Szívelektrofiziológiai alapjelenségek 2. Dr. Tóth András 2018
|
|
|
- Zsombor Orbán
- 5 évvel ezelőtt
- Látták:
Átírás
1 Szívelektrofiziológiai alapjelenségek 2. Dr. Tóth András 2018
2 Témák Ioncsatornák Helyi és akciós potenciálok A stimulus intra- és extracelluláris terjedése
3 4 Ioncsatornák
4 4.1 Alaptulajdonságok
5 Ioncsatorna Protein (vagy protein komplex), mely a sejt, vagy organellum membránban pórusként működik és lehetővé teszi (facilitálja) egyes ionok szelektív transzportját. Fiziológiás aktiválódása következtében transzmembrán elektromos áram generálódik, amely egyaránt lehet befelé mutató ( inward ) vagy kifelé mutató ( outward ). Az ioncsatornák (igen gyakran genetikus okok következtében kialakuló) kiesése, vagy hibás működése súlyos betegségeket okozhat. Nagy számban azonosítottak humán betegségeket, melyek egyes ioncsatornák nem megfelelő működésének következtében alakulnak ki ezek az ioncsatorna betegségek ( channelopathies ).
6 Ioncsatornák alaptulajdonságai 1. Tézis: A sejtek normális működéséhez számos ion precízen szabályozott, szinkronizált transzportja létfontosságú. Ioncsatornák: megtalálhatók a sejtplazma membránjában és számos organellumban. Belsejükben vezető csatorna található, amely az ionok mozgását azok méretétől és/vagy töltésétől függően teszi, vagy nem teszi lehetővé (szelektív permeabilitás). Egy adott ioncsatorna működése lehet szabályozott (gated), vagy nem szabályozott (nongated). Egyéb iontranszport proteinektől az ioncsatornákat két lényeges tulajdonságuk különbözteti meg: (1) A relatív transzportsebesség ( transport rate ) nagyon nagy (gyakran > 10 6 ion/s). (2) Az ionok elektrokémiai gradiensük irányában haladnak át a csatornán (amely a koncentrációgradiens és membránpotenciál függvénye). Metabolikus energiára (ATP, aktív transzport/kotranszport) a "downhill", iontranszporthoz nincs szükség. Mivel képesek az ionok ki- vagy beáramlását szabályozni, az ioncsatornák direkt vagy sok esetben indirekt(!!!) célpontjai különböző gyógyszereknek.
7 Ioncsatornák alaptulajdonságai 2. Tézis: Egyes csatornák működése szabályozott ( gated channels), másoké lényegében nem szabályozott ( nongated leakage channels). Gated ion csatorna specifikus stimulus hatására nyílik vagy (esetenként) záródik. Ez a specifikus stimulus lehet: (1) membránpotenciál (feszültség) változás (voltage gated) (2) specifikus ligand vagy szignálmolekula kötődés ( ligand gated csatornák) (hormon, neurotranszmitter, lokális hormon); (3) mechanikai hatás (deformáció, nyomás, stretch), (4) fényenergia (foton). Mivel a gated ioncsatorna szabályozott csak akkor nyílik, amikor az adekvát trigger szignál beérkezik. Záródása viszont egyaránt lehet spontán, vagy szabályozott. Nongated (leakage) ion csatorna - mindig nyitott (vagy szivárog) és lehetővé teszi egy, vagy több ion áthaladását azok elektrokémiai grádiensének megfelelő irányban. A nongated csatorna nem szabályozott (azaz a transzport-fluxus az elektrokémiai potenciálon kívül csak a hőmérséklettől függ).


8 Egy ioncsatorna elvi struktúrája 1 csatorna domain-ek (tipikusan csatornánként négy) 2 külső vesztibula 3 a selectivity filter 4 a selectivity filter átmérője 5 foszforilációs hely 6 sejt membrán A legtipikusabb csatorna-pórus legszűkebb pontján csak egy-két atomnyi széles és szelektív a rá specifikus ionra (Na +, H +, K + ). Ugyanakkor jónéhány ioncsatorna kevésbé szelektív és több azonos előjelű töltéssel rendelkező ion számára is permeábilis lehet (azaz kation vagy anion szelektív).
9 Biológiai szerep Az ioncsatornák számos, kritikus jelentőségű sejtszintű funkcióval rendelkeznek, pl. kémiai szignálok, transzcelluláris transzport, ph szabályozás, sejttérfogat szabályozása. Kiemelkedő jelentőségük van az idegrendszer működése szempontjából (pl. idegi impulzusok kialakulása/terjedése, traszmitter-aktivált csatornák a szinapszisokban. (A legtöbb állati méreg és toxin pók, skorpió, kígyó, tengeri csiga, méh/darázs, stb. idegméreg és ioncsatornák konduktanciáját és/vagy kinetikáját modulálja.) Ezenkívül számos élettani folyamatban kulcsszereplők: pl. szív-, váz- és simaizom excitáció-kontrakció csatolása, tápanyagok és ionok epitéliális transzportja, immunrendszer aktiválása, a pankreász béta-sejtek inzulin felszabadítása, stb. Ezért nem meglepő, hogy számos ioncsatorna a gyógyszerkutatások kiemelkedő célpontja.
10 Diverzitás Egyetlen élő sejtben több, mint 300 aktív ioncsatorna típus található. Osztályozásuk többféleképpen történhet: a gating mechanizmus szerint, az általuk a póruson szelektiven átengedett ion(ok) szerint, vagy az alkotó csatorna proteinek struktúrája és/vagy lokalizációja szerint. További heterogenitást jelent, ha az adott áramot több, eltérő szerkezettel rendelkező csatorna együttes aktiválódása alakítja ki. A csatornát alkotó, vagy szabályozó alegységek hiánya vagy mutációja funkcióvesztésben nyilvánulhat meg, amely számos betegség hátterében megtalálható.
11 Kérdés: Mi az elvi különbség a membránreceptor és az ioncsatorna között?
12 Kérdés: Van-e olyan membránreceptor, amely ugyanakkor ioncsatorna is?
13 4.2 Vizsgálati technikák

14 Bert Sakmann Nobel díjas a patch clamp módszer kidolgozója

")
15 A patch clamp technika alapelve Szükséges hozzá: Pipetta + Elektród Speciális pipettaoldat Gigaseal (R > 1GOhm) Feszültségparancs Árammérés
16 A patch clamp mérés lépései
17 A patch clamp technika fontosabb konfigurációi Vizsgálható: Egyetlen csatorna árama Egy csatornatípus vagy - csatorna csoport árama Speciális pipettaoldat IC miliő módosítása

