Celluláris és molekuláris neurobiológia SzJDI őszi félév. Neuron-specifikus génműködés. Szabó Gábor MTA KOKI
|
|
|
- Zsófia Faragó
- 7 évvel ezelőtt
- Látták:
Átírás

1 Celluláris és molekuláris neurobiológia SzJDI őszi félév Neuron-specifikus génműködés Szabó Gábor MTA KOKI


2 Génexpressziós szintek Génhez való hozzáférés kromatin DNS metiláció kromatin modifikáció epigentika Átírás cisz és transzelemek akivitás függő elemek RNS érés (processing) alternatív splicing mrns szabályozás mikrorns-ek editálás Transzláció lokális aktivitás függő epigentika

3 Génexpresszió kapcsolt lépései

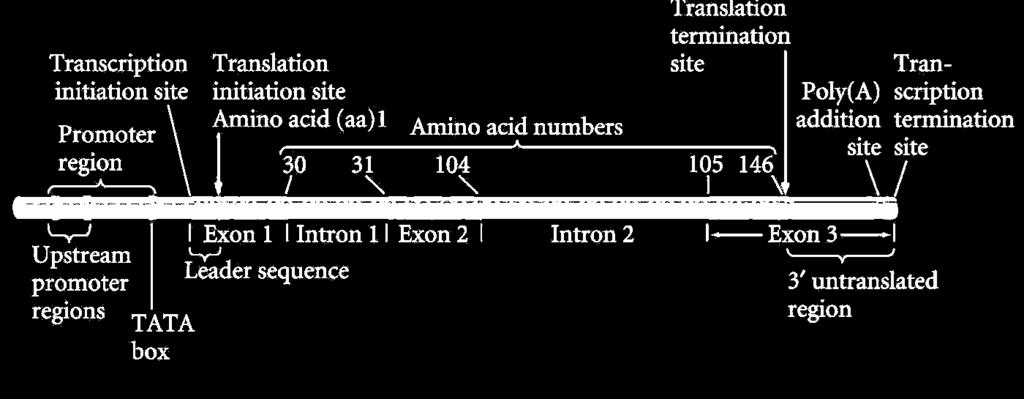
4 Transzkripciós egység-génszerkezet 5 3 szabályozó régió kódoló régió

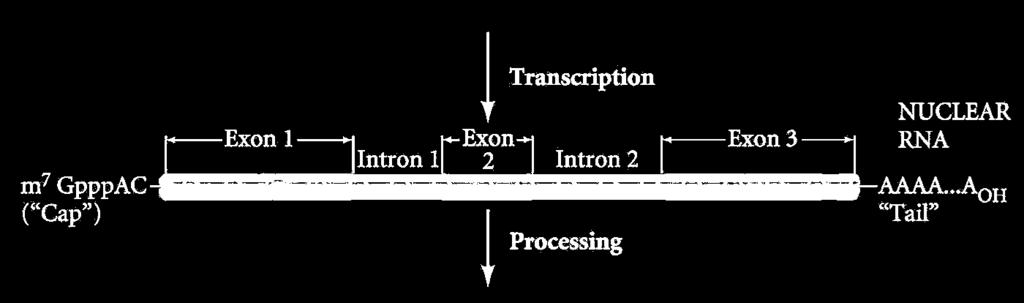
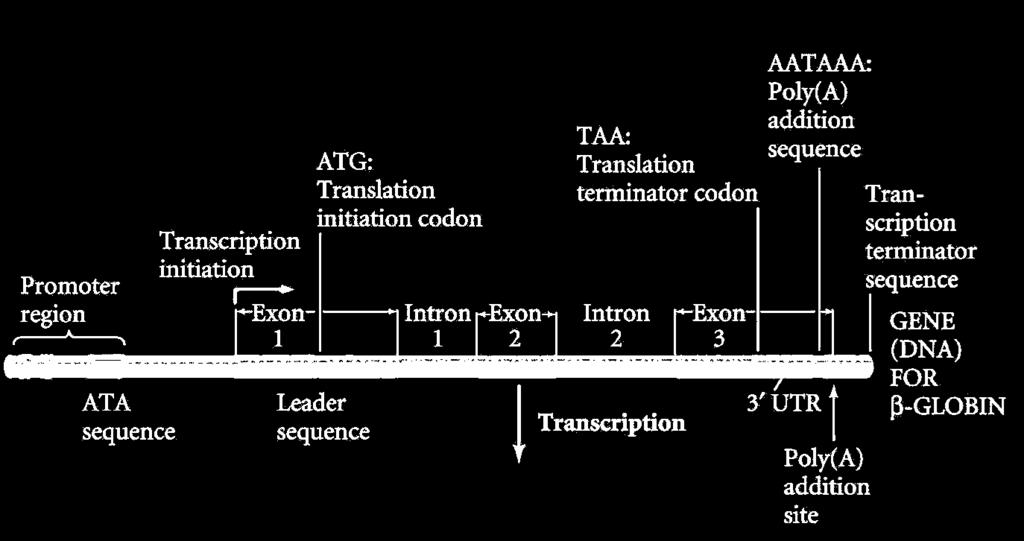

5 mrns képződés Transzkripció+ processing + transzport
 - mrns leader szekvenciák transz elemek - az adott géntől független faktorok - fehérjék (más gének termékei) (lehetnek RNS-ek is microrns) - transzkripciós faktorok (aktivátorok,")
6 Transzkripció szabályozásának elemei cisz elemek - DNS szabályozó elemek gének részei- promoterek, transzkripciós faktor kötőhelyek, enhancerek (silencerek) (elhelyezkedésük 5 -vég, 3 -vég, intronok, kódoló régió) - mrns leader szekvenciák transz elemek - az adott géntől független faktorok - fehérjék (más gének termékei) (lehetnek RNS-ek is microrns) - transzkripciós faktorok (aktivátorok, represszorok) cisz elemekhez kötődnek - kofatorok (ko-aktivátorok, ko-represszorok) transzkripciós faktorokhoz kötődnek
 promoter proximális régió - upstream regulációs elemeket tartalmaz promoter Promoter típusok house")

7 enhancers upstream promoter elements core promoter Promoter felépítése core promoter ( -40 to +40 a TSS-hez viszonyítva) promoter proximális régió - upstream regulációs elemeket tartalmaz promoter Promoter típusok house keeping - minden/legtöbb sejtben működnek (TATA nélküli CpG sziget) szövetspecifikus- TATA/CAAT szövetspecifikus elemek indukálható/fejlődés szabályozott Gének átírása polimeráz II- RNS szintetizáló enzim mrns-kódoló gének mikrorns-kódoló gének
 BRE - TFIIB")

8 Promoter elemek (RNS polimeráz II) BRE - TFIIB kötő elem DPE - downstream promoter elem Inr - iniciátor régió


9 Általános upstream promoter elemek Az iniciáció frekvenciáját növelik: elősegítik azt, hogy a bazális transzkripciós faktorok iniciációs komplex-szé szerveződjenek. upstream promoter elemek

