SALSOLINOL: A PROLAKTIN ELVÁLASZTÁS ÚJ REGULÁTORA ÉS LEHETSÉGES HATÁSMECHANIZMUSA. dr. Radnai Balázs doktori (Ph.D.) értekezése
|
|
|
- Norbert Szőke
- 6 évvel ezelőtt
- Látták:
Átírás
1 HO HO SALSOLINOL: A PROLAKTIN ELVÁLASZTÁS ÚJ CH 3 NH REGULÁTORA ÉS LEHETSÉGES HATÁSMECHANIZMUSA dr. Radnai Balázs doktori (Ph.D.) értekezése Témavezető: Prof. Dr. Nagy György Semmelweis Egyetem Humánmorfológiai és Fejlődésbiológiai Intézet Sejt- és Molekuláris Neuroendokrinológiai Laboratórium Semmelweis Egyetem Idegtudományok Doktori Iskola (vezető: Prof. Dr. Réthelyi Miklós) Neuroendokrinológia programja (vezető: Prof. Dr. Halász Béla) Budapest, 2004
2 Édesanyám emlékére
3 A DOLGOZATBAN HASZNÁLT RÖVIDÍTÉSEK JEGYZÉKE: AL hypophysis mellső lebeny (anterior lobe) c koncentráció (concentration) camp ciklikus adenozin-monofoszfát CL kapilláris hurok (capillary loop) DA Dopamin DAerg Dopaminerg DAT Dopamin transzpoter DOPA 3,4- Dihidroxifenilalanin EM eminentia mediana ICV intracerebroventricularis IL hypophysis közti lebenye (intermediate lobe) IZ-AL Hypophysis mellső lebenyének belső zónája (Internal Zone of Anterior Lobe) LPV hosszú portális vénák (long portal vein) LTC Laktotrop sejtek (lactrotroph cells) NIL hypophysis közti-hátsó lebeny (neuro-intrmediate lobe) NL hypophysis hátsó lebeny (neural lobe) NSB nem specifikus kötés (non specific bound) OZ-AL Hypophysis mellső lebenyének külső zónája (Outer Zone of Anterior Lobe) PHDA periventriculo-hypophysealis dopaminerg rendszer PIF prolaktin elválasztást gátló PRF PRL faktorok (prolactin inhibiting factor) prolaktin elválasztást serkentő faktor (prolactin releasing factor) Prolaktin SB Specifikus kötés (specific SEM SPV bound) Az átlag hibája (standard error of mean) Rövid portális vénák (short portal vein) THDA Tubero-hypophysealis dopaminerg rendszer TIDA Tubero-infundibularis dopaminerg rendszer VMAT Vezikuláris monoamin transzporter - 3 -
4 Technikai megjegyzések: - A dolgozat megírásakor az ORVOSI HELYESÍRÁSI SZÓTÁR (Akadémia kiadó, 1992) ajánlását vettem alapul. A szabályosként elfogadott esetekben az eredeti terminológiát igyekeztem alkalmazni. - A rövidítések jegyzékében csak a gyakran előforduló, rendszeresen használt rövidítések találhatók, egyéb esetben a rövidítés jelentésének feloldására az első említéskor kerül sor, zárójelben jelölve azt. - Az irodalmi hivatkozások az első előfordulás szerinti növekvő számozással kerültek jelzésre, így az irodalomjegyzék is előfordulás szerinti, és nem alfabetikus sorrendben olvasható. - A szövegtörzs Times New Roman karakterkészlet 12 pontos méretében, míg a tartalomjegyzék 10 pontos méretben íródtak. - A számszerű eredményeket ismertető táblázatok sorszámozás nélkül kerültek ismertetésre, tekintettel arra, hogy azokra többszöri hivatkozás nem történik. - Statisztikai eredmények ismertetésekor az átlag ± átlag szórása formátumot alkalmaztam. Az ettől való eltérést külön jeleztem. - Az eredmények ismertetésekor a diszkusszió alapjául szolgáló következtetéseket a bekezdés melletti függőleges piros vonallal jelöltem. - Az értekezésben saját kísérleteimet ismertettem, így a diszkusszió alapjául is ezek szolgálnak. Munkacsoportunk salsolinollal kapcsolatos kutatási eredményeit a komplex kép kialakulása céljából célszerűnek láttam az értekezés végén, a VII. fejezetben külön alfejezetként tárgyalni. Ezen eredmények döntő része az egész munkacsoport összehangolt munkájának eredménye, a saját vizsgálatokat vastag szedéssel jelöltem
5 TARTALOMJEGYZÉK ÁBRÁK JEGYZÉKE ÉS MAGYARÁZATA 7 TÁBLÁZATOK JEGYZÉKE 8 ÖSSZEFOGLALÁS 9 SUMMARY 11 I. BEVEZETÉS 13 II. IRODALMI ÁTTEKINTÉS A prolaktin A prolaktin kémiai szerkezete, szintézise A PRL receptor A prolaktin biológiai hatásai Prolaktin és laktáció A prolaktin egyéb biológiai hatásai A prolaktin elválasztás szabályozása A prolaktin elválasztást gátló tényezők, hatásmechanizmusok Dopamin A dopamin anyagcseréje és hatásmechanizmusa A dopamin kémiai szerkezete, bioszintézise, eliminációja dopamin receptorok A dopamin szerepe a neuroendokrinológiai szabályozó mechanizmusokban Neuroendokrin dopaminerg pályarendszerek és azok szerepe a prolaktin elválasztás szabályozásában További prolaktin inhibitorok A prolaktin elválasztást fokozó ismert faktorok, azok hatásmechanizmusa A hypophysis közti-hátsó lebenyének szerepe a prolaktin elválasztás szabályozásában A közti lebeny szerepe A hátsó lebeny szerepe A salsolinol A salsolinol kémiai szerkezet, szintézisének módjai A salsolinol hatása a prolaktin elválasztásra A salsolinol és származékainak egyéb ismert hatásai 48 III. CÉLKITŰZÉSEK 49 IV. MÓDSZEREK Kísérleti állatok és műtéti beavatkozások Alkalmazott módszerek Receptor binding assay Kísérleti preparátumok A mért adatok feldolgozása Scatchard analízis Hill plot módszer Dopamin ( 3 H-spiperone) receptor binding assay H-salsolinol receptor binding assay Direkt leszorításos vizsgálatok Direkt leszorításos vizsgálatok D 2 receptor kötési helyéről Direkt leszorításos vizsgálatok salsolinol kötési helyről Intracelluláris camp szint meghatározása
6 3. Statisztikai analízis 59 V. EREDMÉNYEK Salsolinol hatása a dopamin D 2 receptorra Salsolinol specifikus kötőhelyeinek vizsgálata 3 H-salsolinol receptor binding assay módszerrel Salsolinol kötőhelyek a hypophysisben Salsolinol kötőhelyek a hypothalamo-hypophysealis rendszerben laktáló és szoptató állatokban Salsolinol kötőhelyek egyéb agyi régiókban H-salsolinol leszoríthatósága kötőhelyéről homológ leszorítási vizsgálatokkal Szoptatás hatása az intracelluláris camp szintre Salsolinol kezelés hatása az intracelluláris camp szintre 74 VI. DISZKUSSZIÓ A salsolinol specifikus kötőhelye(i) A salsolinol hatása az intracelluláris camp szintre A salsolinol szerepe a szoptatás kiváltotta PRL szint emelkedésben 83 VII. KÖVETKEZTETÉSEK Következtetések A salsolinol PRL elválasztást serkentő hatásának összefoglalása 87 VIII. KÖSZÖNETNYILVÁNÍTÁS 89 IX. IRODALOMJEGYZÉK
7 ÁBRÁK JEGYZÉKE ÉS MAGYARÁZATA 1. ábra A prolaktin előfordulása a szervezetben (bal oldalon a piros színnel a szervek, zöld színnel a szekretáló sejtek, jobb oldalon az elválasztott testnedvek feltüntetésével) ábra A prolaktin térbeli modellje (balra) valamint a prolaktin-receptor komplex térbeli elrendeződése (jobbra, a prolaktin molekula sárga színnel kiemelve). A modellek a Cn3D program 4.1-es verziójával készültek. (Forrás: 3. ábra A prolaktin receptor izoformái és jellemzőik. Módosítva, átvéve (26) ábra A prolaktin receptorai, szignáltranszdukciós mechanizmusai. Módosítva, átvéve (25) ábra A tuberoinfundibularis dopaminerg rendszert befolyásoló hormonok és neurotranszmitterek (bal oldalon a gátló, jobb oldalon a serkentő hatásúak). Módosítva, átvéve (1) ábra A dopamin szintézisének mechanizmusa. A tirozin aktív transzporttal jut a neuron citoplazmájába (1). A tirozin-hidroxiláz enzim (2) L-DOPA szintézisét segíti, mely a DOPA-dekarboxiláz enzim (3) katalízise mellett dopaminná alakul. A dopamin ezután a vezikuláris monoamin transzporter (VMAT) segítségével a szekréciós granulumba kerül (4), ahol tárolódik. Membránfúziót követően (5) ürül az extracelluláris térbe (szinaptikus résbe vagy neuroszekréció esetén az érpályába). A receptorhoz (6) kötődve kifejti hatását, vagy a dopamin-transzporter segítségével (7) visszajut a neuronba. Itt visszajuthat a szinaptikus vezikulumba (8), vagy a mitochondrium külső membránjához asszociált monoamin-oxidáz (MAO) enzim hatására inaktív metabolittá alakul (9). A metabolizmus másik útja a katekolamin-o-metil-transzferáz (COMT) által katalizált reakció (10). Módosítva, átvéve (49) ábra A dopamin eliminációja ábra Dopamin receptorcsaládok és transzporterek szerkezeti tulajdonságai ábra Neuroendokrin dopaminerg pályarendszerek ábra A dopaminerg pályarendszerekre ható modulációs lehetőségek. Módosítva, átvéve (1) ábra A salsolinol szerkezete ábra A dopamin salsolinol szintézisével kiegészített metabolizációs lehetőségei (a salsolinol szintézise pirossal kiemelve) ábra A Scatchard egyenes, melynek meredeksége -1/K D, (1) tengely metszete a B MAX ábra Hill Plot transzformáció. Magyarázat a szövegben ábra: A salsolinol D 2 -es receptorhoz kötődésének jellemzői: kötési kapacitás, affinitás és leszorítás. (A) A D 2 receptor antagonista haloperidol és 10-6 M koncentrációjú salsolinol leszorítási kísérlete D 2 dopamin receptorról. (B) 3 H-Spiperone binding assay: - 7 -
8 salsolinol hatása a receptorkötésre a Scatchard egyenes feltüntetésével. Magyarázat a szövegben ábra 3 H-salsolinol szaturációs görbéje laktáló patkány hypophysisének elülső (A) és közti-hátsó lebenyben (B). A három független kísérletből álló sorozat egyik reprezentatív eredménye. Magyarázat a szövegben ábra 3 H-salsolinol receptor binding assay hypophysis közti-hátsó lebenyén (A) valamint az eminentia mediana (B) területén laktáló és szoptató állatok esetében. Három független kísérlet egyik reprezentatív eredménye. Magyarázat a szövegben ábra Salsolinol leszorítása kötőhelyéről ábra Intracelluláris camp szint patkány hypophysis elülső lebenyének belső (IZ-AL) és külső (OZ-AL) zónájában kontroll állapotban, valamint 10 és 30 percig tartó szoptatást követően. Három független kísérlet eredménye ábra Intracelluláris camp szint patkány hypophysis közti- (IL) és hátsó (NL) lebenyében kontroll állapotban, valamint 10 és 30 percig tartó szoptatást követően. Három független kísérlet eredménye ábra Salsolinol kezelés hatása az eminentia mediana camp koncentrációjára. Magyarázat a szövegben. Három független kísérlet eredménye ábra Salsolinol kezelés hatása hypophysis közti-hátsó, valamint elülső lebenyének camp koncentrációjára. Magyarázat a szövegben. Három független kísérlet eredménye ábra A salsolinol lehetséges támadáspontjai és hatásmechanizmusa. A feltételezett gátló hatások piros, a serkentők kék nyíllal jelölve. Magyarázat a szövegben ábra Hasonlósági szempontok az noradrenalin-adrenalin és a dopamin-salsolinol rendszer között. Magyarázat a szövegben TÁBLÁZATOK JEGYZÉKE 1. táblázat A neuroendokrin szabályozásban szerepet játszó dopamintartalmú sejtcsoportok elhelyezkedése és az általuk alkotott pályarendszerek táblázat 3 H-salsolinol receptor binding assay módszerével mért receptor-ligand kötődési paraméterek különböző agyi struktúrákban táblázat 3 H-salsolinol leszorítása specifikus kötőhelyéről különböző anyagokkal, a leszorítás paramétereinek feltüntetésével táblázat Eddigi leszorítási kísérleteinkben megvizsgált anyagok, és azok hatása. Bal oldalon piros mezőben a leszorító tulajdonságúak, jobb oldalon kék mezőben a leszorítást nem eredményezők
9 ÖSSZEFOGLALÁS A prolaktin egy polipeptid szerkezetű hormon, mely a hypophysis mellső lebenyének mammotrop sejtjeiben képződik. Legismertebb, reprodukcióban betöltött szerepén túl a szervezet általános homeosztázisának fenntartásában is szerepet játszik. Szabályozásában eddigi ismereteink szerint- döntően a medialis-bazális hypothalamus területéről származó, tónusos gátló befolyás érvényesül. Az eddig publikált kísérletes eredmények alátámasztani látszottak azt a nézetet, mely szerint a DA tekinthető a prolaktin elválasztás domináns, gátló szabályozójának. A DA a mellső lebenyt a portális keringési rendszeren keresztül elérve tónusos gátló hatást gyakorol a laktotrop sejtek prolaktin elválasztására. Emlősökben bizonyos külső ingerek hatására (például laktáló állatokban az emlőbimbó ingerlésekor) a plazma prolaktin koncentrációja megemelkedik. Ez a változás bekövetkezhet a hypothalamikus DAerg gátló tónus csökkenésén keresztül és/vagy egy eddig ismeretlen hypothalamikus prolaktin releasing hormon (PRF) hatására. Az egyes fiziológiás állapotokban megfigyelhető gyors és átmeneti plazma prolaktin szintemelkedés nem magyarázható kizárólagosan a hypothalamicus DAerg gátlás csökkenésével, annál is inkább, mivel a legtöbb esetben a prolaktin szint növekedését csak kis mértékben és csupán egy nagyon rövid idejű (5-10 perc) hypothalamicus DA szint csökkenés kísér. Mindezek alapján már rég felmerült, hogy létezhet egy a prolaktin ürítést serkentő hypophyseotrop faktor, ami esetleg egy, a DA elülső lebenyi receptorain ható modulátor anyag is lehet. Ezek a feltételezett, de az eddig nem azonosított faktor(ok) megváltoztathatják a DA receptoriális kötését és/vagy a receptorok intracelluláris szignáltranszdukciós rendszerének működését. A közti-hátsó lebeny sebészeti eltávolítása illetve denervációja hatására a különböző ingerek, így a szoptatás hatására bekövetkező prolaktin szint emelkedés jelentősen csökken vagy elmarad. Ez a megfigyelés arra utal, hogy a prolaktin elválasztását serkentő anyag a hypophysis közti-hátsó lebenyében található. Ezt az elképzelést támasztja alá, hogy a közti-hátsó lebeny perklórsavas kivonata in vitro és in vivo körülmények között is serkenteni képes a prolaktin szekréciót. A próbálkozások ellenére a kivonatból eddig ezt az anyagot nem sikerült izolálni
10 A hypophysis közti-hátsó lebenyének perklórsavas kivonatában talált prolaktinreleasing aktivitással (PRL-RA) rendelkező anyag feltételezésünk szerint a fenti mechanizmusok egyikével képes fokozni a laktotrop sejtek prolaktin ürítését. Munkacsoportunknak a fent említett perklórsavas kivonatban HPLC módszer segítségével azonosított egy in vitro és in vivo is PRL-RA-al bíró anyagot, amely meglepő módon a DA-nak egyik katabolikus köztiterméke, az (R)-salsolinol. Az értekezés alapjául szolgáló kísérletekkel igazoltuk, hogy a salsolinol hatása nem a DA D 2 receptorának ligand kötő helyén létrejövő kompetició következménye. Bizonyítottuk, hogy in vivo salsolinol kezelés hatására az intracelluláris camp szint szignifikánsan változik, valamint azt, hogy ez egy salsolinol-membránreceptor kötődés következménye lehet. E receptort eddig nem sikerült azonosítani, de kísérleteink során, közvetett módon az általunk kidolgozott 3 H-salsolinol binding assay módszerével igazoltuk létezését. További specifikus és telíthető salsolinol kötőhelyeket találtunk a striatum, a cortex, az eminentia mediana és a hypothalamus területén. A K D értéke nanomoláris tartományban volt mérhető. Vizsgáltuk a 3 H-salsolinol leszoríthatóságát kötőhelyéről dopaminnal és származékaival, valamint más ligandumokkal is. Eredményeink alapján valószínűnek tűnik, hogy a salsolinol befolyásolhatja a neuroendokrin DAerg rendszert, megváltoztatva annak intracelluláris képződését illetve ürítését. Eredményeink bizonyítják, hogy a salsolinol intravénás alkalmazását követő 10 perccel, a plazma prolaktin szint emelkedésével párhuzamosan, szignifikánsan emelkedik a mellső lebenyi intracelluláris camp szint