18 Automatizált patch clamp mérések elve
19 4.3 Szabályozási alapelvek
20 Egyszerű, kétállapotú ioncsatorna a background csatornák spontán oszcillálnak a nyitott és zárt állapotok között
21 Komplex, többállapotú ioncsatorna állapotai
22 4.4 Biofizikai tulajdonságok

23 Single channel áram és meghatározása 1. Egyetlen csatorna vagy nyitott,vagy zárt állapotban van. 2. A csatorna nyitott állapota rövid ideig tart a makroszkopikus áramhoz viszonyítva. 3. A nyitott állapot időtartama és latenciája nagyon változó. Előfordul, hogy egy csatorna egyáltalán nem nyílik ki. 4. Egyetlen csatorna nyitási valószínűségeloszlás függvénye hasonló a makroszkópikus áramhoz 5. A csatornák többször is kinyílhatnak, ha nincs inaktiváció
24 Sok feszültségfüggő Na + csatorna, illetve feszültségfüggő késleltetett K + csatorna integrált válasza feszültségugrásra A Na + csatorna spontán inaktiválódik, a K + csatorna nem
25 Az átlagos nyitvatartási idő meghatározása A csatorna modulációja az átlagos nyitvatartási idő változásában nyilvánul meg
26 Inward, illetve outward egyenirányító csatornák vezetési (feszültség-áram) karakterisztikája
27 Két csatorna csoport (Na + & K + ), illetve azok kombinációjának feszültség-áram karakterisztikája A K + áram lineáris, azaz nincsen feszültség-inaktiváció A Na + áram nemlineáris a feszültség-inaktiváció miatt
28 Két ioncsatorna típus áram-feszültség összefüggése pitvarból izolált sejtekben meghatározva

29 Az aktivációs/inaktivációs kinetika lépései A Kv4 típusú K + csatornák lassú inaktivációjának működő strukturális modellje
30 4.5 Ioncsatornák csoportosítása
31 Csoportosítás a kapuzó szignál szerint 1 Feszültségfüggő ioncsatornák Nyitásuk (és sokszor záródásuk is) a membránpotenciál (feszültség-gradiens) változásainak függvénye, azaz spontán oszcillációjuk egyensúlyi állapota (a nyitott/ zárt állapot valószínűsége) feszültségfüggő Főbb típusok - Feszültségfüggő Na + csatornák (Nav) - Feszültségfüggő K + csatornák (Kv) - Feszültségfüggő Ca + csatornák (Cav) - Feszültségfüggő H + csatornák (Hv) - Hyperpolarizáció-aktivált csatornák - Tranziens receptor potenciál (TRP) csatornák Funkciójuk Elsősorban ingerelhető sejtek (izomsejtek, neuronok, mirigysejtek) ingerelhetőségéért és ingerületvezetéséért felelősek, létfontosságúak az idegi impulzusok terjedésében is. Az adott körülményektől függően egyaránt hozzájárulhatnak a depolarizáció, repolarizáció, illetve hiperpolarizáció kialakulásához.
32 Csoportosítás a kapuzó szignál szerint 2 Ligand-függő ioncsatornák Másnéven ionotróp receptorok. Nyitásuk egy speciális ligand valamelyik csatornaproteinhez kötődésének függvénye: pl. neurotranszmitterfüggő, G-protein függő, modulált (kovalens modifikáció foszforiláció) csatornák. A ligand kötése a csatornaprotein konformációváltozásán keresztül facilitálja az ionok beáramlását Ezek a csatornák felelősek pl. az impulzusok inicializálásáért, azaz ingerelhető sejtek (ideg, izom, mirigy) kezdeti depolarizációjáért. A körülményektől függően depolarizáció, repolarizáció, vagy hiperpolarizáció kialakításában részt vehetnek. Példák: Kation-permeábilis nicotinerg acetylcholine receptor, ionotróp glutamát-függő receptorok, ATP-függő P2X receptorok, anion-permeábilis GABA-aktivált receptor. Egyesek a 2-nd messengerek által aktivált ioncsatornákat is ide szokták sorolni, bár a ligand és a 2-nd messenger nem azonos kategória.
33 Csoportosítás a kapuzó szignál szerint 3 Egyéb kapuzási mechanizmusok Ide tartozik a membrán belső oldaláról történő aktiválás/inaktiválás pl. második messengereken keresztül. Egyes, a csatornát közvetlenül aktiváló ionok is második messengernek tekinthetők (pl. befelé egyenirányító K + csatornák, Ca 2+ -függő K + csatornák, két-porúsú K + csatornák A modulált csatornák, lehetnek feszültségfüggőek, de ezenkívül kovalens modifikáció (pl. foszforiláció) is modulálja a nyitott/zárt állapotok valószínűségét A G-protein függő csatornák nyitott és zárt állapotainak valószínűségét a (pl. receptor aktiválás során) aktivált G-protein kötődése modulálja (pl. muszkarin típusú Ach)
 hatására történik. Fényre aktiválódó ioncsatornák (2) Fényérzékeny sejtekben nyitásuk fotonabszorbció következtében történik.")
 ciklikus nukleotid (camp; cgmp) érzékeny (CNG) csatornák, illetve 2) hiperpolarizáció által aktivált,")
34 Csoportosítás a kapuzó szignál szerint 4 Mechanoszenzitív ioncsatornák (1) 1 Nyitásuk (és sokszor záródásuk is) a csatornára ható mechanikai stimulus (nyomás, stretch, vibráció) hatására történik. Fényre aktiválódó ioncsatornák (2) Fényérzékeny sejtekben nyitásuk fotonabszorbció következtében történik. 2 Hőérzékeny csatornák Egyes TRP receptorok (TRPV1, TRPM8) vagy magas (pl. kapszaicin receptor), vagy alacsony hőmérsékletre nyílnak. Ciklikus nukleotid-függő csatornák Két család: 1) ciklikus nukleotid (camp; cgmp) érzékeny (CNG) csatornák, illetve 2) hiperpolarizáció által aktivált, ciklikus nukleotid-függő (HCN) csatornák.
35 Csoportosítás a transzportált ion szerint 1 Nátrium csatornák Feszültségfüggő nátrium csatornák (Nav) Epitheliális nátrium csatornák (ENaC) Kálium csatornák Feszültségfüggő kálium csatornák (Kv) Kalcium-aktivált kálium csatornáks (BKCa, SK, stb.) Befelé-egyenirányító (inward-rectifier) kálium csatornák (Kir) Two-pore-domain kálium csatornák ( leak channels) Kalcium csatornák Feszültségfüggő kalcium csatornák (CaV)
36 Csoportosítás a transzportált ion szerint 2 Proton csatornák Feszültségfüggő proton csatornák (Hv), erős ph függéssel és nagyon kicsi konduktanciával Klorid csatornák Kevéssé ismert anioncsatorna szuper-család legalább 13 taggal. (pl. ClC, CLIC, Bestrophin, CFTR). Kevéssé, vagy egyáltalán nem szelektív kisméretű anionokra. Mivel a klorid a legjellemzőbb transzportált anion, klorid csatornáknak nevezik. Nem-szelektív kation csatornák Többféle kationra permeábilisak ( főleg Na +, K + and Ca 2+ ) A TRP (transient receptor potential) csatornacsalád több tagja is ide tartozik
37 További csoportosítások Kevéssé általános tulajdonságok, pl. a pórusok száma, illetve a válasz időtartama alapján történik. Kétpórusú (two pore) csatornák Kb. 2 tagú kation-szelektív ioncsatorna család, amely két Kv-szerű (6-transzmembrán domain) egységet tartalmaz. A membránban valószínűleg dimerként működik. Tranziens receptor potenciállal rendelkező (TRP) csatornák Legalább 28 tagú család, rendkívül diverz aktivációs tulajdonságokkal Egyesek állandóan nyitottak, mások feszültség-, [Ca 2+ ]-, ph-, redox állapot-, osmolaritás- vagy stretch-függőek. Szelektivitásuk is változó, Ca 2+ -szelektív, illetve kevésbé szelektív kation csatornák. 6 alcsalád: kanonikus (TRPC), vanilloid (TRPV), melastatin (TRPM), polycystin (TRPP), mucolipin (TRPML), és ankyrin (TRPA) transzmembrán proteinnel.