10 Transzkripciós faktorok -transz ható elemek - regulációs fehérjék aktivátorok, vagy inhibitorok Funkcionális doménjeik - DNS kötő domén - promoter, vagy enhancer DNS elemekhez kötődik - Aktivációs domén - transzkripciós faktorokkal, ko-aktivátorokkal, ko-represszorokkal, vagy közvetlenül az RNS polimeráz II komplex-szel lép kapcsolatban - Dimerizációs domén: dimér formájában működnek Egyéb faktorokkal transzkripciós komplexet képeznek és serkentik, vagy gátolják az RNS polimeráz II kötődését a promoterhez.
11 Transzkripciós faktorok osztályozása 1. Általános transzkripciós faktorok TFIID, TFIIB, TFIIA, TFIIH, TFIIE, TFIIF Közvetlenül az RNS polimeráz II működéséhez kell. Minden sejtben megtalálható. 2. Minden sejtben jelenlévő konstitutív transzkripciós faktorok sok gén transzkripcióját aktiválja (SP1, CCAAT-box kötő fehérje ) 3. Szövetspecifikus transzkripciós faktorok Specifikus trasz-elemhez kötődik Adott sejttípusban fejeződik ki Szövetspecifikus géneket aktivál 4. Szignál függő/aktiválható transzkripciós faktorok
12

13 Transzkripciós faktorok kötődése a cisz DNS elemekhez
 Science 291,")
14 A humán genomban található transzkripciós faktor gének Venter et al al al (2001) Science 291, 1304-
15 Cisz regulációs elemek: enhancerek/silencerek enhancers upstream promoter elements core promoter Cis regulációs elemek: aktivátorok (enhancerek), gátlók ( silencerek ) Promoterektől a következőkben térnek el: 1. Enhancerek működéséhez szükségesek a promoterek (promoterek enhancerek nélkül is működnek, bár gyengén) 2. Az enhancerek a transzkripciós starthelytől távol is működnek, a promoterek nem 3. fordított orientációban is működnek 4. a transzkripciós kezdőhely mögött is működnek 5. enhancerek nagyon távoli helyekről is működnek A transzkripciót térben és időben szabályozottan fokozzák, vagy gátolják. Működésükhöz szükséges a transzkripciós alap-apparátus. A transzkripció iniciációját serkentik, vagy gátolják.


16 Az enhancerhez kötődő aktivátor fehérjék a transzkripciós komplexhez kötődnek és növelik a transzkripció hatásfokát Lehetnek: szövet/sejttípus specifikusak fejlődés specifikusak aktiválhatók
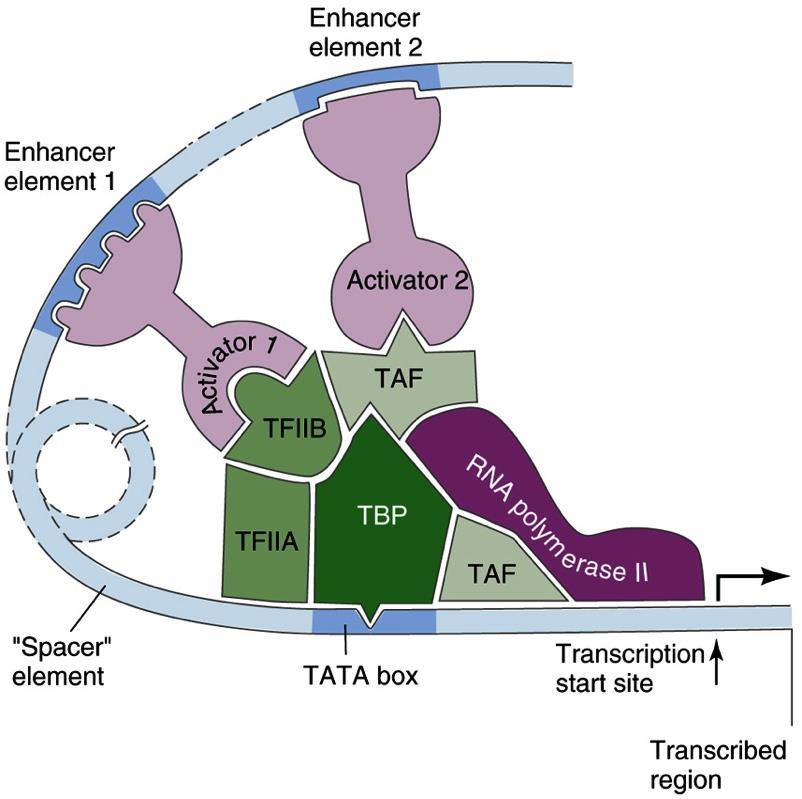
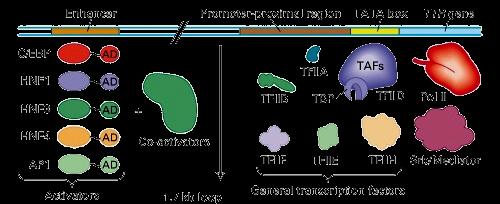
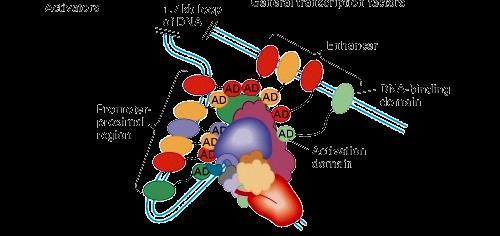
17 Komplex enhancerek general transcription factors


18 Represszorok Represszorok a silencerekhez kötődnek gátolják az aktivátorok kötődését közvetlenül gátolják az aktivátort közvetlenül kötődnek az általános transzkripciós faktorokhoz


19 A transzkripció szabályozásában résztvevő transz-elemek

20 Tipikus emlős génregulációs régió Az emlős géneket a promotereken kívül több enhancer és silencer szabályozza. Egy tipikus enhancer 500 bp hosszú 10 kötőhelyet tartalmaz, amelyhez legalább háromféle transzkripciós faktor, valamint 2 különböző aktivátor és egy represszor kötődik.

21 Szövetspecfikus transzkripciós szabályozás: szövetspecifikus enhancerek-transzkipicós faktorok
22 Ligand-függő transzkripciós szabályozás




23 Nukleáris hormon receptorok: ligand függő transzkripciós faktorok szteroid hormonok tiroid hormonok retinidok D vitamin

24 Nukleáris hormon receptorok szerkezete
 Indirekt transzkripciós")
25 Nukleáris hormonreceptorok működési mechanizmusa 1: NR ligand nélkül inaktív komplexet képez a citoplzmában (hsp90) 2: Ligand kötés-nukleáris transzlokáció : DNS elemhez kapcsolódik koaktivátorokkal és más fehérjékkel 4: Gén transzkripció beindul 5: Más transzkripciós faktorokhoz is kapcsolódhat (NFkB, AP-1) Indirekt transzkripciós szabályozás
26 Aktivitás függő transzkripciós szabályozás az idegrendszerben

27 Szignál specifikus transzkripciós komplexek az idegsejtekben kinases Különböző stimulusok hatására CaMP és Ca ++ reszponzív elemekhez kapcsolódnak a foszforiláció után a faktorok. Beindítják a c-fos és Bdnf transzkripciót.

28 CREB függő gén expressziós kaszkád aktivitása szükséges a neuronális plaszticitáshoz és a memóriához szignálok neurotranszmittertek növekedési faktorok depolarizáció stress kinázok CREB foszforiláció C/EBP TF gén aktiváció késői gének aktivációja C/EBP=CAAT enhancer binding protein plaszticitás memória


29 Az aktivitás függő Bdnf transzkripció szerepe a GABAerg szinapszisok szabályozásában Bdhf gén IV. exon
30 RNS összeszerelődés (processing) 2 processing 1 transcription cap splicing polia

31 Alternatív splicing típusok

32 Mire jó az alternatív splicing? A gének exonjaiból a kódoló régiók több féle módon szerelődhetnek össze A genom kódoló kapacitását növeli Fehérjék funkcionális komplexitását növeli eltérő N-, vagy C-eltérő domén szerkezet funkcionális eltérés változik a szubcelluláris lokalizáció poszt-transzlációs modifikáció fehérje-fehérje kölcsönhatás ligand kötés mrns-ek nem-kódoló régióinak diverzitása RNS szintű szabályozás RNS szint, targetálás, stabilitás transzláció szabályozása (microrns kötődés) Lehet fejlődés és szövetspecifikus és aktivitás függő is
33 Splicing mechanizmus

34 Alternatív splicing mechanizmussal egy génről többféle mrns és fehérje képződik -tropomyosin gén

 A DScam1")

35 Splicing világbajnok a Drosophila Dscam1 (Down Syndrom cell adhesion molecule) gén 2 Alt. (38, 016) A DScam1 izoformák a dendrit arborizációt a dendrit típusok között átfedést szabályozzák.