11 SUMMARY Prolactin is a polypeptide hormone that is synthesized in and secreted from mammotropes of the anterior lobe of the pituitary gland. This hormone not only sub serves multiple roles during reproduction but it also plays an essential role in the general homeostasis of the organism. The secretion of prolactin from mammotropes is under a dominant inhibitory control that originates in the medial-basal hypothalamus. The bulk of previous and recent evidence support the view that dopamine is the neurohormone which is the physiological prolactin-inhibiting hormone. Dopamine is delivered to the anterior lobe through the vascular connection between the hypothalamus and the pituitary gland and maintains mammotropes in their tonically suppressed secretory state. In mammals, however, several exteroceptive stimuli such as suckling of the nipples of lactating mothers by their litters sharply elevate plasma prolactin. These stimuli may act by decreasing the inhibitory influence of hypothalamic dopamine and/or enhancing the activity of unknown hypothalamic neurons that secrete a prolactin-releasing hormone (PRF). Surgical removal or denervation of the neurointermediate lobe of the pituitary gland blocks or attenuates the secretory bursts of prolactin induced by different physiological stimuli including suckling. These observations have suggested strongly that a PRF may exist in the neuro-intermediate lobe. Indeed, it has been also shown that perchloric acid extracts of the neurointermediate lobe can stimulate the release of prolactin both in vitro and in vivo. In spite of relentless efforts to isolate and identify this PRF from neurointermediate lobe, the identity of this substance has remained elusive. Analyzed the perchloric acid extract of the neuro-intermediate lobe and of the median eminence, a dopamine-derived compound, R-salsolinol (SAL), has been detected in high concentration that has a selective and dose-dependent prolactinreleasing activity both in vivo and in vitro. Moreover, the concentration of SAL in neuro-intermediate lobe extracts varies in parallel with the suckling-induced prolactin secretion and is markedly reduced following disruption of dopaminergic innervation of the neuro-intermediate lobe. Lack of interference of SAL with 3 H-spiperone binding to AP homogenates indicates that SAL does not act at the dopamine D 2 receptor. Moreover, 3 H-SAL binds
12 specifically to homogenate of AL as well as neuro-intermediate lobe obtained from lactating rats. These data clearly indicate that SAL does not act at the dopamine (DA) D 2 receptors, and suggest that SAL supposedly has a binding site through which the secretion of PRL may be affected. Therefore, binding of 3 H-SAL to different regions of the central nervous system (CNS) has been investigated. Specific and saturable binding have been detected in the striatum, cortex, median eminence and in the hypothalamus as well as in the AL and the neuro-intermediate lobe (NIL) of the pituitary gland. K D values of the bindings were in the nanomolar range in all tissue tested. 3 H-SAL displacing activity of several agonists and antagonists of known DA receptors have also been tested. Salsolinol may regulate DAergic neurotransmission of hypothalamic neuroendocrine dopaminergic (NEDA) system by an altered intracellular or intraterminal synthesis and/or distribution of hypophysiotropic DA. Our findings show that 10 minutes after SAL injection, parallel to the rapid and marked elevation in plasma PRL, there is a significant increase in camp concentration of the AL. However, we assume that this rapid increase at the level of camp of the AL is related to the initiation of the secretory burst of pituitary mammotropes caused by SAL
13 I. BEVEZETÉS Eltérően a többi hypophysealis hormontól eddigi ismereteink szerint a prolaktin szekréció szabályozásában egy tónusos gátló hatás játssza a meghatározó szerepet. Ezért a tónusos hypothalamikus gátlásért a tubero-infundibuláris DAerg (TIDA) neuroncsoportból az eminentia mediana területén a hosszú portális erekbe ürülő dopamin (DA) tehető felelőssé. A TIDA rendszer mellett a hypothalamikus DAerg (DAerg) neuronoknak további két csoportja ismert. Az egyik rendszer idegsejtjei a nucleus arcuatusban a TIDA neuronoktól rosztrálisan helyezkednek el, axonjai a hypophysis közti és hátsó lebenyében végződnek. E DAerg neuronok képezik a hypothalamus ozmoszenzitív, úgynevezett tuberohypophysealis DAerg rendszerét (THDA). A nucleus arcuatus legrosztrálisabb része a periventriculárisan elhelyezkedő neuronokkal együtt pedig kizárólag a közti lebenyben végződő, az úgynevezett periventriculo-hypophysealis (PHDA) neuronális rendszert alkotja. Az eddig ismert irodalmi adatok alapján, a prolaktin elválasztás hypothalamikus szabályozása szempontjából a prolaktin szint emelkedését alapvetően egyrészt a tónusos gátló hatás csökkenése, másfelől egy esetleges prolaktin releasing hormon serkentő hatása okozhatja. Az egyes fiziológiás állapotokban megfigyelhető gyors és átmeneti plazma prolaktin szint emelkedése nem magyarázható kizárólagosan a hypothalamikus DAerg gátlás csökkenésével, annál is inkább, mivel a legtöbb esetben a prolaktin szint növekedését csak kis mértékben, és csupán egy nagyon rövid ideig (5-10 perc) tartó hypothalamikus DA szint csökkenés kísér. Mindezek alapján már régóta felmerült, hogy létezhet egy a prolaktin ürítést serkentő hypophyseotrop faktor, ami esetleg egy, a DA elülső lebenyi receptorain ható modulátor anyag is lehet. Ezek a feltételezett de az eddig nem azonosított faktor(ok) megváltoztathatják a DA receptoriális kötését és/vagy a receptorok intracelluláris szignáltranszdukciós rendszerének működését, esetlegesen a sejten belüli másodlagos hírvivő milieu -t, akár saját receptoriális hatásukon keresztül. A hypophysis közti-hátsó lebenyének perklórsavas kivonatában talált prolaktinreleasing aktivitással (PRL-RA) rendelkező anyag feltételezésünk szerint a fenti mechanizmusok egyikével képes fokozni a laktotrop sejtek prolaktin ürítését. Munkacsoportunk a fent említett perklórsavas kivonatban HPLC módszer segítségével
14 azonosított egy PRL-RA-al bíró anyagot, amely meglepő módon a DA metabolizmus egyik terméke, a salsolinol. Munkánkban a salsolinol hatásának tanulmányozása során választ kerestünk arra, hogy a salsolinol in vivo is szerepet játszik-e a PRL elválasztás szabályozásában, illetve ezt a hatást milyen mechanizmussal fejti ki. Fontos megválaszolandó kérdés volt annak tisztázása, hogy a salsolinol a már ismert DAerg aktivitás modulációjával éri-e el hatását (például a D 2 receptoriális kompeticióval), vagy esetleg létezik salsolinol kötőhely, mely kulcsszerepet játszhat a PRL elválasztás regulációjában. Dolgozatomban az e célból eddig végzett kísérletek eredményeit ismertetem
15 II. IRODALMI ÁTTEKINTÉS 1. A prolaktin A prolaktin (PRL) a tejelválasztásban betöltött szerepén túl a szervezet homeosztázisában is fontos szabályozó szerepet tölt be (1). Ez az összetett funkció, valamint a hormon elválasztásának szabályozásában rejlő, eddig megválaszolatlan kérdések napjainkban is számos munkacsoport kutatásai középpontjában állnak. A PRL legismertebb származási helye az agyalapi mirigy, ahol annak meghatározott sejttípusában, a laktotrop sejtekben termelődik. E hypophysealis PRL elválasztó neuronok egy része azonban olyan átmeneti sejtpopulációból is származhat, amelyik mind PRL-t mind növekedési hormont képes termelni (2). Ezek az úgynevezett somatomammotrop sejtek, melyek tehát kettős funkciójúak. További jellemzőjük, hogy az újszülött patkányok agyalapi mirigyében túlsúlyban vannak és ösztrogén hatására képesek laktotrop fenotípusú sejtekké differenciálódni (3). PRL immunreaktivitás a központi idegrendszer számos területén, sőt egyéb szövetekben is kimutatható (ld. 1. ábra), így a nagyagyban (agykéreg, hippocampus, amygdala, septum területein), az agytörzsben, a kisagyban, a gerincvelőben illetve a circumventricularis szervek területén (4, 5). Patkányban a hypothalamuson belül a dorsomedialis, a ventromedialis, supraoptikus és paraventricularis magokban is megtalálható a PRL (6). Hím állatokban a hypothalamikus PRL immunreaktivitás mennyisége a hypophysis műtéti eltávolítása után sem változik, míg nőstényekben csökken, jóllehet teljesen nem tűnik el. Ez a megfigyelés arra utal, hogy a hypothalamusban a PRL lokálisan, a hypophysistől függetlenül szintetizálódik (7), sőt az utóbbi idők vizsgálatai kimutatták, hogy a hypothalamikus és hypophysealis PRL elsődleges kémiai szerkezete megegyezik (8). A központi idegrendszerben termelődő PRL hatásának pontos mechanizmusa napjainkban még nem teljes mértékben ismert. A patkány placentában is termelődik számos PRL-szerű molekula, amelyek szerkezeti hasonlóságot mutatnak a hypophysealis PRL-nal. Ezek az úgynevezett placentális laktogének vagy PRL-szerű fehérjék. A decidua sejtekben is termelődik PRL, amely patkányban szerkezetileg egy kissé különbözik a hypophysisben termelődő
16 hormontól. Ez utóbbinak elválasztása érthető módon független a hypothalamikus PRL serkentő ill. gátló faktoroktól. A termelődő PRL az amnion folyadékba diffundálhat, és szerepet játszhat az embrionális ozmoregulációs és immunfolyamatokban. A méh szövetében, mégpedig a nem terhes patkányok myometriumában is kimutatható a PRL. Fiziológiás jelentősége ma még nem ismert (1). Laktáló emlősökben a PRL megtalálható az emlőmirigy epitheliális sejtjeiben, illetve az anyatejben is. A tejben található PRL egy része az agyalapi mirigyben termelődik és a vérkeringés útján az emlőmirigybe jut, ahol az epitheliális sejtek felveszik majd exocitózissal a mirigyvégkamrákba ürítik (9). Ugyanakkor az epithel sejtek önmagukban is képesek a PRL szintézisére (10). Az immunrendszerben a limfociták illetve a csecsemőmirigy valamint a lép immunkompetens sejtjei is képesek a hypophysealis PRL-hoz hasonló szerkezetű PRL szintézisére, melynek funkciója napjainkban is folyó, igen intenzív kutatás tárgya (1)
.")
17 1. ábra A prolaktin előfordulása a szervezetben (bal oldalon a piros színnel a szervek, zöld színnel a szekretáló sejtek, jobb oldalon az elválasztott testnedvek feltüntetésével)
18 1.1. A prolaktin kémiai szerkezete, szintézise A PRL (ld. 2. ábra) egyetlen, 199 aminosavat valamint három diszulfid-hidat tartalmazó láncból álló, 23,5 kda molekulasúlyú fehérje. Poszttranszlációs módosulások következtében különböző molekulasúlyú izoformák alakulnak ki, melyek receptor affinitása, biológiai hatása eltérő, az egyes izoformák aránya más lehet normális és patológiás körülmények között (11). Különbség mutatkozik az egyes sejttípusok által termelt PRL izoformák között is (12). A PRL a prolaktin/növekedési hormon/placentális laktogén családba tartozik, szerkezetét tekintve a citokin/hemopoetikus család tagjaival mutat rokonságot (13). 2. ábra A prolaktin térbeli modellje (balra) valamint a prolaktin-receptor komplex térbeli elrendeződése (jobbra, a prolaktin molekula sárga színnel kiemelve). A modellek a Cn3D program 4.1-es verziójával készültek. (Forrás: A PRL receptor A PRL receptora (PRL-R) a citokin/hemopoetikus receptor család tagja, mely családba többek között az interferon receptorok, az interleukin (IL) 2, IL-3, IL-4, IL-6,
19 granulocyta-makrofág kolónia-stimuláló faktor (GM-CSF), granulocyta kolóniastimuláló faktor (G-CSF), gp-130 (14, 15) sorolhatók. A PRL-R-nak többféle izoformája ismert, melyek alternatív hasítás útján jönnek létre (ld. 3. ábra). A hosszú, közepes és rövid forma közül az utóbbi nem képes a jelátvitelre az intracellulárs domén hiánya miatt (16). 3. ábra A prolaktin receptor izoformái és jellemzőik. Módosítva, átvéve (26) A jelátvitel A PRL receptorhoz történő kapcsolódása a receptor homodimerizációja után a Jak/STAT rendszer aktiválódását idézi elő (ld. 4. ábra), mely a sejt transzkripciós aktivitásának fokozódását eredményezi (17, 18). Az egyes sejteken a PRL más-más Stat fehérje aktiválódását indukálja. Nb2 sejteken a Stat1, Stat3 és Stat5-öt aktiválja (18). A jelátvitel a Ras, Raf/MAPK útvonalon is történhet (19). A PRL target génjei közül az interferon-regulatory factor-1 (IRF-1) az egyik legjelentősebb (20, 21). Aktiválódása T
.")
20 lymphocytákon például nélkülözhetetlen a sejtfejlődés, proliferáció, apoptózis szempontjából. A PRL által indukált Stat1 az IRF-1 gén promoterének pozitív, a Stat5 negatív mediátora: a két Stat faktor arányától függ tehát az IRF-gén aktivációjának fokozódása, vagy csökkenése (20, 22). A receptor stimuláció másik fontos eredménye a sejtnövekedés és differenciálódás szempontjából az ornitin dekarboxiláz jelentősen fokozott termelődése (23). A PRL nemcsak a sejtfelszíni receptorhoz történő kapcsolódás és jelátviteli mechanizmusok beindítása révén képes hatását kifejteni, hanem ismert az a tény is, hogy egyes sejtek internalizálni képesek a PRL-t. Az ilyen módon a sejtbe bejutott PRL a magba transzlokálódik (24). 4. ábra A prolaktin receptorai, szignáltranszdukciós mechanizmusai. Módosítva, átvéve (25)
21 1.3. A prolaktin biológiai hatásai Prolaktin és laktáció A PRL már az emlőmirigy növekedésében és fejlődésében, a tejtermelés megindításában és fenntartásában is alapvető szerepet játszik. Ezt bizonyítja, hogy PRL illetve PRL receptor génhiányos egerekben az emlőmirigy fejlődése abnormális, ezek az állatok tejtermelésre sem képesek (27). A terhesség alatti hypophysis eltávolítás után, mesterségesen előidézett PRL hiányos állapotban a tejtermelés nem indul meg, PRL kezelés azonban képes teljesen helyreállítani azt, bizonyítva ezzel a PRL szerepét. A tejtermelés során a PRL serkenti egyes aminosavak, valamint a glükóz táplálékként történő felvételét, a tejfehérjék (kazein és α-laktalbumin), a tejcukor és a tejben lévő zsírsavak termelését (28) A prolaktin egyéb biológiai hatásai A legtöbb rágcsálóban a PRL luteotrop hatása révén a párzás után hat napig fenntartja a sárgatest szerkezeti és funkcionális integritását (29). Patkány sárgatestében fokozza a progeszteron termelést és gátolja az enzimatikus inaktiválását (30). PRL-receptor hiányos egerek a sérült sárgatest funkciók és elégtelen petesejtek érési folyamatok miatt sterilek (25). A PRL-nak luteotrop funkciója mellett azonban luteolítikus hatása is van, amelynek mechanizmusa még nem pontosan ismert (31). Számos adat utal arra, hogy a PRL hozzájárul a szaporodási magatartásformák kialakulásához, amelyek azonban az alkalmazott modellek sokfélesége miatt még nem kellőképpen tisztázottak. DA antagonistával előidézett PRL szint emelkedésnek nincs hatása a párzási viselkedésre, ugyanakkor a szopás által indukált PRL elválasztás csökkenti a szexuális magatartást. Ezzel ellentétben a proösztrusz délutánján spontán