38 Példa: a K + csatornák népes családja
39 Kérdések: Hogyan lehetséges, hogy a Na + átmehet egy ioncsatornán, a K + pedig nem? Hogyan lehetséges, hogy a K + átmehet egy ioncsatornán, a Na + pedig nem? Segítség: Ion Atomsugár (Angström) Hidratációs energia (kcal/mol ) Na K
40 4.6 Szerkezet
41 Néhány ioncsatorna szupercsalád jellemző képviselője
42 Az S 4 hélixek a feszültségfüggő csatornák feszültségszenzorai jelentős aminosav homológia jellemző

43 A feszültségfüggő Na + csatorna 2D modelje
44 A feszültségfüggő Ca 2+ csatorna 2D modelje
 Az újabb paddle modell: A töltött S4 és S3 szegmensek alkotta hajtű a csatornán kívül helyezkedik el. A csatorna aktivációja ezek elfordulásával jön létre.")
45 Feszültségfüggő csatornák aktivációs hipotézisei a) A konvencionális gating modell: A töltött S4 szegmens (feszültség-szenzor) transzmembrán mozgást végez. b) Az újabb paddle modell: A töltött S4 és S3 szegmensek alkotta hajtű a csatornán kívül helyezkedik el. A csatorna aktivációja ezek elfordulásával jön létre. c) A transzporter modell: Ez az újabb modell azon alapul, hogy a feszültség-szenzor (S4) töltései nem végeznek transzmembrán mozgást, hanem elfordulnak hossztengelyük körül, ezáltal a gating töltések extracelluláris helyzetből intracelluláris pozícióba kerülnek.
 és négy homológ domain-ból állnak (I-IV), melyekben 6 transzmembrán szegmens (S1- S6) található.")
46 Feszültségfüggő Na + csatornák (Nav) Legalább 9 tagú csatornacsalád, melynek nagy szerepe van az akciós potenciál kialakításában és terjedésében. A pórusképző α alegységek rendkívül nagyok (kb. 4,000 amino csoport) és négy homológ domain-ból állnak (I-IV), melyekben 6 transzmembrán szegmens (S1- S6) található. Ezek az α alegységek szabályozó szereppel bíró β alegységekkel egészülnek ki, melyek egyetlen transzmembrán egységet alkotnak.

47 Feszültségfüggő Ca 2+ csatornák (Cav) A Cav csatornacsalád legalább 10 tagból áll. Fontos szerepük van az izmok excitáció-kontrakció csatolásának biztosításában, illetve neuronok transzmitter-felszabadulásának szabályozásában Az α 1 alegység szekezete nagyon hasonlít a Na + csatorna α alegységéhez. További szabályozó alegységek is kapcsolódnak hozzá (α 2, δ, β, γ).

48 Feszültségfüggő K + csatornák (Kv) Közel 40 tagú ioncsatorna család, amely 12 alcsaládból áll. Ezek a csatornák főleg a membrán akciós potenciált követő repolarizációjáért felelősek. Az α alegységek (a Na + csatornákhoz hasonlóan) 6 transzmembrán szegmensből állnak (6TM) és ugyanúgy tetramert alkotva hoznak létre működő csatornát a membránban.

49 A feszültség-függő K + csatorna funkcionális modellje

50 "Birth of an Idea", m x 0.90 m x 0.90 m Steel, glass, wood Sculpture by Julian Voss- Andreae based on potassium channel Photo by Dan Kvitka Sculpture commissioned and owned by Roderick MacKinnon Please use only with link to
51 4.7 Struktúra-funkció kapcsolat
52 A feszültségérzékelő S 4 hélix funkcionális modelje Összesen 6 töltésnek kell elmozdulnia a membránban a csatorna nyílásához

53 A Na + csatorna felülnézeti képe; látható, hogy a négy domén ugyanazon hélixei veszik körbe a centrális ioncsatornát

54 A szívben található néhány fontosabb ioncsatorna
55 A szív akciós potenciálja és a legfontosabb ionáramai, ioncsatornái közötti kapcsolat
56 Kérdés: Melyek az ioncsatornák legfontosabb tulajdonságai?
57 Válasz: Integráns membrán proteinek Hidrofil pórust alkotnak a bilipid membránban Szelektív permeabilitással rendelkeznek Single channel (ON-OFF) konduktanciát mutatnak Rectification - egyenirányító működést mutat(hat)nak Gating - oszcilláció vezető és nem vezető állapotok között Reguláció - azaz feszültség-, ligand-, vagy kovalens módosulásfüggő vezetés Farmakológiai modulálhatóság!!! Klinikailag fontos hatóanyagok célpontjai!!!
58 5 Lokális és akciós potenciálok
59 5.1 Lokális válasz
60 Kérdés: Mi a különbség az elektrokémiai potenciál és a membránpotenciál között?
")
61 A lokális (küszöb alatti) válasz
62 Kérdés: Melyek a lokális válasz legfontosabb jellemzői?