36 Alternatív splicing az idegrendszerben Alternatív splicing eredményeképpen képződő receptor, ioncsatorna, enzim, jelátviteli fehérje strukturális fehérje (sejtváz, sejtadhézió) formáknak eltérőek lehetnek: funkcionális tulajdonságai lokalizációja, membrán kötődése interakciói Vannak idegsejt specifikus splicing faktorok. Az alternatív splicing-et a neuronális aktivitás is szabályozhatja.


37 A glutamáterg és GABAerg szinapszis képződést (gátló vagy serkentő szinaptikus specializáció) a preszinaptikus neurexin és a posztszinaptikus Neuroligin alternatív spilce -formái határozzák meg 3 neurxin génről összesen több mint 1000 izoforma képződhet (5 altenetív exon és 2 promoter) 4/5 neuroligin gén 2-2 splice formája 8-10 izoformát kódol. A kettő nagyszámú kombinációja sokféle gátló és serkentő szinaptikus kapcsolat kialakítását szabályozhatja.

38 AChE alternatív splicing: különböző szerkezetű és lokalizációjú enzimet eredményez intron retenció alternatív 3 exon alternatív 3 exon hidrifil monomér membrán-kötött dimér VVS membrán kapcsolt tetramer szinaptikus membrán kötött


39 AMPA receptor alternatív splicing: flip/flop variáns Q/R editing a Ca ++ permeábilitást szabályozza Flip/Flop AS és az R/G editing a deszenzitzácót szabályozza

40 Idegrendszerben működő splicing faktorok
")
41 A hippocampus CA1 régiójában az NMD receptor aktiváció által kiváltott LTP-hez szükséges a NOVA2 splicing faktor NOVA2 KO egér: sipsc, LTP NOVA2 alternatív splicing NMDAR GABABR2 GIRK2 sipsc (lassú gátló posztszinaptikus áram) LTP

42 Szinaptikus mrns-ek, amelyek alternatív splicing-jét a NOVA szabályozza

43 Génműködés epigenetikus szabályozása Olyan változások, amelyek a genom változtatása nélkül hatással vannak a fenotípusra. Egy sejt tulajdonságainak örökletes megváltozása, amely nem jár együtt a genetikai információ megváltozásával. Génműködés epigenetikus szabályozása: nem függ közvetlenül az adott gén regulációs DNS elemeitől: kromatin (hiszton) módosítás DNS metiláció RNS interferencia RNS editálás
44 Epigenetikus hatások: kapcsolat a környezet és a genom között Környezeti hatások eltérő egyedi következményei: egyedi epigenetikus variabilitás környezet (toxinok, gyógyszerek, táplálkozás, sport, szociális hatások) epigenetikus változás személyek közötti epigenetikus variabilitás génexpressziós programozás FENOTIPIKUS VARIABILITÁS

45
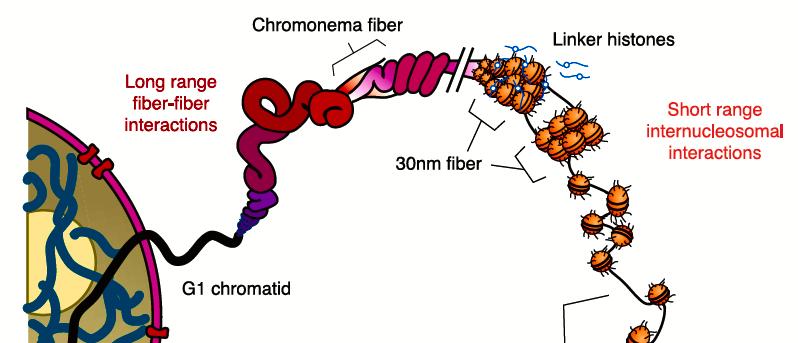
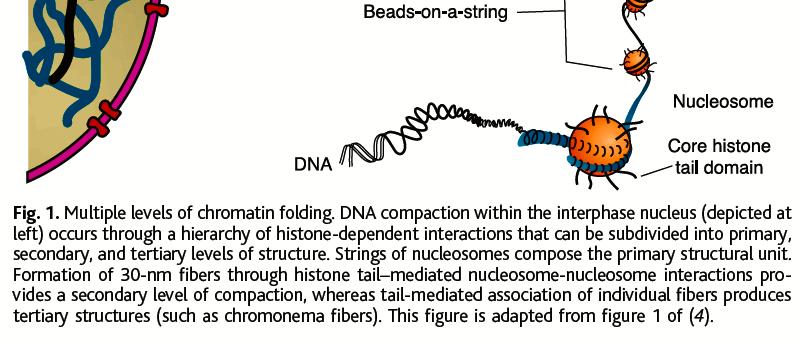

46 DNS-től a kromoszómákig: A DNS kromatin formájában található a sejtmagban

47 Kromatinszerkezet és szerveződés nucleosome
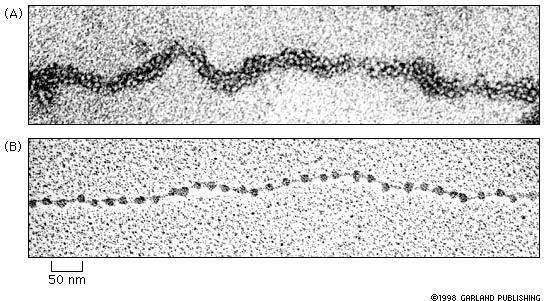
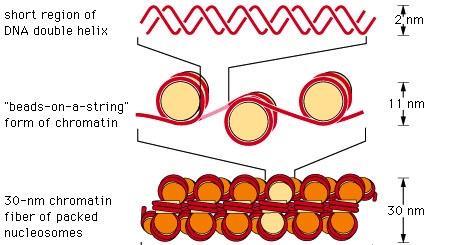
48
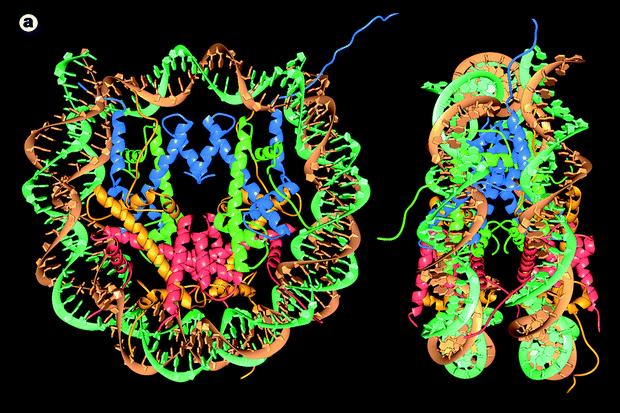
49 Nukleoszóma szerkezet
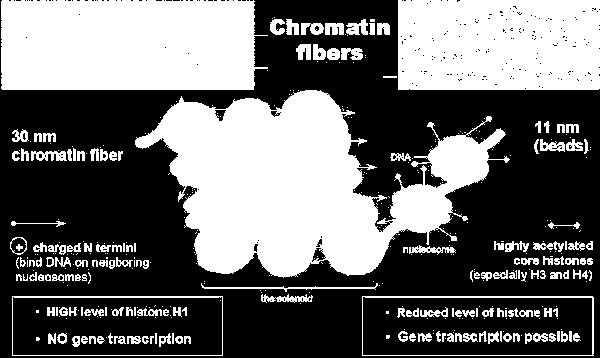
 riboziálció metiláció acetiláció hiperacetiláció hiszton acetiláz laza nukleoszómák transzkripciós aktiváció hipoacetiláció hiszton deacetiláz szoros nukleoszóma")
50 Hisztonok (H1, H2A, H2B, H3, H4) kis fehérjék arginin- vagy lizingazdag: pozitívan töltött kötődik a negatívan töltött DNS-hez poszt-transzlációs módosítás - csökkenti a pozitív töltést foszforiláció poli(adp) riboziálció metiláció acetiláció hiperacetiláció hiszton acetiláz laza nukleoszómák transzkripciós aktiváció hipoacetiláció hiszton deacetiláz szoros nukleoszóma transzkripciós gátlás