22 módon megemelkedő PRL koncentráció gátlása (DA agonistával) nagymértékben csökkenti a szexuális fogékonyságot (32). A PRL a szülés után meggyorsítja az anyai viselkedésformák megjelenését, bár önmagában nem vált ki anyai viselkedést. Ezt a hatását a hypothalamikus medialis preoptikus területen belül lévő idegi struktúrákra hatva fejtheti ki. A PRL a reprodukcióban betöltött funkciói mellett, szerepet játszik számos más, ugyancsak a szervezet belső egyensúlyának fenntartását szolgáló folyamatokban is. Részt vesz például a csontnövekedés, az immunrendszer, a só-vízháztartás és az érképződés szabályozásában (1)
 okozta")
23 2. A prolaktin elválasztás szabályozása A PRL szekrécióját a külső és belső környezet változásai folyamatosan befolyásolják, szabályozzák. A PRL szekréciót kiváltó fiziológiás ingerek közül a legfontosabbak és a legtöbbet tanulmányozottak a szopási inger, a stressz és a megemelkedett ovariális szteroidszint (főleg az ösztrogén) okozta PRL elválasztás fokozódás (33, 34, 35). Ezen ingerek hatásában (nagyság, időtartam) számos különbség mutatkozik, és jelentősen eltérnek a szervezet homeosztázisában játszott szerepük, illetve jelentőségük tekintetében is. Mai ismereteink szerint, ennek megfelelően a szabályozó mechanizmusok is több ponton különböznek. Közös bennük viszont az, hogy e stimulusok nagy része a hypothalamuson keresztül fejti ki hatását, ahol PRF-ek és PIFek felszabadulását befolyásolják. Emlősökben a hypophysis PRL szekréciójának hypothalamikus szabályozása dominánsan gátló jellegű (36). Ugyanakkor a hypothalamus részt vesz a PRL elválasztás serkentésének szabályozásában is, amely a gátló hatások kikapcsolásával és/vagy a beérkező stimuláló hatások felerősítésével érhető el (ld. 5. ábra) 5. ábra A tuberoinfundibularis DAerg rendszert befolyásoló hormonok és neurotranszmitterek (bal oldalon a gátló, jobb oldalon a serkentő hatásúak). Módosítva, átvéve (1)
24 2.1. A prolaktin elválasztást gátló tényezők, hatásmechanizmusok A ma általánosan elfogadott nézet szerint a laktotrop sejtek spontán és magas szekréciós aktivitással rendelkeznek, így a PRL elválasztás hypothalamikus tónusos gátló hatása elengedhetetlen feltétele az úgynevezett bazális PRL ürítésnek. Ezt a megállapítást több megfigyelés is alátámasztja. A hypophysis és a mediobasalis hypothalamus közötti kapcsolat sebészi megszüntetése után (nyélátvágással vagy az eminentia mediana sértésével) a plazma PRL koncentráció folyamatosan növekedve egy hét után maximális szintet ér el (37, 38). A PRL szekréció magas szintre áll be akkor is, ha a hypophysis elülső lebenyét olyan helyre transzplantáljuk, ahol nincs érrendszeri, illetve idegi összeköttetése a hypothalamusszal (pl. vesetok alá) (39). In vitro sejtkultúrában is igen magas az agyalapi mirigy sejtjeinek PRL elválasztása (34) Dopamin A DA-nak a PRL termelésre és elválasztásra kifejtett gátló hatását bizonyítja, hogy a katekolamin metabolizmust befolyásoló szerek a PRL szekréciót is jelentősen megváltoztatják (40) valamint, a DA magas koncentrációban való jelenléte az eminentia mediana területén, valamint a hosszú portális erekben (36, 41). Számos in vivo és in vitro kísérlet bizonyítja a DA-nak a PRL elválasztásra kifejtett gátló hatását (42, 43) A dopamin anyagcseréje és hatásmechanizmusa A dopamin kémiai szerkezete, bioszintézise, eliminációja A DA kémiai szerkezetét tekintve β-3,4-dihidroxifenil-etil-amin. Az 1950-es évekig a katekolamin szintézis egyik közti termékének tartották, később igazolták neurotranszmitter voltát. A DA bioszintézisének helyszíneit és folyamatát a 6. ábra mutatja. A szintézis sebességét meghatározó lépés az L-3,4-dihidroxifenilalanin (L- DOPA) képződése a neuronok által felvett tirozinból. Ezt a lépést a tirozin-hidroxiláz
25 (TH) enzim katalizálja (6. ábra 2-el jelölt enzim). Az enzim aktivitása többek közöttvégtermékei által szabályozott, így megvalósítva a feed-back mechanizmust. A keletkezett L-DOPA-t ezután az aromás aminosav dekarboxiláz enzim (6. ábra 3- al jelölt enzim) közreműködésével alakul DA-ná. A szintézis folyamatának részletei a 6. ábra magyarázatában olvasható. A TH immuncitokémiai vizsgálatokkal megállapították, hogy a TH immunpozitív neuronok valamivel nagyobb területen helyezkednek el, mint az a katekolamin fluoreszcencia alapján várható volt (44, 45). Az enzimreakciók során képződő aminok elleni antitestek használatával egyértelművé vált, hogy a DA és az L-DOPA immunreaktivitás eltérő eloszlást mutat, amelynek alapján feltételezhető egy olyan neuronális rendszer megléte, amely csak TH-t tartalmaz, de aromás aminosav dekarboxilázt nem. Ezek a neuronok így DA helyett csak L-DOPA termelésére képesek (46, 47, 48)
26 Tirozin L-DOPA Dopamin Dopaminerg idegvégződés Postszinaptikus neuron Intracelluláris jelátvitel D 2 R VMAT DAT Szekréciós granulum 6. ábra A dopamin szintézisének mechanizmusa. A tirozin aktív transzporttal jut a neuron citoplazmájába (1). A tirozin-hidroxiláz enzim (2) L-DOPA szintézisét segíti, mely a DOPA-dekarboxiláz enzim (3) katalízise mellett dopaminná alakul. A dopamin ezután a vezikuláris monoamin transzporter (VMAT) segítségével a szekréciós granulumba kerül (4), ahol tárolódik. Membránfúziót követően (5) ürül az extracelluláris térbe (szinaptikus résbe vagy neuroszekréció esetén az érpályába). A receptorhoz (6) kötődve kifejti hatását, vagy a dopamintranszporter segítségével (7) visszajut a neuronba. Itt visszajuthat a szinaptikus vezikulumba (8), vagy a mitochondrium külső membránjához asszociált monoamin-oxidáz (MAO) enzim hatására inaktív metabolittá alakul (9). A metabolizmus másik útja a katekolamin-o-metil-transzferáz (COMT) által katalizált reakció (10). Módosítva, átvéve (49)
27 Az axon-terminálisokból a szinaptikus résbe ürülő DA a jól karakterizált DA receptorokon fejtik ki hatását. A hatástartam regulációjában kulcsfontosságú a szinaptikus résben lévő transzmitter-koncentráció, illetve annak csökkentése. Ebben fontos szerepet játszanak a visszavételi, vagyis reuptake mechanizmusok, mely DA esetében leginkább a DA transzporteren (DAT, ld. 6. ábra 7-es számú képlete) keresztül valósul meg. Számos psychostimuláns drog, így a kokain és az amfetamin is ezt a transzportert képes gátolni, így megnövelve a DA mennyiségét a szinaptikus résben, ezáltal elnyújtott hatást kiváltva. A preszinaptikus neuronba visszajutva a DA a monoamin-oxidáz (MAO), az aldehid-dehidrogenáz (AD), valamint a katekol-o-metil-transzferáz (COMT) enzimek hatására alakul fő metabolitjaivá, vagyis dihidroxi-fenil-acetáttá (DOPAC), valamint homovanillinsavvá (HVA). A részletes folyamatot a 7. ábra mutatja. Az elimináció alternatív útját jelenti a salsolinol-szintetáz enzim, mely a salsolinol szintézisét katalizálja dopaminból és acetaldehidből (ld. 12. ábra, 46.oldal). A fentiek szerint MAO gátlás esetén a DA eliminációja döntően a salsolinol szintézis irányába folyik tovább, így a klasszikus metabolitok képződése helyett az (R)- salsolinol koncentrációjának emelkedését detektálhatjuk
28 3,4-dihidroxi-acetaldehid Dopamin 3,4-dihidroxifenilacetát (DOPAC) homovanillinsav 3-metoxi-tiramin 3-metoxi-4-hidroxi-fenilacetaldehid 7. ábra A dopamin eliminációja dopamin receptorok A szinaptikus résbe (illetve a neuroendokrin DAerg pályarendszerek esetében a portális erekbe illetve az intersticiális térbe) ürülő DA molekuláris biológiai módszerekkel jól identifikálható receptorokhoz kapcsolódik. A szerkezetük és az általuk aktivált másodlagos hírvivő mechanizmus alapján két fő csoportba (D 1 és D 2 - szerű DA receptorok), valamint több alcsoportba (D 1-5 ) sorolható receptorok (ld. 8. ábra) mindegyike a G fehérjéhez kapcsolt receptorcsaládba (GPCR) tartozik. Míg a D 1 receptorcsalád tagjai (D 1 és D 5 ) az intracelluláris camp szintet emelik, addig a D 2 receptorcsalád tagja (D 2, D 3, D 4 ) gátolják a camp képződését, vagyis az adenilát-cikláz enzim működését. A DA által kiváltott PRL szekréció gátlás a D 2 -es DAerg receptoron keresztül valósul meg

29 8. ábra Dopamin receptorcsaládok és transzporterek szerkezeti tulajdonságai A dopamin szerepe a neuroendokrinológiai szabályozó mechanizmusokban A központi idegrendszerben számos DA neurotranszmisszióval működő pályarendszer ismert. Ezek között említést érdemel a substantia nigra és a striatum közötti nigrostiatalis pálya, a mezolimbikus és a mezokortikális pályarendszerek. Ezek jórészt az extrapiramidális mozgató pályarendszerben (nigrostriatalis rendszer), másrészt az emocionális, kognitív magatartásformák kialakításában vesznek részt, nem rendelkeznek közvetlen hatással a neuroendokrin rendszerre. A neuroendokrin szabályozásban szerepet játszó DA tartalmú neuronok az alábbi magcsoportokban helyezkednek el (Dahlstorm és Fuxe-féle alfa-numerikus jelölés feltüntetésével, 50):
30 Magcsoport (alfanumerikus) Magcsoport (nomina anatomica) Célterület Pályarendszer A11 Posterior hypothalamus Gerincvelő Diencephalo-spinalis rendszer A12 Nucleus arcuatus Hypophysis közti és hátsó lebenye, Eminentia mediana Tubero-hypophysealis és tubero-infundibularis rendszer A13 Zona incerta Hypothalamus Incerto-hypothalamikus rendszer A14 Nucleus periventricularis Hypophysis közti lebeny Periventriculohypophysealis rendszer A15 Lateralis és ventralis hypothalamus?? 1. táblázat A neuroendokrin szabályozásban szerepet játszó dopamin tartalmú sejtcsoportok elhelyezkedése és az általuk alkotott pályarendszerek. Az A11-es és A15-ös rendszer funkciója nem tisztázott. Az A13-as DAerg pályarendszer a gonadotrop-serkentő hormon (GnRH) regulációjában játszik szerepet. A tubero-hypophysealis (THDA, A12), a tubero-infundibularis (TIDA, A12) valamint a periventriculo-hypophysealis (PHDA, A14) DAerg rendszer fontos szerepet játszik a PRL elválasztás szabályozásában, így a továbbiakban döntően ezekről esik szó Neuroendokrin dopaminerg pályarendszerek és azok szerepe a prolaktin elválasztás szabályozásában A DA receptorcsalád D 2 alosztályába tartozó receptorok megtalálhatóak az agyalapi mirigy sejtjeinek felszínén. (51, 52). Az agyalapi mirigy működését befolyásoló DAerg neuronok (1. táblázat) a hypothalamikus nucleus periventricularis (A14 katekolaminerg
31 sejtcsoport) és a nucleus arcuatus (A12 katekolaminerg sejtcsoport) területén helyezkednek el (53, 54, 55, 56). Ez a két neuronális sejtcsoport három, egymástól mind morfológiailag mind funkcionálisan elkülönülő rendszerre osztható (ld. 9. ábra). A periventriculo-hypophysealis DAerg (PHDA) neuronok a hypothalamus elülső periventricularis részében helyezkednek el és az agyalapi mirigy IL-ében végződnek. Ezek a neuronok a IL-ben termelődő α-melanocita-stimuláló hormon (α MSH) szabályozásában vesznek részt (53, 54). A második csoportba tartozó neuronok a nucleus arcuatus rostralis részében találhatóak, a PHDA rendszer hátsó részének szomszédságában, és mind a közti mind a hátsó lebenybe vetülnek (36). Ezek az idegsejtek alkotják az úgynevezett ozmoszenzitív tubero-hypophysealis DAerg rendszert (THDA) (57, 55, 58), amely szintén részt vesz a PRL elválasztás szabályozásában (59, 60, 61). Az A12 tubero-infundibularis DAerg (TIDA) rendszert alkotó neuronok a nucleus arcuatus középső és hátsó részében helyezkednek el, és az EM külső zónájába vetülnek. Innen a DA, bekerülve a hosszú portális erekbe, eléri az agyalapi mirigy sejtjeit. Az e sejtek által termelt DA a PRL általánosan elfogadott hypothalamikus szabályozója (36)

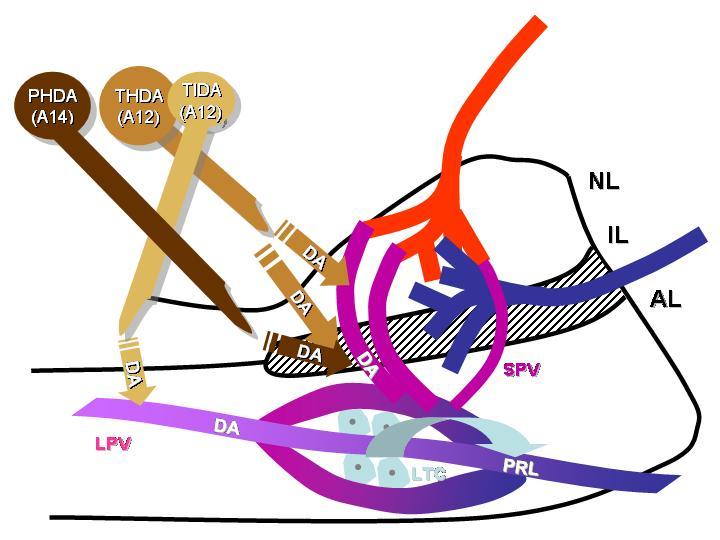
32 9. ábra Neuroendokrin dopaminerg pályarendszerek
33 Az immunhisztokémiai vizsgálatok egyértelműen bizonyították a TH és az aromás aminosav dekarboxiláz (AADC) enzim, valamint a DA jelenlétét is a PHDA, THDA és a TIDA neuronokban, de ez utóbbinak csupán a dorsomedialis részében (DM-TIDA). A DA transzporter mrns-t is csupán ezen a területen azonosították (62). Ezek az idegsejtek tehát az enzimtartalmuk alapján végtermékként dopamint képesek termelni. Ugyanakkor a TIDA neuron rendszer ventrolateralis részében (VL-TIDA) csak TH immunreaktivitás találtak, aromás aminosav dekarboxiláz aktivitást nem (184, 48, 63), L-DOPA azonban detektálható ezen a területen (48, 63). Ezek a VL-TIDA rendszerben elhelyezkedő sejtek tehát csak L-DOPA szintézisére képesek, amely az aromás aminosav dekarboxiláz hiányában nem alakulhat tovább dopaminná. Ezen megfigyelések alapján feltételezhető, hogy az L-DOPA, mint transzmitter végtermékként képződik, az eminentia medianaba ürül és a hosszú portális erekbe kerül, esetleg más idegsejtek felveszik, és további enzimatikus folyamatok szubsztrátjaként dopaminná (64), vagy más anyaggá alakul. Az arcuatus mag két részének eltérő funkcionális jelentőségére utal az a megfigyelés, hogy a DM-TIDA részt vesz álterhes nőstény patkányokban a PRL napi ritmusának szabályozásában, ugyanis az itt elhelyezkedő neuronok aktivitása csökkenést mutat a napi PRL csúcs kezdetekor, míg a VL-TIDA területén elhelyezkedő neuronokban hasonló változás nem figyelhető meg (65). A TIDA rendszer DAerg neuronjainak aktivitáscsökkenését eredményezi a laktáció (mely 24 órás elválasztás után visszaáll a korábbi mértékre), továbbá a szopási stimulus (66, 67, 68). A fenti, fiziológiai állapotokkal összefüggő különbségeken túl, nemi eltérések is tapasztalhatók a TIDA neuronok működésében. Nőstényekben a neuronok válaszkészsége a PRL feed-back iránt kifejezettebb (69, 70). A neuronok aktivitása kasztrációt követően nőstényekben csökken, míg hímekben növekszik (71). Hormonpótlással az előbb említett hatások kivédhetők (72). Hím és nőstény állatok stresszre adott eltérő válasza szintén szembetűnő: nőstényekben a TIDA aktivitás csökken, míg hímekben változatlan marad (73)
34 A TIDA rendszertől eltérően a THDA és a PHDA neuronok aktivitása független a keringő gonadalis szteroidoktól, illetve THDA neuronok aktivitásában nem tapasztalhatók nemi különbségek sem (74). A hypophysis NIL-ének műtéti eltávolítása illetve nyélnyomorítás után, amikor az AL vérellátása és így a TIDA rendszerből érkező DA mennyisége sem változik, a plazma PRL szint mégis 3-4 szeresére emelkedik mind hím, mind ciklusos és laktáló nőstény patkányokban, valamint ezzel párhuzamosan lecsökken a hypophysis AL DA tartalma. Ez a hatás tehát pusztán a THDA és PHDA rendszerek funkciócsökkenésével függ össze (75, 59, 76). A korábban vázolt DAerg sejtcsoportok és pályarendszerek közvetlen PRL szekréciót gátló hatásukon túl fontos csomópont szerepet töltenek be, amennyiben azok különböző környezeti tényezők és kémiai ágensek hatására bekövetkező aktivitásváltozása a PRL elválasztás befolyásolását eredményezik. Az említett hormonok hatásait foglalja össze a 10. ábra. A legfontosabb gátló tényező maga a PRL, míg a serkentő hatások közül ösztradiol emelendő ki
, így a portális erekben megemelkedő DA koncentráció a PRL elválasztásának gátlását eredményezi (77).")