63 Időbeli szummáció

64 Térbeli szummáció
65 Kérdés: Melyek a lokális válasz speciális esetei?
66 5.2 Akciós potenciál

67 Az akciós potenciál fázisai

68 Különböző típusú akciós potenciálok
69 Kérdés: Miben különbözik a lokális válasz az akciós potenciáltól?
70 5.3 Akciós potenciálok a szívben
71 Az emlősszívben mérhető ionkoncentrációk

72 A gyors és lassú válasz szívben
73 Az akciós potenciál alakjának regionális variabilitása a szívben
74 Kérdés: Mi a magyarázata a különböző szívsejtekben mért akciós potenciálok jelentősen eltérő kinetikájának?
75 Ionáramok! Gyors Nátrium Funny Delayed rectifier Kálcium I L T+L 0 0 Tranziens outward Background Nátrium Inward rectifier Az eltérő akciós potenciálok magyarázata
76 Kérdés: Hogyan lehetne megváltoztatni az akciós potenciál alakját?

77 Tetrodotoxin hatása a gyors válaszra
78 Kérdés: Mi a tetrodotoxin hatása a gyors válaszra a szívizomsejtekben?
79 6 Az ingerület terjedése
80 6.1 Az ingerületvezetés alapelvei
81 Kérdés: Miért éri meg, sejtjeink nyugalmi potenciálját fenntartani, ha az ára jelentős mértékű energiafelhasználás (ATP)?
")
82 A különböző távolságban regisztrált (mért) potenciálváltozások

83 A változás regisztrált (mért) maximuma a távolság függvényében
84 Modell RC-körre számított potenciálváltozásai

85 Az axonmembrán elektromos modellje

86 R m R i C A membrán mért időkonstansa
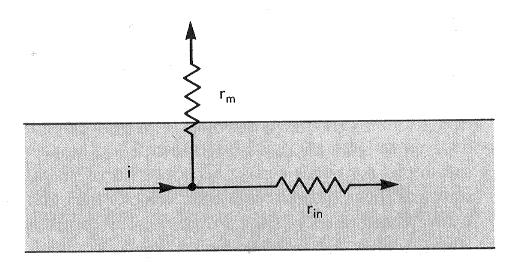
87 Modell feszültségosztó (rezisztencia-hányados)

88 R R m i A membrán mért térkonstansa
89 A lokális (küszöb alatti) válasz terjedésének modellje

90 A lokális válasz terjedésének modellje
91 Kérdés: Miért éppen az axondombon alakul ki a posztszinaptikus akciós potenciál?

92 Az akciós potenciál terjedési modellje velőhüvely nélküli rostban

93 Az akciós potenciál szaltatórikus terjedése

94 Akciós potenciál myelinhüvellyel rendelkező és myelin-hüvely nélküli idegrostban mért terjedési sebessége
95 Kérdés: Milyen tényezőktől függ a myelinhüvelyes rostok vezetési sebessége? És a myelinhüvely nélkülieké?
96 Kérdés: Mi a magyarázata a myelinhüvelyes rostok jóval nagyobb vezetési sebességének?
97 6.2 Ingerületvezetés szívizomban
98 Kérdés: Mit értünk azon a kifejezésen, hogy a szívizom funkcionális szincitiumot képez?
")
99 MW 1500 Ca 2+ ph E m Az elektromos szinapszis (gap junction) szerkezete
100 Kérdés: Hol találhatók a szervezetben elektromos szinapszisok?
101 Kérdés: Melyek a legfontosabb funkcionális eltérések az elektromos és kémiai szinapszisok között?
102 A szívizomsejt elektromos modellje
103 A mikroszkópikus ingerületterjedés számítógépes szimulációja
104 Kérdés: Mitől függ, hogy milyen irányban és milyen pillanatnyi sebességgel terjed az AP a háromdimenziós szívizomban?
105 Kérdés: Mi a magyarázata annak, hogy az AV csomón nagyon lassan halad át az ingerület?
106 Kérdés: Van-e gyors, illetve lassú akciós potenciál terjedés? Miért?
107 Bónusz!!! A gap junction rendszer jelentősége a szívizom fiziológiás ingerületvezetésében

108 Impulzusterjedés szubcelluláris szinten
109 Intra- és intercelluláris aktivációs késleltetések közötti különbségek - egyetlen sejt szélességű sejthálózat
110 Intra- és intercelluláris aktivációs késleltetések közötti különbségek - néhány sejt szélességű sejthálózat

111 Impulzusvezetés (izokron vonalak) fiziológiás gap junction csatolás esetén (homogén AP-populáció)

112 Impulzusvezetés (izokron vonalak) a gap junction hálózat súlyos szétkapcsolása esetén (heterogén AP-populáció)
 is csökkenhet (36.7 cm/s helyett 0.31 cm/s)")
113 Súlyos gap junction szétkapcsolás esetén a vezetési sebesség KÉT nagyságrenddel (!!!) is csökkenhet (36.7 cm/s helyett 0.31 cm/s)

114 Normális gap junction csatolás esetén viszonylag egyenletesen elhelyezkedő izokron vonalak és homogén AP-populáció

115 Kritikus gap junction szétkapcsolódás esetén az akciós potenciálok klasztereket alkotnak, köztük a késleltetés jelentős

116 Az egyes klasztereket alkotó sejtek elhelyezkedése kritikus mértékű szétkapcsolás esetén- az ingerület helyenkénti visszafordulása reentry kialakulásához vezethet
117 VÉGE
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András
 Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Ioncsatornák 1 Transzmembrán transzport 1 A membrántranszport-folyamatok típusai 2 J: diffúziós
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Ioncsatornák 1 Transzmembrán transzport 1 A membrántranszport-folyamatok típusai 2 J: diffúziós
Elektrofiziológiai alapjelenségek. Dr. Tóth András
 Elektrofiziológiai alapjelenségek Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Lokális és akciós potenciálok Az ingerület terjedése
Elektrofiziológiai alapjelenségek Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Lokális és akciós potenciálok Az ingerület terjedése
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András
 Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika
 Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Szívelektrofiziológiai alapjelenségek. Dr. Tóth András 2018
 Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Érzékszervi receptorok
 Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
Membránpotenciál, akciós potenciál
 A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
Az akciós potenciál (AP) 2.rész. Szentandrássy Norbert
 2.rész. Szentandrássy Norbert") Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Transzportfolyamatok a biológiai rendszerekben
 A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
IONCSATORNÁK. Osztályozás töltéshordozók szerint: pozitív töltésű ion: Na+, K+, Ca2+ negatív töltésű ion: Cl-, HCO3-
 Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák
 A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
A szívizomsejt ioncsatornái és azok működése
 A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
A sejtek közötti kommunikáció módjai és mechanizmusa. kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok
 A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György
 Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György Nyugalmi membránpotenciál: TK. 284-285. Akciós potenciál: TK. 294-301. Elektromos ingerelhetőség:
Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György Nyugalmi membránpotenciál: TK. 284-285. Akciós potenciál: TK. 294-301. Elektromos ingerelhetőség:
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
- Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál. ellenállás. kondenzátor
 - Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál ellenállás kondenzátor Alap struktúra: 4x6TM S5-S6: vizes pórus, szelektivitás S1-S4: feszültség-függés gating: általában
- Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál ellenállás kondenzátor Alap struktúra: 4x6TM S5-S6: vizes pórus, szelektivitás S1-S4: feszültség-függés gating: általában
Membránpotenciál. Nyugalmi membránpotenciál. Akciós potenciál
 Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Az ingerületi folyamat sejtélettani alapjai
 Az ingerületi folyamat sejtélettani alapjai Dr. Oláh Attila DEOEC Élettani Intézet 2011.09.15. Alapvetések I. Mi az a membránpotenciál? Az intakt sejtmembrán elektromosan szigetel -> a rajta keresztül
Az ingerületi folyamat sejtélettani alapjai Dr. Oláh Attila DEOEC Élettani Intézet 2011.09.15. Alapvetések I. Mi az a membránpotenciál? Az intakt sejtmembrán elektromosan szigetel -> a rajta keresztül
CELLULÁRIS SZÍV- ELEKTROFIZIOLÓGIAI MÉRÉSI TECHNIKÁK. Dr. Virág László
 CELLULÁRIS SZÍV- ELEKTROFIZIOLÓGIAI MÉRÉSI TECHNIKÁK Dr. Virág László Intracelluláris mikroelektród technika Voltage clamp technika Patch clamp technika Membrane potentials and excitation of impaled single
CELLULÁRIS SZÍV- ELEKTROFIZIOLÓGIAI MÉRÉSI TECHNIKÁK Dr. Virág László Intracelluláris mikroelektród technika Voltage clamp technika Patch clamp technika Membrane potentials and excitation of impaled single
In vitro elektrofiziológiai technikák Mike Árpád
 In vitro elektrofiziológiai technikák Mike Árpád 2011-05-20 1. A sejt szintű elektrofiziológia alapjai: Története Technikák Ionáramok szelektivitása, iránya, nagysága, hatása a membránpotenciálra 2. FAQ
In vitro elektrofiziológiai technikák Mike Árpád 2011-05-20 1. A sejt szintű elektrofiziológia alapjai: Története Technikák Ionáramok szelektivitása, iránya, nagysága, hatása a membránpotenciálra 2. FAQ
A sejtek közöti kommunikáció formái. BsC II. Sejtélettani alapok Dr. Fodor János
 A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
1. Mi jellemző a connexin fehérjékre?
 Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
Ioncsatorna szerkezetek
 Jellegzetes Ioncsatorna szerkezetek Ördög Balázs Farmakológiai és Farmakoterápiai Intézet Kapuzás Feszültség szabályozott Voltage-gated Fesz. szab. Na +, +, Ca 2+ 2+,, K + + csatornák channels Transiens
Jellegzetes Ioncsatorna szerkezetek Ördög Balázs Farmakológiai és Farmakoterápiai Intézet Kapuzás Feszültség szabályozott Voltage-gated Fesz. szab. Na +, +, Ca 2+ 2+,, K + + csatornák channels Transiens
Membránszerkezet, Membránpotenciál, Akciós potenciál. Biofizika szeminárium
 Membránszerkezet, Membránpotenciál, Akciós potenciál Biofizika szeminárium 2013. 09. 09. Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens
Membránszerkezet, Membránpotenciál, Akciós potenciál Biofizika szeminárium 2013. 09. 09. Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
4. Egy szarkomer sematikus rajza látható az alanti ábrán. Aktív kontrakció esetén mely távolságok csökkenése lesz észlelhető? (3)
") Budapesti Műszaki és Gazdaságtudományi Egyetem, Budapest, 2009. jan. 6. Villamosmérnöki és Informatikai Kar Semmelweis Egyetem Budapest Egészségügyi Mérnök Mesterképzés Felvételi kérdések orvosi élettanból
Budapesti Műszaki és Gazdaságtudományi Egyetem, Budapest, 2009. jan. 6. Villamosmérnöki és Informatikai Kar Semmelweis Egyetem Budapest Egészségügyi Mérnök Mesterképzés Felvételi kérdések orvosi élettanból
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az idegsejt elektrokémiai és
 Mottó: Mert az angyal a részletekben lakik. Petri György: Mosoly Az idegsejt elektrokémiai és fiziológiai működésének alapjai. ELTE, 2006. október 6. Tartalom Az idegsejt felépítése Az idegi elektromosság
