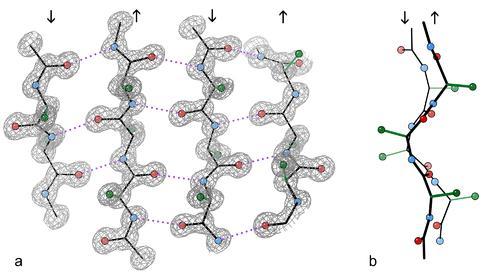
33 A béta redő Béta redő Béta fordulat

34 A fehérje szerkezetének jóslása a hidrofób aminosavak pozíciója alapján Hidrofobicitási skála Hidrofób aminosavak belül, hidrofilok kívül



35 A fehérjék harmadlagos szerkezete másodlagos szerkezeti elemekből épül fel Másodlagos szerkezeti elemek Modulok Harmadlagos szerkezet
36 Az Anfinsen kísérlet RNáz RNáz RNáz Natív forma (100 % aktivitás) Redukálás Széttekerés (8M urea) Denaturált forma (0 % aktivitás) Urea eltávolítása Oxidálás Natív forma (90 % aktivitás) RNáz RNáz RNáz Natív forma (100 % aktivitás) Redukálás Széttekerés (8M urea) Denaturált forma (0 % aktivitás) Oxidálás Urea eltávolítása Véletlenszerűen feltekeredett forma (1-2 % aktivitás)
37 Szabad energia csökken TÁMOP /1/A Lehetséges konformációk száma csökken A fehérjék feltekeredése metastabil köztes állapotokon keresztül Folding funnel
38 A rendezetlen fehérjék szerkezete 1. alegység 2. alegység Rendezetlen régió

39 A rendezetlen fehérjék szerkezete a fehérje-fehérje interakciók során módosulhat TÁMOP /1/A

40 A GroEL chaperon kristályszerkezeti képe TÁMOP /1/A

41 A chaperonok működési mechanizmusa Chaperon Hibás szerkezet Natív szerkezet
42 A chaperonok szerepe a fehérjék feltekeredésének elősegítésében
43 A chaperonok osztályozása szerkezetük alapján Hsp oligomerek Hsp 60 oligomerek Hsp 70 monomerek Hsp 90 dimerek Hsp 110 oligomerek
44 A Hsp60 működése Hibás szerkezet Anfinsen kalitka Natív szerkezet hsp10 ATP ADP 15 s ATP ATP ADP ADP ADP ADP ATP ATP hsp60 ADP ATP
45 A Hsp70 működése TÁMOP /1/A
46 A Hsp90 működése Hsp70 Hsp40 HOP DNS kötő domén Hsp90 Hip Hsp90 HOP SHR Éretlen receptor hormon kötésre nem képes Hsp70 HOP Hsp40 Hip Hsp90 p23 SHR Foldoszóma Érett receptor hormon kötésre képes Hormon SHR Hip p23 SHR SHR DNS kötés ATP Immunofilin peptidil-prolil cisz-transz izomeráz
47 A Hsp110 működése Natív fehérje Natív fehérje Denaturált fehérje ATP Hsp 110 Aggregálódott fehérje hsp70

48 A kalnexin és kalretikulin működése TÁMOP /1/A
49 A protein diszulfid izomeráz (PDI) működése HS- PDI SH HS- S S SH PDI SH SH PDI SH S S S S 1. konformáció 2. konformáció
50 A peptidil-prolil izomeráz működése cisz Pro transz Pro


51 Szabadenergia TÁMOP /1/A A folding során kialakuló abnormális szerkezetű fehérjék megjelenése Entrópia abnormális állapot natív állapot
52 A rossz prionok számának növekedése Natív, endogén PrP c PrP sc interakciója endogén PrP c -vel Spontán PrP sc keletkezik PrP sc interakciója endogén PrP c -vel
 denaturált aggregáció Amiloid rostok Amiloid plakkok Amiloid rostok")
53 Az amiloid plakkok kialakulásának feltételezett mechanizmusa olvadt (molten globule) natív olvadt (molten globule) denaturált aggregáció Amiloid rostok Amiloid plakkok Amiloid rostok natív
54 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 3. előadás A FEHÉRJÉK CÉLBAJUTTATÁSA (SORTING ÉS TARGETING)
55 A fehérjék irányítása az endoplazmatikus retikulumba I. ER membrán 5. SRP receptor SRP ER lumen mrns 3 citoszól
56 A fehérjék irányítása az endoplazmatikus retikulumba II. ER membrán Szignál peptidáz 6. ER lumen SRP receptor mrns citoszól 5 5. SRP 7. 3
57 A fehérjék irányítása az endoplazmatikus retikulumba III. ER membrán 10. SRP receptor 8. ER lumen 5 mrns SRP citoszól UAA 9. 3
58 A fehérjék endoplazmatikus retikulumba történő irányítása és kotranszlációs módosítása
59 A fehérjék kotranszlációs N-glikozilációja az endoplazmatikus retikulumban
60 Az N-glikoziláció mechanizmusa, az oligoszacharid lánc kialakulása dolikol-foszfáton TÁMOP /1/A N-Acetil glükózamin Mannóz Glükóz Reorientáció P Dolikol-foszfát Citoszól P P P P P P P ER membrán P P P P AsnXxxSer/Thr ER lumen 3 4
61 Minőség-ellenőrzés az endoplazmás retikulumban Helyesen feltekeredett fehérje Hibásan feltekeredett fehérje PDI Grp78 ERAD chaperon ER lumen Citoszól Fehérje lebontása U U U U U Proteoszóma
62 A fehérjék útja a különböző kompartmentek között sejtmag 1. DER 2. Plazma membrán Lizoszóma 7. citoszól Cisz Golgi hálózat Extracelluláris tér Sejtmag Golgi ciszternák Szekretoros vezikula 7. Citoszól Transz Golgi hálózat
63 A fehérjék módosítása a Golgi kompartmentjeiben N-Acetil glükózamin P Mannóz Glükóz P P P P
64 A Golgi készülékben változatos cukortartalmú N-glikozilált fehérjék keletkeznek N-Acetil glükózamin Mannóz Glükóz Galaktóz Mannóz gazdag típus Lizoszóma Sziálsav Komplex típus Extracelluláris tér/plazma membrán
65 A fehérjék irányítása a mitokondriumba C Hsp 70 C Külső membrán N C N Import receptor Belső membrán mhsp 70 N Mitokondriális mátrix N C
66 A fehérjék irányítása a sejtmagba TÁMOP /1/A
67 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 4. előadás A FEHÉRJESZERKEZET TANULMÁNYOZÁSA: RÖNTGEN KRISZTALLOGRÁFIA, NMR, TÖMEGSPEKTROMETRIA


68 A kristályszerkezet és a fehérje rendezettsége közötti viszony Rendezettség jóslás a szekvencia alapján - IUPred (iupred.enzim.hu) A fehérje kristályszerkezete

69 A fehérjeszerkezet megállapítása Röntgen krisztallográfiával I Fehérjék kristályosítása Megfelelő kristályok kiválasztása Nagy energiájú Röntgen sugárzás A fehérje kristályban levő elektronokon ütközik a Röntgen sugár és jellegzetes diffrakciós képet hoz létre
 Diffrakciós kép")
70 A fehérjeszerkezet megállapítása Röntgen krisztallográfiával II Elektroneloszlási kép (Fourier transzformációt felhasználva) Diffrakciós kép Illesztett szerkezet