35 10. ábra A dopaminerg pályarendszerekre ható modulációs lehetőségek. Módosítva, átvéve (1). A PRL, a hypothalamus szintjén feed-back mechanizmussal képes saját elválasztását befolyásolni. A plazma PRL szintjének emelkedése fokozza a hypothalamusban a DA szintézist (70), így a portális erekben megemelkedő DA koncentráció a PRL elválasztásának gátlását eredményezi (77). Újabb vizsgálatok PRL receptorokat mutattak ki a TIDA, PHDA és a THDA neuronok felszínén (78), amely a feed-back mechanizmus molekuláris biológiai alapjának bizonyítéka. A PRL mindhárom DAerg rendszerre serkentőleg hat (60). A laktáció középső szakaszában azonban nem működik a visszaható mechanizmus, a TIDA neuronok érzéketlenné válnak a megemelkedett mennyiségű PRL-ra. Ez a jelenség a korai laktációban nem tapasztalható, később pedig hosszú ideig tartó
36 elválasztás után megszűnik (79, 80). Érdekes megfigyelés, hogy a feed-back mechanizmus teljes hiánya esetén (például a PRL hiányos egerekben), a TIDA neuronok száma csökken, míg más hypothalamikus noradrenerg sejtcsoportban (A13, A14) nem tapasztalható sejtszám-csökkenés. Ezt a jelenséget így nem a mutáció elsődleges hatása magyarázza, hanem a PRL hiányra adott válasz, amely az egyedfejlődés korai szakaszában alkalmazott PRL kezeléssel megakadályozható (81, 82) További prolaktin inhibitorok Habár a DA PRL elválasztást gátló hatása teljes mértékben bizonyított, számos kísérletes megfigyelés támasztja alá azt, hogy önmagában nem elegendő a PRL szekréció teljes gátlásához, vagyis további PIF-ek létezése valószínűsíthető. Így például, míg hímekben a portális erekben 5-7-szer alacsonyabb a DA koncentráció, mint nőstényekben, a PRL szintekben nem tapasztalható ekkora különbség (36, 83). Laktáció során sem találtak teljesen ellentétes változásokat az EM illetve a portális erek DA tartalma és a plazma PRL koncentrációja között (84, 85). A klasszikus neurotranszmitterek közül az egyik lehetséges PIF az acetil-kolin. Szisztémás kolinerg aktiválás illetve acetil-kolin agonista icv adása után csökken a plazma PRL szintje. Kolinerg agonista adásával meggátolható a szopás illetve ösztradiol által indukált PRL elválasztás is (1). Az ugyancsak klasszikus neurotranszmitternek tekinthető noradrenalin is szóbajön, mint a PRL elválasztás egyik gátlója. A központi noradrenerg rendszer ugyanis valószínűleg α1 adrenerg receptorokon keresztül (86) tónusos gátló hatást fejt ki a bazális és ösztrogén indukált PRL elválasztásra (87). Másrészről viszont a proösztrusz délutánján megjelenő PRL csúcs illetve a stressz indukált PRL elválasztás gátolt a centrális noradrenerg pályák mechanikai vagy kémiai roncsolása után (88). További lehetséges PIF a γ-amino-vajsav (GABA), mely a mediobasalis hypothalamusban elhelyezkedő GABAerg neuronokból felszabadulva, az EM külső zónájában a hosszú portális erekbe ürülve eljut az hypophysis AL sejtjeihez (89, 90), ahol receptorain keresztül, direkten a laktotrop sejteken hatva gátolja a PRL elválasztást
37 (91, 92). További bizonyíték, hogy a PRF-ek által kifejtett hatások közvetítésében (43) szerepet játszó paraventricularis és supraopticus magvak gazdag GABAerg innervációban részesülnek. A hypothalamikus preoptikus és elülső periventricularis areában illetve a paraventricularis magban termelődő szomatosztatin in vitro körülmények között gátolja a bazális illetve a TRH vagy VIP által kiváltott PRL elválasztást (93). Az endothel sejtek által termelt vazoaktív peptidek közül az endothelin-1 és az endothelin-3 is képes in vitro a PRL szekréció dózisfüggő gátlására, amely hatás a D 2 receptor aktiválása mellett is fennmarad (43). A pankreász polipeptidek közé tartozó neuropeptid Y in vitro gátolja laktáló vagy ciklusos nőstény állatból származó hypophysis mellső lebeny sejtjeiből a PRL elválasztást, valamint az egyidejűleg adott DA gátló hatását is fokozza (94) A prolaktin elválasztást fokozó ismert faktorok, azok hatásmechanizmusa A rendelkezésre álló kísérletes adatok alapján általánosan elfogadott nézet, hogy a PRL elválasztás főképp a DA gátló hatása alatt áll, ugyanakkor ezzel párhuzamosan számos kutatás folyik a PRL elválasztást serkentő anyagok iránt. A PRF-ek létezésének feltételezése előtt a PRL szekréció fokozódására a hypothalamus tónusos gátló hatás megszűntét követő, spontán módon bekövetkező PRL szint emelkedést tartották domináns hatásnak (34). Ezt az elképzelést támasztotta alá, hogy a TH gátlásával felfüggesztett DA szekréció önmagában a PRL elválasztás fokozódásához vezetett. Ennek mértéke hasonló a szopási inger, illetve stressz esetén tapasztalthoz (41). In vitro kísérletben a DA infúzió felfüggesztése gyors PRL szekréciónövekedést eredményez. Az emlőideg elektromos ingerlése azonban csak egy néhány percig tartó DA ürítés csökkenést okoz, amit azután gyors pulzusokban történő DA elválasztás követ a szoptatás egész időtartama alatt. A szopási inger után a többszörös plazma PRL szint emelkedés mellett a portális vér DA tartalma csak kismértékben csökken (43, 34)
38 A fenti megfigyelések arra utalnak, hogy a DA szekréció csökkenése önmagában nem elégséges a szopásra adott PRL válasz létrejöttéhez. A jelenlegi eredmények alapján úgy tűnik, hogy ellentétben a többi adenohypophysealis hormonnal, nincs egy kitüntetett fő PRL serkentő hormon, hanem számos anyag rendelkezik PRL elválasztást fokozó hatással. A nucleus paraventricularis parvocelluláris neuronjaiban képződő, majd az EM területén a hosszú portális erekbe ürülő thyreotroph releasing hormont (TRH) eredetileg, az hypophysis AL sejtjeiben termelődő, thyroid serkentő hormon (TSH) elválasztását serkentő hypophyseotrop faktorként írták le (95, 96). A TRH in vitro és in vivo körülmények között is képes a PRL elválasztás dózisfüggő serkentésére (77, 97). PRF aktivitással rendelkezik a vazoaktív intesztinális peptid (VIP) is, amelynek a jelenléte a nucleus paraventricularisban és az eminentia medianaban is bizonyított (98, 99). A portális vérben megtalálható peptid a laktotrop sejteken található VIP receptorokon keresztül hatva képes a PRL elválasztás serkentésére (98, 99, 100). A VIP-pel homológiát mutató peptid hisztidin-izoleucin ugyancsak kimutatható a portális vérben (100), és szintén képes a PRL szekréciót serkentésére (101, 102). A több, mint egy évtizede juh hypothalamusból azonosított és izolált hypophysealis adenilát ciklázt aktiváló polipeptid (PACAP) a VIP neuropeptid család egyik tagja (103, 104). Két biológiailag aktív formája ismert: a 27 aminosavból álló PACAP-27, illetve a hosszabb PACAP-38. Mindkét peptid szoros homológiát mutat a VIP N- terminális aminosavszekvenciájával. Hatásukat tekintve nagymértékű ciklikus AMP (camp) felhalmozódást okoz tenyésztett ALi sejtekben, feltehetően a nagy affinitású receptorokon keresztül hatva (105). A hypophysealis portális érrendszerben a PACAP-38 koncentrációja magasabb, mint a szisztémás keringésben. Szisztémás PACAP-38 injekció szignifikánsan és dózisfüggően emeli a plazma PRL és növekedési hormon szinteket hím és elválasztott laktáló nőstényekben is, ugyanakkor folyamatosan szoptató anyákban nincs hatása a plazma PRL koncentrációjára (106, 107, 108). Laktálókban a PACAP-38 a szopás indukált PRL válaszhoz hasonló mértékű PRL elválasztást okoz. Hímekben a PACAP a hypothalamus léziója után is serkentette a PRL és a növekedési hormon elválasztását,
39 ami közvetlen hypophysealis hatásmechanizmust sejtet (109). Ezzel ellentétben, sejtkultúrában és reverz hemolítikus plakk assay-ben a PACAP-38 dózisfüggően gátolta a PRL leadást, míg a növekedési hormon elválasztást serkentette (109, 107, 108). Ez utóbbi megfigyelések arra utalnak, hogy a PACAP-38 nem közvetlenül a laktotrop sejteken hatva fejti ki a PRL serkentő hatását, hanem esetleg indirekten, egy más típusú hypophysealis sejten keresztül, parakrin módon. Ezek lehetnek a follikulus stimuláló hormont termelő (FSH) sejtek, amelyek szintén rendelkeznek PACAP kötőhellyel (110) illetve a melanotrop sejtek, amelyekben PACAP hatására megnő a camp koncentráció és az αmsh elválasztás (111). PACAP-38 kezeléskor megnőtt az interleukin-6 (IL-6) szekréció az FSH sejtekből, amely peptid képes in vitro a PRL elválasztás serkentésére. Így az IL-6 lehet egy, a PACAP serkentő hatását közvetítő, parakrin faktor (112, 113). A szerotonin (5HT) PRL elválasztás szabályozásában való részvételét is számos eredmény látszik alátámasztani. Centrálisan vagy perifériásan adott szerotonin illetve szerotonin prekurzor (5-hidroxi-triptofán, 5HTP) hatására emelkedik ugyanis a plazma PRL koncentrációja (114, 115, 116). Ez a hatás azonban csak intakt NIL és hypothalamus esetén figyelhető meg (117, 118, 119). Az 5HT receptor 1 agonisták in vivo is képesek a PRL szint emelésére (40, 120). A szerotonin szintézis gátlása intakt állatokban nem befolyásolta a PRL elválasztást, ugyanakkor laktáló nőstényekben teljesen megakadályozta a szopási inger által kiváltott PRL elválasztást, mely hatás kivédhető volt szopási stimulus előtt adott 5HTP-al (121). Bár szerotonin receptorokat kimutattak a hypophysis elülső lebenyében is, de a szerotonin in vitro nem serkenti a PRL szekréciót a laktotrop sejtekből (122, 123, 114, 124). Mindezen adatok arra utalnak, hogy a szerotoninerg pályáknak fontos szerepet játszanak a szopási stimulus közvetítésében, és így a szopás kiváltotta PRL válaszban, azonban a szerotonin nem neurohormonként közvetlenül, hanem indirekten, hypothalamikus szinten hatva befolyásolja a PRL szekréciót. Az angiotenzin II részt vehet a PRL fiziológiás szabályozásában, mind a hypothalamus (125, 126) mind a hypophysis szintjén (127). Centrális hatással a hisztamin is serkenti a PRL elválasztást (főképpen a stresszre adott PRL választ hím patkányokban) döntően H 2 receptorokon hatva (128, 129)
40 A P-anyag, a tachykinin peptidcsalád tagja, serkenti a PRL elválasztást központi és perifériás kezeléskor, illetve hypophysis sejtkultúrában is (130). Az endogén opioidok három, különböző gének által kódolt, peptidcsaládba tartoznak, ezek az endorfinok, dinorfinok és enkefalinok (131). Az enkefalin immunpozitív neuronok megtalálhatóak a hypothalamikus paraventricularis valamint supraopticus magban, ahonnan az eminentia medianaba és a hypophysis hátsó lebenyébe vetítenek. Laktáció alatt a TIDA neuronok is szintetizálnak enkefalint, így ciklusos nőstényhez képest az EM és a portális vér nagy mennyiségű enkefalint tartalmaz (132, 133). Az endogén opioidok, feltehetőleg µ és/vagy κ opiát receptoron keresztül (134), gátolják a TIDA neuronokat, így serkentik a PRL elválasztást (135, 136, 137). Hypophysis sejtkultúrában alkalmazva a galanin is képes a PRL elválasztást serkenteni (138). A galanint tartalmazó idegsejtek megtalálhatóak a mediobasalis hypothalamus területén, az EM külső rétegében pedig immunpozitív rostokat is találtak (139). A 13 aminosavból álló neurotenzin főleg a nucleus arcuatus neuronjaiban termelődik, ahonnan az eminentia medianaban ürülve eléri az AL sejtjeit. Sejtkultúrában illetve in vivo perifériásan alkalmazva serkenti, míg centrálisan csökkenti PRL ürítést és gátolja a stressz, illetve szerotonin kezelés okozta PRL elválasztást. (1, 140). A központi idegrendszeren belül megtalálható serkentő aminosavaknak is szerepet tulajdonítanak a PRL elválasztás szabályozásában. Ezek közül a nucleus arcuatus neuronjai felé szinte kizárólag a glutamát közvetíti a gyors serkentő impulzusokat. A nucleus paraventricularisban is gazdag glutamáterg rostvégződés található (141, 142). A glutamáterg receptorok megtalálhatóak a nucleus paraventricularis neuronjain, sőt az AL laktotrop sejtjein is kimutatták jelenlétét (143, 144). Érdemes megemlíteni, hogy a DA is képes a PRL szekréció növelésére, nagyon alacsony, pikomoláris koncentrációban, in vitro sejtkultúrában alkalmazva (145, 146, 147). Az állatok in vivo fiziológiás állapota meghatározhatja a laktotrop sejtek in vitro válaszkézségét a DA iránt. Így például egy rövid szopási ingert követően laktálókból
41 kivett (148) illetve ösztrogén és progeszteron-kezelt, ovariektomizált nőstény állatokból származó sejteken (149) a DA alacsony koncentrációban serkentőleg hat. Az elválasztott állatokból származó sejteknél a dopaminnak viszont csak a gátló hatása érvényesül (148, 150) A hypophysis közti-hátsó lebenyének szerepe a prolaktin elválasztás szabályozásában Számos eredmény támasztja alá, hogy a hypophysis közti és hátsó lebenye részt vesz laktotrop sejtek szabályozásában (45). Morfológiai sajátosságok alapján az itt termelődő anyagok, így az α-msh, oxitocin, vazopresszin, DA illetve eddig nem identifikált molekulák a rövid portális ereken keresztül elérhetik az AL sejtjeit, így részt vehetnek a PRL elválasztás szabályozásában. A hypophysis NIL-ének műtéti eltávolítása illetve denervációja megemelte a bazális PRL szintet nőstény és laktáló patkányban egyaránt (75, 76). Mivel a THDA és PHDA neuronok a NIL-ben végződnek, ezért kézenfekvőnek tűnt az innen felszabaduló DA valamint az oxitocin és vazopresszin hiányával magyarázni a fenti észleletet. Későbbi kísérletek azonban kimutatták, hogy ekkor a szopási stimulus által kiváltott PRL szekréció oxitocin és a vazopresszin egyidejű pótlása esetén is gátolt. A NIL perklórsavas kivonata mind in vivo mind in vitro serkentette a PRL elválasztást, amely hatás egy héttel a hypophysisnyél átmetszése után, a NIL és a hypothalamus közötti neurális kapcsolat megszűntével párhuzamosan (151, 152, 153, 154) megszűnt. Specifikus oxitocin receptor antagonista kezelés nem befolyásolta a NIL-i kivonat PRF aktivitását (153). Mindezekből arra következtettek, hogy a NIL tartalmaz eddig ismeretlen hypothalamikus PRF-et, és ez biztosan nem az oxitocin A közti lebeny szerepe
42 A IL melanotrop sejtjei által termelt α-msh szerepet játszik a laktotrop sejtek funkciójának szabályozásában. A IL-ből a rövid portális ereken keresztül az AL sejtjeihez jutva receptoraihoz kötődni képes. Ezek legnagyobb számban a laktotropokon találhatók (155). Valószínűleg azonban az α-msh nem rendelkezik direkt PRL serkentő hatással, hanem a responsiveness faktorok közé tartozik. In vitro csökkenti a laktotrop sejtek nagy dózisban adott DA gátló hatása iránti válaszkészséget, illetve növelni képes a TRH-ra, ANG-I-re és a serkentő hatású, alacsony dózisban adott dopaminra (148, 150) adott PRL-release választ. α-msh immunneutralizáció nagymértékben csökkentette a szoptatás alatti PRL elválasztás (156, 42, 148). Ezzel ellentétben szopási inger hatására nem változott sem a plazma, sem a hypophysis elülső lebenyének α-msh koncentrációja (157, 158). Ez utóbbi megfigyelés alapján nem valószínű, hogy az α-msh részt vesz a szopási ingert követő PRL szekrécióban A hátsó lebeny szerepe A NIL kivonat korábban említett PRL serkentő hatásáért a hypothalamikus oxitocin is felelős lehet (159). Az oxitocin koncentrációja a portális vérben ugyanis ször magasabb, mint a szisztémás keringésben, a nagy affinitású oxitocin receptorok pedig megtalálhatóak az elülső lebenyben (160, 161). A hypothalamus paraventricularis és a supraopticus magjainak magnocelluláris neuronjai által termelt oxitocin axontranszport révén a raktározás helyére, a hypophysis hátsó lebenyébe kerül. Az eminentia mediana-n keresztülhaladó axonok, szinaptikus buttonok kialakítása révén, kapcsolatba kerülnek a kapilláris hurkokkal, sőt az oxitocin a hosszú portális erekbe is ürülhet (160) Az oxitocin a hátsó lebenyből a rövid portális ereken keresztül is elérheti az AL sejtjeit. A szopásra adott PRL válaszban betöltött hypophysealis szerepét vizsgálva, munkacsoportunk megállapította hogy a szopási stimulus után nagymértékben megnőtt a IL-ben az oxitocin koncentráció, viszont az elülső lebenyben nem változott a
43 kiinduláskor is nagyon alacsony oxitocin szint (158). Az oxitocin fiziológiás szerepe a IL-ben még nem ismert, oxitocin receptor itt nem található (162). A nagy dózisban adott oxitocin fokozza a PRL szekréciót hím és ovariektomizált nőstény állatokban, de laktáló patkányokban csökkenti a szopásra adott PRL választ (163, 164). Az oxitocin antiszérummal végzett passzív immunizáció csökkenti a szopási inger vagy az ösztrogén által kiváltott PRL emelkedést (159). Ugyanakkor specifikus oxitocin antagonista beadásakor, mely gátolja a szopás által indukált tejbelövellést, nem változik a szopási ingert követő PRL emelkedés (165). Úgy tűnik tehát, hogy bizonyos fiziológiás állapotokban PRF szerep tulajdonítható az oxitocinnak, de valószínűleg nem vesz részt a szopásra adott PRL válaszban. Az arginin-vazopresszin a hypothalamikus nucleus paraventricularis és supraopticus mag hátulsó magnocelluláris sejtjeiben termelődik. A vazopresszin immunreaktivitás emellett megtalálható a nucleus paraventricularis mediális kissejtes részében is. A magnocelluláris neuronok axonjai a hypophysis hátsó lebenyében, a parvocelluláris neuronokból kiinduló axonok pedig az EM külső rétegében végződnek, így a vazopresszin elérheti az AL sejtjeit a hosszú és a rövid portális ereken keresztül is (166). Korábbi kísérletek megmutatták, hogy a só- és vízháztartás felborítása a hypophysis hátsó lebeny szintjén jelentős változásokat okoz a PRL elválasztásban. A chiasma opticum mögötti kétoldali elülső deafferentáció, a nucleus paraventricularis teljes roncsolása illetve a hátsó lebeny denerválása után diabetes insipidus alakul ki és gátolt a szopásra adott PRL válasz (167, 168, 157, 158). Homozigóta Brattleboro patkányoknál, amelyekben genetikai hiba miatt nem termelődik vazopresszin és így szintén diabetes insipidusban szenvednek, nincs PRL szint emelkedés a szopás inger hatására (169, 170). A vazopresszin. in vivo serkenti a PRL elválasztást (171), in vitro azonban nincs PRL ürítést fokozó hatása (172). A neurofizin II képes emelni a PRL szintet, de valószínűleg nem direkt hypophysealis hatással (173). A 39 aminosavból álló glikopeptid, a vazopresszinneurofizin glikopeptid prekurzor karboxi terminális darabja, a PRL elválasztásban játszott szerepe még bizonytalan, in vitro körülmények között a PRL elválasztást serkentő, gátló illetve nem befolyásoló hatását is leírták már. Mindezen adatok arra