51 Hiszton kód: hiszton fehérjék poszt-transzlációs módosítása


52 Hiszton acetiláció a kromatin kondenzációt szabályozza Hiszton acetiláció aktiválja a transzkripciót

53 A transzkripciós aktivátorok és represszorok a kromatin szerkezetének módosításán keresztül is hatnak
54 DNS metiláció: citozin metiláció

55 A CpG dinukleotidok alulreprezentáltak a genomban A CpG előfordulása alacsonyabb a vártnál várt gyakoriság 1/16= 6.25% oka Met-citozin spontán deaminációja timidint eredményez- csökken a CpG aránya Lehetséges dinukleotid párok GG GA GT GC AG AA AT AC TG TA TT TC CG CA CT CC megfigyelt gyakoriság 1 % O NH 2 NH 2 N N CH 3 N O N deamination O O HN CH 3 HN O O N N C mc T U A genomban vannak olyan régiók ahol a CpG arány megtartott, illetve magas. Itt a CpG nem metilált. CpG szigetek. Ezek regulációs régiók.
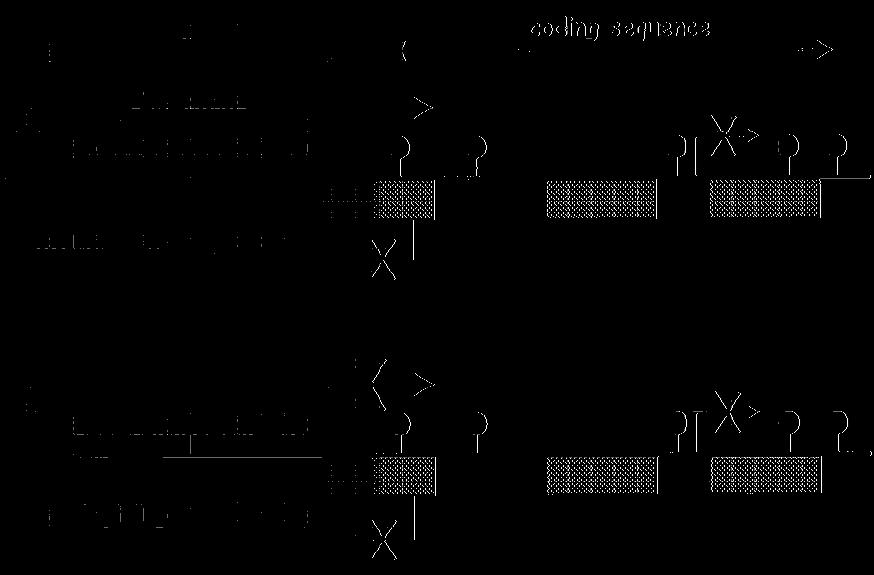
56 A CpG szigetek kijelölik a promotereket CpG szigetek = DNS régió a várt CpG frekvenciával hossza = 200 bp -tól néhány kb-ig. CpG a szigeten belül döntően nem-metilált, míg a CpG szigeten kívül metilált. A humán genomban CpG sziget van (a genom 1-2%-a) CpG szigetek majdnem mindig promotereket és/vagy exonokat tartalmaznak. ( a gének 50-60% tartalmaz CpG szigetet. CpG szigetben található promoterek nem tartalmaznak TATA vagy DPE elemeket, de többszörös GC motívumokat tartalmaznak. - gyakran többszörös és gyenge transzkripciós starthellyel rendelkeznek (többszörös Sp1+Inr miatt ) Ez a metilációs mintázat a genomot transzkripciósan aktív és inaktív zónákra osztja.
57 A CpG szigetek kijelölik a promotereket A metilált és nem metilált CpG nem random található a genomban Metilált Nem-metilált TSS AGCGAGCGAGCGTGTATGTTCTCATTAGGGGACGATC TCGCTCGCTCGCACATACAAGAGTAATCCCCTGCTAG Hemimetilát legtöbb CpG metilált az emlős sejtekben CpG promóterekhez kapcsoltak

58 A konstitutívan működő háztartási gének promoterei CpG szigetben találhatók. Van olyan szövetspecifikus gén is, amelyiknek a promotere szintén CpG szigetben helyezkedik el. A CpG szigetben a CpG nem metilált- a promoter aktív. Metilációval a promoter inaktiválható.

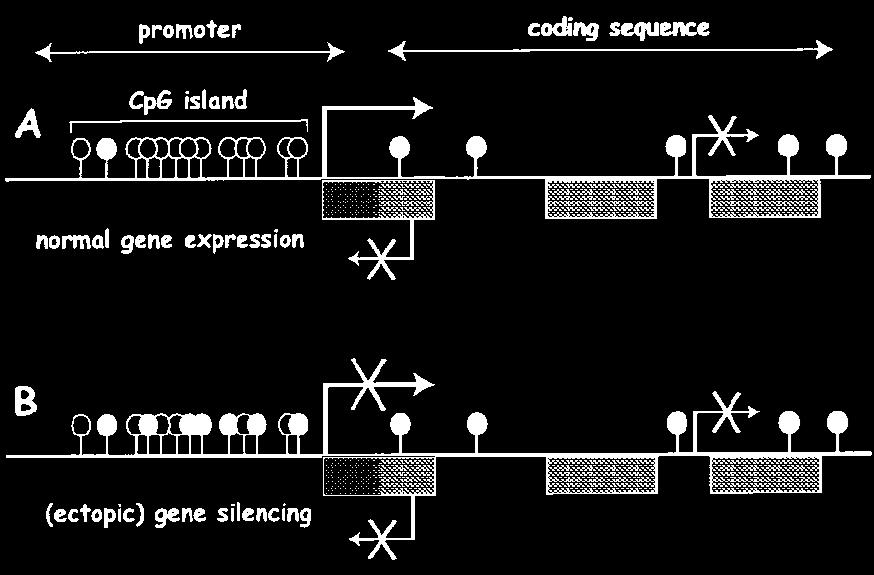

59 Gének ki-be kapcsolása DNS metilációval BE: CpG sziget demetiláció KI: CPG sziget metiláció

60 DNA metiláció funkcionális következménye Metilált promoter inaktív A a CpG sziget mentes szövetspecifikus gének automatikusan metiláltak, ennek a következménye a globális inaktiváció, aminek a fenntartásához nem kellenek további elemek. Metilkötő fehérje A metilált CpG-ket a metilkötő fehérjék ismerik fel (MBP), amelyek különböző hisztonmódosító enzimeket vonzanak (HDAC-hiszton deacetiláz, HMT hiszton metil transzferáz)- következmény kromatin kondenzáció-inaktiváció. A DNS metilációnak direkt szerepe lehet a nukleoszómák pozicionálásában is.