71 Az aminosav-sorrend illesztés minősége meghatározza a kristályszerkezet minőségét 5 Å 3 Å 1,5 Å


72 A fehérjeszerkezet megállapítása mágneses magrezonancia (NMR) segítségével
")
73 A megállapított fehérjeszerkezetek tárolása a PDB (Protein Data Bank) adatbázisban
74 A tömegspektrométerek felépítése Minta bemenet Elektronika Ion forrás Tömeg analizátor Detektor Adatfeldolgozó rendszer Vákuum rendszer Tömeg spektrum
75 A MALDI Mátrix által segített lézer deszorpció ionizáció (Matrix Assisted Laser Desorption Ionization) működési elve Mintatartó lemez Minta Lézer AH Analizátor Detektor Mátrix kv
76 TÁMOP /1/A Az elektrosprey ionizáció (ESI) elve Szárító gáz Fűtött lemez LC 2-4 kv Kapilláris Analizátor Detektor
77 Az ionok útja a repülési idő (Time-Of-Flight) TOF analizátorban Ionforrás Analizátor Detektor
78 A tömegspektrométer felbontásának javítása a reflektron segítségével Reflektron detektor
79 Az ionok útja a kvadrupólban Kvadrupól Detektor
80 Az ionok útja az ioncsapdában Ioncsapda Detektor
81 Aminosavszekvencia meghatározása elektrospray ionizációs tandem MS (ESI MS/MS) készülékkel m/z m/z ESI ionforrás Q1 Ütközési cella Q3 Detektor QFIENGSEFAQK
82 Intenzitás Intenzitás TÁMOP /1/A Az ionok útja a nagyfelbontású tömegspektrométerben (HDMS) IMS T-hullám Ionforrás Detektor (GRGDS) ,7 Å 2 (SDGRG) ,7 Å 2 Sodródási idő Érkezési idő (ms)
83 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 5. előadás FEHÉRJÉK TISZTÍTÁSA (KROMATOGRÁFIÁS TISZTÍTÁSI MÓDSZEREK) ÉS ANALÍZISE (SDS- PAGE, 2DE, TÖMEGSPEKTROMETRIA)
84 Lépések száma A megfelelő fehérjetisztítási stratégia kiválasztásának szempontjai Tisztaság 4. A kisebb szennyeződések eltávolítása 3. A nagyobb szennyeződések eltávolítása 2. Izolálás, koncentrálás és stabilizálás 1. Extrakció
85 Fehérjék tisztítása affinitás kromatográfiával A megfelelő fehérje hozzákötődése az oszlophoz A nem kötődött fehérjék lemosása A specifikusan kötődött fehérjék elúciója
86 Fehérjék tisztítása/elválasztása analitikai gélszűréssel Vizes oldat Vizes oldat Vizes oldat Vizes oldat
87 Fehérjék sómentesítése dialízissel TÁMOP /1/A
88 SDS-poliakrilamid gélektroforézis - Vándorlási irány +
89 Fehérjék izoelektromos fókuszálása ph 3 ph 10

90 Kétdimenziós gélelektroforézis TÁMOP /1/A



91 A gélben levő fehérjék vizualizálása különféle festési módszerekkel SyproRuby festés oomassie festés ph 3 ph 10 4% Ezüst festés 18%
92 A fehérjék tisztítása immunprecipitációval (IP) Y Y Y Protein A/G beadek + Antitest Mosás Y Y Indirekt IP specifikus kötődése Y Y Y Y Mosás Y Centrifugálás Ag-Ab kötés hasítása Direkt IP

93 A fehérjék vizsgálata Western blot segítségével Blottolás PVDF membrán Megfelelő antitest hozzáadása


94 A proteomikai munkafolyamat szemléltetése Fehérjét tartalmazó foltok kivágása Enzimes (tripszin) emésztés A sejt lízise 2DE Fehérjék azonosítása Tömegspektrometriás analízis
95 Kvalitatív és kvantitatív különbségek analízise kétdimenziós gélelektroforézis segítségével A Minta 2DE B Minta 2DE gélanalízis kvalitatív és kvantitatív különbségek
 Jelölés DIGE festékkel H. influenzae (aktinonin kezelt) Minták összekeverése 2DE")
96 Kvalitatív és kvantitatív különbségek analízise differenciál gélelektroforézis (DIGE) segítségével H. influenzae (Kontrol) Jelölés DIGE festékkel H. influenzae (aktinonin kezelt) Minták összekeverése 2DE
97 intenzitás TÁMOP /1/A SILAC - Sejtkultúrák metabolikus jelölése stabil izotópok segítségével Tápfolyadék normál Arg-el (könnyű Arg) Tápfolyadék 13 C tartalmú Arg-el (nehéz Arg) Sejtek lízise és homogenizálása Minta összekeverése MS/MS 6 Da m/z
98 Az itraq jelölőanyag szerkezete Jelölő csoport (114, 115, 116, 117) Kiegyenlítő csoport Fehérjéhez való kötődésért felelős csoport Izobár csoport
99 intenzitás intenzitás TÁMOP /1/A Kémiai jelölés itraq módszerrel Kontroll Kezelt sejt 60 min Kezelt sejt 120 min Sejtek összekeverése MS 117 Kezelt sejt 240 min m/z MS/MS 115 Fehérje mennyiségi adat a görbe alatti területből számolva m/z
100 Célzott fehérjék kimutatása MRM segítségével m/z m/z ESI ionforrás Q1 Ütközési cella Q3 Detektor Komplex fehérje elegy Meghatározott m/z-jű ionok kiválasztása és fragmentációja Meghatározott m/z-jű ionok kiválasztása és detektálása
101 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 6. előadás A FEHÉRJÉK POSZT-TRANSZLÁCIÓS MÓDOSÍTÁSA ÉS A MÓDOSÍTÁSOK KIMUTATÁSA PROTEOMIKAI MÓDSZEREKKEL



102 Glikálás Glükóz Glükóz + Glükóz HbA1c


103 A fehérjék módosítása foszforiláció és defoszforiláció révén Ser Tyr ATP Kináz ADP Tyr Kináz Tyr Foszfatáz P Thr Ser/Thr Kináz Ser/Thr Foszfatáz Foszfatáz Pi
104 A fehérjék módosítása preniláció révén Farneziláció Geraniláció 3
105 A fehérjék módosítása zsírsav módosítások révén Palmitinsav Mirisztinsav
106 A fehérjék módosítása proteolízis révén Limitált proteolízis Degradáció

107 A proteolitikus hasítás helye TÁMOP /1/A
108 A poszt-transzlációs módosítások géntranszkripciót befolyásoló szerepe TÁMOP /1/A
109 Izopeptid kötés létrehozása a transzglutaminázok által katalizált reakcióban Gln Lys Ca 2+ Ca 2+ Ca 2+ TG2 Gln Lys Ca 2+ Ca 2+ Ca 2+