44 utalnak, hogy az vazopresszin és neurofizin II valószínűleg szerepet játszik a PRL elválasztás szabályozásában. A PRL elválasztás szabályozása során ismertetett tényezők, azok sokszínűsége alapján elmondható tehát, hogy a szervezet homeosztázisát szabályozó PRL nem pusztán gátló illetve serkentő hormon által szabályozott, hanem regulációjában a neuroendokrin milieu játszik döntő szerepet
45 3. A salsolinol Az emberi agyban a salsolinolt (11. ábra) először Sjöquist és munkatársai mutattak ki (174). Míg az agyban, és a cerebrospinalis folyadékban csak az (R) enantiomer található meg, addig a perifériás vérben mind az (S) mind az (R) enantiomer kimutatható (174). A salsolinol vér-agy gáton való áthaladásáról eltérőek a vélemények. Korábbi kísérletekben intravénásan beadott salsolinolról kimutatták, hogy nem jutott át a vér-agy gáton. Sjöquist és Magnusson által végzett újabb vizsgálatokban azt találták, hogy az intraperitonealisan adott salsolinol hatására csökkent a nigrostriatalis rendszer DA szintje patkányban. Ez viszont azt sugallja, hogy a salsolinol átjut a vér-agy gáton. Az emberi agyban a salsolinol legnagyobb mennyiségben a striatumban, a hypothalamusban található. HO HO NH CH 3 Salsolinol (SAL) 1-metil-6,7-dihidroxi- 1,2,3,4-tetrahidroizokinolin 11. ábra A salsolinol szerkezete
46 3.1. A salsolinol kémiai szerkezet, szintézisének módjai A salsolinol szerkezetét tekintve a tetrahydroisoquinolinok közé sorolható: 1-metil- 6,7-dihidroxi-1,2,3,4-tetrahydroisoquinolin (lásd 11. ábra). Az emberi szervezetbe a táplálékfelvételen túl (például kakaó, banán, szója egyes vörösborok és sajtok nagy mennyiségben tartalmazzák) dopaminból és acetaldehidből a salsolinol szintetáz enzim katalizálása mellett szintetizálódni is képes (lásd 12. ábra). Ez a 34 kda molekulatömegű enzim kétféle reakció katalízisére képes: egyrészt a fent említett salsolinol szintézisét segíti elő dopaminból és acetaldehidből, másrészt dopaminból és piruvátból salsolinol-1-karboxilsav képződését gyorsítja. Ez utóbbit azonban eddigi ismereteink szerint nem alakítja tovább salsolinollá (176,177,178). 12. ábra A dopamin salsolinol szintézisével kiegészített metabolizációs lehetőségei (a salsolinol szintézise pirossal kiemelve)
47 3.2. A salsolinol hatása a prolaktin elválasztásra Munkacsoportunk a közelmúltban hím, intakt és ovariectomizált nőstény állatokból származó NIL-ből és eminentia medianaból PRF-ként viselkedő anyagként (R)- salsolinolt izolált. A salsolinol (S) enantiomerje számos növényben megtalálható, így táplálékként bekerülhet az állati szervezetbe, míg az (R)-salsolinol enzimatikus úton állatok és az ember központi idegrendszerében is szintetizálódik, dopaminból és acetaldehidből (ld fejezet). A striatum és a hypothalamus DAerg sejtjei tartalmaznak egy úgynevezett salsolinol-szintetáz enzimet, amely képes szelektíven (R)-salsolinolt szintetizálni (75). A NIL denervációja után, a TH immunreaktivitás eltűnésével párhuzamosan nagymértékben lecsökkent a salsolinol mennyisége a NIL-i kivonatban, feltételezhető tehát, hogy a salsolinol a hypothalamikus DAerg neuronokban szintetizálódik, majd a NIL-be jut. A legnagyobb koncentrációban laktáló állatokból származó NIL-ben detektálták az elválasztási periódus előtt. Elválasztás után szintje nagymértékben csökkent, majd szopás hatására szignifikánsan nőtt, párhuzamosan a plazma PRL szintben megfigyelhető változásokkal. Szisztémás salsolinol kezelés gyors, szelektív és dózisfüggő módon serkentette a PRL elválasztást mind hím mind laktáló állatokban, míg a többi elülső lebenyi hormon ürítésére nem volt hatással. Mivel a salsolinol a véragy gáton nem jut át (179), így hypopysealis vagy egyéb vér-agy gáton kívüli hatóhely feltételezhető. A salsolinol in vitro is serkenti a PRL szekréciót, de az in vivo hatásnál kisebb mértékben. Mindezen adatok egyértelműen arra utalnak, hogy a salsolinol szerepet játszhat a PRL elválasztás szabályozásában, azonban ennek helye és mechanizmusa még nem ismert. A salsolinol az eddigi kutatások alapján a DAerg terminálisokon is hathat, mivel az itt fiziológiás körülmények között megtalálható és szekretálható DA mennyiségének jelentős csökkentésével a salsolinol PRL elválasztást serkentő hatása gátlódik (180)
48 A jelen munkában ismertetett kísérleteink célja a salsolinol hatásmechanizmusának tisztázása A salsolinol és származékainak egyéb ismert hatásai A salsolinol metilált származékáról, az N-methyl-salsolinolról korábban kimutatták, hogy neurotoxikus hatással bír, mely kifejezett a nigrostriatalis DAerg sejteken. Az N- metil-salsolinol gátolni képes a tirozin-hidroxiláz aktivitását, mely a DA szintézis sebesség-meghatározó enzime. Emellett gátolja a mitokondriális komplex I és az alfaketoglutarát dehidrogenáz aktivitását is, mely hatásokon keresztül a sejtek energiatermelésére is hatással van (181). Fentieken kívül az N-metil-salsolinol direkt DNS károsító hatással is rendelkezik, valamint káros hatással lehet a szerotonin anyagcserére is. A fent említett mechanizmusokon keresztül főleg a nekrózist és apoptózist indukáló hatásán keresztül a DAerg sejteket elhalásához vezethet. E hatása miatt számos kutatás tárgya az N-metilsalsolinol lehetséges kóroki szerepe a Parkinson-kór kialakulásában (182). Lényeges megemlíteni, hogy az (R)- és (S)-salsolinol jelenlegi ismereteink szerint nem rendelkezik toxikus hatással
49 III. CÉLKITŰZÉSEK Kísérleteinkben a következő kérdésekre kerestünk választ: 1. A salsolinol a PRL elválasztást befolyásoló hatásáról ismert D 2 DA receptoron fejti-e ki hatását, esetleges antagonizmuson keresztül? a., Képes-e specifikus ligandját leszorítani a D 2 -es DA receptorról? b., Képes-e a D 2 receptor kötési paramétereit (K D, B MAX ) befolyásolni? 2. Létezik-e telíthető, specifikus salsolinol kötőhely különböző agyi régiókban illetve a hypophysisben? a., Salsolinol receptor binding assay kifejlesztése a specifikus kötőhelyek vizsgálatához. b., Különböző ligandumok hatásának vizsgálata az esetlegesen létező salsolinol kötőhelyre (leszorításos vizsgálatok). 3. Amennyiben létezik, milyen hatásmechanizmussal képes az in vitro és in vivo észlelt hatását kifejteni? Intracellularis camp szint meghatározása kontroll és salsolinol kezelés esetén. 4. Lehet-e a szoptatással összefüggő PRL elválasztás fokozódását előidéző neurotranszmitter vagy neuromodulátor? Laktáló állatban szoptatás hatására fellépő cmap szint mérése, a salsolinol kezelés alkalmával észlelhető változás összevetése céljából
50 IV. MÓDSZEREK 1. Kísérleti állatok és műtéti beavatkozások Kísérleteink során g testtömegű felnőtt hím, valamint elsőszülő laktáló nőstény Sprague-Dawley patkányokat alkalmaztunk. Az állatokat standard laboratóriumi körülmények között tartottuk 22 C-os szobahőmérsékleten 14:10 valamint 12:12 órás nappal-éjszakai ciklussal. A kísérleteket 10:00 és 14:00 óra között végeztük. A táplálékot és a vizet az állatok szabadon felvehették. Az anyákat egyedi ketrecekben tartottuk a kölykeikkel, a kölykök számát a szülést követő második napon nyolcra csökkentettük. Az állatokat dekapitációval pusztítottuk el. Az agyat gyors eltávolítását követően folyamatosan hűtöttük, majd Glowinski és Iversen módszere szerint (183) preparáltuk. Kísérleteink tervezése és kivitelezése, valamint a kísérleti állatok tartása során a National Institute of Health Guide for Care and Use of Laboratory Animals és a Semmelweis Egyetem Etikai Kódex szerint jártunk el. 2. Alkalmazott módszerek A következőkben ismertetett módszerekkel legkevesebb három, független kísérletet végeztünk. 2.1 Receptor binding assay Kísérleteink során egy viszonylag új - az intézetben eddig még nem alkalmazott - módszert használtunk. A receptor binding assay során a receptorok ligandkötésének paraméterei határozhatóak meg. Mérhető a receptor-ligand kötés erőssége (K A vagy K D ) valamint a kötési kapacitás (B MAX ). A ligand koncentrációjának növelésekor emelkedik a kötődő mennyiség addig, míg szabad ligandkötő hely rendelkezésre áll. Miután telítődtek a receptorok a kötött ligandum mennyisége nem növekszik tovább. A telítési koncentráció ismeretében
51 meghatározható a receptorok mennyisége (B MAX ), valamint az 50 %-os telitettséghez tartozó ligandkoncentráció (K D ). A természetes kötőhelyek mellett a ligand egyéb helyekhez is kapcsolódni képes, de míg a receptorhoz specifikusan kapcsolódik ezek nem specifikus kötések. Szemben a specifikus kötőhelyekkel, a nem specifikusakról a ligand nem szorítható le, így meghatározásukhoz (vagyis a specifikus kötőhelyekről történő leszorítás céljából) feleslegben adott receptor agonistát vagy antagonistát használtunk. A specifikus kötés meghatározásához az így kapott, nem specifikus kötést jellemző értéket az összes kötésből le kell vonni Kísérleti preparátumok 6-8 patkány adenohypophysisét, eltávolításuk után 2 ml jéghideg 0.05 M koncentrációjú nátrium-foszfát pufferrel (ph=7.4) üveg-teflon szöveti homogenizátorban homogenizáltuk, majd a fehérjebontó enzimek aktivitásának csökkentése céljából 2 ml PMSF, DTT, Szójabab Tripszin Inhibitor, Gordox tartalmú proteáz inhibitor koktélba szuszpendáltunk (a proteáz aktivitást gátló koktél pontos összetételét a es fejezetben ismertetem) A mért adatok feldolgozása Scatchard analízis A kapott adatok feldolgozásakor a telítési görbe matematikai átalakítások segítségével egyenessé alakítható, amiről a K D értéke könnyen meghatározható (Scatchard analízis) (317, 329). Az átalakítás lényege, hogy a kötött/szabad radioaktív ligand mennyiségét a kötött függvényében ábrázoljuk. Így egyenest kapunk, melynek meredeksége az -1/K D (-K A ) értékét adja meg, (1) tengely metszete pedig a kötési kapacitást (B MAX ) mutatja
52 Az egyensúlyra vezető biokémiai folyamatok mindegyikére felírható az alábbi egyenlet: A + B AB (1. egyenlet) Ahol A és B... reagáló anyagok AB... keletkezett vegyület Mivel a receptor-ligandum kötése is egy egyensúlyi folyamat, ezért erre az esetre adaptált egyenlet a következő: K 1 R + S < > RS K -1 A fenti folyamatra az egyensúlyi egyenlet: (2. egyenlet) Ahol: R... Receptor S... Szubsztrát RS... Kötött szubsztrát K 1 [RS] 1 = = = K A K -1 [R] [S] K D (3. egyenlet) Mivel a teljes receptorszám a szabad receptorok (R) és a kötött receptorok (RS) számának összege, ezért: [R t ] = [R] + [RS] A (3)-as és a (4)-es egyenletekből adódik: (4. egyenlet) [RS]=K A [S] [R t ] - K A [S] [RS] (5. egyenlet)
53 Az (5)-ös egyenlet átrendezve: [RS] K A [S] = [R t ] 1+K A [S] Ahol [RS]/[R t ] a kötött receptorok frakciója (továbbiakban r ) (6. egyenlet) A (6)-os egyenlet grafikonja a r ([S]) koordináta rendszerben hiperbolát eredményez, melyet telítési görbeként ábrázolunk. Az (3)-as egyenletet átalakítva a (4)-es segítségével a következőt kapjuk: [RS] K A [R t ] - K A [RS] = [S] (7. egyenlet) Ábrázolva [RS]-t [RS]/[S] függvényében egy egyenest kapunk, melynek paraméterei: Az (1)-es tengely metszete a kötési kapacitást adja meg: K [R A t ] - K [RS] = 0 A K [R A t ] = K [RS] A [R t ] = [RS] Ez szemlélteti tehát azt a helyzetet, melyben az összes receptor kötve van. A meredekség (tgα ) pedig a -K A -t adja meg. Az ilyen módon ábrázolt egyenest nevezzük Scatchard egyenesnek (13. ábra). A transzformációhoz alkalmazott módszer a Scatchard Plot
54 13. ábra A Scatchard egyenes, melynek meredeksége -1/K D, (1) tengely metszete a B MAX Hill Plot módszer Amennyiben egy enzim vagy receptor több kötőhelyet tartalmaz (allosztérikus reakció) azok egymással kooperálhatnak, mely lehet pozitív és negatív egymásrahatás is. Matematikai transzformációk sorozatával (melyek ismertetésétől eltekintünk) elvégezhető a Hill Plot transzformáció. Ennek eredményét grafikusan ábrázolva egyenest kapunk (ld. 14. ábra), melynek meredeksége a Hill koefficienst (n H ) jelenti. Amennyiben az n H értéke eggyel egyenlő, akkor nem létezik allosztérikus kötőhely, illetve nincs kooperatív hatás (vagyis kísérleteinkben valóban egyetlen receptorligandkötőhely reakcióról beszélhetünk). Amennyiben n H <1 negatív kooperáció áll fent, n H >1 esetében a kötőhelyek közötti kooperáció pozitív hatású
55 14. ábra Hill Plot transzformáció. Magyarázat a szövegben Dopamin ( 3 H-spiperone) receptor binding assay Az adenohypophysealis D 2 DA receptorok affinitásának és kötési kapacitásának meghatározásához a 3 H-spiperone receptor binding assay módszert alkalmaztuk (184, 185, 186, 187, 188,189). A 200 µl végtérfogatú elegyeket ( mg/ml fehérjekoncentráció) 7-12 különböző koncentrációjú 3 H-spiperone (c= nm, SA= 20 Ci/mmol, Amersham International) izotóppal inkubáltuk. A minták feléhez a nem specifikus kötés (NSB) meghatározása céljából haloperidol-t (c=10-4 M) mértünk. Az inkubációt a homogenátum hozzáadásával indítottuk 37 o C-os vízfürdőben a stady state állapot elérése céljából 30 percig. A membránreceptorokhoz kötött izotóp szeparációját Skatron Cell Harvester segítségével Whatman GF/B filteren keresztül jéghideg 0.05 M koncentrációjú nátrium-foszfát pufferrel (ph=7.4) végeztük. A radioaktivitás detektálása céljából a filterhez 2 ml Ultima Gold TM (Packard) szcintillációs folyadékot mértünk, majd 24 óra múlva a szcintillációs beütésszámot β-counter készülékkel
56 detektáltuk. A mért adatokból a Scatchard analízist Prism software segítségével végeztük (GraphPad Software Inc.) H-salsolinol receptor binding assay Az általunk kidolgozott módszerrel (első publikáció: B. E. Tóth, K. Homicskó, B. Radnai, W. Maruyama, J. E. DeMaria, M. Vecsernyés, M. I. K. Fekete, F. Fülöp, M. Naoi, M. E. Freeman, G. M. Nagy: Salsolinol is a Putative Endogenous Neurointermediate Lobe Prolactin-Releasing Factor. Journal of Neuroendocrinology 13: , 2001) a salsolinol kötőhely kötési tulajdonságai (affinitás, receptorszám) vizsgálható. Dekapitált patkány agyszövetének különböző területeit homogenizáltunk folyamatos hűtés mellett üveg-teflon homogenizátorban, 5 mm MgCl 2 -ot, 1 mm dithiotreitolt (DTT), 100 µm fenil-metil-szulfonil-fluoridot (PMSF), 50 µg/ml szójabab tripszin inhibitort, 40 KIU/ml aprotinint tartalmazó 50 mm-os TRIS pufferben (ph=7,4, 1:40 arányban) g-n 20 percig, 4 ºC-on folytatott centrifugálást követően az üledéket pufferben (50 mm-os ph=7,4 NaCl-ot nem tartalmazó TRIS pufferben 1:75 arányban) reszuszpendáltuk, majd ismételt centrifugálást követően ugyanabban a pufferben 1:75 arányban szuszpendáltuk. A kötésvizsgálathoz az így nyert preparátum 100 µl-ét használtuk 250 µl-es végtérfogatban, három párhuzamos méréssel. A mérőelegy tartalmazott még 50 µl eltérő koncentrációjú 3 H-salsolinol (SA=997,8 GBq/mmol) izotópot (Fülöp Ferenc Szegedi Tudományegyetem, Gyógyszerkémiai Intézet; Kertész István, Tóth Géza MTA Szegedi Biológiai Központ, 190), valamint 100 µl puffert. A nem specifikus kötés vizsgálata 10 µm-os jelöletlen ligand jelenlétében történt. Az inkubáció 90 percig 4 ºC-on történt. A szűrést gyors vákuum filtrációs módszerrel Skatron Cell Harvesterrel Whatman GF/B filteren keresztül, 3x5 ml jéghideg 50 mm-os TRIS-HCl pufferrel (ph=7,4) történő mosás mellett végeztük. Szcintilláció eléréséhez Ultima Gold elegyet (Packard) alkalmaztunk. A mérést 12 óra
57 equilibrium-idő után β-counter-ben végeztük. Az eredményeket Graphpad Prism (Version ) program segítségével értékeltük. A fehérjekoncentráció mérését Lowry módszerével végeztük Direkt leszorításos vizsgálatok Direkt leszorítási vizsgálatok alkalmával a receptorokat azokra specifikus, radioaktívan jelölt ligandummal telítjük, majd növekvő koncentrációban, jelöletlen, az általunk vizsgált anyaggal próbáljuk meg leszorítani kötési helyéről. A különböző jelöletlen ligandum-koncentráció mellett mért beütésszámok alapján felvett görbéből következtethetünk a leszorítás tényére, annak biokémiai paramétereire (pl. IC 50 ) Direkt leszorításos vizsgálatok D 2 receptor kötési helyéről A 200 µl végtérfogatú elegyben ( mg/ml fehérjekoncentráció) található receptorkötő helyeket 10-8 M-os végkoncentrációjú 3 H-spiperone izotóppal telítettük (SA=20 Ci/mmol, Amersham International). A minták feléhez kontroll leszorítási görbe elérése céljából 7 féle koncentrációban ( M) D 2 receptor antagonista Haloperidol-t (C=2 mg/ml, Richter) adtunk. A párhuzamos csövekhez a vizsgálati anyagot mértünk, változó koncentrációban. Az inkubációt a homogenátum hozzáadásával indítottuk 37 o C-os vízfürdőben a egyensúlyi állapot elérése céljából 30 percig. A membránreceptorokhoz kötött izotóp szeparációját Skatron Cell Harvester segítségével Whatman GF/B filteren keresztül jéghideg 0.05 M koncentrációjú nátrium-foszfát pufferrel (ph=7.4) végeztük. A radioaktivitás detektálása céljából a filterhez 2 ml Ultima Gold TM (Packard) szcintillációs folyadékot mértünk, majd 24 óra múlva a szcintillációs beütésszámot β- counter készülékkel detektáltuk. A mért adatokból a Scatchard analízist Prism software segítségével végeztük (GraphPad Software Inc.)