61 DNS és hiszton modifikáció együttes szerepe a kromatin-szerkezet szabályozásában
62 CpG-ben gazdag promoterrel rendelkező idegrendszeri gének: epigenetikus szabályozás alatt állhatnak

63 Epigenetikus szabályozás az idegrendszerben egyedi gének sejtek: idegi őssejt differenciáció (fejlődés alatt felnőttben) jelátviteli folyamatok szinaptikus folyamatok plaszticitás memória hálózati aktivitás oszcilláció magatartás kóros idegi tevékenység
64 Szinaptikus plaszticitás epigenetikus szabályozása

65 Epigenetikus szabályozás zavara: idegrendszeri betegségek

66



67 Reelin és a GAD76 promoter hipermetilációja skizofréniás betegek preforntális cortexében DNMT promoter metiláció Reelin/GAD67 csökkent gátlás dendrit tüske változások oscillációs zavarok kognitív zavarok
 mrns-t szinetet csökkent (degradál) Szabályozó nem fehérjekódoló RNS-ek (npc) Eukarióta gén mrns + npc RNS")
68 Génműködés poszt-transcripciós szabályozása RNS interferenciával (géncsendesítés) RNS-RNS interakción alapszik poszt-transzkripciós szinten hat mrns-t inaktivál (transzlációt gátol) mrns-t szinetet csökkent (degradál) Szabályozó nem fehérjekódoló RNS-ek (npc) Eukarióta gén mrns + npc RNS Fehérje

69 A genomban fehérjét nem-kódoló RNS gének is találhatók, amelyek szabályozó RNS-eket kódolnak



70 Génműködés szabályozása genom kódolt mikro-rns-ekkel MicroRNS-ek Kisméretű nem-kódoló RNS család, amely szekvencia specifikusan szabályozza a génexpresszió különböző szintjeit.

71 Micro-RNS-ek képződés és hatásának mechanizmusa
72
73

74 A micro RNS-ek az idegrendszer fejlődésének minden lépést szabályozzák


75 Micro RNS-ek szerepe a szinaptikus tüske plaszticitás szabályozásában micrns-ek transzkripciós faktorokon, deacetilázokon, kinázokon és növekedési faktorokon keresztül szabályozzák az aktin citoszkeleton komponenseit serkentő gátló

76 Micro-RNS-ek és az Alzheimer kór mirns-ek szabályozzák az APP mrns és a BACE1 processzáló enzimet kódoló mrns szintjét.
77 An overview of selected mirnas associated with neurological disorders mirna Disease Dysregulation Target mir-9/9* Huntington disease Downregulated mir-29a AD Downregulated NAV3 REST (mir-9) CoREST (mir-9*) Consequence of mirna dysregulation Excessive amount of REST and CoREST in the nucleus Inactivation of neuron-specific genes NAV3 coexpressed tau neurofibrillary tangles in pyramidal neurons mir-29a,-29b-1 Sporadic AD Downregulated BACE1 Increase of amyloid-β mir-106b AD Downregulated APP Increase of amyloid-β mir-107 AD Downregulated BACE1 Increase of amyloid-β mir-124 FXS Downregulated Decreased neuronal maturation mir-133b PD Upregulated Pitx3 mir-146a/b Rett syndrome Downregulated Irak1 Repression of neuronal maturation An increase in the amount of Irak1 puts the brain into an inflammatory state Upregulated in GRIA2 mir-181b Schizophrenia Decrease in neuronal outgrowth prefrontal cortex VSNL1 mir-298 and -328 AD Unknown BACE1 Increase of amyloid-β mir-342-3p Prion disease Upregulated mir-485-5p AD Downregulated in entorhinal cortex and hippocampus BACE1 Increase of amyloid-β
")
78 Poszttranszkripciós szabályozás: RNS editálás (editing) RNS-ben a DNS által kódolt nukleotid (bázis) enzimatikus reakcióval történő megváltoztatása C-U A-I editing guanozin a kodon szempontjából adenozin dezaminálás inozin kodon változás adenozin dezamináz (ADAR) aminósav csere

 szbályozása funkció")
79 A/I editálás az RNS különböző helyein történhet mrns kódoló régió- kodon változás- aminósav csere - fehérje funkció változás UTR megváltozik az mrns transzlációja, lokalizációja vagy stabilitás intron/splice brach site - megválozik az altertnative splicing - új fehérje izoforma STOP kodon eliminálódik- kódoló régió hosszabb Nem-kódoló RNS- mirns biogenezis (szint) szbályozása funkció szabályozása?
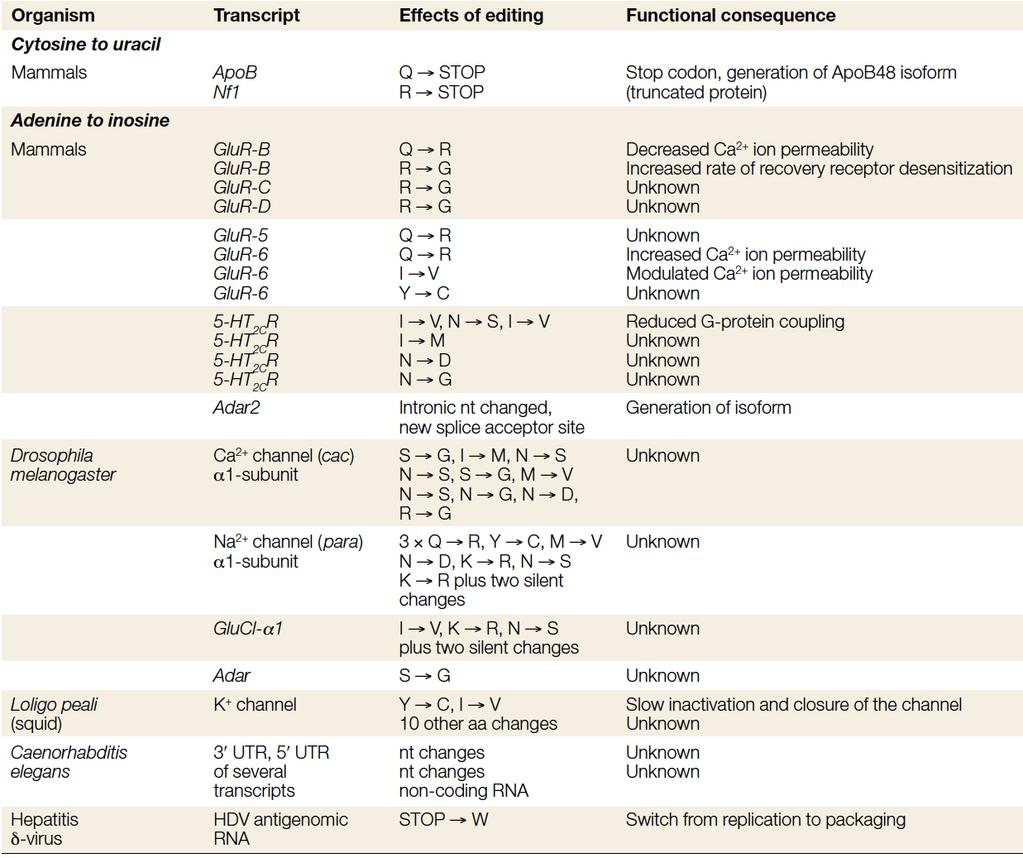
80
-V csere G-protein kötés GABA A 3 I/M")
81 A/I RNS editálás hatása neuronális receptor és csatorna funkcióra K V 1.1 I/V csere kapcsolat a K V b1.1-el Ca v 1.3 IQ-MR csere calmodulin kötés 0 GluA2- Q/R csere Ca ++ permeábilitás ER exit 5-HT 2C I-N-I/ V-S(N)-V csere G-protein kötés GABA A 3 I/M csere membrán trafficing
82 editált genom kódolt
83 mrns anatómiája Az mrns a fehérje kódoláson kívül olyan szekvenciákat és struktúrákat tartalmaz, amelyekhez fehérjék kötődve szabályozzák a transzlációt, transzportot, lokalizációt és stabilitást. 5 -UTR+ faktorok transzláció hatásfoka 3 -UTR+ faktorok mrns transzport stabilitás transzláció