Mottó: Mert az angyal a részletekben lakik. Petri György: Mosoly Az idegsejt elektrokémiai és fiziológiai működésének alapjai. ELTE, 2006. október 6. Tartalom Az idegsejt felépítése Az idegi elektromosság
Membránszerkezet Nyugalmi membránpotenciál
 Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Az idegi működés strukturális és sejtes alapjai
 Az idegi működés strukturális és sejtes alapjai Élettani és Neurobiológiai Tanszék MTA-ELTE NAP B Idegi Sejtbiológiai Kutatócsoport Schlett Katalin a kurzus anyaga elérhető: http://physiology.elte.hu/agykutatas.html
Az idegi működés strukturális és sejtes alapjai Élettani és Neurobiológiai Tanszék MTA-ELTE NAP B Idegi Sejtbiológiai Kutatócsoport Schlett Katalin a kurzus anyaga elérhető: http://physiology.elte.hu/agykutatas.html
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.
. Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.") Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV
 KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus
 Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Sejtek membránpotenciálja
 Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt
 az információ kémiailag kódolt") Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
klorid ioncsatorna az ABC (ATP Binding Casette) fehérjecsaládba tartozik, amelyek általánosságban részt vesznek a gyógyszerek olyan alapvetı
 fehérjecsaládba tartozik, amelyek általánosságban részt vesznek a gyógyszerek olyan alapvetı") Szegedi Tudományegyetem Farmakológiai és Farmakoterápiai Intézet Igazgató: Prof. Dr. Varró András 6720 Szeged, Dóm tér 12., Tel.: (62) 545-682 Fax: (62) 545-680 e-mail: varro.andras@med.u-szeged.hu Opponensi
Szegedi Tudományegyetem Farmakológiai és Farmakoterápiai Intézet Igazgató: Prof. Dr. Varró András 6720 Szeged, Dóm tér 12., Tel.: (62) 545-682 Fax: (62) 545-680 e-mail: varro.andras@med.u-szeged.hu Opponensi
Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén keresztül
 Dr. Miklós Zsuzsanna Semmelweis Egyetem, ÁOK Klinikai Kísérleti Kutató- és Humán Élettani Intézet Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén
Dr. Miklós Zsuzsanna Semmelweis Egyetem, ÁOK Klinikai Kísérleti Kutató- és Humán Élettani Intézet Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Transzporterek vizsgálata lipidmembránokban Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest
 Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Biofizika I. DIFFÚZIÓ OZMÓZIS
 1. KÍSÉRLET 1. kísérlet: cseppentsünk tintát egy üveg vízbe Biofizika I. OZMÓZIS 2012. szeptember 5. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet 1. megfigyelés: a folt lassan szétterjed és megfesti az egész
1. KÍSÉRLET 1. kísérlet: cseppentsünk tintát egy üveg vízbe Biofizika I. OZMÓZIS 2012. szeptember 5. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet 1. megfigyelés: a folt lassan szétterjed és megfesti az egész
Membránszerkezet. Membránszerkezet, Membránpotenciál, Akciós potenciál. Folyékony mozaik modell. Membrán-modellek. Biofizika szeminárium
 Membránszerkezet, Membránpotenciál, Akciós potenciál Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens kötésekkel összetartott lipidekből
Membránszerkezet, Membránpotenciál, Akciós potenciál Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens kötésekkel összetartott lipidekből
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Bari Ferenc egyetemi tanár SZTE ÁOK-TTIK Orvosi Fizikai és Orvosi Informatikai Intézet
 A membránpotenciál eredete. A diffúziós potenciál, Donnan-potenciál, Goldmann-potenciál, a Nernst-Planckegyenlet. A nyugalmi és akciós potenciál (általános jellemzői, ionáramok). Bari Ferenc egyetemi tanár
A membránpotenciál eredete. A diffúziós potenciál, Donnan-potenciál, Goldmann-potenciál, a Nernst-Planckegyenlet. A nyugalmi és akciós potenciál (általános jellemzői, ionáramok). Bari Ferenc egyetemi tanár
A plazmamembrán felépítése
 A plazmamembrán felépítése Folyékony mozaik membrán Singer-Nicholson (1972) Lipid kettősréteg Elektronmikroszkópia Membrán kettősréteg Intracelluláris Extracelluláris 1 Lipid kettősréteg foszfolipidek
A plazmamembrán felépítése Folyékony mozaik membrán Singer-Nicholson (1972) Lipid kettősréteg Elektronmikroszkópia Membrán kettősréteg Intracelluláris Extracelluláris 1 Lipid kettősréteg foszfolipidek
Receptorok, szignáltranszdukció jelátviteli mechanizmusok
 Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL
 Eke András, Kollai Márk FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL Szerkesztette: Ivanics Tamás Semmelweis Kiadó www.semmelweiskiado.hu B u d a p e s t, 2 0 0 7 Szerkesztette: Ivanics Tamás egyetemi docens, Semmelweis
Eke András, Kollai Márk FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL Szerkesztette: Ivanics Tamás Semmelweis Kiadó www.semmelweiskiado.hu B u d a p e s t, 2 0 0 7 Szerkesztette: Ivanics Tamás egyetemi docens, Semmelweis
Potenciálok. Elektrokémiai egyensúly
 Potenciálok Elektrokémiai egyensúly 2/14 edény szemipermeábilis hártyával elválasztva KCl oldat, negatív ion nem tud átlépni kvantitatív jellemzés: elektrokémiai potenciál = + RTlnc + zfe ha ez egyenlő
Potenciálok Elektrokémiai egyensúly 2/14 edény szemipermeábilis hártyával elválasztva KCl oldat, negatív ion nem tud átlépni kvantitatív jellemzés: elektrokémiai potenciál = + RTlnc + zfe ha ez egyenlő
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A membránpotenciál. A membránpotenciál mérése
 A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A diffúzió leírása az anyagmennyiség időbeli változásával A diffúzió leírása a koncentráció térbeli változásával
 Kapcsolódó irodalom: Kapcsolódó multimédiás anyag: Az előadás témakörei: 1.A diffúzió fogalma 2. A diffúzió biológiai jelentősége 3. A részecskék mozgása 3.1. A Brown mozgás 4. Mitől függ a diffúzió erőssége?
Kapcsolódó irodalom: Kapcsolódó multimédiás anyag: Az előadás témakörei: 1.A diffúzió fogalma 2. A diffúzió biológiai jelentősége 3. A részecskék mozgása 3.1. A Brown mozgás 4. Mitől függ a diffúzió erőssége?
BIOFIZIKA. Membránpotenciál és transzport. Liliom Károly. MTA TTK Enzimológiai Intézet
 BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
A Sejtmembrán Szerkezete Nyugalmi Membránpotenciál
 A Sejtmembrán Szerkezete Nyugalmi Membránpotenciál 2012.09.25. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej
A Sejtmembrán Szerkezete Nyugalmi Membránpotenciál 2012.09.25. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej
A transzportfolyamatok és a sejtek közötti kommunikáció
 A transzportfolyamatok és a sejtek közötti kommunikáció A sejtmembrán I.véd II.szelektál (átmenő anyagtranszport szigorúan szabályozott) III.elválaszt (barrier) extracelluláris (sejten kívüli) intracelluláris
A transzportfolyamatok és a sejtek közötti kommunikáció A sejtmembrán I.véd II.szelektál (átmenő anyagtranszport szigorúan szabályozott) III.elválaszt (barrier) extracelluláris (sejten kívüli) intracelluláris
Kálium ioncsatornák eltérő funkciói
 Kálium ioncsatornák eltérő funkciói Elektrofiziológia kurzus 2016/17 őszi szemeszter 2016. 10. 12 Prorok János, PhD Farmakológiai és Farmakoterápiai Intézet Tartalmi áttekintés A K+ csatornákat érintő
Kálium ioncsatornák eltérő funkciói Elektrofiziológia kurzus 2016/17 őszi szemeszter 2016. 10. 12 Prorok János, PhD Farmakológiai és Farmakoterápiai Intézet Tartalmi áttekintés A K+ csatornákat érintő
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A sejtek membránpotenciálja (MP)
") A sejtek membránpotenciálja (MP) XVIII. sz. Galvani, Aldani: "állati elektromosság" az izom és az idegszövet elektromosan ingerlékeny az izom és az idegszövet elektromosan vezetıképes 1939, Hodgkin és
A sejtek membránpotenciálja (MP) XVIII. sz. Galvani, Aldani: "állati elektromosság" az izom és az idegszövet elektromosan ingerlékeny az izom és az idegszövet elektromosan vezetıképes 1939, Hodgkin és
Az agyi értónust befolyásoló tényezők
 2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
Folyadékkristályok; biológiai és mesterséges membránok
 Folyadékkristályok; biológiai és mesterséges membránok Dr. Voszka István Folyadékkristályok: Átmenet a folyadékok és a kristályos szilárdtestek között (anizotróp folyadékok) Fonal, pálcika, korong alakú
Folyadékkristályok; biológiai és mesterséges membránok Dr. Voszka István Folyadékkristályok: Átmenet a folyadékok és a kristályos szilárdtestek között (anizotróp folyadékok) Fonal, pálcika, korong alakú
A szívizomsejtek ionáramai
 A szívizomsejtek ionáramai Dr. Szentesi Péter DE OEC Élettani Intézet 2009 A szivet alkotó szívizomsejtek A sejtmembrán szerkezete Csatornák Pórus Szőrı Kapu A Patch Clamp módszer Egyedi csatorna izolálása
A szívizomsejtek ionáramai Dr. Szentesi Péter DE OEC Élettani Intézet 2009 A szivet alkotó szívizomsejtek A sejtmembrán szerkezete Csatornák Pórus Szőrı Kapu A Patch Clamp módszer Egyedi csatorna izolálása
MEMBRÁNSZERKEZET, MEMBRÁNPOTENCIÁL, AKCIÓS POTENCIÁL. Biofizika szeminárium
 MEMBRÁNSZERKEZET, MEMBRÁNPOTENCIÁL, AKCIÓS POTENCIÁL Biofizika szeminárium 2012. 09. 24. MEMBRÁNSZERKEZET Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens
MEMBRÁNSZERKEZET, MEMBRÁNPOTENCIÁL, AKCIÓS POTENCIÁL Biofizika szeminárium 2012. 09. 24. MEMBRÁNSZERKEZET Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYEK KÁLIUM TÁPLÁLKOZÁSÁNAK GENETIKAI ALAPJAI előadás áttekintése A kálium szerepe a növényi szervek felépítésében
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYEK KÁLIUM TÁPLÁLKOZÁSÁNAK GENETIKAI ALAPJAI előadás áttekintése A kálium szerepe a növényi szervek felépítésében
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Érzékelési folyamat szereplői. Az érzékelés biofizikájának alapjai. Receptor felépítése. Az inger jellemzői MILYEN? HOL? MENNYI? MEDDIG?
 külső, belső környezet ei Érzékelési folyamat szereplői Az érzékelés biofizikájának alapjai specifikus transzducer központi idegrendszer Az jellemzői Receptor felépítése MILYEN? HOL? MENNYI? MEDDIG? Magasabb
külső, belső környezet ei Érzékelési folyamat szereplői Az érzékelés biofizikájának alapjai specifikus transzducer központi idegrendszer Az jellemzői Receptor felépítése MILYEN? HOL? MENNYI? MEDDIG? Magasabb
Ioncsatorna funkciók mérése in vitro körülmények között. Dr. Nagy Norbert Tudományos munkatárs SZTE Farmakológiai és Farmakoterápiai Intézet
 Ioncsatorna funkciók mérése in vitro körülmények között Dr. Nagy Norbert Tudományos munkatárs SZTE Farmakológiai és Farmakoterápiai Intézet Témák: -Alapkutatásban alkalmazott elektrofiziológiai technikák
Ioncsatorna funkciók mérése in vitro körülmények között Dr. Nagy Norbert Tudományos munkatárs SZTE Farmakológiai és Farmakoterápiai Intézet Témák: -Alapkutatásban alkalmazott elektrofiziológiai technikák
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A szív élettana. Aszív élettana I. A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG
 A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa
A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa
Az elmúlt években végzett kísérleteink eredményei arra utaltak, hogy az extracelluláris ph megváltoztatása jelentősen befolyásolja az ATP és a cink
 A cystás fibrosis (CF) a leggyakoribb autoszomális, recesszív öröklődés menetet mutató halálos kimenetelű megbetegedés a fehérbőrű populációban. Hazánkban átlagosan 2500-3000 élveszületésre jut egy CF
A cystás fibrosis (CF) a leggyakoribb autoszomális, recesszív öröklődés menetet mutató halálos kimenetelű megbetegedés a fehérbőrű populációban. Hazánkban átlagosan 2500-3000 élveszületésre jut egy CF
Vázizom elektrofiziológia alapjai. Tóth András, PhD
 Vázizom elektrofiziológia alapjai Tóth András, PhD Témák Struktúra Kontrakció és relaxáció Aktiváció Excitáció-kontrakció csatolás Akciós potenciál Ioncsatornák* Ca 2+ homeosztázis Struktúra Vázizom
Vázizom elektrofiziológia alapjai Tóth András, PhD Témák Struktúra Kontrakció és relaxáció Aktiváció Excitáció-kontrakció csatolás Akciós potenciál Ioncsatornák* Ca 2+ homeosztázis Struktúra Vázizom
Sáry Gyula SZTE ÁOK Élettani Intézet
 A szenzoros transzdukció celluláris alapjai: a szenzoros inger neurális aktivitás összefüggés általános törvényszerűségei, a szenzoros (generátor) potenciál keletkezése különböző szenzoros modalitásokban,
A szenzoros transzdukció celluláris alapjai: a szenzoros inger neurális aktivitás összefüggés általános törvényszerűségei, a szenzoros (generátor) potenciál keletkezése különböző szenzoros modalitásokban,
Látás. Látás. A környezet érzékelése a látható fény segítségével. A szem a fényérzékelés speciális, páros szerve (érzékszerv).
.") Látás A szem felépítése és működése. Optikai leképezés a szemben, akkomodáció. Képalkotási hibák. A fotoreceptorok tulajdonságai és működése. A szem felbontóképessége. A színlátás folyamata. 2014/11/18
Látás A szem felépítése és működése. Optikai leképezés a szemben, akkomodáció. Képalkotási hibák. A fotoreceptorok tulajdonságai és működése. A szem felbontóképessége. A színlátás folyamata. 2014/11/18
SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár
 SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár TDK felelős: Dr. Pataricza János egyetemi docens tel.: (62) 545-674 fax: (62) 545-680 E-mail: Pataricza@phcol.szote.u
SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár TDK felelős: Dr. Pataricza János egyetemi docens tel.: (62) 545-674 fax: (62) 545-680 E-mail: Pataricza@phcol.szote.u
OZMÓZIS. BIOFIZIKA I Október 25. Bugyi Beáta PTE ÁOK Biofizikai Intézet
 BIOFIZIKA I 2011. Október 25. Bugyi Beáta PTE ÁOK Biofizikai Intézet Áttekintés 1. Diffúzió rövid ismétlés 2. Az ozmózis jelensége és leírása 4. A diffúzió és ozmózis orvos biológiai jelentősége Diffúzió
BIOFIZIKA I 2011. Október 25. Bugyi Beáta PTE ÁOK Biofizikai Intézet Áttekintés 1. Diffúzió rövid ismétlés 2. Az ozmózis jelensége és leírása 4. A diffúzió és ozmózis orvos biológiai jelentősége Diffúzió
Nyugalmi potenciál, akciós potenciál és elektrotónusos potenciálok. - Ionális mechanizmusok -
 Nyugalmi potenciál, akciós potenciál és elektrotónusos potenciálok - Ionális mechanizmusok - Támpontok:4-6 Sántha Péter 2017.09.11. Transzmembrán potenciál (E m ) Nyugalmi potenciál (E 0 ): Elektromos
Nyugalmi potenciál, akciós potenciál és elektrotónusos potenciálok - Ionális mechanizmusok - Támpontok:4-6 Sántha Péter 2017.09.11. Transzmembrán potenciál (E m ) Nyugalmi potenciál (E 0 ): Elektromos
Intracelluláris módszerek a szív elektrofiziológiai tulajdonságainak vizsgálatára. Tóth András DSc. 2018
 Intracelluláris módszerek a szív elektrofiziológiai tulajdonságainak vizsgálatára Tóth András DSc. 2018 Témák (a ***-gal jelölt fejezetek a vizsgához nem szükségesek) Preparátumok Sejt izolálás Multicelluláris
Intracelluláris módszerek a szív elektrofiziológiai tulajdonságainak vizsgálatára Tóth András DSc. 2018 Témák (a ***-gal jelölt fejezetek a vizsgához nem szükségesek) Preparátumok Sejt izolálás Multicelluláris
Érzékelési folyamat szereplői. Az érzékelés biofizikájának alapjai. Inger Modalitás Receptortípus. Az inger jellemzői MILYEN? HOL? MENNYI? MEDDIG?
 külső, belső környezet ei Érzékelési folyamat szereplői Az érzékelés biofizikájának alapjai specifikus transzducer központi idegrendszer Az jellemzői MILYEN? HOL? MENNYI? MEDDIG? Magasabb szintű kódolás
külső, belső környezet ei Érzékelési folyamat szereplői Az érzékelés biofizikájának alapjai specifikus transzducer központi idegrendszer Az jellemzői MILYEN? HOL? MENNYI? MEDDIG? Magasabb szintű kódolás
A transzportfolyamatok és a sejtek közötti kommunikáció
 A transzportfolyamatok és a sejtek közötti kommunikáció A sejtmembrán protektív és szelektív barrier kompartmentalizáció: sejtfelszín és sejtorganellumok borítása 1926 szénhidrát 1943 zsírsav 1972 poláros
A transzportfolyamatok és a sejtek közötti kommunikáció A sejtmembrán protektív és szelektív barrier kompartmentalizáció: sejtfelszín és sejtorganellumok borítása 1926 szénhidrát 1943 zsírsav 1972 poláros
-Két fő korlát: - asztrogliák rendkívüli morfológiája -Ca szignálok értelmezési nehézségei
 Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Orvosi élettan. Bevezetés és szabályozáselmélet Tanulási támpontok: 1.
 Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 anyagcsere hőcsere Az élőlény és környezete nyitott rendszer inger hő kémiai mechanikai válasz mozgás alakváltoztatás
Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 anyagcsere hőcsere Az élőlény és környezete nyitott rendszer inger hő kémiai mechanikai válasz mozgás alakváltoztatás
Az idegsejtek diverzitása
 Az idegsejtek diverzitása Készítette Dr. Nusser Zoltán előadása és megadott szakirodalma alapján Walter Fruzsina II. éves PhD hallgató A neurobiológia hajnalán az első idegtudománnyal foglalkozó kutatók
Az idegsejtek diverzitása Készítette Dr. Nusser Zoltán előadása és megadott szakirodalma alapján Walter Fruzsina II. éves PhD hallgató A neurobiológia hajnalán az első idegtudománnyal foglalkozó kutatók
SZAGLÁS 2
 AZ ÉRZÉKELÉS BIOLÓGIÁJA 1 SZAGLÁS 2 ÍZLELÉS 3 HALLÁS 4 ÉRINTÉS EGYENSÚLY 5 FÁJDALOM 6 túl az emberi érzékelésen 7 HOGYAN ALAKÍTJÁK ÁT A RECEPTOR SEJTEK A KÜLÖNBÖZŐ STIMULUSOKAT AKCIÓS POTENCIÁLLÁ? HOGYAN
AZ ÉRZÉKELÉS BIOLÓGIÁJA 1 SZAGLÁS 2 ÍZLELÉS 3 HALLÁS 4 ÉRINTÉS EGYENSÚLY 5 FÁJDALOM 6 túl az emberi érzékelésen 7 HOGYAN ALAKÍTJÁK ÁT A RECEPTOR SEJTEK A KÜLÖNBÖZŐ STIMULUSOKAT AKCIÓS POTENCIÁLLÁ? HOGYAN
OZMÓZIS, MEMBRÁNTRANSZPORT
 OZMÓZIS, MEMBRÁNTRANSZPORT Vig Andrea PTE ÁOK Biofizikai Intézet 2014.10.28. ÁTTEKINTÉS DIFFÚZIÓ BROWN-MOZGÁS a részecskék rendezetlen hőmozgása DIFFÚZIÓ a részecskék egyenletlen (inhomogén) eloszlásának
OZMÓZIS, MEMBRÁNTRANSZPORT Vig Andrea PTE ÁOK Biofizikai Intézet 2014.10.28. ÁTTEKINTÉS DIFFÚZIÓ BROWN-MOZGÁS a részecskék rendezetlen hőmozgása DIFFÚZIÓ a részecskék egyenletlen (inhomogén) eloszlásának
A LÁTÁS BIOFIZIKÁJA AZ EMBERI SZEM GEOMETRIAI OPTIKÁJA. A szem törőközegei. D szem = 63 dioptria, D kornea = 40, D lencse = 15+
 A LÁTÁS BIOFIZIKÁJA A SZÍNLÁTÁS ELMÉLETE ELEKTRORETINOGRAM Két kérdés: Sötétben minden tehén fekete Lehet-e teniszt játszani sötétben kivilágított hálóval, vonalakkal, ütőkkel és labdával? A szem törőközegei
A LÁTÁS BIOFIZIKÁJA A SZÍNLÁTÁS ELMÉLETE ELEKTRORETINOGRAM Két kérdés: Sötétben minden tehén fekete Lehet-e teniszt játszani sötétben kivilágított hálóval, vonalakkal, ütőkkel és labdával? A szem törőközegei
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
Szerkezet és funkció kapcsolata a membránműködésben. Folyadékkristályok típusai (1) Dr. Voszka István
 Dr. Voszka István") MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2015/2016 II. félév Időpont: szerda 17 30-19 00 Helyszín Elméleti Orvostudományi Központ Szent-Györgyi Albert előadóterme II. 3. Szerkezet
MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2015/2016 II. félév Időpont: szerda 17 30-19 00 Helyszín Elméleti Orvostudományi Központ Szent-Györgyi Albert előadóterme II. 3. Szerkezet