58 Direkt leszorításos vizsgálatok salsolinol kötési helyről A 200 µl végtérfogatú elegyben ( mg/ml fehérjekoncentráció) található receptorkötő helyeket 5*10-8 M-os végkoncentrációjú 3 H-salsolinol izotóppal telítettük (SA=997,8 GBq/mmol). A minták feléhez standard leszorítási görbe elérése céljából 12 eltérő koncentrációban (2,5*10-4 2,5*10-15 M) jelöletlen salsolinol-t adunk, tekintettel arra, hogy egyéb bizonyítottan elfogadható agonista vagy antagonista egyelőre nem áll rendelkezésre. A párhuzamos csövekhez a vizsgálati anyagot mértünk, változó koncentrációban. Az inkubációt a homogenátum hozzáadásával indítottuk 37 o C-os vízfürdőben a stady state állapot elérése céljából 30 percig. A membránreceptorohoz kötött izotópot szeparációját Skatron Cell Harvester segítségével Whatman GF/B filteren keresztül jéghideg 0.05 M koncentrációjú nátrium-foszfát pufferrel (ph=7.4) végeztük. A radioaktivitás detektálása céljából a filterhez 2 ml Ultima Gold TM (Packard) szcintillációs folyadékot mértünk, majd 24 óra múlva a szcintillációs beütésszámot β- counter készülékkel detektáltuk. A mért adatokból a Scatchard analízist Prism software segítségével végeztük (GraphPad Software Inc.) Intracelluláris camp szint meghatározása A sejten belüli camp koncentrációjának meghatározásához Gottshall és munkatársai (192) RIA elvére alapuló módszerét alkalmaztuk. A radioimmunassay vizsgálathoz laktoperoxidáz reakcióval előállított 125 I-Tirozil-cAMP-t használtunk, melyet szintézise után reverz fázisú HPLC módszerrel tisztítottunk Vydac C 18 oszlopon. A mintákat 0,5 ml 5%-os trifluor-acetilsavban homogenizáltuk üveg-teflon homogenizátorban, majd g-vel szobahőmérsékleten 30 percig centrifugáltuk. A felülúszót kétszer liofilizáltuk. A száraz üledéket RIA pufferben feloldva alkalmaztuk a camp meghatározáshoz. A camp antiszérum guanozin-5 -monofoszfáttal, adenozin-trifoszfáttal, valamint 8- bromo-camp-vel végbemenő keresztreakciója kevesebb, mint 0,1%-ban fordul elő
59 3. Statisztikai analízis A statisztikai feldolgozást ANOVA alkalmazásával végeztük, amit Student- Newman-Keuls teszt követett. Két csoport összehasonlítása esetén kétmintás t-próbát használtunk. Szignifikancia megállapításához 95 %-os konfidencia intervallumot (p 0,05) alkalmaztunk
60 V. EREDMÉNYEK 1. Salsolinol hatása a dopamin D 2 receptorra Első vizsgálataink alkalmával arra kerestük a választ, hogy a salsolinol a már ismert DAerg gátló hatás gátlásán keresztül fejti-e ki in vivo észlelhető PRL elválasztást serkentő hatását. Ezek alapján először a D 2 -es DAerg receptorra kifejtett hatást vizsgáltuk 3 H-spiperone receptor binding assay ( módszer), valamint homológ leszorításos vizsgálatok ( módszer) segítségével. A 15. ábra segítségével a háromszor ismételt egybevágó hatást mutatókísérletsorozat egyik reprezentatív eredményét ábrázoltuk. A kísérleteket laktáló állat adenohypophysisének membránpreparátumán végeztük. A salsolinol preinkubáció a radioaktív ligand hozzáadását megelőzően 5 percig tartott. Az (A) ábrán látható kísérlet alkalmával a 3 H-spiperon D 2 -es DA receptorról történő leszorítását vizsgáltuk. Kontrollként az ismert D 2 receptor antagonista Haloperidolt vizsgáltuk, mely látható módon koncentrációfüggő leszorítást eredményez. Ezzel szemben a salsolinol nem képes a radioaktívan jelölt ligandum leszorítására, így joggal felmerül annak lehetősége, hogy a salsolinol kötőhelye a D 2 DA receptortól eltérő. Az előzőekben ismertetett kísérletsorozat eredménye alapján a továbbiakban azt vizsgáltuk, hogy különböző koncentrációban alkalmazott salsolinol képes-e -és amennyiben képes, hogyan és milyen koncentrációban- megváltoztatni a D 2 -es DA receptor 3 H-spiperone kötését, vagyis affinitási konstansát (K A ). A (B) ábrán a 10-6 M koncentráció mellett mért telítési görbét, és származtatott Scatchard-egyenest ábrázoltuk. Ekkor -mint ahogy más, az ábrán nem jelölt koncentráció (10-10 M<C salsolinol <10-4 M) mellett sem- nem látható különbség a kontroll és a salsolinolt tartalmazó minták mellett felvett egyenesek meredeksége között, azaz a receptor affinitásában sem mutatkozik különbség
61 (A) (B) 15. ábra: A salsolinol D 2 -es receptorhoz kötődésének jellemzői: kötési kapacitás, affinitás és leszorítás. (A) A D 2 receptor antagonista haloperidol és 10-6 M koncentrációjú salsolinol leszorítási kísérlete D 2 dopamin receptorról. (B) 3 H- spiperone binding assay: salsolinol hatása a receptorkötésre a Scatchard egyenes feltüntetésével. Magyarázat a szövegben
62 A 15. ábra (B) grafikonján ábrázolt Scatchard egyenes paraméterei a következők: Kísérleti állat Egyenes meredeksége Receptorkötési jellemzők K D B MAX KONTROLL ± , ,01793 SALSOLINOL ± , ,02189 A fenti kísérletekkel igazoltuk tehát, hogy a salsolinol nem a D 2 -es receptorhoz kötődve fejti ki hatását, mivel annak ligandját nem képes leszorítani a receptorról, és affinitási konstansát sem változtatja meg
63 2. Salsolinol specifikus kötőhelyeinek vizsgálata 3 H-salsolinol receptor binding assay módszerrel Folytatásként specifikus salsolinol kötőhelyek létezését próbáltuk igazolni, melyhez 3 H-salsolinol receptor binding assay módszert ( módszer) dolgoztunk ki. Ezt a módszert a világon elsőként publikáltuk Salsolinol kötőhelyek a hypophysisben Ezen kísérleteket laktáló állatokból, dekapitációt követően eltávolított hypophysisekből előállított membránpreparátumokon végeztük. Makroszkóposan szeparálva az elülső- és a NIL-t, külön vizsgáltuk a két terület izotópkötő képességét. A 16. ábra hat független kísérlet eredményeit tartalmazza az átlag szórásának feltüntetésével. A Hill koefficiens értéke egyhez közelít (n H =0,95), mely valódi és nem allosztérikus receptor-kötőhely kapcsolódásra utal. Jól látható, hogy a salsolinol a laktáló állatok hypophysisének elülső, valamint NILének membránjához telíthető módon, specifikusan kötődik. A statisztikai adatokat a következő táblázat foglalja magában: Kötési kapacitás B MAX (pmol/mg fehérje) p Disszociációs konstans K D (nm) Elülső lebeny 5,99±0,62 0,013 78,0±7,8 Közti-hátsó lebeny 7,5±0,3 0, ,8±27,9-63 -
64 16. ábra 3 H-salsolinol szaturációs görbéje laktáló patkány hypophysisének elülső (A) és közti-hátsó lebenyben (B). A három független kísérletből álló sorozat egyik reprezentatív eredménye. Magyarázat a szövegben
65 2.2. Salsolinol kötőhelyek a hypothalamo-hypophysealis rendszerben laktáló és szoptató állatokban Ebben a kísérletsorozatban arra kerestük a választ, hogy laktáló állatokban szoptatás hatására változik-e a 3 H-salsolinol-specifikus kötőhely kötődési paraméterei. Vizsgálatainkban laktáló, illetve 10 percig szoptató patkányok hypophysisének NILét, valamint az EM régióját használtuk membránpreparációhoz, ezeken a es módszerrel vizsgálva az izotópkötő képesség változását szoptatás hatására. 17. ábra 3 H-salsolinol receptor binding assay hypophysis közti-hátsó lebenyén (A) valamint az eminentia mediana (B) területén laktáló és szoptató állatok esetében. Három független kísérlet egyik reprezentatív eredménye. Magyarázat a szövegben
66 Ahogy azt a 17. ábra mutatja, 10 perces szoptatási stimulus hatására a NIL-ben (A) a kötőhelyek affinitásában (K D ) és kötési kapacitásában (B MAX ) egyaránt változás mutatkozik, míg az eminentia mediana területén (B) pusztán a kötési kapacitásban (vagyis a kötőhelyek számában) tapasztaltunk szignifikáns változást. A hypophysis elülső lebenyében valamint a striatumban egyik paraméter esetében sem találtunk szignifikáns változást (az ábrán nincs feltüntetve)
67 2.3 Salsolinol kötőhelyek egyéb agyi régiókban A különböző fiziológiás állapotokban (pl. szoptatás) tapasztalható eltérés mellet érdemes megvizsgálni különböző agyi régió 3 H-salsolinol kötési képességét. Ennek érdekében végeztük az alább ismertetett kísérletsorozatot. Minden szövet esetében három független kísérletet végeztünk. Ebben különböző agyi régiókból készítettünk membránpreparátumot, melyeket a es módszer szerint kötési vizsgálatokat végeztünk. A vizsgált szövetek esetében kapott eredményeket ismerteti az alábbi táblázat: Szövet K D (nm) ± SEM B MAX (pmol/mg fehérje) Hypophysis Elülső lebeny 155,80 ± 27,94 40,30 ± 4,14 Közti-hátsó lebeny 78,00 ± 7,82 34,60 ± 1,40 Eminentia mediana 107,60 ± 34,57 61,30 ± 12,20 Hypothalamus 368,00 ± 91,18 100,50 ± 18,10 Striatum 209,00 ± 37,79 27,10 ± 3,08 Frontalis cortex 91,00 ± 34,97 24,40 ± 2,78 2. táblázat 3 H-salsolinol receptor binding assay módszerével mért receptor-ligand kötődési paraméterek különböző agyi struktúrákban Látható, hogy specifikus, telíthető 3 H-salsolinol kötődés észlelhető a hypophysis mellett az eminetia mediana, a hypothalamus, striatum, valamint a frontalis cortex területén is
68 A legalacsonyabb affinitású kötőhelyek a a hypothalamusban találhatók, míg a legmagasabb K D értéket a hypophysis NIL-ében mértünk. A legmagasabb kötési kapacitással (vagyis a legtöbb receptorral) a hypothalamus rendelkezik
69 3. 3 H-salsolinol leszoríthatósága kötőhelyéről homológ leszorítási vizsgálatokkal Leszorítási vizsgálatok alkalmával különböző kémiai ágensek specifikus kötőhelyért zajló kompeticiója vizsgálható. Ezekből a vizsgálatokból következtethetünk arra, hogy milyen más vegyületek képesek a korábban talált specifikus 3 H-salsolinol kötőhelyhez kötődni. A mérések során a es módszert alkalmaztuk három független kísérletben. A leszorításhoz használt anyagokat a 18. ábra, valamint az 4. táblázat tartalmazza. 18. ábra Salsolinol leszorítása kötőhelyéről
70 A 18. ábra az ott feltüntetett anyagok leszorítóképességét mutatja grafikusan ábrázolva. A leszorítás paramétereit (K i, IC 50 ) az alábbi táblázatban foglaltuk össze: Leszorító anyag IC 50 (M) ± SEM K i (nm) Salsolinol 1,10 ± 0,19 x Dopamin 2,00 ± 0,20 x Apomorphine 5,92 ± 0,68 x L-DOPA 1,72 ± 0,15 x α-methyl-dopa 7,20 ± 0,09 x Carbidopa 2,38 ± 0,16 x Benserazide 2 x 10-7 Noradrenalin 2,31 ± 0,19 x táblázat 3 H-salsolinol leszorítása specifikus kötőhelyéről különböző anyagokkal, a leszorítás paramétereinek feltüntetésével. A 3. táblázatból leolvasható, hogy a 3 H-salsolinolt kötőhelyéről leginkább az α- methyl-dopa, apomorphin, DA, L-DOPA valamint a benserazide, kisebb inhibíciós hatással ugyan, de a noradrenalin, a carbidopa képes leszorítani. A DA transzporter ismert inhibitora, a mazindol nem képes a leszorításra, akárcsak az α 2 receptor agonista clonidine, valamint a további biogén aminok (szerotonin, hisztamin), melyek szintén nem rendelkeznek leszorítóképességgel (a táblázatban nincsenek feltüntetve). A 4. táblázat a leszorítási kísérletekben vizsgált ágenseket mutatja. Piros mezőben a leszorításra képes, kék mezőben a hatással nem bíró vegyületeket foglaltuk össze. Ezen kísérletek egy részét dr. Homicskó Krisztián PhD hallgató végezte az ismertetett módszer felhasználásával
71 L-Dopa + Haloperidol - SAL + Domperidone - Dopamin + Butachlamol - Apomorphine + Sulpiride - Carbidopa + Quinelorane - Benserazide + RGH α-methyl-dopa + 7-OH-DPAT - Clonidine - Mazindol - Papaverin - 5HTP - Tyrosine - Phenylalanine - Tryptophane - NSD táblázat Eddigi leszorítási kísérleteinkben megvizsgált anyagok, és azok hatása. Bal oldalon piros mezőben a leszorító tulajdonságúak, jobb oldalon kék mezőben a leszorítást nem eredményezők

72 4. Szoptatás hatása az intracelluláris camp szintre Kísérleteink során a 4 órás elválasztást követő, 10 percig, valamint további 20 percig tartó szopási stimulus hatására bekövetkező intracelluláris camp szint változást vizsgáltuk (2.3. módszer) laktáló patkány hypophysis elülső lebenyének két, laktotrop sejteket tartalmazó területén, a belső és külső zónájában (IZ-AL és OZ-AL) valamint a közti- és hátsó lebenyében. A hypophysisben mért adatokat szemlélteti a 19. ábra. 19. ábra Intracelluláris camp szint patkány hypophysis elülső lebenyének belső (IZ- AL) és külső (OZ-AL) zónájában kontroll állapotban, valamint 10 és 30 percig tartó szoptatást követően. Három független kísérlet eredménye. A 10 percig tartó szopási stimulus hatására szignifikáns és szelektív camp koncentráció-emelkedést észleltünk a belső zónában (IZ-AL), melyet a 30 percig tartó stimulust követően vizsgált hatás nem követett, az akkor mért camp szint visszaállt a nem szoptatott állatban tapasztalt szintre
73 A külső zónában (OZ-AL) statisztikailag szignifikánsnak minősíthető különbséget nem találtunk. A hypophysis közti- és hátsó lebenyében mért értéket a 20. ábra mutatja. 20. ábra Intracelluláris camp szint patkány hypophysis közti- (IL) és hátsó (NL) lebenyében kontroll állapotban, valamint 10 és 30 percig tartó szoptatást követően. Három független kísérlet eredménye. Ellentétben az adenohypophysisben tapasztalttal, a hátsó lebenyben szignifikáns camp koncentráció csökkenést találtunk mind a 10, mind a 30 percig tartó szoptatási periódust követően. A bazálisan magasabb camp szint ellenére, a IL-ben szignifikáns camp szint változás nem észlelhető. Adataink a minták teljes camp szintet mutatják, így nincs direkt bizonyítékunk arra vonatkozóan, hogy ez a szopás stimulált változás előfordul-e valamennyi agyalapi mirigy sejtpopulációban, vagy csak a laktotropokra korlátozódik
 a kontrollcsoporttal (i.v. fiziológiás sóoldatot kapott) szemben. A kísérleti állatokat a vizsgálat napján reggel 9:00-kor anyjuktól elválasztottuk.")