84 Lokális fehérjeszintézis az idegrendszer sejtjeinek nyúlványaiban Előfordul: dedritek axonok- növekedés, cél-megtatlás, sérülés glia-nyúlványok oligodendroglia- mielinizáció Asztrocita-GFAP Aktivitás függő transzláció a dendritekben lehetővé tesz szinapszisok morfológiájának és aktivitásának egyedi szabályozását
85 RNS transzport és lokális fehérjeszintézis szignál mrns inaktív aktív
86 Lokalizációs szignálok idegsejt-specifikus mrns-ek 3 - nem-transzlálódó (3 -UTR) régiójában

87 Transzláció szabályozása a dendritekben Neurotranszmitter receptor kapcsolt útvonalak serkentő szinapszisok esetében BDNF R szignál függő kinázok foszforiláció RNS kötő fehérjék mirns általános RNS specifikus

88 Hosszú 3 -UTR-t tartalmazó BDNF mrns a dendritekbe transzportálódik és itt transzlálódik. Dendritikus BDNF szabályozza a dendrit tüske morfológiáját és a szinaptikus plaszticitást.

89 A CamK2 mrna hosszú 3 -UTR-t eltávolítva megszűnik a dendritikus lokalizáció és zavart szenved a szinaptikus plaszticitás.

90 Szinaptikus aktivitás stimulálja az Arc transzkripcióját és a lokális transzlációját. Arc modulálja szinaptikus plaszticitást az AMPA receptorok endocitózisán keresztül.
91 CREB lokális transzlációja az axonban NGF hatására. Majd retrográd transzportálódik a sejtmagba és NGF reszponzív géneket aktivál.

92 A RanBP1 és importinb lokális szintézise az axonba hozzájárul a retrográd transzport jelátviteli komplex létrehozásában a sérülést követően.

93 A denritekben lokálisan szintetizálódott fehérjék szerepe a hippocampalis LTP-ben


94

95 A c-fos gén transzkripciójának Ca++-függő szabályozása

96 Neuronális aktivitás RNS kötő fehérjék Ca++ függő foszforilációján és hiszton modifikáción keresztül szabályozza az alternatív splicing-ot

97 Szövetspecifkus alternatív exon a fehérje-fehérje kölcsönhatást is szabályozza

98 Az RNS editálás megváltoztatja humán felszültségfüggő kálium csatorna tulajdonságait
Celluláris és molekuláris neurobiológia SzJDI 2014. őszi félév. Neuron-specifikus génműködés. Szabó Gábor MTA KOKI szabog@koki.hu
 Celluláris és molekuláris neurobiológia SzJDI 2014. őszi félév Neuron-specifikus génműködés Szabó Gábor MTA KOKI szabog@koki.hu Génexpressziós szintek Génhez való hozzáférés kromatin DNS metiláció kromatin
Celluláris és molekuláris neurobiológia SzJDI 2014. őszi félév Neuron-specifikus génműködés Szabó Gábor MTA KOKI szabog@koki.hu Génexpressziós szintek Génhez való hozzáférés kromatin DNS metiláció kromatin
Epigenetikai Szabályozás
 Epigenetikai Szabályozás Kromatin alapegysége a nukleoszóma 1. DNS Linker DNS Nukleoszóma mag H1 DNS 10 nm 30 nm Nukleoszóma gyöngy (4x2 hiszton molekula + 146 nukleotid pár) 10 nm-es szál 30 nm-es szál
Epigenetikai Szabályozás Kromatin alapegysége a nukleoszóma 1. DNS Linker DNS Nukleoszóma mag H1 DNS 10 nm 30 nm Nukleoszóma gyöngy (4x2 hiszton molekula + 146 nukleotid pár) 10 nm-es szál 30 nm-es szál
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
TÉMAKÖRÖK. Ősi RNS világ BEVEZETÉS. RNS-ek tradicionális szerepben
 esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
esirna mirtron BEVEZETÉS TÉMAKÖRÖK Ősi RNS világ RNS-ek tradicionális szerepben bevezetés BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek BIOLÓGIAI MOLEKULÁK FEHÉRJÉK NUKLEINSAVAK DNS-ek RNS-ek
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Az X kromoszóma inaktívációja. A kromatin szerkezet befolyásolja a génexpressziót
 Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
- Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=>
, így próbálta leírni a sejt specializációt=>") 1 - Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=> a sejtek sorsa meg van határozva, mint egy üveggolyó,
1 - Conrad Hal Waddington számára a gének fizikai háttere még ismeretlen volt (Watson-Crick-Franklin 1953), így próbálta leírni a sejt specializációt=> a sejtek sorsa meg van határozva, mint egy üveggolyó,
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
DNS replikáció. DNS RNS Polipeptid Amino terminus. Karboxi terminus. Templát szál
 DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Fehérje expressziós rendszerek. Gyógyszerészi Biotechnológia
 Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Fehérje expressziós rendszerek Gyógyszerészi Biotechnológia Expressziós rendszerek Cél: rekombináns fehérjék előállítása nagy tisztaságban és nagy mennyiségben kísérleti ill. gyakorlati (therapia) felhasználásokra
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
RNS-ek. 1. Az ősi RNS Világ: - az élet hajnalán. 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek
 RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
RNS-ek RNS-ek 1. Az ősi RNS Világ: - az élet hajnalán 2. Egy már ismert RNS Világ: - a fehérjeszintézis ben résztvevő RNS-ek 3. Egy újonnan felfedezett RNS Világ: - szabályozó RNS-ek 4. Transzkripció Ősi
Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában
 Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában Czimmerer Zsolt, Csenki-Bakos Zsolt, Urbányi Béla TOX 2017 Tudományos Konferencia 2017.10.12. Bükfürdő A sejtek
Epigenetikai mintázatok biomarkerként történő felhasználási lehetőségei a toxikológiában Czimmerer Zsolt, Csenki-Bakos Zsolt, Urbányi Béla TOX 2017 Tudományos Konferencia 2017.10.12. Bükfürdő A sejtek
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét. Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet
 Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
CzB 2010. Élettan: a sejt
 CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Molekuláris biológiai alapok
 Biokémiai és Molekuláris Biológiai Intézet Molekuláris biológiai alapok Sarang Zsolt Dimenziók a biológiában Fehérjék (kb. 50 ezer különböző fehérje a szervezetben 21 féle aminosavból épül fel) Élő szervezetek
Biokémiai és Molekuláris Biológiai Intézet Molekuláris biológiai alapok Sarang Zsolt Dimenziók a biológiában Fehérjék (kb. 50 ezer különböző fehérje a szervezetben 21 féle aminosavból épül fel) Élő szervezetek
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
Sejtmag, magvacska magmembrán
 Sejtmag, magvacska magmembrán Láng Orsolya Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Kompartmentalizáció Prokaryóta Cytoplazma Eukaryóta Endomembrán Kromatin Plazma membrán Eredménye
Sejtmag, magvacska magmembrán Láng Orsolya Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Kompartmentalizáció Prokaryóta Cytoplazma Eukaryóta Endomembrán Kromatin Plazma membrán Eredménye
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
I. A sejttől a génekig
 Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
Gén A gének olyan nukleinsav-szakaszok a sejtek magjainak kromoszómáiban, melyek a szervezet működését és növekedését befolyásoló fehérjék szabályozásához és előállításához szükséges információkat tartalmazzák.
Epigenetikai szabályozás
 Epigenetikai szabályozás 1. A Transzkripció epigenetikai szabályozása Dia 1 A kromatin szerkezete A sejtciklus S fázisában egyetlen sejtben kb 4x1 méter hosszúságú DNS található 4x23 darabra vágva (4N
Epigenetikai szabályozás 1. A Transzkripció epigenetikai szabályozása Dia 1 A kromatin szerkezete A sejtciklus S fázisában egyetlen sejtben kb 4x1 méter hosszúságú DNS található 4x23 darabra vágva (4N
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások)
") 3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
3. Sejtalkotó molekulák III. Fehérjék, enzimműködés, fehérjeszintézis (transzkripció, transzláció, poszt szintetikus módosítások) 3.1 Fehérjék, enzimek A genetikai információ egyik fő manifesztálódása
Viselkedésélettan: tanulás és memória. Zachar Gergely Semmelweis Egyetem, Anatómiai Intézet
 Viselkedésélettan: tanulás és memória Zachar Gergely Semmelweis Egyetem, Anatómiai Intézet Tanulás: Egy rendszer viselkedésének adekvát, rövidebbhosszabb távú megváltozása külső és belső stimulus hatására
Viselkedésélettan: tanulás és memória Zachar Gergely Semmelweis Egyetem, Anatómiai Intézet Tanulás: Egy rendszer viselkedésének adekvát, rövidebbhosszabb távú megváltozása külső és belső stimulus hatására
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Epigenetika. szomatikus sejt (emlőszövet sejt) magjának enukleált (magjától megfosztott) petesejtbe ültetésével hozták létre
 magjának enukleált (magjától megfosztott) petesejtbe ültetésével hozták létre") Epigenetika Dolly a klónbirka szomatikus sejt (emlőszövet sejt) magjának enukleált (magjától megfosztott) petesejtbe ültetésével hozták létre A sejtdifferenció a genomi ekvivalencia (genomi megegyezőség)
Epigenetika Dolly a klónbirka szomatikus sejt (emlőszövet sejt) magjának enukleált (magjától megfosztott) petesejtbe ültetésével hozták létre A sejtdifferenció a genomi ekvivalencia (genomi megegyezőség)
2007/11/05 Molekuláris biológia előadások - Putnoky 1-1
 1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
1-1 Fehérje transzportmechanizmusok az eukariota sejtben: 1) transzmembrán transzport kitekert formában, egyedi fehérjék transzportja célzottan - citoszol ER, citoszol MT 2) póruson keresztüli transzport
A genetikai lelet értelmezése monogénes betegségekben
 A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
Degeneráció és regeneráció az idegrendszerben
 Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
Új szignalizációs utak a prodromális fázisban. Oláh Zita
 Új szignalizációs utak a prodromális fázisban Oláh Zita 2015.10.07 Prodromális fázis Prodromalis fázis: De mi történik?? Beta-amiloid: OK vagy OKOZAT? Beta-amiloid hogyan okozhat neurodegenerációt? Tau
Új szignalizációs utak a prodromális fázisban Oláh Zita 2015.10.07 Prodromális fázis Prodromalis fázis: De mi történik?? Beta-amiloid: OK vagy OKOZAT? Beta-amiloid hogyan okozhat neurodegenerációt? Tau
Egy vagy több nukleotid mutációja megváltoztathatja a fehérje szerkezetét és működését
 Egy vagy több nukleotid mutációja megváltoztathatja a fehérje szerkezetét és működését Mutáció: egy sejt genetikai anyagában létrejövő hirtelen véletlenszerű változás, aminek hatására az a sejt és az abból
Egy vagy több nukleotid mutációja megváltoztathatja a fehérje szerkezetét és működését Mutáció: egy sejt genetikai anyagában létrejövő hirtelen véletlenszerű változás, aminek hatására az a sejt és az abból
3. Főbb Jelutak. 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3.
 Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A tanulási és emlékezési zavarok pathofiziológiája. Szeged,
 A tanulási és emlékezési zavarok pathofiziológiája Szeged, 2015.09.09 Szerkezet, működés, információáramlás, memória, tanulás: 1. Neokortex 2. Limbikus rendszer Limbikus rendszer és a memória Paul Broca
A tanulási és emlékezési zavarok pathofiziológiája Szeged, 2015.09.09 Szerkezet, működés, információáramlás, memória, tanulás: 1. Neokortex 2. Limbikus rendszer Limbikus rendszer és a memória Paul Broca
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Az agy betegségeinek molekuláris biológiája. 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5.
 Az agy betegségeinek molekuláris biológiája 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5. Alzheimer kór 28 Prion betegség A prion betegség fertőző formáját nem egy genetikai
Az agy betegségeinek molekuláris biológiája 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5. Alzheimer kór 28 Prion betegség A prion betegség fertőző formáját nem egy genetikai
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
A jel-molekulák útja változó hosszúságú lehet. A jelátvitel. hírvivő molekula (messenger) elektromos formában kódolt információ
 elektromos formában kódolt információ") A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
Gerinces és növényi ortológ promóter adatbázisok fejlesztése és elemzése. Doktori (Ph.D.) értekezés. Készítette: Sebestyén Endre
 értekezés. Készítette: Sebestyén Endre") Gerinces és növényi ortológ promóter adatbázisok fejlesztése és elemzése Doktori (Ph.D.) értekezés Készítette: Sebestyén Endre Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola
Gerinces és növényi ortológ promóter adatbázisok fejlesztése és elemzése Doktori (Ph.D.) értekezés Készítette: Sebestyén Endre Eötvös Loránd Tudományegyetem Természettudományi Kar Biológia Doktori Iskola
3. Sejtalkotó molekulák III.
 3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
3. Sejtalkotó molekulák III. Fehérjék, fehérjeszintézis (transzkripció, transzláció, posztszintetikus módosítások). Enzimműködés 3.1 Fehérjék A genetikai információ egyik fő manifesztálódása Számos funkció
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA NOVEMBER
 Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterkézés megfeleltetése az Euróai Unió új társadalmi kihívásainak a écsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMO-4.1.2-08/1/A-2009-0011 Az orvosi biotechnológiai
Az orvosi biotechnológiai mesterkézés megfeleltetése az Euróai Unió új társadalmi kihívásainak a écsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMO-4.1.2-08/1/A-2009-0011 Az orvosi biotechnológiai
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Bevezetés Áttekintés
 Bevezetés Áttekintés Fogalmak Egy lymphocita életének áttekintése Human Ig-izotípusok morfológiája Human Ig-izotípusok funkciói Miért? Válasz: más Ig effektor funkciók C fixáció FcR Polymerizáció Izotípusváltás
Bevezetés Áttekintés Fogalmak Egy lymphocita életének áttekintése Human Ig-izotípusok morfológiája Human Ig-izotípusok funkciói Miért? Válasz: más Ig effektor funkciók C fixáció FcR Polymerizáció Izotípusváltás
A replikáció mechanizmusa
 Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
Az öröklődés molekuláris alapjai A DNS megkettőződése, a replikáció Szerk.: Vizkievicz András A DNS-molekula az élőlények örökítő anyaga, kódolt formában tartalmazza mindazon információkat, amelyek a sejt,
1b. Fehérje transzport
 1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301)
") Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Biokémia és molekuláris biológia I. kurzus (bb5t1301) Tematika 1 TEMATIKA Biokémia és molekuláris biológia IB kurzus (bb5t1301) 0. Bevezető A (a biokémiáról) (~40 perc: 1. heti előadás) A BIOkémia tárgya
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Asztrociták: a központi idegrendszer sokoldalú sejtjei. 2009.11.04. Dr Környei Zsuzsanna
 Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
Receptorok, szignáltranszdukció jelátviteli mechanizmusok
 Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Hamar Péter. RNS világ. Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. www.meetthescientist.hu 1 26
 Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
Hamar Péter RNS világ Lánczos Kornél Gimnázium, Székesfehérvár, 2014. október 21. 1 26 Főszereplők: DNS -> RNS -> fehérje A kód lefordítása Dezoxy-ribo-Nuklein-Sav: DNS az élet kódja megkettőződés (replikáció)
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei)
") Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
Antiszenz hatás és RNS interferencia (a génexpresszió befolyásolásának régi és legújabb lehetőségei) Az antiszenz elv története Reverz transzkripció replikáció transzkripció transzláció DNS DNS RNS Fehérje
A KOLESZTERIN SZERKEZETE. (koleszterin v. koleszterol)
") 19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
DOKTORI ÉRTEKEZÉS TÉZISEI. Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben.
 DOKTORI ÉRTEKEZÉS TÉZISEI Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben Tóth Mónika Témavezető: Dr. Bálint Éva SZTE TTIK Biológia Doktori Iskola
DOKTORI ÉRTEKEZÉS TÉZISEI Az MDR1 gén kromatin szerkezetének tanulmányozása gyógyszerérzékeny és gyógyszerrezisztens humán sejtekben Tóth Mónika Témavezető: Dr. Bálint Éva SZTE TTIK Biológia Doktori Iskola
Biológus MSc. Molekuláris biológiai alapismeretek
 Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Biológus MSc Molekuláris biológiai alapismeretek A nukleotidok építőkövei A nukleotidok szerkezete Nukleotid = N-tartalmú szerves bázis + pentóz + foszfát N-glikozidos kötés 5 1 4 2 3 (Foszfát)észter-kötés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
TDK lehetőségek az MTA TTK Enzimológiai Intézetben
 TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Kromoszómák, Gének centromer
 Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
Sejt - kölcsönhatások. az idegrendszerben és az immunrendszerben
 Sejt - kölcsönhatások az idegrendszerben és az immunrendszerben A sejttől a szervezetig A sejtek között, ill. a sejtek és környezetük közötti jelátviteli folyamatok összessége az a struktúrált kölcsönhatásrendszer,
Sejt - kölcsönhatások az idegrendszerben és az immunrendszerben A sejttől a szervezetig A sejtek között, ill. a sejtek és környezetük közötti jelátviteli folyamatok összessége az a struktúrált kölcsönhatásrendszer,
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
A sejtek közöti kommunikáció formái. BsC II. Sejtélettani alapok Dr. Fodor János
 A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
2006 1. Nemszinaptikus receptorok és szubmikronos Ca2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra.
 2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE
 ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE") Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A doktori értekezés tézisei. A növényi NRP fehérjék lehetséges szerepe a hiszton defoszforiláció szabályozásában, és a hőstressz válaszban.
 A doktori értekezés tézisei A növényi NRP fehérjék lehetséges szerepe a hiszton defoszforiláció szabályozásában, és a hőstressz válaszban. Bíró Judit Témavezető: Dr. Fehér Attila Magyar Tudományos Akadémia
A doktori értekezés tézisei A növényi NRP fehérjék lehetséges szerepe a hiszton defoszforiláció szabályozásában, és a hőstressz válaszban. Bíró Judit Témavezető: Dr. Fehér Attila Magyar Tudományos Akadémia
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
A biológiai mozgás molekuláris mechanizmusai
 BIOLÓGIAI MOZGÁSOK A biológiai mozgás molekuláris mechanizmusai Kollektív mozgás Szervezet mozgása ( Az évszázad ugrása ) Szerv mozgás BIOLÓGIAI MOZGÁSOK BIOLÓGIAI MOZGÁSOK Ritmusosan összehúzódó szívizomsejt
BIOLÓGIAI MOZGÁSOK A biológiai mozgás molekuláris mechanizmusai Kollektív mozgás Szervezet mozgása ( Az évszázad ugrása ) Szerv mozgás BIOLÓGIAI MOZGÁSOK BIOLÓGIAI MOZGÁSOK Ritmusosan összehúzódó szívizomsejt
TRANSZPORTFOLYAMATOK 1b. Fehérjék. 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS
 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
Génexpresszió prokariótákban 1
 β-galaktozidáz-szint laktóz elfogy a laktóz Génexpresszió prokariótákban 1 14. A GÉNEXPRESSZIÓ SZABÁ- LYOZÁSA PROKARIÓTÁKBAN Enzimindukció, indukálható operon. Policisztronos. Katabolit represszió, represszálható
β-galaktozidáz-szint laktóz elfogy a laktóz Génexpresszió prokariótákban 1 14. A GÉNEXPRESSZIÓ SZABÁ- LYOZÁSA PROKARIÓTÁKBAN Enzimindukció, indukálható operon. Policisztronos. Katabolit represszió, represszálható
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Korai stressz és sérülékenység. Tóth Máté, PhD MTA KOKI Magatartás Neurobiológia Osztály
 Korai stressz és sérülékenység Tóth Máté, PhD MTA KOKI Magatartás Neurobiológia Osztály Korai stressz és sérülékenység + = Biológiai dinamika folyamatos kölcsöhatásban akörnyezettel Pszichiátriai zavarok
Korai stressz és sérülékenység Tóth Máté, PhD MTA KOKI Magatartás Neurobiológia Osztály Korai stressz és sérülékenység + = Biológiai dinamika folyamatos kölcsöhatásban akörnyezettel Pszichiátriai zavarok
A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban
 A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban Doktori (PhD) értekezés Siklódi Erika Rozália Biológia Doktori Iskola Iskolavezető: Prof. Erdei Anna, tanszékvezető egyetemi
A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban Doktori (PhD) értekezés Siklódi Erika Rozália Biológia Doktori Iskola Iskolavezető: Prof. Erdei Anna, tanszékvezető egyetemi
Semmelweis Egyetem / Élettani Intézet / Budapest. Bioinformatika és genomanalízis az orvostudományban. Szekvenciaelemzés. Cserző Miklós 2017
 Bioinformatika és genomanalízis az orvostudományban Szekvenciaelemzés Cserző Miklós 2017 A mai előadás Szekvencia analízis statisztikus szempontból Annotálás homológia alapján Az annotálás szempontjai
Bioinformatika és genomanalízis az orvostudományban Szekvenciaelemzés Cserző Miklós 2017 A mai előadás Szekvencia analízis statisztikus szempontból Annotálás homológia alapján Az annotálás szempontjai
Hemoglobin - myoglobin. Konzultációs e-tananyag Szikla Károly
 Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere
 I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere Kopper László A fejezet célja, hogy megismerje a hallgató a daganatok kialakulásában szerepet
I./1. fejezet: Jelátviteli utak szerepe a daganatok kialakulásában A daganatkeletkezés molekuláris háttere Kopper László A fejezet célja, hogy megismerje a hallgató a daganatok kialakulásában szerepet