110 Poszt-transzlációs módosítások kimutatására szolgáló specifikus festési eljárások Sypro Ruby Minden fehérjét megfest ProQ Diamond Csak a foszfoproteineket festi ProQ Emerald Csak a glikoproteineket festi
111 A fehérjék oldalláncán levő foszfát csoport sorsa a tömegspektrometriás analízis során Ser P Foszfoproteinek Dúsítás (TiO2, IMAC) Ser P Ser P Ser P CID PO 3-79 Da H 3 PO 4 vesztés 98 Da
112 Prekurzor ion keresés m/z m/z 79 ESI ionforrás Q1 Ütközési cella Q3 Detektor
113 Semleges vesztés m/z m/z ESI ionforrás Q1 Ütközési cella Q3 Detektor
114 Poszt-transzlációs módosítások kimutatása MRM segítségével ESI ionforrás m/z Q1 Ütközési cella m/z Q3
115 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 7. előadás FEHÉRJE-FEHÉRJE KÖLCSÖNHATÁSOK KIALAKULÁSA ÉS VIZSGÁLATI LEHETŐSÉGEI

116 Interakciós hálózat Csomópontok hub-ok
117 Fehérje-fehérje interakciók vizsgálata ko-immunoprecipitációval Antitest a komplex egyik tagja ellen Fehérje komplex Immunprecipitáció Y Sávok kivágása és tripszines emésztése Tripszines emésztés SDS-PAGE A peptid keverék LC-MS/MS analízise A peptidek MS/MS analízise Az ismert fehérjével kapcsolatban levő fehérjék azonosítása
118 Fehérje-fehérje interakciók vizsgálata pull-down technikával x Y X elleni antitest Fehérje komplex IP x Y X ellenes antitest Y ellenes antitest SDS-PAGE Western blot
119 Y TÁMOP /1/A Fehérje-fehérje interakciók vizsgálata far-western technikával Immobilizált X fehérje x x Y x Y Y + Y fehérje + Y fehérje elleni antitest Előhívás Membrán
120 Dithiobisz-szulfoszukcinimidil-propionát TÁMOP /1/A
121 Bisz-szulfoszukcinimidil-szuberát TÁMOP /1/A
122 Fehérje-fehérje interakciók vizsgálata fotoaktív keresztkötő ágensek segítségével TÁMOP /1/A Foto-Leu Foto-Met tartalmú tápfolyadék UV Keresztkötés alakul ki az egymástól pár Å-re levő fehérjék között (interakciós partnerek) Interakciós partnerek azonosítása Keresztkötött fehérjék további analízise
123 Fehérje-fehérje interakciók vizsgálata élesztő két hibrid rendszer segítségével Gal4BD domén Gal4AD domén BD lacz Y Y X lacz BD X X lacz Y + BD Y X lacz
124 Fehérje-fehérje kölcsönhatások kimutatása fehérje chipek alkalmazásával Y Indirekt módszer Direkt módszer Y Y Üvegfelület Y Y Y Y Üvegfelület Y Üvegfelület Antitest chip Antitest chip A pozitív jelet adó helyek detektálása
125 Fehérje chipeken megkötött fehérjék analízise SELDI technika segítségével Fehérje chip Komplex fehérje elegy Mátrix hozzákeverése lézer SELDI lemez Detektor Analizátor MALDI-TOF
126 Fág bemutató rendszer Megfelelő fágok kötődése Specifikus elúció és felszaporítás mátrix
127 Fehérje-fehérje interakciók vizsgálata FRET fluoreszcencia rezonancia energia transzfer segítségével h 436 nm Y CFP Donor csoport 480 nm h 436 nm Y CFP 480 nm FRET X YFP Akceptor csoport X YFP 535 nm
128 Fehérje-fehérje interakciók vizsgálata felületi plazmon rezonancia (surface plasmon resonance) elvén Fényforrás Szenzor chip P-polarizált fény Prizma Visszavert fény Detektor Felületi arany réteg Minta Y Y Y Y Felületi plazmon Áramlási cella Immobilizált ligand
129 Felületi plazmon rezonancia elvén alapuló fehérje-fehérje interakciók vizsgálata Biacore készülékkel Rezonancia jel (kru) 18 Y Y Y Y Disszociáció Y Y Y Y x Y k d k a x Y k d Y Y Y Y 12 Asszociáció Regenerálás Y Y Y Y k a Koncentráció Y Y Y Y Idő (s)
130 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 8. előadás HETEROLÓG EXPRESSZIÓ I
 E.")
131 Az expressziós vektor részei az expresszáló organizmusban működő replikációs origó (replikatív vektorok) egyéb szekvenciák (pl. a plazmid stabil replikációját, vagy integrácóját segítő gének, a promóter működését szabályozó gének) E. coli szelekciós marker gén (shuttle vektorok) a hatékony expresszióhoz szükséges szekvenciák (promóter régió, riboszómakötő szekvenciák, transzkripcó terminációs szekvenciák stb.) az expresszálandó gén (cdns, intronokkal mesterségesen kiegészített cdns is lehet; szignál szekvencia beépítése, eltávolítása, ritka kodonok számának növelése, csökkentése, tag szekvenciák beépítése stb.) E. coli replikációs origó (shuttle vektorok) az expresszáló organizmusban működő szelekciós marker gén (replikatív vektoroknál fontos)
132 Az RNS I és RNS II szerepe a plazmidok replikációjában RNS II RNS I trns ori rop replikáció nincs replikáció fokozott replikáció
133 Levan sucrase szelekción alapuló integráció kétszeres rekombináció egyszeres rekombináció integrálódott kazetta integrálódott plazmid
134 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 9. előadás HETEROLÓG EXPRESSZIÓ II
135 A Sec és a Tat szekréciós útvonalak összehasonlítása extracelluláris tér Sec folding Tat citoszol transzláció transzláció és folding
136 A Sec-útvonal felépítése és működése extracelluláris tér SPáz sece secy secg secdf citoszol seca ftsy yajc b- SRP
137 A S. carnosus expressziós kazetta felépítése promóter SP pro gén CWA
138 A Saccharomyces cerevisiaere és a Pichia pastorisra jellemző N-glikozid oldalláncok P S. cerevisiae mannán -típus 2 NacGlc és 50 < Man n x Asn S. cerevisiae core -típus 2 NacGlc és <15 Man b-1,4-n-acetil glükózamin b-1,4-mannóz a-1,3-mannóz a-1,6-mannóz a-1,2-mannóz a-mannóz Asn Asn P. pastoris
139 Az integráció alternatívája: mesterséges kromoszómák centromer replikációs origó, marker gén, shuttle vektror szekvenciák inzertált gének (a megfelelő promóterrel és segítő szekvenciákkal ellátva) telomer szekvenciák (a közöttük lévő szakaszt vágják ki a transzformáció előtt, hogy linearizálják a DNS-t)
140 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 10. előadás HETEROLÓG EXPRESSZIÓ III
141 Irányított integráció, funkcionalizált sejtek A B C
142 Néhány jellegzetes növényi N-glikozid oldallánc Az ER lumenben kialakuló alapváz: Asn Asn Asn Asn Asn b-1,4-n-acetil glükózamin b-1,4-mannóz a-1,3-mannóz a-1,6-mannóz a-1,2-mannóz b-1,2-n-acetil glükózamin a-1,3-fukóz b-1,2-xilóz b-1,3-galaktóz a-1,4-fukóz

143 T DNS Az Agrobacterium tumefaciens Ti plazmid részei
144 Transzformálás Agrobacterium tumefaciens segítségével növényi sejt T-DNS Agrobacterium
145 N-glikoziláció rovar sejtekben Az ER lumenben kialakuló alapváz: Asn Asn Asn Asn Asn b-1,4-n-acetil glükózamin b-1,4-mannóz a-1,3-mannóz a-1,6-mannóz a-1,2-mannóz b-1,2-n-acetil glükózamin a-1,3-fukóz a-1,6-fukóz
146 Az expressziós kazetta beépítése a bacmidba 1. Tn7L és R szekvenciák E. coli donor plazmid transzpozázt kódoló segítő plazmid bacmid LacZ gén - Tn7 célszekvencia LacZ promóter E. coli donor plazmid 2. transzpozázt kódoló segítő plazmid a bacmidba beépült expressziós kazetta bacmid
147 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 11. előadás PROTEIN ENGINEERING

148 A de novo protein design jelentősége TÁMOP /1/A
149 Helix: -Gly-Glu-Leu-Glu-Glu-Leu-Leu-Lys-Lys-Leu-Lys-Glu-Leu-Leu-Lys-Gly Loop: Pro-Arg-Arg Egy de novo protein design segítségével létrehozott fehérje szerkezete csak helix helix-loop-helix végső szerkezet NH 2 -Met-helix-loop-helix-loop-helix-loop-helix-COOH
150 A receptor tirozin kináz (RTK) működése ligand RTK membrán P P P P P intracelluláris hatások
151 Mesterséges növekedési faktorok oligomerizációs scaffold linker receptor kötő modul oligomer (dimer)
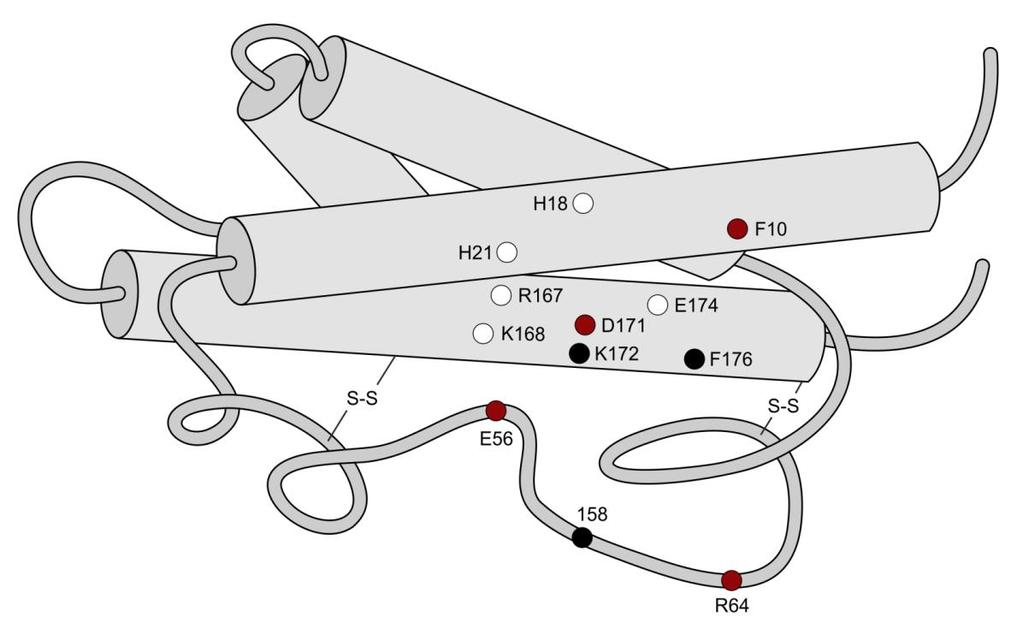
152 Receptor-specifikus peptid hormon változatok kifejlesztése A B
153 PCR alapú site directed mutagenesis deléció pont mutáció inzerció kivágásra szánt régió pont mutáció a primerben nem komplementer loop a primerben
154 Irányított evolúció gén vad típusú fehérje mutáns gének könyvtára ismétlés mutáns fehérjék könyvtára screening/szelekció kedvező tulajdonságú változat
155 DNA Shuffling 1 gén 1 gén 2 restrikciós emésztés ligálás rekombináns génváltozatok
156 DNA Shuffling 2 gén 1 gén 2 DNáz kezelés a fragmensek összekeverése, de- és renaturálása PCR primer nélkül PCR az eredeti végekre tervezett primerekkel
157 Staggered extension process (StEP) A B C D E
158 Exon shuffling homológ gének különböző láncvégződésű domainek az eredeti domaineket vegyesen tartalmazó rekombináns gének
159 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 12. előadás HUMÁNTERÁPIÁS FEHÉRJÉK ELŐÁLLÍTÁSA
160 Az inzulin szerkezete Gly Ile Val Glu Gln Cys Cys Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asp Val Phe Glu Hys Leu Cys Gly Ser Hys Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Thr Lys Pro Thr Tyr Phe Phe Gly Arg Glu Asn Tyr Cys Asn
161 vérszérum koncentráció TÁMOP /1/A Különféle inzulin változatok farmakokinetikai tulajdonsága aspart, lispro, glulisine inzulin NPH detemir glargine idő (h)

162 A hepatitis B vírus felépítés TÁMOP /1/A

163 A hepatitis B vírus egyszerűsített életciklusa TÁMOP /1/A

164 A hepatitis B vírus genotípusok földrajzi eloszlása TÁMOP /1/A
165 A Hansenula poliyorpha expressziós vektora TÁMOP /1/A
166 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 13. előadás HUMÁNTERÁPIÁS ENZIMEK ELŐÁLLÍTÁSA
167 Urát oxidáz (hugysav hidroxiláz) - H N O O N urát H N N H O O H H O 2 O + O H 2 N O H N + H N O 2 2 O N 5-hidroxiizourát xantin allantoin ureát + glioxilát
168 A human -galaktozidáz P P P P P P N-acetilglükózamin mannóz galaktóz sziálsav fukóz
169 A humán glükocerebrozidáz H O H O H O O O O O H N O H n 1 2 glükozilceramid +H 2 O H O H O H O O O H O H O H O N O H n glükóz ceramid
170 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Emri Tamás Protein biotechnológia 14. előadás DIAGNOSZTIKÁBAN HASZNÁLT ENZIMEK ELŐÁLLÍTÁSA
171 A GOX (glükóz oxidáz) által katalizált reakció O H O H H O O O H GOX O + H O O + H 2 O 2 O 2 H O O H b -D-glükóz H O +H 2 O O H D-glükono- d -lakton H O O H D-glükonsav O H C O O H H O O H
172 A GOX működése O H H 2 O 2 FAD H O H O O O H O H GOX O H O 2 FADH 2 H O O O H O O H

173 A GOX gyártása süllyesztett kultúrákban TÁMOP /1/A
174 A GAOX (galaktóz oxidáz) által katalizált reakció O H O H O O O H GAOX O + O 2 H O O + H 2 O 2 H O O H D-galaktóz H O O H 6-oxo-D-galaktóz
175 A GAOX működése +O 2 Cu + H O S Cys Tyr -H 2 O 2 + R-OH. O Cu 2+ S Cys Tyr -R=O
176 A ChOX (koleszterol oxidáz) által katalizált reakciók koleszterol H O FAD CHOX O FADH + H+ kolest-5-én-3-on CHOX O 2 H 2 O 2 O kolest-4-én-3-on
177 A HrP (tormaperoxidáz) által katalizált reakció O O O O O O H H 2 O H 2 O gvajakol O O O O tetragvajakol (vörösesbarna)
178 A HrP felhasználása immunoassay rendszerekben tormaperoxidázzal jelölt szekunder antitest O N H N H + 2OH - 2H 2 O 2 antigén HRP N H 2 O hv (~428 nm) C O O - primer antitest C O O N H H 2 O N 2 luminol oxidálása H 2 O 2 jelenlétében fotopapír
179 A HrP tartalmú bioszenzorok elektród H O O O H 2e - 2H + N O O OH H O O O H O Amplex Red N resorufin N H dihidroresorufin oxidáz szubsztrát H 2 O 2 2 HO. 2 H 2 O HRP
180 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP /1/A Csősz Éva Protein biotechnológia 15. előadás A TERÁPIÁS FEHÉRJÉK FELHASZNÁLÁSA, JÖVŐBENI LEHETŐSÉGEK, PERSPEKTÍVÁK. A TERÁPIÁS FEHÉRJÉK ALKALMAZÁSÁVAL KAPCSOLATOS NEMZETKÖZI ÉS HAZAI KÖVETELMÉNYEK
181 A génterápia és fehérjeterápia összehasonlítása Génterápia Fehérje terápia DNS izolálása a beteg szervezetéből Mutálódott gén kicserélése egy normál génre Fehérje bejuttatása intravénásan, intramuszkulárisan, per os A normál gén beépítése vírus vektorba A normál gén bejuttatása a beteg szervezetébe a vektor segítségével
182 Antitest függő citotoxicitás mechanizmusa Tumorellenes antitestek Fc receptor Tumor sejt Ölő sejt (NK sejt vagy monocita)
183 Her2 ellenes antitestek alkalmazása emlőrák kezelésében Trasztuzumab Her2 Her2 Her2 Her2 Her2 Fc receptor Tumor sejt Ölő sejt (NK sejt vagy monocita)

184 A TNF alfát gátló fehérjeterápiás készítmények hatása Adalimumab Infliximab Golimumab Cetrolizumab pegol monoklonális antitestek Etanercept szolubilis TNF alfa receptor TNF alfa Pszoriázis Reumatoid arthritisz Crohn betegség Spondilitisz TNF alfa gátlás
Fehérjebiotechnológia Emri, Tamás Csősz, Éva Tőzsér, József Szerkesztette Tőzsér, József, Debreceni Egyetem
 Fehérjebiotechnológia Emri, Tamás Csősz, Éva Tőzsér, József Szerkesztette Tőzsér, József, Debreceni Egyetem Fehérjebiotechnológia írta Emri, Tamás, Csősz, Éva, Tőzsér, József, Tőzsér, József, és Szerzői
Fehérjebiotechnológia Emri, Tamás Csősz, Éva Tőzsér, József Szerkesztette Tőzsér, József, Debreceni Egyetem Fehérjebiotechnológia írta Emri, Tamás, Csősz, Éva, Tőzsér, József, Tőzsér, József, és Szerzői
1b. Fehérje transzport
 1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
Fehérjebiotechnológia
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Fehérjebiotechnológia Emri Tamás Csősz Éva Tőzsér József
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Fehérjebiotechnológia Emri Tamás Csősz Éva Tőzsér József
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI AZ AMINOSAVAK ÉS FEHÉRJÉK 1. kulcsszó cím: Aminosavak Egy átlagos emberben 10-12 kg fehérje van, mely elsősorban a vázizomban található.
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE
 ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE") Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak?
 TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis,
TRANSZLÁCIÓ és fehérje transzport Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis,
3. Sejtalkotó molekulák III.
 3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
Proteomika Peptid szekvenálás. Dr. Csősz Éva Debreceni Egyetem ÁOK Biokémiai és Molekuláris Biológiai Intézet Proteomika Szolgáltató Laboratórium
 Proteomika Peptid szekvenálás Dr. Csősz Éva Debreceni Egyetem ÁOK Biokémiai és Molekuláris Biológiai Intézet Proteomika Szolgáltató Laboratórium Fókuszterületek Omikák, rendszerbiológia hipotézis vezérelt
Proteomika Peptid szekvenálás Dr. Csősz Éva Debreceni Egyetem ÁOK Biokémiai és Molekuláris Biológiai Intézet Proteomika Szolgáltató Laboratórium Fókuszterületek Omikák, rendszerbiológia hipotézis vezérelt
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét. Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet
 Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
DNS replikáció. DNS RNS Polipeptid Amino terminus. Karboxi terminus. Templát szál
 DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek
 Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
A sejtek élete. 5. Robotoló törpék és óriások Az aminosavak és fehérjék R C NH 2. C COOH 5.1. A fehérjeépítőaminosavak általános
 A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
A sejtek élete 5. Robotoló törpék és óriások Az aminosavak és fehérjék e csak nézd! Milyen protonátmenetes reakcióra képes egy aminosav? R 2 5.1. A fehérjeépítőaminosavak általános képlete 5.2. A legegyszerűbb
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak?
 A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
A TRANSZLÁCIÓ Hogyan lesz a DNS-ben kódolt információból fehérje? A DNS felszínén az aminosavak sorba állnak? mrns, trns, riboszómák felfedezése A GENETIKAI KÓD 20 AS és csak 4 bázis, a kódolás hogy lehetséges?
TRANSZPORTFOLYAMATOK 1b. Fehérjék. 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS
 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
Fehérjeszerkezet, és tekeredés
 Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Fehérjeszerkezet, és tekeredés Futó Kinga 2013.10.08. Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
15. Fehérjeszintézis: transzláció. Fehérje lebontás (proteolízis)
") 15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
DER (Felületén riboszómák találhatók) Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.
 Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.") Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK
 POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
Transzláció. Leolvasás - fehérjeszintézis
 Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
Transzláció Leolvasás - fehérjeszintézis Fehérjeszintézis DNS mrns Transzkripció Transzláció Polipeptid A trns - aminosav kapcsolódás 1 A KEZDETEK ELŐTT Az enzim aktiválja az aminosavat azáltal, hogy egy
Bioinformatika 2 5.. előad
 5.. előad adás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 03. 21. Fehérje térszerkezet t megjelenítése A fehérjék meglehetősen összetett
5.. előad adás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 03. 21. Fehérje térszerkezet t megjelenítése A fehérjék meglehetősen összetett
A fehérjék hierarchikus szerkezete
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A tananyag felépítése: A BIOLÓGIA ALAPJAI. I. Prokarióták és eukarióták. Az eukarióta sejt. Pécs Miklós: A biológia alapjai
 A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A fehérjék hierarchikus szerkezete
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Bay Péter
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Bay Péter Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
Bioinformatika előadás
 10. előadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat Genomika vs. proteomika A genomika módszereivel nem a tényleges fehérjéket vizsgáljuk,
10. előadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat Genomika vs. proteomika A genomika módszereivel nem a tényleges fehérjéket vizsgáljuk,
Fehérjeszerkezet, és tekeredés. Futó Kinga
 Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
Fehérjeszerkezet, és tekeredés Futó Kinga Polimerek Polimer: hasonló alegységekből (monomer) felépülő makromolekulák Alegységek száma: tipikusan 10 2-10 4 Titin: 3,435*10 4 aminosav C 132983 H 211861 N
ENZIMEK BIOTECHNOLÓGIAI ELŐÁLLÍTÁSA
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ENZIMEK BIOTECHNOLÓGIAI
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ENZIMEK BIOTECHNOLÓGIAI
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Fehérjeszerkezet, fehérjetekeredés
 Fehérjeszerkezet, fehérjetekeredés A fehérjeszerkezet szintjei A fehérjetekeredés elmélete: Anfinsen kísérlet Levinthal paradoxon A feltekeredés tölcsér elmélet 2014.11.05. Aminosavak és fehérjeszerkezet
Fehérjeszerkezet, fehérjetekeredés A fehérjeszerkezet szintjei A fehérjetekeredés elmélete: Anfinsen kísérlet Levinthal paradoxon A feltekeredés tölcsér elmélet 2014.11.05. Aminosavak és fehérjeszerkezet
Zsírsav szintézis. Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P. 2 i
 Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
A piruvát-dehidrogenáz komplex. Csala Miklós
 A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
Bioinformatika 2 10.el
 10.el őadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 04. 24. Genomikavs. proteomika A genomika módszereivel nem a tényleges fehérjéket
10.el őadás Prof. Poppe László BME Szerves Kémia és Technológia Tsz. Bioinformatika proteomika Előadás és gyakorlat 2009. 04. 24. Genomikavs. proteomika A genomika módszereivel nem a tényleges fehérjéket
Korszerű tömegspektrometria a. Szabó Pál MTA Kémiai Kutatóközpont
 Korszerű tömegspektrometria a biokémi miában Szabó Pál MTA Kémiai Kutatóközpont Tematika Bevezetés: ionizációs technikák és analizátorok összehasonlítása a biomolekulák szemszögéből Mikromennyiségek mintaelőkészítése
Korszerű tömegspektrometria a biokémi miában Szabó Pál MTA Kémiai Kutatóközpont Tematika Bevezetés: ionizációs technikák és analizátorok összehasonlítása a biomolekulák szemszögéből Mikromennyiségek mintaelőkészítése
Hemoglobin - myoglobin. Konzultációs e-tananyag Szikla Károly
 Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
A fehérjék szerkezete és az azt meghatározó kölcsönhatások
 A fehérjék szerkezete és az azt meghatározó kölcsönhatások 1. A fehérjék szerepe az élõlényekben 2. A fehérjék szerkezetének szintjei 3. A fehérjék konformációs stabilitásáért felelõs kölcsönhatások 4.
A fehérjék szerkezete és az azt meghatározó kölcsönhatások 1. A fehérjék szerepe az élõlényekben 2. A fehérjék szerkezetének szintjei 3. A fehérjék konformációs stabilitásáért felelõs kölcsönhatások 4.
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
CzB 2010. Élettan: a sejt
 CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
 9. Előadás Fehérjék Előzmények Peptidkémia Analitikai kémia Protein kémia 1901 E.Fischer : Gly-Gly 1923 F. Pregl : Mikroanalitika 1952 Stein and Moore : Aminosav analizis 1932 Bergman és Zervas : Benziloxikarbonil
9. Előadás Fehérjék Előzmények Peptidkémia Analitikai kémia Protein kémia 1901 E.Fischer : Gly-Gly 1923 F. Pregl : Mikroanalitika 1952 Stein and Moore : Aminosav analizis 1932 Bergman és Zervas : Benziloxikarbonil
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL
 TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa. Gergely Pál 2009
 FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
FEHÉRJESZINTÉZIS: a transzláció mechanizmusa és a polipeptidlánc további sorsa Gergely Pál 2009 Fehérjeszintézis és poszttranszlációs módosítások A kódszótár A riboszóma szerkezete A fehérjeszintézis (transzláció)
Elválasztástechnikai és bioinformatikai kutatások. Dr. Harangi János DE, TTK, Biokémiai Tanszék
 Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Röntgendiffrakció, tömegspektrometria, infravörös spektrometria.
 A biomolekuláris szerkezet és dinamika vizsgálómódszerei: Röntgendiffrakció, tömegspektrometria, infravörös spektrometria. Smeller László A molekuláris szerkezet és dinamika vizsgáló módszereinek áttekintése
A biomolekuláris szerkezet és dinamika vizsgálómódszerei: Röntgendiffrakció, tömegspektrometria, infravörös spektrometria. Smeller László A molekuláris szerkezet és dinamika vizsgáló módszereinek áttekintése
Élettan. előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45
 Élettan előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45 oktató: Dr. Tóth Attila, adjunktus ELTE TTK Biológiai Intézet, Élettani és Neurobiológiai tanszék
Élettan előadás tárgykód: bf1c1b10 ELTE TTK, fizika BSc félév: 2015/2016., I. időpont: csütörtök, 8:15 9:45 oktató: Dr. Tóth Attila, adjunktus ELTE TTK Biológiai Intézet, Élettani és Neurobiológiai tanszék
A biokémia alapjai. Typotex Kiadó. Wunderlich Lívius Szarka András
 A biokémia alapjai Wunderlich Lívius Szarka András Összefoglaló: A jegyzet elsősorban egészségügyi mérnök MSc. hallgatók részére íródott, de hasznos segítség lehet biomérnök és vegyészmérnök hallgatók
A biokémia alapjai Wunderlich Lívius Szarka András Összefoglaló: A jegyzet elsősorban egészségügyi mérnök MSc. hallgatók részére íródott, de hasznos segítség lehet biomérnök és vegyészmérnök hallgatók
Biopolimer 12/7/09. Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. DNS. Polimerek. Kardos Roland DNS elsődleges szerkezete
 Biopolimerek Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. Osztódó sejt magorsófonala Kardos Roland 2009.10.29. Dohány levél epidermális sejtjének aktin hálózat Bakteriofágból kiszabaduló
Biopolimerek Makromolekulák szerkezete. Fehérje szerkezet, és tekeredés. Osztódó sejt magorsófonala Kardos Roland 2009.10.29. Dohány levél epidermális sejtjének aktin hálózat Bakteriofágból kiszabaduló
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
CIÓ A GENETIKAI INFORMÁCI A DNS REPLIKÁCI
 A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
A GENETIKAI INFORMÁCI CIÓ TÁROLÁSA ÉS S KIFEJEZŐDÉSE A DNS SZERKEZETE Két antiparalel (ellentétes lefutású) polinukleotid láncból álló kettős helix A két lánc egy képzeletbeli közös tengely körül van feltekeredve,
A fehérjék szerkezeti hierarchiája. Fehérje-szerkezetek! Klasszikus szerkezet-funkció paradigma. szekvencia. funkció. szerkezet! Myoglobin.
 Myoglobin Fehérje-szerkezetek! MGLSDGEWQLVLNVWGKVEADIPGGQEVLIRLFK GPETLEKFDKFKLKSEDEMKASE DLKKGATVLTALGGILKKKGEAEIKPLAQSA TKKIPVKYLEFISECIIQVLQSK PGDFGADAQGAMNKALELFRKDMASNYKELGFQG Fuxreiter Mónika! Debreceni
Myoglobin Fehérje-szerkezetek! MGLSDGEWQLVLNVWGKVEADIPGGQEVLIRLFK GPETLEKFDKFKLKSEDEMKASE DLKKGATVLTALGGILKKKGEAEIKPLAQSA TKKIPVKYLEFISECIIQVLQSK PGDFGADAQGAMNKALELFRKDMASNYKELGFQG Fuxreiter Mónika! Debreceni
A genetikai lelet értelmezése monogénes betegségekben
 A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói
 1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
A replikáció mechanizmusa
 Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
Szerkesztette: Vizkievicz András
 Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
Fehérjék A fehérjék - proteinek - az élő szervezetek számára a legfontosabb vegyületek. Az élet bármilyen megnyilvánulási formája fehérjékkel kapcsolatos. A sejtek szárazanyagának minimum 50 %-át adják.
FEHÉRJÉK A MÁGNESEKBEN. Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium. Alkímia Ma, Budapest,
 FEHÉRJÉK A MÁGNESEKBEN Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium Alkímia Ma, Budapest, 2013.02.28. I. FEHÉRJÉK: L-α aminosavakból felépülő lineáris polimerek α H 2 N CH COOH amino
FEHÉRJÉK A MÁGNESEKBEN Bodor Andrea ELTE, Szerkezeti Kémiai és Biológiai Laboratórium Alkímia Ma, Budapest, 2013.02.28. I. FEHÉRJÉK: L-α aminosavakból felépülő lineáris polimerek α H 2 N CH COOH amino
Áttekintő tartalomjegyzék
 4 Áttekintő tartalomjegyzék Új trendek a kromatográfiában (Gyémánt Gyöngyi, Kurtán Tibor, Lázár István) 5 Új technikák és alkalmazási területek a tömegspektrometriában (Gyémánt Gyöngyi, Kéki Sándor, Kuki
4 Áttekintő tartalomjegyzék Új trendek a kromatográfiában (Gyémánt Gyöngyi, Kurtán Tibor, Lázár István) 5 Új technikák és alkalmazási területek a tömegspektrometriában (Gyémánt Gyöngyi, Kéki Sándor, Kuki
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
Biológus MSc. Molekuláris biológiai alapismeretek
 Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
A Proteomika Szolgáltató Laboratóriumban elérhető szolgáltatások
 A Proteomika Szolgáltató Laboratóriumban elérhető szolgáltatások Dr. Csősz Éva Proteomika Szolgáltató Laboratórium műszerpark MALDI-TOF (ABSciex) PSZL ESI-4000 QTRAP (ABSciex) Kétdimenziós elektroforézis
A Proteomika Szolgáltató Laboratóriumban elérhető szolgáltatások Dr. Csősz Éva Proteomika Szolgáltató Laboratórium műszerpark MALDI-TOF (ABSciex) PSZL ESI-4000 QTRAP (ABSciex) Kétdimenziós elektroforézis
Aminosavak általános képlete NH 2. Csoportosítás: R oldallánc szerkezete alapján: Semleges. Esszenciális aminosavak
 Aminosavak 1 Aminosavak általános képlete N 2 soportosítás: oldallánc szerkezete alapján: Apoláris Poláris Bázikus Savas Semleges Esszenciális aminosavak 2 (apoláris) Glicin Név Gly 3 Alanin Ala 3 3 Valin
Aminosavak 1 Aminosavak általános képlete N 2 soportosítás: oldallánc szerkezete alapján: Apoláris Poláris Bázikus Savas Semleges Esszenciális aminosavak 2 (apoláris) Glicin Név Gly 3 Alanin Ala 3 3 Valin
Proteomika az élelmiszer-előállításában
 DEBRECENI EGYETEM Proteomika az élelmiszer-előállításában Czeglédi Levente Gulyás Gabriella Csősz Éva 2014., korrig 2015. TÁMOP-4.1.1.C-12/1/KONV-2012-0014 Élelmiszerbiztonság és gasztronómia vonatkozású
DEBRECENI EGYETEM Proteomika az élelmiszer-előállításában Czeglédi Levente Gulyás Gabriella Csősz Éva 2014., korrig 2015. TÁMOP-4.1.1.C-12/1/KONV-2012-0014 Élelmiszerbiztonság és gasztronómia vonatkozású
A fehérjék hierarchikus szerkezete. Szerkezeti hierarchia. A fehérjék építőkövei az aminosavak. Fehérjék felosztása
 Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
Fehérjék felosztása A fehérjék hierarchikus szerkezete Smeller László Semmelweis Egyetem Biofizikai és Sugárbiológiai Intézet Biológiai funkció alapján Enzimek (pl.: tripszin, citokróm-c ) Transzportfehérjék
A molekuláris biológia eszközei
 A molekuláris biológia eszközei I. Nukleinsavak az élő szervezetekben Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje DNS feladata: információ tárolása és a transzkripció
A molekuláris biológia eszközei I. Nukleinsavak az élő szervezetekben Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje DNS feladata: információ tárolása és a transzkripció
INFORMATIKA EMELT SZINT%
 Szövegszerkesztés, prezentáció, grafika, weblapkészítés 1. A fényképezés története Táblázatkezelés 2. Maradékos összeadás Adatbázis-kezelés 3. Érettségi Algoritmizálás, adatmodellezés 4. Fehérje Maximális
Szövegszerkesztés, prezentáció, grafika, weblapkészítés 1. A fényképezés története Táblázatkezelés 2. Maradékos összeadás Adatbázis-kezelés 3. Érettségi Algoritmizálás, adatmodellezés 4. Fehérje Maximális
Biotechnológiai gyógyszergyártás
 Biotechnológiai gyógyszergyártás Dr. Greiner István 2013. november 6. Biotechnológiai gyógyszergyártás Biotechnológiai gyógyszerek Előállításuk és analitikájuk Richter és a biotechnológia Debrecen A jövő
Biotechnológiai gyógyszergyártás Dr. Greiner István 2013. november 6. Biotechnológiai gyógyszergyártás Biotechnológiai gyógyszerek Előállításuk és analitikájuk Richter és a biotechnológia Debrecen A jövő
Gáspári Zoltán. Élő molekulák az élet molekulái
 Gáspári Zoltán Élő molekulák az élet molekulái Invokáció Kajtár Márton 1929-1991 www.eotvoskiado.hu Élő és élettelen? Élő és élettelen: a kemoton Élő kémiai rendszer, de nem élőlény (Gánti, 1975) Autokatalitikus
Gáspári Zoltán Élő molekulák az élet molekulái Invokáció Kajtár Márton 1929-1991 www.eotvoskiado.hu Élő és élettelen? Élő és élettelen: a kemoton Élő kémiai rendszer, de nem élőlény (Gánti, 1975) Autokatalitikus
A TÖMEGSPEKTROMETRIA ALAPJAI
 A TÖMEGSPEKTROMETRIA ALAPJAI web.inc.bme.hu/csonka/csg/oktat/tomegsp.doc alapján tömeg-töltés arány szerinti szétválasztás a legérzékenyebb módszerek közé tartozik (Nagyon kis anyagmennyiség kimutatására
A TÖMEGSPEKTROMETRIA ALAPJAI web.inc.bme.hu/csonka/csg/oktat/tomegsp.doc alapján tömeg-töltés arány szerinti szétválasztás a legérzékenyebb módszerek közé tartozik (Nagyon kis anyagmennyiség kimutatására
BIOLÓGIA ALAPJAI. Anyagcsere folyamatok 2. (Felépítő folyamatok)
") BIOLÓGIA ALAPJAI Anyagcsere folyamatok 2. (Felépítő folyamatok) A molekuláris biológiai alapjai DNS replikáció RNS transzkripció Fehérje szintézis (transzláció) (Az ábrák többsége Dr. Lénárd Gábor Biológia
BIOLÓGIA ALAPJAI Anyagcsere folyamatok 2. (Felépítő folyamatok) A molekuláris biológiai alapjai DNS replikáció RNS transzkripció Fehérje szintézis (transzláció) (Az ábrák többsége Dr. Lénárd Gábor Biológia
Biomolekuláris kölcsönhatások vizsgálata felületi plazmonrezonancia elvén működő Biacore keszülékkel
 Biomolekuláris kölcsönhatások vizsgálata felületi plazmonrezonancia elvén működő Biacore keszülékkel Biomolekuláris interakciók Fehérje-fehérje Fehérje-ligand Fehérje-DNS/RNS fehérje/ligand-lipid Alegység-kölcsönhatások,
Biomolekuláris kölcsönhatások vizsgálata felületi plazmonrezonancia elvén működő Biacore keszülékkel Biomolekuláris interakciók Fehérje-fehérje Fehérje-ligand Fehérje-DNS/RNS fehérje/ligand-lipid Alegység-kölcsönhatások,
Anyagszerkezet vizsgálati módszerek
 Kromatográfia Folyadékkromatográfia-tömegspektrometria Anyagszerkezet vizsgálati módszerek Pannon Egyetem Mérnöki Kar Anyagszerkezet vizsgálati módszerek Kromatográfia 1/ 25 Folyadékkromatográfia-tömegspektrometria
Kromatográfia Folyadékkromatográfia-tömegspektrometria Anyagszerkezet vizsgálati módszerek Pannon Egyetem Mérnöki Kar Anyagszerkezet vizsgálati módszerek Kromatográfia 1/ 25 Folyadékkromatográfia-tömegspektrometria
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
(11) Lajstromszám: E 007 952 (13) T2 EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA
 Lajstromszám: E 007 952 (13) T2 EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA") !HU00000792T2! (19) HU (11) Lajstromszám: E 007 92 (13) T2 MAGYAR KÖZTÁRSASÁG Magyar Szabadalmi Hivatal EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA (21) Magyar ügyszám: E 04 73892 (22) A bejelentés napja:
!HU00000792T2! (19) HU (11) Lajstromszám: E 007 92 (13) T2 MAGYAR KÖZTÁRSASÁG Magyar Szabadalmi Hivatal EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA (21) Magyar ügyszám: E 04 73892 (22) A bejelentés napja:
Enzimek. Enzimek! IUBMB: szisztematikus nevek. Enzimek jellemzése! acetilkolin-észteráz! legalább 10 nagyságrend gyorsulás. szubsztrát-specificitás
 Enzimek acetilkolin-észteráz! Enzimek! [s -1 ] enzim víz carbonic anhydrase 6x10 5 10-9 karbonikus anhidráz acetylcholine esterase 2x10 4 8x10-10 acetilkolin észteráz staphylococcal nuclease 10 2 2x10-14
Enzimek acetilkolin-észteráz! Enzimek! [s -1 ] enzim víz carbonic anhydrase 6x10 5 10-9 karbonikus anhidráz acetylcholine esterase 2x10 4 8x10-10 acetilkolin észteráz staphylococcal nuclease 10 2 2x10-14
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
A fehérjéket felépítő húsz standard aminosav Fehérjék szerkezetének kialakulása
 A fehérjéket felépítő húsz standard aminosav Fehérjék szerkezetének kialakulása Osváth Szabolcs Semmelweis Egyetem szabolcs.osvath@eok.sote.hu reakció t 1/2 25 ºC-on t 1/2 100 ºC-on DNS hidrolízis Biopolimerek
A fehérjéket felépítő húsz standard aminosav Fehérjék szerkezetének kialakulása Osváth Szabolcs Semmelweis Egyetem szabolcs.osvath@eok.sote.hu reakció t 1/2 25 ºC-on t 1/2 100 ºC-on DNS hidrolízis Biopolimerek
Gyógyszerrezisztenciát okozó fehérjék vizsgálata
 Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
A fehérjéket felépítő húsz standard aminosav
 Fehérjék szerkezetének kialakulása Osváth Szabolcs Semmelweis Egyetem szabolcs.osvath@eok.sote.hu Biopolimerek reakció t 1/2 25 ºC-on t 1/2 100 ºC-on egy polimerben lévő kötések tipikus száma hányfajta
Fehérjék szerkezetének kialakulása Osváth Szabolcs Semmelweis Egyetem szabolcs.osvath@eok.sote.hu Biopolimerek reakció t 1/2 25 ºC-on t 1/2 100 ºC-on egy polimerben lévő kötések tipikus száma hányfajta
Biokémiai kutatások ma
 Nyitray László Biokémiai Tanszék Hb Biokémiai kutatások ma Makromolekulák szerkezet-funkció kutatása Molekuláris biológia minden szinten Redukcionista molekuláris biológia vs. holisztikus rendszerbiológia
Nyitray László Biokémiai Tanszék Hb Biokémiai kutatások ma Makromolekulák szerkezet-funkció kutatása Molekuláris biológia minden szinten Redukcionista molekuláris biológia vs. holisztikus rendszerbiológia
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Tematika. Korszerű tömegspektrometria a. Ionforrás. Gyors atom bombázás. Szabó Pál MTA Kémiai Kutatóközpont. Cél: Töltött részecskék előállítása
 Tematika Korszerű tömegspektrometria a biokémi miában Szabó Pál MTA Kémiai Kutatóközpont Bevezetés: ionizációs technikák és analizátorok összehasonlítása a biomolekulák szemszögéből Mikromennyiségek mintaelőkészítése
Tematika Korszerű tömegspektrometria a biokémi miában Szabó Pál MTA Kémiai Kutatóközpont Bevezetés: ionizációs technikák és analizátorok összehasonlítása a biomolekulák szemszögéből Mikromennyiségek mintaelőkészítése
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
Genomika. Mutációk (SNP-k) és vizsgálatuk egyszerű módszerekkel. DNS szekvenálási eljárások. DNS ujjlenyomat (VNTR)
 és vizsgálatuk egyszerű módszerekkel. DNS szekvenálási eljárások. DNS ujjlenyomat (VNTR)") Genomika (A genom, génállomány vizsgálata) Mutációk (SNP-k) és vizsgálatuk egyszerű módszerekkel DNS szekvenálási eljárások DNS ujjlenyomat (VNTR) DNS chipek statikus és dinamikus információk vizsgálata
Genomika (A genom, génállomány vizsgálata) Mutációk (SNP-k) és vizsgálatuk egyszerű módszerekkel DNS szekvenálási eljárások DNS ujjlenyomat (VNTR) DNS chipek statikus és dinamikus információk vizsgálata
2007/11/05 Molekuláris biológia előadások - Putnoky 1-1
 1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Klónozás: tökéletesen egyforma szervezetek csoportjának előállítása, vagyis több genetikailag azonos egyed létrehozása.
 Növények klónozása Klónozás Klónozás: tökéletesen egyforma szervezetek csoportjának előállítása, vagyis több genetikailag azonos egyed létrehozása. Görög szó: klon, jelentése: gally, hajtás, vessző. Ami
Növények klónozása Klónozás Klónozás: tökéletesen egyforma szervezetek csoportjának előállítása, vagyis több genetikailag azonos egyed létrehozása. Görög szó: klon, jelentése: gally, hajtás, vessző. Ami
Az aminosav anyagcsere orvosi vonatkozásai Csősz Éva
 Az aminosav anyagcsere orvosi vonatkozásai Csősz Éva E-mail: cseva@med.unideb.hu Általános reakciók az aminosav anyagcserében 1. Nitrogén eltávolítás: transzaminálás dezaminálás: oxidatív nem oxidatív
Az aminosav anyagcsere orvosi vonatkozásai Csősz Éva E-mail: cseva@med.unideb.hu Általános reakciók az aminosav anyagcserében 1. Nitrogén eltávolítás: transzaminálás dezaminálás: oxidatív nem oxidatív