74 5. Salsolinol kezelés hatása az intracelluláris camp szintre Kísérleteinkben kölykeiktől 4 órára elválasztott patkányoknak 10 mg/ttkg dózisban intravénásan adott salsolinol camp szintet befolyásoló hatását vizsgáltuk (2.3 módszer) a kontrollcsoporttal (i.v. fiziológiás sóoldatot kapott) szemben. A kísérleti állatokat a vizsgálat napján reggel 9:00-kor anyjuktól elválasztottuk. 4 órával később az anyák 10mg/ttkg dózisban intravénásan salsolinolt kaptak egyszeri, bólus adagolásban. 15 perc elteltével az állatokat dekapitáltuk, az eminentia mediana-t valamint a hypophysist gyorsan eltávolítottuk, majd az elülső lebenyt és a NIL-t szem ellenőrzése mellett szeparáltuk. A kezelést követő tizenötödik percben dekapitált állatok eminentia mediana területéről származó szövetében mért camp értékeit a 21. ábra, míg a közti-hátsó és elülső lebenyben kapott mérési eredményeket a 22. ábra foglalja össze. 21. ábra Salsolinol kezelés hatása az eminentia mediana camp koncentrációjára. Magyarázat a szövegben. Három független kísérlet eredménye
75 22. ábra Salsolinol kezelés hatása hypophysis közti-hátsó, valamint elülső lebenyének camp koncentrációjára. Magyarázat a szövegben. Három független kísérlet eredménye. Az eminentia mediana területén szignifikáns camp szint csökkenést tapasztaltunk, míg az elülső lebenyben szignifikáns camp szint emelkedést lehetett kimutatni. A NILben nem találtunk szignifikáns eltérést
76 VI. DISZKUSSZIÓ A klasszikus neuroendokrinológiai nézet szerint a hypothalamus arcuatus magjában képződő és az eminentia mediana területén a hosszú portális erekbe szekretálódó DA tónusos gátló hatást fejt ki az agyalapi mirigy laktotrop sejtjeire. Eszerint a PRL elválasztás serkentése a gátló hatású DAerg pályarendszerek gátlásának útján alakul ki. Jimmy D. Neill és munkacsoportja az emlő érző idegeinek ingerlésével szopási stimulust szimulált, eközben vizsgálva a hypophysis-nyél és az eminentia mediana DA tartalmát (193, 194, 85). Vizsgálataikkal igazolták, hogy a szopási stimulusra létrejövő PRL-szint emelkedést csak annak első 3-5 percében kíséri a neuroendokrin DAerg rendszer aktivitásának 60-70%-os csökkenése, ezután gyors, sorozatos DA pulzációt figyeltek meg, mely az ingerlés végéig fennmaradt. Ezen kísérletek alapján megállapítható, hogy a PRL elválasztás fokozódását a DA aktivitás csökkenése legfeljebb beindítja, de annak fenntartásában más, PRF-nak is szerepet kell játszania. Habár számos, PRL elválasztást fokozó (úgynevezett PRL release aktivitással rendelkező) anyagot sikerült azonosítani (ld. Irodalmi áttekintés 2.2.), de ezek mindegyike más funkciójáról ismert, így ez a hatás legfeljebb járulékos jelentősséggel bír. Számos munkacsoport kutatásainak célja ezért a PRL releasing faktor(ok) (PRF) azonosítása. A jelen értekezés alapjául szolgáló kísérletekben az előkísérleteink során a NIL perklórsavas kivonatából (mely a PRL elválasztást serkenteni képes) azonosított, és in vivo PRL elválasztást fokozó hatással rendelkező salsolinol hatásának mechanizmusára kerestünk magyarázatot. A salsolinol a TIDA vagy a PHDA pályarendszereken keresztül ürülhet (akár a DA-al kotranszmitterként) a NIL területén, ahonnan a rövid portális erek útján elérheti az adenohypophysis laktotrop sejtjeit. A DA-nal ellentétes salsolinol hatás kifejeződése több szinten, többféle mechanizmussal képzelhető el (a lehetséges mechanizmusokat a 23. ábra foglalja össze): 1. Az intracellulárisan DA-ból képződött salsolinol gátolni képes a DA szintézisét, ezzel csökkentve a DA leadást. Ezt a mechanizmust támasztja alá
77 Minami és munkatársai (195) megfigyelése, mely szerint a salsolinol képes gátolni a TH enzimaktivitást. 2. A salsolinol a VMAT gátlása révén csökkenti a szinaptikus vezikulum DA tartalmát. 3. A DA-nal kotranszmitterként ürülve, vagy önmagában felszabadulva a. képes a DA hatásért felelős D 2 DAerg receptorhoz DA antagonistaként kötődni. b. saját receptorához kötődve a DA által kiáltott másodlagos hírvivő mechanizmusokat ellentétes hatással módosítani. 4. A salsolinol képes a DA neuronalis visszavételét a DAT-en keresztül fokozni, ezáltal annak extraneuronális térből történő eliminációját elősegíteni

78 23. ábra A salsolinol lehetséges támadáspontjai és hatásmechanizmusa. A feltételezett gátló hatások piros, a serkentők kék nyíllal jelölve. Magyarázat a szövegben
Hypothalamikus struktúrák és faktorok jelentősége a prolaktin. elválasztás szabályozásában. Bodnár Ibolya. Doktori (Ph.D.
 Hypothalamikus struktúrák és faktorok jelentősége a prolaktin elválasztás szabályozásában Bodnár Ibolya Doktori (Ph.D.) értekezés Témavezető: Prof. Dr. Nagy György Semmelweis Egyetem, Doktori Iskola Idegtudományok
Hypothalamikus struktúrák és faktorok jelentősége a prolaktin elválasztás szabályozásában Bodnár Ibolya Doktori (Ph.D.) értekezés Témavezető: Prof. Dr. Nagy György Semmelweis Egyetem, Doktori Iskola Idegtudományok
AZ ÖSZTROGÉN ÉS A DEHIDROEPIANDROSZTERON SZEREPE A SZINAPTIKUS ÁTRENDEZŐDÉSBEN
 AZ ÖSZTROGÉN ÉS A DEHIDROEPIANDROSZTERON SZEREPE A SZINAPTIKUS ÁTRENDEZŐDÉSBEN c. PhD-értekezés magyar nyelvű összefoglalója Csákvári Eszter Témavezető: Dr. Párducz Árpád Magyar Tudományos Akadémia Szegedi
AZ ÖSZTROGÉN ÉS A DEHIDROEPIANDROSZTERON SZEREPE A SZINAPTIKUS ÁTRENDEZŐDÉSBEN c. PhD-értekezés magyar nyelvű összefoglalója Csákvári Eszter Témavezető: Dr. Párducz Árpád Magyar Tudományos Akadémia Szegedi
HYPOTHALAMIKUS STRUKTÚRÁK ÉS FAKTOROK JELENTOSÉGE A PROLAKTIN ELVÁLASZTÁS SZABÁLYOZÁSÁBAN. Bodnár Ibolya. Doktori (Ph.D.) értekezés tézisei
 értekezés tézisei") HYPOTHALAMIKUS STRUKTÚRÁK ÉS FAKTOROK JELENTOSÉGE A PROLAKTIN ELVÁLASZTÁS SZABÁLYOZÁSÁBAN Bodnár Ibolya Doktori (Ph.D.) értekezés tézisei Témavezeto: Prof. Dr. Nagy György Semmelweis Egyetem, Doktori Iskola
HYPOTHALAMIKUS STRUKTÚRÁK ÉS FAKTOROK JELENTOSÉGE A PROLAKTIN ELVÁLASZTÁS SZABÁLYOZÁSÁBAN Bodnár Ibolya Doktori (Ph.D.) értekezés tézisei Témavezeto: Prof. Dr. Nagy György Semmelweis Egyetem, Doktori Iskola
Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában. Doktori tézisek. Dr. Oláh Márk
 Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában. Doktori tézisek Dr. Oláh Márk Semmelweis Egyetem Szentágothai János Idegtudományok Doktori Iskola Témavezető:
Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában. Doktori tézisek Dr. Oláh Márk Semmelweis Egyetem Szentágothai János Idegtudományok Doktori Iskola Témavezető:
Endokrinológia. Közös jellemzők: nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások. váladékuk a hormon
 Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az élő szervezetek menedzserei, a hormonok
 rekkel exponálunk a munka végén) és azt utólag kivonjuk digitálisan a képekből. A zajcsökkentés dandárját mindig végezzük a raw-képek digitális előhívása során, mert ez okozza a legkevesebb jelvesztést
rekkel exponálunk a munka végén) és azt utólag kivonjuk digitálisan a képekből. A zajcsökkentés dandárját mindig végezzük a raw-képek digitális előhívása során, mert ez okozza a legkevesebb jelvesztést
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegrendszer és a hormonális rednszer szabályozó működése
 Az idegrendszer és a hormonális rednszer szabályozó működése Az idegrendszer szerveződése érző idegsejt receptor érző idegsejt inger inger átkapcsoló sejt végrehajtó sejt végrehajtó sejt központi idegrendszer
Az idegrendszer és a hormonális rednszer szabályozó működése Az idegrendszer szerveződése érző idegsejt receptor érző idegsejt inger inger átkapcsoló sejt végrehajtó sejt végrehajtó sejt központi idegrendszer
2006 1. Nemszinaptikus receptorok és szubmikronos Ca2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra.
 2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
Belső elválasztású mirigyek
 Belső elválasztású mirigyek Szekréciós szervek szövettana A különböző sejtszervecskék fejlettsége utal a szekretált anyag jellemzőire és a szekréciós aktivitás mértékére: Golgi komplex: jelenléte szekrétum
Belső elválasztású mirigyek Szekréciós szervek szövettana A különböző sejtszervecskék fejlettsége utal a szekretált anyag jellemzőire és a szekréciós aktivitás mértékére: Golgi komplex: jelenléte szekrétum
Az adenohipofizis. Az endokrin szabályozás eddig olyan hormonokkal találkoztunk, amelyek közvetlen szabályozás alatt álltak:
 Az adenohipofizis 2/10 Az endokrin szabályozás eddig olyan hormonokkal találkoztunk, amelyek közvetlen szabályozás alatt álltak: ADH, aldoszteron, ANP inzulin, glukagon szekretin, gasztrin, CCK, GIP eritropoetin
Az adenohipofizis 2/10 Az endokrin szabályozás eddig olyan hormonokkal találkoztunk, amelyek közvetlen szabályozás alatt álltak: ADH, aldoszteron, ANP inzulin, glukagon szekretin, gasztrin, CCK, GIP eritropoetin
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
Homeosztázis és idegrendszer
 Homeosztázis és idegrendszer Magatartás és homeosztázis a hipotalamusz és a limbikus rendszer ingerlése összehangolt motoros-vegetatívendokrin változásokat indít ezek a reakciók a homeosztázis fenntartására,
Homeosztázis és idegrendszer Magatartás és homeosztázis a hipotalamusz és a limbikus rendszer ingerlése összehangolt motoros-vegetatívendokrin változásokat indít ezek a reakciók a homeosztázis fenntartására,
megerősítik azt a hipotézist, miszerint az NPY szerepet játszik az evés, az anyagcsere, és az alvás integrálásában.
 Az első két pont a növekedési hormon (GH)-felszabadító hormon (GHRH)-alvás témában végzett korábbi kutatásaink eredményeit tartalmazza, melyek szervesen kapcsolódnak a jelen pályázathoz, és már ezen pályázat
Az első két pont a növekedési hormon (GH)-felszabadító hormon (GHRH)-alvás témában végzett korábbi kutatásaink eredményeit tartalmazza, melyek szervesen kapcsolódnak a jelen pályázathoz, és már ezen pályázat
Zárójelentés. A) A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.
 A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.") Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
Az elért eredmények. a) A jobb- és baloldali petefészek supraspinális beidegzése
 A jobb- és baloldali petefészek supraspinális beidegzése") Az elért eredmények A perifériás belsőelválasztású mirigyek működésének szabályozásában a hypothalamushypophysis-célszerv rendszer döntő szerepet játszik. Közvetett, élettani megfigyelések arra utaltak,
Az elért eredmények A perifériás belsőelválasztású mirigyek működésének szabályozásában a hypothalamushypophysis-célszerv rendszer döntő szerepet játszik. Közvetett, élettani megfigyelések arra utaltak,
Kolin-acetiltranszferáz
 Kolin-acetiltranszferáz Neurotranszmitter-kritériumok: Szintetizáló enzim-készlet ( kulcs-enzimek ) Tároló-rendszer (vezikuláris transzporterek) Felvevő /lebontó rendszer Adagolással posztszinaptikus válasz
Kolin-acetiltranszferáz Neurotranszmitter-kritériumok: Szintetizáló enzim-készlet ( kulcs-enzimek ) Tároló-rendszer (vezikuláris transzporterek) Felvevő /lebontó rendszer Adagolással posztszinaptikus válasz
Az endokrin szabályozás általános törvényszerűségei
 Az endokrin szabályozás általános törvényszerűségei a kémiai és idegi szabályozás alapelvei hormonok szerkezete, szintézise, tárolása, szekréciója hormonszintet meghatározó tényezők hormonszekréció szabályozása
Az endokrin szabályozás általános törvényszerűségei a kémiai és idegi szabályozás alapelvei hormonok szerkezete, szintézise, tárolása, szekréciója hormonszintet meghatározó tényezők hormonszekréció szabályozása
Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában.
 Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában. Doktori értekezés Dr. Oláh Márk Semmelweis Egyetem Szentágothai János Idegtudományi Doktori Iskola Témavezető:
Új hypothalamo-hypophysealis mechanizmusok szerepe a hypophysis hormonürítésének szabályozásában. Doktori értekezés Dr. Oláh Márk Semmelweis Egyetem Szentágothai János Idegtudományi Doktori Iskola Témavezető:
A Humánmorfológiai és Fejlődésbiológiai Intézet rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2012/2013-es tanévre
 témái a 2012/2013-es tanévre") A Humánmorfológiai és rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2012/2013-es tanévre 1. Neokortikális interneuronok thalamikus inputjának quantitativ analízise Témavezető:
A Humánmorfológiai és rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2012/2013-es tanévre 1. Neokortikális interneuronok thalamikus inputjának quantitativ analízise Témavezető:
A Humánmorfológiai és Fejlődésbiológiai Intézet rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2011/2012-es tanévre
 témái a 2011/2012-es tanévre") A Humánmorfológiai és rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2011/2012-es tanévre 1. Neokortikális interneuronok thalamikus inputjának quantitativ analízise Témavezető:
A Humánmorfológiai és rektori pályatételei, szakdolgozati és tudományos diákköri (TDK) témái a 2011/2012-es tanévre 1. Neokortikális interneuronok thalamikus inputjának quantitativ analízise Témavezető:
Központi idegrendszeri vizsgálatok.
 Központi idegrendszeri vizsgálatok. Újszülött patkány endorfin imprintingje után, felnőtt állatban a szexuális magatartást, szteroid hormon kötést és az agyi szerotonin szinteket vizsgáltuk. A nőstények
Központi idegrendszeri vizsgálatok. Újszülött patkány endorfin imprintingje után, felnőtt állatban a szexuális magatartást, szteroid hormon kötést és az agyi szerotonin szinteket vizsgáltuk. A nőstények
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
A köztiagy (dienchephalon)
") A köztiagy, nagyagy, kisagy Szerk.: Vizkievicz András A köztiagy (dienchephalon) Állománya a III. agykamra körül szerveződik. Részei: Epitalamusz Talamusz Hipotalamusz Legfontosabb kéregalatti érző- és
A köztiagy, nagyagy, kisagy Szerk.: Vizkievicz András A köztiagy (dienchephalon) Állománya a III. agykamra körül szerveződik. Részei: Epitalamusz Talamusz Hipotalamusz Legfontosabb kéregalatti érző- és
A nem. XY XX nemi kromoszómapár. here - petefészek. férfi - nő. Női nemi szervek. Endometrium. Myometrium
 Reproduktív funkciók Nemi hormonok: Androgének Ösztrogének Gesztagének Támpont: 86 (nyomokban 90 is) A nem Genetikai nem Gonadális nem Nemi fenotípus XY XX nemi kromoszómapár here - petefészek férfi -
Reproduktív funkciók Nemi hormonok: Androgének Ösztrogének Gesztagének Támpont: 86 (nyomokban 90 is) A nem Genetikai nem Gonadális nem Nemi fenotípus XY XX nemi kromoszómapár here - petefészek férfi -
Hormonális szabályozás
 Hormonális szabályozás A Bárdos-Husvéth-Kovács: Gazdasági állatok anatómiájának és élettanának alapjai (Mezőgazda Kiadó, 2007) tankönyv ábráinak felhasználásával Szabályozás: kommunikáció! Vezérlés és
Hormonális szabályozás A Bárdos-Husvéth-Kovács: Gazdasági állatok anatómiájának és élettanának alapjai (Mezőgazda Kiadó, 2007) tankönyv ábráinak felhasználásával Szabályozás: kommunikáció! Vezérlés és
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Az agy betegségeinek molekuláris biológiája. 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5.
 Az agy betegségeinek molekuláris biológiája 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5. Alzheimer kór 28 Prion betegség A prion betegség fertőző formáját nem egy genetikai
Az agy betegségeinek molekuláris biológiája 1. Prion betegség 2. Trinukleotid ripít betegségek 3. ALS 4. Parkinson kór 5. Alzheimer kór 28 Prion betegség A prion betegség fertőző formáját nem egy genetikai
A hormonok a sejtek közötti kémiai kommunikációt biztosítják, mely összehangolja a szervek működését és az aktuális igényekhez igazítja azt.
 71. Az endokrin szabályozás alapelvei 72. A hypothalamo-hypophysealis (neuroendokrin) rendszer általános jellemzése A hormonok a sejtek közötti kémiai kommunikációt biztosítják, mely összehangolja a szervek
71. Az endokrin szabályozás alapelvei 72. A hypothalamo-hypophysealis (neuroendokrin) rendszer általános jellemzése A hormonok a sejtek közötti kémiai kommunikációt biztosítják, mely összehangolja a szervek
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Tóth-Petrovics Ágnes: Szaporasági teljesítmények növelése exogén hormonális kezelések nélkül
 Tóth-Petrovics Ágnes: Szaporasági teljesítmények növelése exogén hormonális kezelések nélkül Tejtermelő tehenek szaporításának nehézségei tenyésztői szemmel A tejtermelés világszerte szinte kizárólag holstein
Tóth-Petrovics Ágnes: Szaporasági teljesítmények növelése exogén hormonális kezelések nélkül Tejtermelő tehenek szaporításának nehézségei tenyésztői szemmel A tejtermelés világszerte szinte kizárólag holstein
Reproduktív funkciók 1. Androgén hormonok
 Reproduktív funkciók 1. Androgén hormonok Mellékvesekéreg (Zona reticularis) Here (Leydig sejtek) (Petefészek) Androgének Dehidroepiandroszteron-szulfát Dehidroepiandroszteron Androszténdion Androgének
Reproduktív funkciók 1. Androgén hormonok Mellékvesekéreg (Zona reticularis) Here (Leydig sejtek) (Petefészek) Androgének Dehidroepiandroszteron-szulfát Dehidroepiandroszteron Androszténdion Androgének
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai
 Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
Dózis-válasz görbe A dózis válasz kapcsolat ábrázolása a legáltalánosabb módja annak, hogy bemutassunk eredményeket a tudományban vagy a klinikai gyakorlatban. Például egy kísérletben növekvő mennyiségű
NÖVÉNYÉLETTAN. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Auxinok Előadás áttekintése 1. Az auxinok felfedezése: az első növényi hormon 2. Az auxinok kémiai szerkezete és
NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Auxinok Előadás áttekintése 1. Az auxinok felfedezése: az első növényi hormon 2. Az auxinok kémiai szerkezete és
MAGYAR NYELVŰ ÖSSZEFOGLALÁS
 MAGYAR NYELVŰ ÖSSZEFOGLALÁS Bevezetés Ph.D. munkám során az agynak a neurodegeneratív folyamatok iránti érzékenységét vizsgáltam, különös tekintettel a korai neonatális fejlődést befolyásoló különböző
MAGYAR NYELVŰ ÖSSZEFOGLALÁS Bevezetés Ph.D. munkám során az agynak a neurodegeneratív folyamatok iránti érzékenységét vizsgáltam, különös tekintettel a korai neonatális fejlődést befolyásoló különböző
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Orvosi élettan. Bevezetés és szabályozáselmélet Tanulási támpontok: 1.
 Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 anyagcsere hőcsere Az élőlény és környezete nyitott rendszer inger hő kémiai mechanikai válasz mozgás alakváltoztatás
Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 anyagcsere hőcsere Az élőlény és környezete nyitott rendszer inger hő kémiai mechanikai válasz mozgás alakváltoztatás
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
PhD vizsgakérdések április 11. Próbálja meg funkcionális szempontból leírni és példákon bemutatni az intralimbikus kapcsolatok jelentőségét.
 PhD vizsgakérdések 2012. április 11 1 Mi a szerepe a corpus geniculatum lateralé-nak a látásban? Próbálja meg funkcionális szempontból leírni és példákon bemutatni az intralimbikus kapcsolatok jelentőségét.
PhD vizsgakérdések 2012. április 11 1 Mi a szerepe a corpus geniculatum lateralé-nak a látásban? Próbálja meg funkcionális szempontból leírni és példákon bemutatni az intralimbikus kapcsolatok jelentőségét.
A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására
 OTKA 62707 A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására Zárójelentés A gesztációs diabetes mellitus (GDM) egyike a leggyakoribb terhességi komplikációknak, megfelelő
OTKA 62707 A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására Zárójelentés A gesztációs diabetes mellitus (GDM) egyike a leggyakoribb terhességi komplikációknak, megfelelő
OTKA ZÁRÓJELENTÉS
 NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
NF-κB aktiváció % Annexin pozitív sejtek, 24h kezelés OTKA 613 ZÁRÓJELENTÉS A nitrogén monoxid (NO) egy rövid féléletidejű, számos szabályozó szabályozó funkciót betöltő molekula, immunmoduláns hatása
A hypophysis prolaktin elválasztásának szabályozása: intracelluláris mechanizmusok és a glükokortikoidok szerepe. Dr. Horváth Katalin Mónika
 A hypophysis prolaktin elválasztásának szabályozása: intracelluláris mechanizmusok és a glükokortikoidok szerepe Dr. Horváth Katalin Mónika A doktori (Ph.D.) értekezés tézisei Semmelweis Egyetem, Doktori
A hypophysis prolaktin elválasztásának szabályozása: intracelluláris mechanizmusok és a glükokortikoidok szerepe Dr. Horváth Katalin Mónika A doktori (Ph.D.) értekezés tézisei Semmelweis Egyetem, Doktori
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben Tartalék energiaforrás, membránstruktúra alkotása, mechanikai
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben Tartalék energiaforrás, membránstruktúra alkotása, mechanikai
A sejtek közöti kommunikáció formái. BsC II. Sejtélettani alapok Dr. Fodor János
 A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A magatartás szabályozása A vegetatív idegrendszer
 A magatartás szabályozása A vegetatív idegrendszer A magatartási válasz A külső vagy belső környezetből származó ingerekre adott komplex (szomatikus, vegetatív, endokrin) válaszreakció A magatartási választ
A magatartás szabályozása A vegetatív idegrendszer A magatartási válasz A külső vagy belső környezetből származó ingerekre adott komplex (szomatikus, vegetatív, endokrin) válaszreakció A magatartási választ
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
HORMONÁLIS SZABÁLYOZÁS
 HORMONÁLIS SZABÁLYOZÁS Hormonok: sejtek, sejtcsoportok által termelt biológiailag aktív kémiai anyagok, funkciójuk a szabályozás, a célsejteket a testnedvek segítségével érik el. Kis mennyiségben hatékonyak,
HORMONÁLIS SZABÁLYOZÁS Hormonok: sejtek, sejtcsoportok által termelt biológiailag aktív kémiai anyagok, funkciójuk a szabályozás, a célsejteket a testnedvek segítségével érik el. Kis mennyiségben hatékonyak,
A fiziológiás terhesség hátterében álló immunológiai történések
 A fiziológiás terhesség hátterében álló immunológiai történések APAI Ag ANYAI Ag FERTŐZÉS AUTOIMMUNITÁS MAGZATI ANTIGEN ALACSONY P SZINT INFERTILITAS BEÁGYAZÓDÁS ANYAI IMMUNREGULÁCIÓ TROPHOBLAST INVÁZIÓ
A fiziológiás terhesség hátterében álló immunológiai történések APAI Ag ANYAI Ag FERTŐZÉS AUTOIMMUNITÁS MAGZATI ANTIGEN ALACSONY P SZINT INFERTILITAS BEÁGYAZÓDÁS ANYAI IMMUNREGULÁCIÓ TROPHOBLAST INVÁZIÓ
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
AZ UTÓDGONDOZÁS ÉLETTANA
 VISELKEDÉSÉLETTAN 7. ELŐADÁS AZ UTÓDGONDOZÁS ÉLETTANA Dobolyi Árpád ELTE, Élettani és Neurobiológiai Tanszék A REPRODUKCIÓS CIKLUS AZ ANYÁK VISELKEDÉSI ÉS FIZIOLÓGIÁS VÁLTOZÁSAI AZ OXYTOCIN ÉS VAZOPRESSIN
VISELKEDÉSÉLETTAN 7. ELŐADÁS AZ UTÓDGONDOZÁS ÉLETTANA Dobolyi Árpád ELTE, Élettani és Neurobiológiai Tanszék A REPRODUKCIÓS CIKLUS AZ ANYÁK VISELKEDÉSI ÉS FIZIOLÓGIÁS VÁLTOZÁSAI AZ OXYTOCIN ÉS VAZOPRESSIN
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
A NŐI NEMI MŰKÖDÉS ÉS A LAKTÁCIÓ SZABÁLYOZÁSÁNAK STRESSZFÜGGÉSE
 A NŐI NEMI MŰKÖDÉS ÉS A LAKTÁCIÓ SZABÁLYOZÁSÁNAK STRESSZFÜGGÉSE 2013.Február 27. Oláh Márk Semmelweis Egyetem Humánmorfológiai és Fejlődésbiológiai Intézet olah.mark@med.semmelweis-univ.hu MELY HYPOTHALAMO-HYPOPHYSEALIS-
A NŐI NEMI MŰKÖDÉS ÉS A LAKTÁCIÓ SZABÁLYOZÁSÁNAK STRESSZFÜGGÉSE 2013.Február 27. Oláh Márk Semmelweis Egyetem Humánmorfológiai és Fejlődésbiológiai Intézet olah.mark@med.semmelweis-univ.hu MELY HYPOTHALAMO-HYPOPHYSEALIS-
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
NÖVÉNYÉLETTAN. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010
 NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Sejtfal szintézis és megnyúlás Környezeti tényezők hatása a növények növekedésére és fejlődésére Előadás áttekintése
NÖVÉNYÉLETTAN Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 Sejtfal szintézis és megnyúlás Környezeti tényezők hatása a növények növekedésére és fejlődésére Előadás áttekintése
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Pszichiátriai zavarok neurobiológiai alapjai
 Pszichiátriai zavarok neurobiológiai alapjai Kéri Szabolcs 1 1. Alapfogalmak: anatómia, fiziológia 2. Funkcionális lokalizáció az agyban 3. Szinapszisok és neurotranszmitterek 4. A neurotranszmisszió molekuláris
Pszichiátriai zavarok neurobiológiai alapjai Kéri Szabolcs 1 1. Alapfogalmak: anatómia, fiziológia 2. Funkcionális lokalizáció az agyban 3. Szinapszisok és neurotranszmitterek 4. A neurotranszmisszió molekuláris
Gonádműködések hormonális szabályozása áttekintés
 Gonádműködések hormonális szabályozása áttekintés Hormonok szerepe a reproduktív működésben érett ivarsejtek létrehozása és fenntartása a megtermékenyítés körülményeinek optimalizálása a terhesség fenntartása,
Gonádműködések hormonális szabályozása áttekintés Hormonok szerepe a reproduktív működésben érett ivarsejtek létrehozása és fenntartása a megtermékenyítés körülményeinek optimalizálása a terhesség fenntartása,
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI. - autokrin. -neurokrin. - parakrin. -térátvitel. - endokrin
 A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
A KÉMIAI KOMMUNIKÁCIÓ ALAPELVEI - autokrin -neurokrin - parakrin -térátvitel - endokrin 3.1. ábra: Az immunreakciók főbb típusai és funkciójuk. IMMUNVÁLASZ TERMÉSZETES ADAPTÍV humorális sejtes HUMORÁLIS
ÖNINGERLÉS PRANCZ ÁDÁM
 ÖNINGERLÉS PRANCZ ÁDÁM Az agyi jutalmazási ( revard ) rendszere: Egyes fiziológiai tevékenységek, mint az evés, ivás, nemi kontaktus 2jutalmazottak, örömmel, gyönyörrel, kellemes érzéssel vagy kielégüléssel
ÖNINGERLÉS PRANCZ ÁDÁM Az agyi jutalmazási ( revard ) rendszere: Egyes fiziológiai tevékenységek, mint az evés, ivás, nemi kontaktus 2jutalmazottak, örömmel, gyönyörrel, kellemes érzéssel vagy kielégüléssel
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
[S] v' [I] [1] Kompetitív gátlás
![[S] v' [I] [1] Kompetitív gátlás](/thumbs/93/114506146.jpg "[S] v' [I] [1] Kompetitív gátlás") 8. Szeminárium Enzimkinetika II. Jelen szeminárium során az enzimaktivitás szabályozásával foglalkozunk. Mivel a klinikai gyakorlatban használt gyógyszerhatóanyagok jelentős része enzimgátló hatással bír
8. Szeminárium Enzimkinetika II. Jelen szeminárium során az enzimaktivitás szabályozásával foglalkozunk. Mivel a klinikai gyakorlatban használt gyógyszerhatóanyagok jelentős része enzimgátló hatással bír
Kutatási beszámoló ( )
") Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Kutatási beszámoló (2008-2012) A thrombocyták aktivációja alapvető jelentőségű a thrombotikus betegségek kialakulása szempontjából. A pályázat során ezen aktivációs folyamatok mechanizmusait vizsgáltuk.
Jegyzőkönyv. dr. Kozsurek Márk. A CART peptid a gerincvelői szintű nociceptív információfeldolgozásban szerepet játszó neuronális hálózatokban
 Jegyzőkönyv dr. Kozsurek Márk A CART peptid a gerincvelői szintű nociceptív információfeldolgozásban szerepet játszó neuronális hálózatokban című doktori értekezésének házi védéséről Jegyzőkönyv dr. Kozsurek
Jegyzőkönyv dr. Kozsurek Márk A CART peptid a gerincvelői szintű nociceptív információfeldolgozásban szerepet játszó neuronális hálózatokban című doktori értekezésének házi védéséről Jegyzőkönyv dr. Kozsurek
Testtömeg szabályozás. Palicz Zoltán
 Testtömeg szabályozás Palicz Zoltán A hypothalamus fontosabb magcsoportjai Alapfogalmak Orexigén projekció: táplálékfelvételt indukáló idegpálya Anorexigén projekció: táplálékfelvételt gátló idegpálya
Testtömeg szabályozás Palicz Zoltán A hypothalamus fontosabb magcsoportjai Alapfogalmak Orexigén projekció: táplálékfelvételt indukáló idegpálya Anorexigén projekció: táplálékfelvételt gátló idegpálya
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Vásárhelyi Barna. Semmelweis Egyetem, Laboratóriumi Medicina Intézet. Az ösztrogénekimmunmoduláns hatásai
 Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Vásárhelyi Barna Semmelweis Egyetem, Laboratóriumi Medicina Intézet Az ösztrogénekimmunmoduláns hatásai Ösztrogénhatások Ösztrogénhatások Morbiditás és mortalitási profil eltérő nők és férfiak között Autoimmun
Az erek simaizomzatának jellemzői, helyi áramlásszabályozás. Az erek működésének idegi és humorális szabályozása. 2010. november 2.
 Az erek simaizomzatának jellemzői, helyi áramlásszabályozás. Az erek működésének idegi és humorális szabályozása 2010. november 2. Az ér simaizomzatának jellemzői Több egységes simaizom Egy egységes simaizom
Az erek simaizomzatának jellemzői, helyi áramlásszabályozás. Az erek működésének idegi és humorális szabályozása 2010. november 2. Az ér simaizomzatának jellemzői Több egységes simaizom Egy egységes simaizom
NYÁRÁDY ERAZMUS GYULA ORSZÁGOS MAGYAR KÖZÉPISKOLAI BIOLÓGIA TANTÁRGYVERSENY XI. OSZTÁLY MAROSVÁSÁRHELY 2013. május 11. FELADATLAP
 NYÁRÁDY ERAZMUS GYULA ORSZÁGOS MAGYAR KÖZÉPISKOLAI BIOLÓGIA TANTÁRGYVERSENY XI. OSZTÁLY MAROSVÁSÁRHELY 2013. május 11. FELADATLAP A feladatlap kitöltésére 1.5 órád van. A feladatlapon 60 sorszámozott tesztfeladatot
NYÁRÁDY ERAZMUS GYULA ORSZÁGOS MAGYAR KÖZÉPISKOLAI BIOLÓGIA TANTÁRGYVERSENY XI. OSZTÁLY MAROSVÁSÁRHELY 2013. május 11. FELADATLAP A feladatlap kitöltésére 1.5 órád van. A feladatlapon 60 sorszámozott tesztfeladatot
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban
 A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban Doktori (PhD) értekezés Siklódi Erika Rozália Biológia Doktori Iskola Iskolavezető: Prof. Erdei Anna, tanszékvezető egyetemi
A humán tripszinogén 4 expressziója és eloszlási mintázata az emberi agyban Doktori (PhD) értekezés Siklódi Erika Rozália Biológia Doktori Iskola Iskolavezető: Prof. Erdei Anna, tanszékvezető egyetemi
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
Sejtek közötti kommunikáció:
 Sejtek közötti kommunikáció: Mi a sejtek közötti kommunikáció célja? Mi jellemző az endokrin kommunikációra? Mi jellemző a neurokrin kommunikációra? Melyek a közvetlen kommunikáció lépései és mi az egyes
Sejtek közötti kommunikáció: Mi a sejtek közötti kommunikáció célja? Mi jellemző az endokrin kommunikációra? Mi jellemző a neurokrin kommunikációra? Melyek a közvetlen kommunikáció lépései és mi az egyes
I. Spinális mechanizmusok vizsgálata
 Az OTKA pályázat keretében több irányban végeztünk kutatásokat a fájdalom mechanizmusok tisztázása érdekében. Az egyes kutatások eredményeit külön fejezetekben ismertetem. I. Spinális mechanizmusok vizsgálata
Az OTKA pályázat keretében több irányban végeztünk kutatásokat a fájdalom mechanizmusok tisztázása érdekében. Az egyes kutatások eredményeit külön fejezetekben ismertetem. I. Spinális mechanizmusok vizsgálata
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
III./2.2.: Pathologiai jellemzők, etiológia. III./2.2.1.: Anatómiai alapok
 III./2.2.: Pathologiai jellemzők, etiológia Ez az anyagrész az önálló fejfájások pathomechanizmusát foglalja össze. A tüneti fejfájások kóreredetét terjedelmi okokból nem tárgyaljuk. III./2.2.1.: Anatómiai
III./2.2.: Pathologiai jellemzők, etiológia Ez az anyagrész az önálló fejfájások pathomechanizmusát foglalja össze. A tüneti fejfájások kóreredetét terjedelmi okokból nem tárgyaljuk. III./2.2.1.: Anatómiai
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
VEGETATIV IDEGRENDSZER AUTONOM IDEGRENDSZER
 VEGETATIV IDEGRENDSZER AUTONOM IDEGRENDSZER A szervezet belső környezetének_ állandóságát (homeostasisát) a belső szervek akaratunktól független egyensúlyát a vegetativ idegrendszer (autonóm idegrendszer)
VEGETATIV IDEGRENDSZER AUTONOM IDEGRENDSZER A szervezet belső környezetének_ állandóságát (homeostasisát) a belső szervek akaratunktól független egyensúlyát a vegetativ idegrendszer (autonóm idegrendszer)
Hormonális szabályozás
 Hormonális szabályozás Alapfogalmak Hormon: sejtek, sejtcsoportok által termelt, biológiailag aktív anyag, amely a célsejteket a testnedvek közvetítésével éri el (humorális szabályozás). Hatására a sejtanyagcsere
Hormonális szabályozás Alapfogalmak Hormon: sejtek, sejtcsoportok által termelt, biológiailag aktív anyag, amely a célsejteket a testnedvek közvetítésével éri el (humorális szabályozás). Hatására a sejtanyagcsere
Aktív életerő HU/KAR/0218/0001
 Aktív életerő HU/KAR/0218/0001 A bizonyítottan javítja az idősödő kutyák életminőségét: élénkebbé teszi az állatokat és ezáltal aktívabb életmódot tesz lehetővé számukra. Az oxigenizáció mellett a szív-
Aktív életerő HU/KAR/0218/0001 A bizonyítottan javítja az idősödő kutyák életminőségét: élénkebbé teszi az állatokat és ezáltal aktívabb életmódot tesz lehetővé számukra. Az oxigenizáció mellett a szív-
Orvosi élettan. Bevezetés és szabályozáselmélet Tanulási támpontok: 1.
 Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 1. Szabályozáselmélet Definiálja a belső környezet fogalmát és magyarázza el, miért van szükség annak szabályozására.
Orvosi élettan Bevezetés és szabályozáselmélet Tanulási támpontok: 1. Prof. Sáry Gyula 1 1. Szabályozáselmélet Definiálja a belső környezet fogalmát és magyarázza el, miért van szükség annak szabályozására.
Az agykéreg és az agykérgi aktivitás mérése
 Az agykéreg és az agykérgi aktivitás mérése Intrakortikális hálózatok Elektromos aktiváció, sejtszintű integráció Intracelluláris sejtaktivitás mérés Sejten belüli elektromos integráció 70 mv mikroelektrod
Az agykéreg és az agykérgi aktivitás mérése Intrakortikális hálózatok Elektromos aktiváció, sejtszintű integráció Intracelluláris sejtaktivitás mérés Sejten belüli elektromos integráció 70 mv mikroelektrod
A KOLESZTERIN SZERKEZETE. (koleszterin v. koleszterol)
") 19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként