DEBRECENI EGYETEM Oktatási segédlet oldal {frissítve } Összeállította : Szentirmai Attila Emeritus egyetemi tanár
|
|
|
- Elvira Faragóné
- 9 évvel ezelőtt
- Látták:
Átírás
1 DEBRECENI EGYETEM Oktatási segédlet oldal {frissítve } Összeállította : Szentirmai Attila Emeritus egyetemi tanár BIOTERMÉK TECHNOLÓGIA Szerkesztési elvek: taxonomiai fajnevek dőlt betüvel; Publikáció (folyóirat.évszám.kötetszám:oldalszám tól-ig); Enzimnév néha dőlt betüvel; TARTALOMJEGYZÉK: BEVEZETÉS EXPOZÍCIÓ? 1 AZ ELJÁRÁSOKHOZ SZÜKSÉGES SZERVEZETEK KIVÁLASZTÁSA 3 SZTEROIDOK DEHIDROGÉNEZÉSE 14 ALKOHOLOK OXIDÁCIÓJA KETONNÁ VAGY ALDEHIDDÉ 18 SZÉN-SZÉN KETTŐSKÖTÉS TELÍTÉSE 21 A KETTŐS KÖTÉS IZOMERIZÁCIÓJA 22 SZTEROIDOK HIDROXILEZÉSE 24 KORTIKOIDOK ELŐÁLLÍTÁSÁT SZOLGÁLÓ EREDMÉNYEK 28 A SZTEROID VÁZ OXIDATÍV LEBOMLÁSA NORSZTEROID TOTÁLSZINTÉZIS 41 ENANTIOMER ÖSZTRON INTERMEDIER ELŐÁLLÍTÁSA 48 6-AMINOPENICILLINSAV (6-APS) ELŐÁLLÍTÁSA 49 KONVERZIÓRA ALKALMAS SEJTTÖMEG FELHASZNÁLÁSA 63 7-AMINO KEFÉMSAV (7-ACA) és szármaszékainak ELŐÁLLÍTÁSA 64 L-AMINOSAK RLŐÁLLÍTÁSA 66 AZ AROMÁS AMINOSAVBIOSZINTÉZIS SZABÁLYOZOTTSÁGA 73 ELJÁRÁSOK TRIPTOFÁN ELŐÁLLÍTÁSÁRA 85 L-ASCORBINSAV ELŐÁLLÍTÁSÁHOZ KAPCSOLÓDÓ TEVÉKENYSÉG 92 GLÜKÓZ-FRUKTÓZ CUKORSZÖRP NAGYIPARI ELŐÁLLÍTÁSA 95 BIOETANOL 100 A CIKLODEXTRIN BIOLÓGIÁJA 105 KÖRNYEZETET SZENNYEZŐ ANYAGOK BIOLÓGIAI HASZNOSÍTÁSA 111 Szakmai dolgozat kérdéseire adható válasz 119 BEVEZETÉS EXPOZÍCIÓ? Az idesorolható biotechnológiai folyamatokat a szakirodalom eredetileg biotranszformációnak nevezve tárgyalta. Ugyanakkor transzformációnak nevezték a mikrobiális genetika művelői a mikroorganizmus genetikai állományában idegen DNS bevitelével előidézett öröklődő változást. Ezért célszerűnek látszik a megkülönböztető biokonverzió elnevezés használata. Ezekben az eljárásokban különféle szerves vagy szervetlen kiindulási vegyületekből egy vagy több enzim specifikus katalitikus aktivitását hasznosítva jutunk el a célvegyületnek tekintett (piacképes) hasznos termékhez. Az eljárással szemben támasztott követelmény, hogy a szubsztrátum jó hatásfokkal, minimális anyagvesztés mellett, nagy koncentrációban, (kis térfogatban) kerüljön átalakításra. A kiválasztott mikroszervezeteket a fenti cél elérése érdekében olyan fermentációs körülmények között dolgoztatják, hogy a túlélés egyetlen lehetősége számukra a kívánt reakciósor hatásos működtetése legyen. Az átalakítandó kiindulási vegyület többnyire valamilyen preparatív kémiai tevékenység köztes terméke (kulcs intermedierje), amely általában a primer anyagcsere enzimei által katalizált biokonverziós reakcióban szubsztrát-analogként vesz részt. 1

2 A gazdaságosságra törekedve biokonverziós eljárást alkalmaznak a kémiailag körülményesen kivitelezhető feladat megoldására. (Az alkohol dehidrogenáz királis vegyületek előállítására kerülhet alkalmazásra. A galaktóz oxidáz katalizálja a királis, vagy prokirális alifás alkoholok sztereospecifikus oxidációját. A peroxidáz az arómás vegyületek hidroxilezésére, a szulfatáz a fenolizomerek szétválasztására.használható. A karbonsav eszterázok a rezolválást egyszerüsítik.) Az eljárások egy részénél az élő sejt jelenléte szükséges, míg más részük akár rögzített enzimekkel is lefolytatható. A mikrobiológus az utóbbi esetben az enzimtermelésre használható mikroorganizmus tömegtenyésztését végzi, kidolgozva a technológiát, amely a kiválasztott, vagy genetikai munkával előállított mikroszervezet gazdaságos alkalmazását lehetővé teszi. A mikrobiális enzimek specificitását hasznosítva környezetszennyezés nélkül vállalkozhatunk különféle vegyipari alapanyagok előállítására is Az optimális eljárásban kémiai lépések és biológiai reakciók váltakozva is részt vehetnek, amint az példaként a CETUS által propénből propilénoxid előállítására kifejlesztett eljárásban tanulmányozható * Az első lépésben az eljárás H 2 O 2 (hidrogénperoxid) igényét kielégítendő az Oudemansiella mucido piranoz oxidáza glükózból szorbozont állít elő, amit a ciklust teljesítve platina katalizátor mellett redukálnak vissza glükózzá. * A Clamidiomyces fumago klórperoxidáza állítja elő propénből a klórpropánt, * amiből H 2 O 2 jelenlétében egy Flavobacterium törzs klórhidrin epoxidáza alakítja ki a propilénoxidot. 2
 Az eljárások egy részénél az élő sejt jelenléte szükséges, míg más részük akár rögzített enzimekkel is lefolytatható.")
3 AZ ELJÁRÁSOKHOZ SZÜKSÉGES SZERVEZETEK KIVÁLASZTÁSA Első feladat a természetes élőhelyükön előforduló populációból kiválasztani az igényeinknek megfelelő tulajdonságú, a reprodukálhatóság szempontjából genetikailag stabil (törzs) klón kiválasztása. A feltétel teljesítése szempontjából előnyös a gyüjteményekben elhelyezett törzsek átvizsgálása, amelyeknél a szabadalmi szempontból kívánatos rendszertani vizsgálatokat már elvégezték. A gyüjtemények katalógusainak tanulmányozásán kívül sokat segít a szakirodalom ismerete és néhány kézikönyv. Ilyen például Cherney és Herzog Microbial transformations of Steroiods /Academic. P. New York 1967/, vagy Klaus Kieslich Microbial transformation of Non-steroid Cyclic Compounds /Georg Thieme Verlag Stuttgart 1976/ Ezekben az átalakítandó vegyületek, illetve a a termékek szempontjából kapunk használható adatot az átalakítást végző mikroorganizmus megjelölésével. Az irodalom tájékoztat arról, hogy bizonyos reakciók elősegítésére milyen rendszertani csoportok részletesebb vizsgálata vezethet eredményre. Az utóbbi időben hasonló összefoglaló munkák nem jelentek meg, mert a különböző számítógéppel elérhető adatbázisok feleslegessé teszik. A kiválasztás kísérleti munkáját indokolja a leírtaknál előnyösebb.törzs izolálásának lehetősége, továbbá a genetikai munkával fejlesztett variánsok alkalmazhatóságának reménye. A kiválasztás rutinszerű.munka. Folyékony táptalajon növesztett tenyészethez adagolják az átalakítandó szubsztrátumot. Meghatározott ideig végzett átalakítási periódus után általában a céltermék kinyerése szempontjából előnyös szerves oldószeres extrakcióval különíthető el a termék. Más esetben a vizes fázisból vett minta összetétele kromatográfiával elválasztva vizsgálható. Könnyebb a törzs kiválasztása, ha az elvégzendő átalakítás a mikróba számára javítja az életkörülményeket. A szterin oldallánc lebontására alkalmas mikroszervezet táptalaja egyedüli szénforrásként koleszterint tartalmazott. Hasonló meggondolás szerint paraffint hasznosító mikroszervezet izolálása érdekében a tenyésztésre használt táptalaj egyedüli szénforrásként normál paraffint tartalmazott. A kinőtt tenyészetet többször átoltva hasonló összetételű táptalajra; a paraffint hasznosító tenyészet oltóanyaga reprodukálhatóan fenntartható. A következő lépésben a tenyészetet addig hígítjuk, amíg egyetlen élősejtből növekedő tenyészethez jutunk. A klón elkülönítése, néha nehéz bonyolult feladat, mert lehetséges, hogy a törzs növekedése egy konzorcium hatásos együttműködését jelenti. A törzs ez esetben tiszta formában nem tartható fenn. 3

4 SZTEROIDOK MIKROBIÁLIS ÁTALAKÍTÁSA A szteroidok mikrobiológiai átalakítását célzó kutatások kezdettől fogva szorosan kapcsolódtak a gyógyszeripar igényeihez. A szteroid hormonok és hormonanalógok előállítására kidolgozott eljárásokban a meglehetősen drága és sok esetben kémiailag érzékeny vegyületekre tekintettel a biológiai lépések az enzimreakciók sztereospecifitása, valamint a biológiai eljárások enyhe reakciókörülményei miatt - kezdettől fogva fontos szerepet nyertek. A világszerte alkalmazott mikrobiológiai eljárásokban oxigenázok, oxidoreduktázok, izomerázok, dehidrogenázok, dehidratázok és acilázok működését hasznosítják. A szteroidok átalakítását célzó kutatómunka irányát szakmai mélységét minden esetben a gyakorlati előnyök, az ipar által támasztott igények szabták és szabják meg ma is. A gazdaságosság és nem kis mértékben a világpiaci ár határozza meg a bevezetendő eljárással szemben támasztott követelményeket. A biokonverziós eljárásban felhasználható mikrobiális reakció legyen tehát - egy termékhez vezető - teljessé tett - viszonylag kis térfogatban végezhető - rövid idő alatt végbemenő folyamat - olcsó segédanyagokat igénylő - egyszerü berendezésekben lefolytatható - a teremék kinyerésére viszonylag egyszerű, hatásos módszer szolgáljon A gazdaságosság a kémiai reakciók és az enzimreakció legelőnyösebb kombinálását, optimális illesztését teszi szűkségessé. A fermentációs eljárás költséges volta miatt sokszor célszerűbb több egymást követő enzimreakciót a köztes termék kinyerése és tisztítása nélkül végrahajtani. Ezen kívánalmaknak megfelelően a mikrobiológus feladata egyrészt az elvárást teljesítő törzs kiválasztása, illetve genetikai módszerekkel történő előállítása; másrészt a kiválasztott törzs élettani tulajdonságainak felderítése után a megismert, genetikailag meghatározott biokémiai adottságainak gazdaságos hasznosítása üzemi körülmények között. Végül is biztonságosan megvalósítható eljárás kidolgozása és beillesztése a szintézisút kémiai lépései közé. Az ipari gyakorlat általában olyan vegyület átalakítására kívánja a kiválasztott mikroorganizmus enzimkészletét használni, amely a normál anyagcserében nem fordul elő, sőt sok esetben a környezetben elő nem forduló szintetikus termékek előállítása a cél. Ezek átalakítására az ad lehetőséget, hogy az enzim felépítésétől függően a természetes szubsztrátumhoz hasonló szerkezetű vegyület is kapcsolódhat az aktív centrumhoz és a reakció sikeres befejeztével onnan el is távozhat. Egy-egy új vegyület mikrobiológiai átalakítására szolgáló eljárás kidolgozása a különböző forrásból származó, különböző fajba sorolható mikroszervezetek aktivitásának összehasonlításával kezdődik. Ezt a munkát jobbára minden új vegyület átalakításakor a törzsek biológiai aktivitásának változatossága miatt célszerű elvégezni, annak ellenére, hogy a szerves vegyületek átalakításáról könnyen kezelhető kézikönyvek, áttekinthatő összefoglalók állnak rendelkezésünkre. Ez azért javasolható, mert az irodalmi adatok, különösen a nemzetkozi gyüjteményekből megszerezhető törzsek szabadalmakban közölt eredményei sok esetben csak a bejelentők reményeit tartalmazzák. A szteroidok mikrobiológiai átalakítására szolgáló enzimek jelenléte illetve hiánya nem köthető taxonomiai bélyegekhez. Ebből következően azonos fajba sorolt törzsek az átalakítóképesség, és a gyakorlati alkalmazhatóság szempontjából igen eltérőek lehetnek. Célszerű a kiválasztott törzs szubsztrátspecifitását alkalmas modellvegyületek adagolásával feltérképezni. A szubsztrátumtól függő technológia kidolgozása érdekében szubsztrátspecifitásra vonatkozó adatokat előnyösen sejtmentes enzimkivonat alkalmazásával gyüjthetünk, jóllehet az ipari gyakorlatban élő, vagy elölt mikroorganizmusok konverziós aktivitását hasznosítjuk. 4

5 Jelen lehet a mikroszervezet csupán az enzim hordozójaként (például a 3,17-dihidroxi-4- pregnen-3,20-dion előállítása Streptomyces griseocarneus elölt miceliumtömegével), sok esetben azonban az enzim működéséhez szükséges kofaktorok folytonos regenerálásához az élő sejt jelenléte nélkülözhetetlen. (hidrokortizon előállítása Absidia orchidis tenyészetével, vagy koleszterin oldallánc lebontása Mycobacterium tenyészettel). A mikrobiológiai átalakítást végző enzim működését befolyásoló tényezők: - szubsztrátum koncentráció hatása - a termék koncentráció hatása - az enzim szubsztrátspecificitása - ph hatása a reakciósebességre - ph hatása a reakció irányára - hőmérséklet hatása a reakciósebességre - kofaktor (ellátás) aktuális koncentráció hatása a reakciósebességre - az enzim működését befolyásoló egyéb tényezők (gátló ill. serkentők) jelenléte A kofaktorellátottságot befolyásoló tényezők: - kofaktort regeneráló rendszer aktivitása - anaerob illetve aerob reakciókörülmények hatása - a regeneráló rendszer szubsztrátum-ellátottságának hatása - a kofaktor másirányú felhasználási lehetősége az anyagcserében Szubsztrátum ellátottságot befolyásoló tényezők: - a sejtburok áteresztőképességének alakulása - a szubsztrátum, illetve a termék aktuális koncentrációja, vizoldékonyságuk Az átalakítást katalizáló enzim képződését befolyásoló tényezők: - indukálhatóság esetén az optimális (szelektív) induktor alkalmazása - represszálhatóság vizsgálata, derepresszió alkalmazása - konstitutívan derepresszált mutáns előállítása - zavaró mellékreakciók hatásának csökkentése. Az átalakítási reakció szempontjából ideális mikroorganizmus kiválasztása: Csak a kívánt reakciót katalizálja. A mellékreakciókat katalizáló enzimek képződése visszaszorítható legyen, Rosszabb esetben a melléktermék képződése szelektíven gátolható legyen. Reverzibilis reakció esetén az egyensúly a kívánt irányba mozdítható legyen Ha lehet a kívánt reakciót katalizáló enzim konstitutívan derepresszált legyen, rosszabb esetben képződése indukálható legyen A kofaktor szint a tenyésztési körülmények változtatásával optimalizálható legyen. * A kutató-fejlesztő munka művelőinek a feladata a fenti szempontok figyelembevételével a kidolgozandó, illetve már kidolgozott eljárások versenyképességét, piaci helyzetét szüntelen javítani. Az ipari tevékenység céljára kiválasztott szervezetek természetes, vagy célzottan előállított öröklött genetikai állományát a bonyolult élettani összefüggések ismeretében kialakított eljárásban lehet hasznosítani. A példaként felsorolt eljárások kidolgozásakor minden esetben nem a mikroorganizmus növekedése, hanem a céltermék képződése szempontjából optimális fizikai és kémiai körülmény meghatározása a feladat. Ezt a kérdést a folytonosan növekedő tenyészetben lehet érdemében vizsgálni, mivel az úgynevezett batsh eljárás változó körülményei között nehéz felismerni a folyamatot meghatározó tényezőt. 5

6 Szakaszos eljárás estén a reakcióközeg összetétele állandóan változik; A szubsztrátum fogy, a termék mennyisége növekedőben van, közben felszaporodnak bizonyos nemkívánt melléktermékek, amelyek zavarhatják a kiválasztott törzs normál anyagcseréjét. Ebből következően, a fejlesztés gazdasági eredményét szolgáló optimális feltételek adatait csak folytonosan növekedő tenyészetben folytatott vizsgálatok segítségével a fizikai és kémiai körülmények változtatásával nyert részeredmények ismeretében határozhatjuk meg. 6

7 SZTERINEK. SZTEROLOK KÉMIÁJA Az alapváz ciklopentano-perhidro-fenantrén. A humán szervezetben előforduló zooszterin, a koleszterin C 27 H 46 O (vízben nemoldódó, szintelen, optikailag aktív, vizes alkoholból *H 2 O-val lemezesen kristályosodó vegyület) Az A gyürű hidroxil csoportjának helyzete 90%-.ban ß-térállású. A 5 telítés esetén az epi szerkezet irányába tolódik az egyensúly. A 10, 13, 17 helyzetű szubsztitució cisz helyzetű. Nyolc asszimetria centrum alapján levezethető 256 sztereoizomer közül biolgiailag csupán egyetlen változat aktív. Átalakításuknál ezért a biológiai módszerek gazdasági előnyt élveznek. Ipari eljárással a koleszterinből ivarhormonok, kéreghormonok, D-3 vitamin készíthető Az emlős szervezetben a máj feladata a genetikailag meghatározott koleszterin szint biztosítása, amelyből egyrészt az epesavak(cholsav származékok) képződnek, másrészt különböző életfontosságú szteroid-hormonokat állítanak elő a belsőelválasztású mirigyek. A gombák membránjában ergoszterol fordul elő diosgenin (C 27 H 48 O 3 ) Ipari szempontból alapanyagként szerepeltek a vizes oldatban habzó növényi steroidsaponinok (hexózhoz kapcsolt sapogenin), amelynek aglükon frakciójából oxidatív módszerrel pregnán vegyület (3β,17α,21-trihidroxi-5-pregnen-20-on.21-acetát) nyerhertő. Példaként említhető a Dioscorea kaktusz gyökeréből nyert saponinból hidrolizált termék a Olcsó gyógyszeripari alapanyag előállítás szempontjából a növényi szterinek közül a szójabab olajból nyert γ-szitoszterin említendő /A gyapotmagolajból nyert β-szitoszterinben a C 24 -re β-helyzetben kapcsolódik az etil!/ γ-szitoszterin 7

8 IZOPRÉN-OLIGOMEREK BIOSZINTÉZISE (a fontosabb köztestermékek szerkezete) acetoacetil-coa-szintetáz 2 acetil-coa >CoA-SH acetoacetil-coa < acetil-coa hidroximetil-glutaril-coa-szintetáz >CoA-SH COOH H-C-H β-hidroxi-β-metilglutaril-coa HO-C-CH 3 < NADPH H-C-H hidroximetil-glutaril-coa-reduktáz >CoA-SH O=C-S-CoA >2 NADP + H H OH H 3 R-mevalonát HO-C--C--C---C--COOH mevalonát-kináz < ATP H H CH 3 H >ADP H H OH H 5-foszfo-mevalonát H 2 O 3 -P-O-C--C--C---C--COOH < ATP H H CH 3 H mevalonátfoszfát-kináz >ADP H H OH H 5-pirofoszfo-mevalonát PP--O-C--C--C---C--COOH < ATP H H CH 3 H 5-pirofoszfo-mevalonát dekarboxiláz >CO >ADP H izomeráz H-C-H H-C-H dimetilallil-pirofoszfát izopentenil-pirofoszfát C-CH 3 C.CH 3 C-H H-C-H dimetilallil H-C-O--PP H-C-O--PP transzferáz H H geranil PP transzferáz geranil-pirofoszfát tetraizoprenil-pirofoszfát < NADPH tetraizoprenil- -PP reduktáz >4 NADP + CH 3 PP--O-HCH-HC=C-HCH-HCH-HC=C CH 3 CH 3 izoprenil PP transzferáz oligoizoprén szintáz farnezil-pirofoszfát >baktoprenol-foszfát retinál fitanil-pirofoszfát szkvalén szintáz < NADPH >NADP + karotinok szkvalén 8

9 ERGOSZTEROL KÉPZŐDÉS VÁZLATA 9
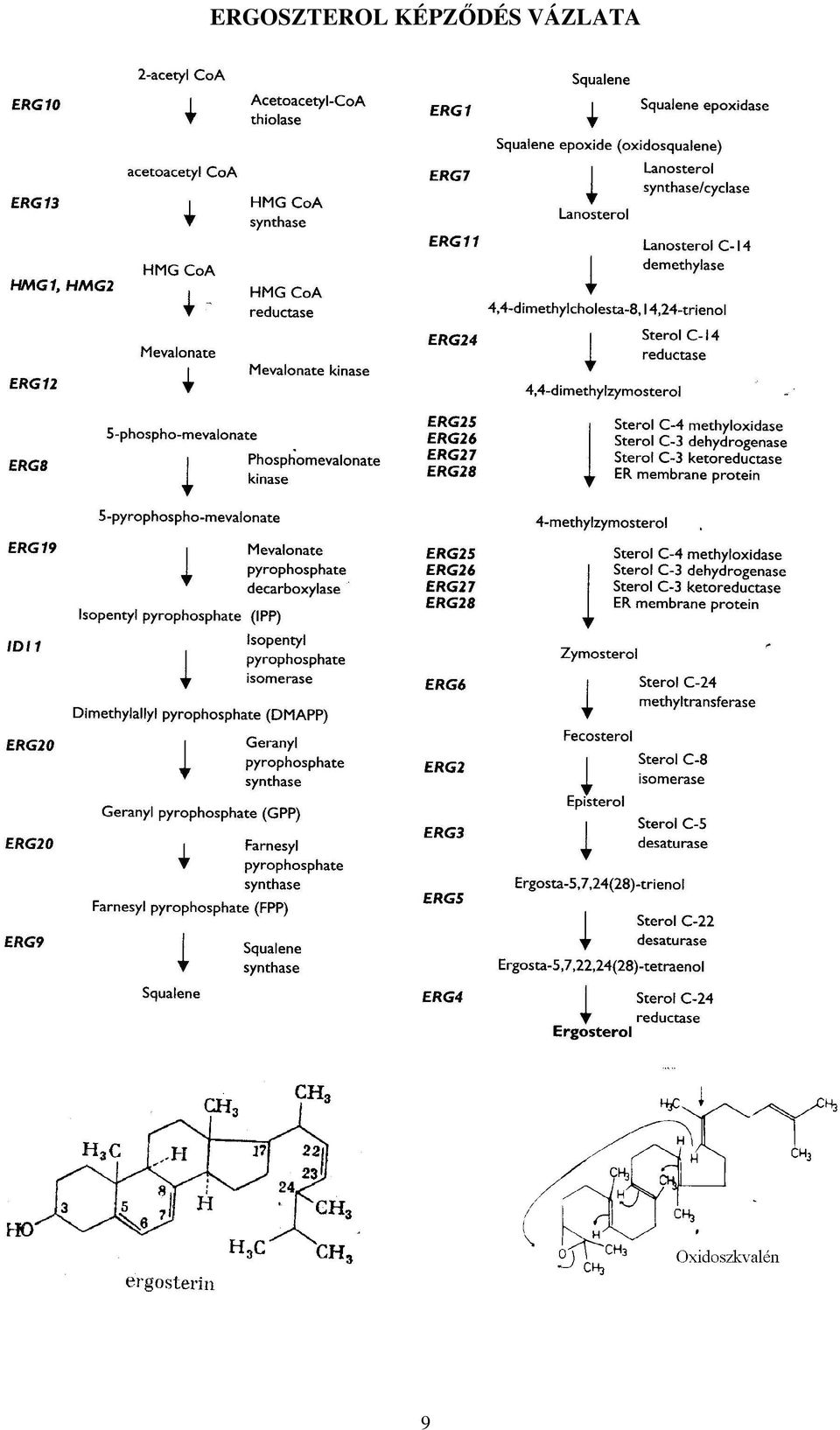
10 Az antibiotikus hatású anyagok tulajdonságaival foglalkozó mikrobiológusok évtizedek óta keresnek hatásos fungisztatikus anyagokat. A Streptomycesek által termelt anyagok nistatin felfedezése mellett a gombavilág egymás elleni küzdelmét szolgáló anyagok felismerése is reményt keltőnek tünt. Valójában először ez a vegyületcsoport mint ismeretlen szerkezetű antifungális anyag került izolálásra. A XX. század utolsó negyedében Merck Sharp and Dohme gondozásában került a piacra a lovastatin hatásos népszerű koleszterin szint csökkentőként. A koleszterin szintézis nélkülözhetetlen építőelemének a mevalonsavnak a képződését gátolva gyakorlatilag a koleszterin képződést lehet akadályozni. A kutatócsoport (A. Endo és munkatársai) a hetvenes években elméleti meggondolásból a fungisztatikus hatású mikroorganizmusok között célzottan szterinszintézist gátló mikrobiális eredetű vegyületet kerestek. Közel 8000 törzs biológiai aktivitását vizsgálták patkány máj enzimrendszerén in vitro. A Penicillium által termelt compactinról - későbbi néven mevastatinról ban jelent meg az első közlemény. Később a compactin metilezett származékát az Aspergillus fajok tenyészlevében is megtalálták. Az első összefoglaló dolgozat a hetvenes évek végén jelent meg. (J. Antibiot. 32: ) a Monascus ruber tenyészlevéből kinyerhető hatásos vegyületekről. A következő évben l980- ban A.V. Alberts és munkatársai Aspergillus terreus tenyészlevében találtak hasonló hatású hexahidro naftalénhez kapcsolódó heptánsavakat. (Proc. Natl. Acad. Sci. USA, 77: ). A pravastatin savként kerül kiszerelésre. A többi "statin" lakton alakja a májban felnyílik és így létrjön a gátlás szempontjából fontos hidroxisav. Jól tudott, hogy a mevalon-sav a szterinszintézis korai köztesanyaga. A lovastatin és rokonvegyületei, mint intracelluláris enzim-gátlók a mevalonsav képződés akadályozásával a magasabb-rendűekben a szterin képződést, a koleszterin szintézist gátolják. A kompetitív gátló hatás már 1 nm tartományban jelentkezik. Ez három negyságrenddel alacsonyabb mint a HMG-CoA (3-hidroxi-3-metilglutaril-CoA) disszociációs állandója. Ezek a vegyületek az izeltlábúakban akadályozzák a juvenilis (fejlődési) hormon képződést, a növényekben és gombákban a szterin képződését. mevalon-sav NADPH-reduktáz 10

11 A vegyületcsoport a zsírsav szintézishez hasonló módon acetil-coa elemekből épül fel. A kabonil csoportok eliminálásához szükséges NADPH intenziv szénhidrát anyagcserét feltételez. Ebből következik, hogy ez a vegyületcsoport nem az idiofázisban képződik hanem a fejlődési ciklusban. 11

12 MIÉRT ÉRDEKELT A GYÓGYSZERIPAR A SZTEROIDOK MIKROBIOLÓGIAI ÁTALAKÍTÁSÁRA SZOLGÁLÓ ELJÁRÁSOK FEJLESZTÉSÉBEN? A hormonhatású vegyületek hatásának befolyásolása analógokkal, a módosított változattal új hatás kiváltása is lehetséges. AZ EMLŐS SZERVEZETBEN KOLESZTERINBŐL KÉPZŐDŐ ÉLETTANILAG JELENTŐS VEGYÜLETEK GESZTAGÉNek képviselője Progeszteron C-3 helyen karbonil konjugált kettős kötéssel C-17-hez szénhidrogén lánc kapcsolódik C-20 karbonillal KORTIKOIDok képviselője Dezoxikortikoszteron C-3 helyen karbonil konjugált kettős kötéssel C-17-hez szénhidrogén lánc kapcsolódik C-20 karbonillal C-21 hidroxilezve GLÜKOKORTIKOID képviselő Hidrokortizon C-3 helyen karbonil konjugált kettős kötéssel C-17-hez szénhidrogén lánc kapcsolódik C-20 karbonillal és α-helyzetű hidroxil csoport C-11 β-helyzetű hidroxil csoport képviselő Kortizon C-3 helyen karbonil konjugált kettős kötéssel C-17-hez szénhidrogén lánc kapcsolódik C-20 karbonillal és α-helyzetű hidroxil csoport C-11 karbonil MINERALOKORTIKOIDok képviselője Aldosteron C-3 helyen karbonil konjugált kettős kötéssel C-17-hez szénhidrogén lánc kapcsolódik C-20 karbonillal C-21 hidroxilezve C-11 β-helyzetű hidroxil csoport ANDROGÉNek képviselője Testosteron C-3 helyen karbonil konjugált kettős kötéssel C-17-hez csak β-helyzetű hidroxil csoport kapcsolódik ÖSZTROGÉNek képviselője Ösztradiol C-3 helyen fenolos hidroxil az aromás A gyürűn C-17-hez csak β-helyzetű hidroxil csoport kapcsolódik A máj epetermelésének befolyásolása 12

13 ÉLETTANILAG JELENTŐSEBB SZTEROID-VEGYÜLETEK A KOLESZTERIN az emlős szervezet nélkülözhetetlen szteroid terméke g az ideális szint; naponta a májban termelődik. 20 %-a szabad formában, zömében azonban linolénsavval észterezve fordul elő általában 0.5g kiürül a széklettel Epesavak 3-5 g vesz részt az emésztésben; naponta 0.8 g képződik koleszterinből a májban (a többi recirkulál) Koleszterin-szint szabályzók (sztatinok): Xeter; Crestor; (10 mg) HORMONHATÁSÚ SZTEROIDOK A szteroid hormonok zömmel fehérjéhez kötve cirkulálnak a szervezetben hormonhatása azonban csak a szabad hormonnak van hatását a receptor fehérjéhez kötődve fejti ki A mellékvesekéreg hormontermelése genetikailag meghatározott amit adott esetben szerkezeti analógokkal, céltermékekkel befolyásolni lehet Kortikoszteroidok 21 C atomot tartalmaznak - Mineralokortikoidok az elektrolit forgalmat befolyásolják (11 desoxi-kortikoid) - Glükokortikoidok az intermedier szénhidrátforgalmat befolyásolják (11 oxikortikokoid) - Az aldoszteron erélyes mineralo és gyenge glükokortikoid hatású - 9a-fluorokortizolban a fluor sokszorozza a mineralokortikoid hatást - A prednizolon ( 1 ) csökkenti a mineralokortikoid hatást, erősíti a gyulladás csökkentő hatást - A progesteron a szexuálhormon hatás mellett csekély mértékben kéreghormonhatású Androgének Nemi jellegre ható hormonhatású szteroidok 19 C atomot tartalmaznak A mellékvese-kéregben és a herében termelődnek A női szervezetben csak a kéreg termeli Ösztrogén hatású hormon 18 C atomot tartalmaz, aromás A gyűrű Az ováriumban termelődik koleszterinből illetve 17-hidroxi-progeszteronból; A hormonhatású szteroidok zöme a májban inaktiválódik, A bomlástermék végül szulfát illetve glükuronid formában ürül a vizelettel 13

14 SZTEROIDOK DEHIDROGÉNEZÉSE (a 1 reakció) A szteroidok 1,2-dehidrogénezését az ötvenes évek első felében Fried, Thoma és Klingsberg (J. Am. Chem. Soc :5764), valamint Vischer és Wettstein (Experientia :371) fedezte fel. Néhány év mulva, 1959-ben Levy és Talalay Pseudomonas testosteroni-ból dehidrogénező enzimkeveréket izolálva (J. Biol. Chem :2014) valószínűsítették, hogy itt direkt dehidrogénezés történik hidroxilezett átmeneti termék képződése nélkül. Az in vitro dehidrogénező reakció nem igényelt oxigént. Megállapították, hogy a fenazinmetoszulfát hatásos oxidáló ágensként alkalmazható az esetben, ha az enzimkészítménnyel anaerob körülmények között dolgoztak. Más dehidrogénezésben használható faktorok (NAD +, NADP +, FMN, FAD, citokrom-c, ferricianid ion) használhatatlanok voltak. Ezek az eredmények, Talalay azon megfigyelésével együtt, hogy az 1α-hidroxi, illetve a 2β-hidroxi származékok nem alakulnak dehidrogénezett származékká, megerősítette a közvetlen dehidrogénezésre vonatkozó elképzeléseket. A dehidrogénezési mechanizmust erősítették meg Gale és munkatársai, amikor kimutatták Bacillus sphaericus-ban a prednizolon képződéshez szűkséges chinon szerű faktort, a K 2 vitamint. Bacillus sphaericus sejtmentes kivonatában ugyanis ez az enzim nem működik, de a K 2 vitamin, menadion, illetve hexahidro-koenzim-q hatására a dehidrogénező aktivitás újra megjelenik (Biochemistry :788). Talalay szerint a chinon szerkezetű vegyületek csak másodlagos szerepet játszanak a redukciós ciklus folyamatában. Az igazi koenzim valamilyen flavin szerkezetű vegyület. Iica és munkatársai olyan sejtmentes enzimkészítményt állítottak elő Bacillus cyclo-oxidans-ból, amelyik reverzibilisen 1,2- dehidrogénezést és 1,2- hidrogénezést katalizált. Az enzimszint növelhető volt a tápközegbe adott androszta- 1,4-dién-3,17-dionnal. A dehidrogénezést menadion stimulálta, a hidrogénezés pedig NADH jelenlétét igényelte. (Steroids Suppl :159) A két enzimaktivitást a hagyományos fehérjetisztítási módszerekkel eddig elválasztani nem sikerült. A szteroid 1,2-dehidrogenáz működését sztereokémiai szempontból Bacillus sphaericus-nál Ringold, Hayano, valamint Stefanovic tanulmányozták részletesen (J. Biol. Chem :1960). Az enzimkészítményük az 1β és a 2α szubsztituált származékokat dehidrogénezte, viszont az 1α és a 2β szubsztituáltakat nem alakította át, amiből következtethető, hogy a dehidrogénezésben az 1α és a 2β helyzetű hidrogén atom vesz részt. Ezt Ringold azon tapasztalata is megerősítette, hogy a dehidrogénezési reakció után is megmaradt az 1β deuterium, míg az 1α deuterium eltünt. 14

15 Az enzim három aktív hellyel egy protondonor, egy protonakceptor és egy hidrid akceptor tulajdonságú ponttal közeledik a szubsztrátumhoz. A protondonor és a protonakceptor hely hatására egy enolizációs folyamat indul a 3-ketonon. Ezt követi a hidrid átvitel az 1α helyzetből, ami végül is az enzimszubsztrátum komplex szétesését vonja maga után. A fermentációs közegben az enolizációt izotopkicserélési módszerrel bizonyították, ami igazolta, hogy az enolizáció a dehidrogénezési reakció szűkséges lépcsője. Az 1,2-dehidrogénezési reakció ( 1 reakció) kényelmes lehetőséget teremtett a 19- norszteroidok aromatizálására (J. Am. Chem. Soc :4627). Ennek ipari jelentősége nagy mértékben csökkent a hatvanas években a norszteroid totálszintézis kifejlesztése és ipari gyakorlatba vétele után. A Bacillus sphaericus-ból nyert kivonat az 5β-androsztán-3,17-dion dehidrogénezésére nem alkalmas, viszont az 5α-androsztán-3,17-dion esetében eredményes. Ezt az eredményt a két vegyület enolizációs képességében rejlő különbség magyarázza. Az 5β származék esetében ugyanis az enolizáció a 4-es szénatom irányába következik be, az 5α származék esetében viszont a 2-es szénatom van kitüntetett helyzetben, ami az 1α helyzetű hidrogén eliminációját segíti. Jerussi és Ringold későbbi vizsgálatai azt mutatták, hogy az enolizációt katalizáló enzim kapcsolatban van egy flavint, illetve más redox rendszert tartalmazó olyan fehérjével, amelyik képes az axiális helyzetű hidrid-ion elvonására az 1-es szénatomról. A telített pregnán dehidrogénezésekor főtermékként minden esetben 1,4-dien származék képződik. A melléktermékként jelentkező 4-androsztén-3,17-dion utólagos telítés eredménye lehet. Fonken és Murray Streptomyxa affinis-szel dehidrogénezték az 5β-pregnán-3,11,20-trion-20-ketált. Ez az eredmény lehetőséget adott a dehidrogénezésben szereplő hidrogén térhelyzetét részletesen vizsgálni 5α-pregnán, illetve 5β-pregnán szerkezetű szubsztrátum esetében. Mindkét pregnán származékban az 1β-hidrogént és a 2αhidrogént deuteriumra cserélték. Az 5α-pregnán- 3,11,20-trionból az 1α,2β helyzetű, az 5βpregnán-3,11,20-trionból viszont az 1β,2β helyzetű szubsztituensek távoztak a dehidrogénezési folyamatban, ami nem meglepő, ha összehasonlítjuk az 5α-pregnán és az 5βpregnán enolizált A gyürüjének térszerkezetét. A dehidrogénezés utolsó momentumaként távozó hidrid ugyanis mindkét esetben axiális helyzetből kerül a dehidrogenáznak a hidridakceptor pomtjára. (J. Org. Chem :1102) Dehidrogénezés a szterán váz különböző (1-, 4-, 5-, 7-, 9-, 14-, 16-) helyein figyelhető meg. Az első ilyen reakciót Horváth János és Krámli András írta le 1947-ben. Azotobacter tenyészetben koleszterolból 7-dehidrokoleszterol képződött (Nature :633). Több közlemény nem jelent meg erről a reakcióról, noha a szakirodalom a mikrobiológiai konverzió mérföldköveként emlékezik meg róla. 15

16 Ezen a téren a kutatás a már említett Fried, Thoma és Klingsberg eredményeinek közlésével indul, amelyben megállapítják, hogy a Cylindrocarpon radicicola tenyészlevében a progesteronból, testosteronból és a Reichstein féle S-vegyületből egyaránt 1,2- dehidrotestololakton képződik. Streptomyces lavendulae viszont progesteronból androszta- 1,4-dien-3,17-dion és 1,2-dehidro-testosteron keverékét állítja elő. A kortikoidok 1,2-dehidrogénezésére 1954 óta világszerte a Corynebacterium simplex, illetve az érvényben levő taxonomiai elnevezés szerint Arthrobacter simplex ATCC 6946 törzset használják. Genetikailag stabil clon-ként élesztőkivonatot és glükózt tartalmazó táptalajon az aktivitás elvesztése nélkül fenntartható. Szabadalmi és gazdasági meggondolásból néhány más törzs biokémiai aktivitását is hasznosítják: Bacterium cyclooxidans ATCC 12673(1958), Bacillus lentus ATCC (1963), Bacillus sphericus ATCC 7055 (1955), Mycobacterium globiforme 193 (1959), Streptomyxa affinis ATCC 6737 (1958). Ezek fontos szerepet kaptak a gyulladást csökkentő szteroid gyógyszerek előállítását szolgáló eljárásokban. Élesztőkivonatot, kazein-hidrolizátumot, kukoricalekvárt, glükózt tartalmazó táptalajon változatlanul az aktvitás maximális szintjét teljesítik. A tenyésztése ph tartományban 28 C-on, aerob kürülmények között óra alatt befejeződik. Az enzim indukcióját a tápközegben szennyezésként nyomokban jelenlevő növényi szteroidok végzik. Az enzimszint értékét előnyösen befolyásolja a növekedési periodus utolsó szakaszában adagolt szubsztrátum. Még ma sem vesztett jelentőségéből a mikrobiológiai eljárás, jóllehet az 1,2-dehidrogénezés kémiai úton is végrehajtható. A 1 dehidrogénezést végző mikróbák általában a szteroid váz lebontására szolgáló enzimekkel (például 9α-hidroxiláz aktivitással) rendelkezve, nemkívánatos veszteséget okozhatnak. Ez a veszély megfelelő enzimhiányos mutáns alkalmazásával, vagy specifikus hatású gátlószer alkalmazásával kivédhető, de kellően magas dehidrogénező szint esetében a biokonverzót végző tenyészet hígitott formában való használata is eredményre vezethet. A tenyészközegbe adagolt induktor ugyanis olyan 1,2- dehidrogenáz szintet eredményezhet, ami a biokonverzióra használható tenyészet százszoros higítását is lehetővé teszi. Ez utóbbi esetben gyakorlatilag csapvízben, tápanyag nélkül folyik a dehidrogénező folyamat. Sejtnövekedés hiányában a szteroid váz lebontására szolgáló enzimek képződésére nincs lehetőség. Nemkívánatos mellékreakcióként bekövetkezhet a pregnán oldallánc keto csoportjának a redukciója. Ez a NADH kofaktort igénylő enzim ugyancsak indukálható. Ezért a növekedési periódusban induktorként célszerű pregnán oldalláncot nem tartalmazó vegyületet (például androszténdiont) használni és az átalakítási szakaszban klóramfenikol adagolásával akadályozni az induktív enzim képződését. A dehidrogénezési folyamathoz nem szükséges élő sejt. Sikerrel használható az indukált tenyészetből előállított acetonos száraz sejtpor. Ekkor azonban elektronakceptorként fenazin-metoszulfát jelenléte szűkséges. Folyamatos tenyésztéssel tartósan fennntartva a növekedési állapotot, magas aktivitású sejttömeg állítható elő. A nagy aktivitású sejttömeg előnyösen használható hidrokortizon kristályszuszpenzó prednizolon kristályszuszpenzióvá alakítására. Literenként g hidrokortizon kristályt tartalmazó szuszpenzióból 95%-os hatásfokkal képződik a prednizolon kristály. A teljes átalakulást akadályozza a szubsztrátumra kristályosodó prednizolon, mert a zárványként jelenlevő hidrokortizon már nem vehet részt a biokonverziós folyamatban. 16

17 Vischer és Wettstein közléséből megtudhatjuk, hogy a Fusarium solani és a Fusarium caucasicum feltűnő aktivitással állítja elő az androsta-1,4-dién-3,17-diont a dezoxikortikoszteronból, 4-androstén- 3,17-dionból, és progeszteronból. Tovább oxidálva végül tesztololakton származékok képződnek. Wix és Albrecht vizsgálatai szerint Fusarium tenyészettel a telített szteroidok, valamint a 3β-hidroxi-5,6-dehidroszteroidok is androsta-1,4-dien-3,17- dionná alakíthatók. (Acta Microbiol. Acad.Sci. Hung :339). A telített szteroidok 4-es helyzetben történő dehidrogénezését először Vischer és Wettstein írták le. Hasonló átalakításról számolnak be a pregnán sorban Nocardia blackvellivel, Stoudt és munkatársai (Arch. Biochem. Biophys :280), valamint Coronelli, Kluepfel és Sensi Corynebacteriun simplex sejtjeit használva (Experientia :441; :208). {*Laskin és Fried szerint az eredetileg hidroxilező fajként leírt Glomerelle fusaroides 9,(11)- helyzetben képes az ösztront dehidrogénezni (U.S. Patent , )}. Az epesavak dehidrogénezésére Hayakawa és munkatársai Streptomyces és Corynebacterium törzseket használtak (J. Biochem. Tokyo :108; :4319; 45:633). A digitoxigenin dehidrogénezése 16-os helyzetben Trichotecium reseum-mal végezhető (J. Biol. Chem :3399). Ez az organizmus 17αhidroxilező aktivitásával vált ismertté. Lehetséges, hogy a digitoxigenin labilis 17α-hidroxi származékából spontán vízkilépéssel képződik a dehidrogénezett végtermék. Herzog és munkatársai a Wojnovicia graminis tenyészlevében is megfigyelték a tesztoszteron dehidrogénezését 14-es helyzetben a 12α-hidroxilezéssel egyidejüleg. Általános azonban az a vélemény, hogy ez nem enzimes folyamat eredménye, hanem a mikrobiális hidroxilezés közben a 12α,14α-diolból képződő műtermék, amely a labilis 14α-hidroxicsoport spontán eliminációjának az eredeménye. Az elimináció mozgató ereje a megjelenő hidroxilcsoportok 1,3- diaxiális kölcsönhatásából adódik. Ezt a megállapítást, a 14α-hidroxi-tesztoszteron melléktermékként való megjelenése támogatja (J. Org. Chem :2177). 17
. A telített szteroidok 4-es helyzetben történő dehidrogénezését először Vischer és Wettstein írták le.")
18 ALKOHOLOK OXIDÁCIÓJA KETONNÁ vagy ALDEHIDDÉ A hidroxiszteroidok reverzibilis oxidációs folyamatait Talalay és munkatársai a Pseudomonas testosteroni indukálható enzimein tanulmányozták részletesen. Elválasztották a 3α-hidroxil csoportot oxidáló enzimtől a 3β- és 17β-hidroxil csoportokat oxidáló enzimet. Megállapították, hogy mindkét enzim NAD + kofaktort igényel működéséhez, viszont szubsztrátspecificitás szempontjából markáns különbséget tehetünk közöttük. Specifikus az enzim a sztereokémiai viszonyokra, függetlenül attól, hogy a hidroxil csoport az adott esetben axiális vagy ekvatoriális helyzetben van. ( The Enzymes Vol. 7:177 Acad. Press New York 1963) Ringold és munkatársai szerint a Pseudomonas testosteroni esetében a 6-os helyzetben levő elektronszívó csoportok a megfordítható reakció egyensúlyát befolyásolhatják, akár meg is fordíthatják. A kísérleti eredmények szerint a karbonil csoporton kialakuló elektronsürüség döntő szerepet játszik az egyensúly kialakulásában. Ez nem meglepő, hiszen a folyamat valójában hidrid-ion átvitelét jelenti a szubsztrátum és a NAD + között. Az alkoholok ketonná oxidálása már Bertrand 1896-ban végzett munkáiból ismert. A szteroloknál először Mamoli és munkatársai alkalmazták, amikor 5-androsztén-3β,17β-diolt oxidáltak 4-androsztén-3,17-dionná egy fertőzött élesztőtenyészetből izolált baktériummal (Berichte :2701). Ugyanezzel a tenyészettel oxidálták a 3β-hidroxi-5-pregnén-20-on vegyületet progeszteronná. Az eredményeken felbuzdulva a mikrobiológusok hasonló céllal különböző baktériumok tiszta tenyészetével végeztek hasonló átalakításokat. Kiderült, hogy a Micrococcus dehydrogenans, a Corynebacterium mediolanum, a Flavobacterium dehydrogenans enyhe reakciókörülmények között nem csak a 3β-hidroxi csoport dehidrogénezését, hanem az acetil csoport hidrolízisét és az 5,6-kettőskötés izomerizálását is elvégzik. A 3β-hidroxil csoport dehidrogénezését a levegő oxigénjével regenerálódó szterinoxidázok is sikeresen végrehajtják. A sugárgombákban aktívan működnek mint irreverzibilisen dehidrogénező enzimek. Régiószelektivitásukból adódóan azonban a szteroidok 17β-hidroxil csoportját természetesen nem dehidrogénezik. Mivel nem igényelnek NAD + kofaktort, vizzel nem elegyedő szerves oldószerek, klórozott szénhidrogének jelenlétében is működőképesek. A különböző helyen hidroxilezett szteroid származékok oxidációját számos esetben megfigyelték. Így a 3α-hidroxiszteroidokon kívül a 6β-hidroxi, a 7α-hidroxi, a 11β-hidroxi, a 16α- és 16β-hidroxi, a 18-hidroxi és a 20-hidroxi származék mikrobiális oxidációjára van ugyan irodalmi adat, ezeknek a kémiai oxidációja azonban általában előnyösebb. Sok példát találunk az irodalomban a karbonil csoportok direkt bevezetésére. Feltételezhető, hogy a primer hidroxilezést közvetlenül követő oxidáció eredményét regisztrálják. 18

19 KARBONIL CSOPORT KÖZVETLEN REDUKCIÓJA ALKOHOLLÁ Az alkohol oxidációjával foglalkozó fejezetben már találkoztunk ezzel a sok esetben megfordíthatónak tekinthető reakcióval. Ha feltesszük, hogy sebességmeghatározó faktor a hidrid-ion átvitele a NADH-ról a karbonil csoportra, akkor nyilvánvaló, hogy az elektronsürűség csökkentése a karbonilcsoporton, kedvez a redukciós folyamatnak. Ringold és munkatársai kimutatták, hogy a Pseudomonas testosteroni sejtmentes kivonata a 2α-metil, illetve a 6β-metil-tesztoszteron 3-as ketocsoportját nem redukálja (Biochem. Biophys. Res. Comm :162). Megváltozik azonban a helyzet elektronszívó csoportok jelenlétében, a 2α-, 4α-, 6α-, vagy 6β- helyzetben fluorozott, vagy a 4-es helyzetben klórozott tesztoszteronból 3α- és 3β-hidroxi származékok keveréke képződik. Robinson, Bruce és Oliveto szerint Flavobacterium dehydrogenans nem redukálja a szubsztituálatlan 17-ketoszteroidokat, viszont a 16,16 difluoro-ösztron metilétert 17α-hidroxi származékká alakítja. Az erősen elektronegatív elem jelenléte elősegíti a hidrid-ion átvitelét a karbonil csoportra (J. Org. Chem :975). A szteroid vegyületek 3-as és 17-es ketocsoportjainak élesztővel történő redukcióját Mamoli és Vercellone írta le 1937-ben (Berichte :470; 70:2073). A redukciós folyamatok katalizálására azért használják az élesztőt, mert általában nem okoz más átalakítást. Az élesztők általában nem redukálják a konjugációban levő ketocsoportokat, bár az 5α-1- androszten-3,17-diont a Saccharomyces cerevisiae 5α-androsztán-3,17-diollá redukálja. A mikrobában általában két NAD + függő hidroxiszteroid dehidrogenáz felelős a ketocsoportok redukciójáért. A szubsztrátum szerkezetétől függ, hogy melyik végzi el a redukciót. Az A-B gyürú transz anellációja a β-enzim működésének kedvez, ezért főleg 3βhidroxiszármazék képződik Ezzel szemben az A-B gyürű cisz anellációja a 3α-hidroxi származék képződését segíti. Sok esetben a redukálandó ketocsoporttól távolabb levő szubsztituens változása megváltoztathatja a biokonverzió lefolyását. Saccharomyces cerevisiae jelenlétében az 5α- illetve az 5βdihidro-11-keto-progeszteronból egyaránt a 3α-hidroxiszármazék képződik, függetlenül az A-B gyürű kapcsolódásától. Ugyanakkor 3β,11α-dihidroxi-5α-dihidroprogeszteron képződik élesztő jelenlétében a 11α-hidroxi-5α-pregnánolonból (Helv.Chim.Acta :1945). A szteroid vegyületekben előforduló ketocsoportok redukciójára is bőven találunk adatot a szakirodalomban. Az epesavakban előforduló 7-keton redukcióját 7α-hidroxillá Streptomyces gelasticus-szal és Corynebacterium fajokkal írták le (Proc. Japan Acad./1957/ 33:221). 19
.")
20 A 9,10-szeko vegyületekben a 9-ketonok 9α-, illetve 9β-hidroxilezett termékké alakulnak Nocardia restrictus, illetve Mycobacterium fajok tenyészlevében (Biochemistry :1238). A 16-os helyen megjelenő oxigénfunkció a lehetséges oxidált és redukált formában jelenik meg, feltehetőleg enzimes és spontán átalakulási folyamatok eredményeként. A 19-karbonil redukcióját Penicillium thomii aktivitásaként Sih és munkatársai közölték 14β-hidroxi-3,18- dioxo-4,20/22/-kardenolid szubsztrátumon. Az egyensúly az alkohol irányába tolódik (J. Org. Chem :854). A pregnán oldallánc 20-ketocsoportjának redukciójával több kutatócsoport foglalkozott. Először Fried, Thoma és Klingsberg írta le Streptomyces lavendulae-val végzett biokonverziós folyamatok nem kívánatos mellékreakciójaként. Hübner és munkatársai Streptomyces hydrogenans-ból izolálták azt az indukálható 20β-dehidrogenázt (Biochem. Biophys. Acta :270), amely széles szubsztrátspecifitása miatt a kortikoidok előállítását célzó biokonverziós eljárások nemkívánt melléktermékének képződését katalizálja. Prokariótákban és fonalas gombákban fordul elő ez az enzim. Algákban, élesztőkben és az emlős szervezetben a 20α-dehidrogenáz működik, így a Rhodotorula longissima tenyészlevében prednizolonból, annak 20α-hidroxi származéka képződik (J.Org.Chem :695). A 22-aldehidek redukcióját először Murray és Peterson írták le Rhizopus fajok esetében, de a Mycobacterium tenyészlevében is megtalálható az oldallánc-lebontás melléktermékei között 20-oximetilpregnán formájában. Megjelenésükből a mikroszervezet sejten belüli oxidoredukciós állapotára következtethetünk (J. Am. Chem. Soc. /954.76:5673). 20
. A pregnán oldallánc 20-ketocsoportjának redukciójával több kutatócsoport foglalkozott.")
21 SZÉN-SZÉN KETTŐSKÖTÉS TELÍTÉSE Telitetlen kötések telítésére szolgáló reakciók két csoportban tárgyalhatók Külön tárgyalandó az izolált kettős kötés redukciója a konjugált rendszerek telítésétől. Nem azonosított féceszből izolált anaerob baktériumokkal a koleszterint 5β-kolesztánná, másnéven koprosztánná lehet redukálni (Arch. Biochem. Biophys :213). A kolsav átalakítása 7-dezoxiepesavvá ugyancsak bendőbaktériumokkal végezhető. Amennyiben a redukálandó kettőskötés keto-csoporttal van konjugálva, a dehidrogénezésnél megismert szabályok érvényesek, azaz a belépő hidrogénatom mindig axiális és transz az 1α,2β-szénatomon. A reakció egyensúlyra vezető természetéből következik, hogy a triamcinolont a Bacillus cyclo-oxidans részben 1,2-dehidro-triamcinolonná redukálja (J. Biol.Chem :965). Konjugált kettőskötés redukcióját Mamoli és Schramm rothasztó baktériumok tenyészetét használva figyelték meg. A Bacillus putrificus vagy a Schubert által használt Clostridium paraputrificum anaerob körülmények között a 4-androszten-3,17-dionból 3-keto-5β-androsztán, és 3α-hidroxi- 5β-androsztán származékok keverékét állítja elő (Z. Naturforschung b:284). Élesztővel az 1,2-dehidro-3-keto szteroidok redukciója 3β-hidroxi-5α-androsztán képződéséhez vezet. A 4,5-kettőskötés telítésére külön enzim szolgál, ami lehetőséget ad 3-keto- 1,4-dien szerkezetű vegyületekből 3-keto-1,2-dehidro-5β-androsztán származékok előállítására. Különleges esetet képvisel a 16-dehidroprogesteron hidroxilezése Rhizopus nigricans, illetve Aspergillus niger hidroxilező képességeinek felhasználásával. Mindkét esetben ugyanis a 11α-hidroxiprogeszteron 17α-epimerje képződik, annak ellenére, hogy a pregnán oldallánc β-helyzete termodinamikailag stabilabb. Az egyik észszerű magyarázat szerint a β-oldalról közelítő enzimfelületen hidrogénaddicióval fejeződik be a reakció, ami a 17-es oldallánc kezdeti planaritását a hidrogénaddicióval kvázi axiális szerkezetüvé módosítja. Ugyanez az eredmény akkor is, ha a jelenséget egy megfordított Ringold mechanizmusként értelmezzük, feltételezve egy átmeneti enol forma létezését, ami spontán protonálódik még az enzimfelület elhagyása előtt. Itt a β-oldalról közelítő enzimfelület nem katalitikus, hanem csak szterikus szerepet játszik (Berichte :1847; :96). 21
22 A KETTŐS KÖTÉS IZOMERIZÁCIÓJA Mamoli 1939-ben írt közleményében (Ann. Rev. Biochem :352) a Corynebacterium mediolanum-mal végzett 3β,21-dihidroxi-5-pregnen-20-on.21-acetát dezoxikortikoszteronná alakítására vezető reakciósorban a dehidrogénező reakciót követő lépésként említi a szteroid izomeráz megfordítható működését. Általában a mikroorganizmusok, amelyek az oxidációs reakciót képesek elvégezni, izomerizálással fejezik be konverziós tevékenységüket. Az irodalom egyetlen esetet ismer, amikor egy Actinoplanes missuriensis a dehidrogénezést követően nem izomerizálta a reakcióterméket. Az enzimkatalizálta izomerizáció mechanizmusát Talalay és Ringold tanulmányozták részletesen. Pseudomonas testosterini-ből nyert indukálható enzimet tisztították, kristályos formában előállították. Az aktív centrumban levő hisztidin imidazol csoportjának fontos szerepét tisztázták( ). A Pseudomonas enzim az 5,10 kettőskötést is átfordítja kialakítva a 3-keto-4,5-dehidro szerkezetet. Deuterium-oxiddal végzett kísérleteikben megállapították, hogy nem hidrogénfelvétel, hanem közvetlen (intramolekuláris) hidrogénátvitel történik a 4-es szénatomról egy átmenetileg létező 8 tagú, illetve 10 tagú gyürűs szerkezetű köztes állapoton keresztül a 6-os szénatomra. Az imidazol csoport szerepe Mellékveséből előállított nyers kivonatban a dehidro-epiandroszteron képződés első lépése a rendszer dekonjugációja, a kettős kötés vándorlása 5-ös helyzetbe és ezt követi a redukciós folyamat. 22
23 ÉSZTERÁZOK REVERZIBILIS MŰKÖDÉSE A szteroid molekula hidroxil csoportjainak a védelmére széleskörüen alkalmaznak kémiai eljárásokat, így az acilezést is. A védőcsoportok eltávolítása sok esetben a mikrobiológus feladata. Az enzimek sztereospecificitása miatt a sztereoszelektív észterhidrolízis sok esetben racemátok szétválasztására is alkalmazható. A 3-keto-1,4-dien szerkezet kialakítása közben általában az észterkötések is hidrolízálódnak. Legkönnyebben a primer alkoholok észterei, amit sorrendben a szekunder acetoxi csoport hidrolízise követ. Végül a molekulán belüli acilvándorlást követően a tercier acetoxi csoport hidrolízise is bekövetkezik. A 21-acetát hidrolízise volt az egyik legrégebben felfedezett szteroidészter hidrolízis. Mamoli Corynebacterium mediolanum aktivitását hasznosította dezoxikortikoid előállításához 21- acetoxi-pregnenolonból. Az észterek hidrolízise szteroid alkoholokká részleteiben nem ismert, noha legtöbb esetben specifikus észteráz hatással találkozunk. Az eddig vizsgált észtert bontó enzimek hisztidint tartalmaznak, továbbá az ismert észterázt gátló vegyületekkel a szteroidésztert bontó enzimek működése is akadályozható. A hidrolízis közben az enzimet az észter savkomponense átmenetileg acilezi, amit egy újabb vízmolekula távolít el az enzimfelületről. Az észterázok reverzibilis működése miatt a tenyészlébe adagolt szteroidokat sok esetben acilezi a mikroszervezet. Az élesztők esetében gyakran a 17-hidroxi származék acetátjával találkozunk. A Streptomyces roseochromogenes amelyet sikerrel használnak hidroxilező szervezetként 16-hidroxikortikoidok előállításához a 21- amino-9α fluoro-11β,17α-dihidroxi- 1,4-pregnadien-3,20-dionból triamcinolon.21-n-acetátot állít elő. A triamcinolon acetonidját például a Trichoderma glauka képes acilezni 21 származékká. 23
24 SZTEROIDOK HIDROXILEZÉSE A mikroorganizmusokban hidroxil csoportok kiépítésére számos példát ismerünk. Leggyakrabban telítetlen kötés kialakitását követő vizfelvétel ad lehetőséget a további feladat teljesítésére kész köztestermék, a hidroxilezett származék megjelenésére. A jól ismert Szent-Györgyi Krebs ciklus enzimei közt szereplő FAD igényes szukcinát dehidogenáz[7] hatására képződő fumársav vízfelvétellel [8] alakul almasavvá, amely NAD + kofaktort igénylő reakció [9] során alakul oxálecetsavvá, a trikarbonsav ciklus nélkülözhetetlen termékévé.. A zsírsav lebontás (béta-oxidáció) hidroxilezett köztes termékének képződését is megelőzi a [6] FAD igényes acil-coa dehidrogenáz működése, a telítetlen kötés kialakítása, amit követ a vízfelvétel [4], és a NAD + igényes β-oh-acilcoa dehidrogenáz [7] működése. ZSÍRSAV SZINTÉZIS ÉS LEBOMLÁS ÖSSZEHASONLÍTÁSA 1); Malonil transzferáz (működését a jelenlevő zsírsavak alloszterikusan gátolják) 2); β-ketoacil-coa szintház 3); D-β-ketoacil-CoA reduktáz 4); D-β-hidroxi-acil-CoA dehidratáz 5); acil-coa reduktáz 6); FAD igényes acil-coa dehidrogenáz 7); NAD + igényes β-oh-acil-coa dehidrogenáz 8); ketoacil-coa tioláz * a transeszteráz az igényeknek megfelelően segít felhasználni a képződött zsírsavat 24
25 Bloom és Shull megállapította, hogy az axiális hidroxilezésre képes mikroorganizmus adott körülmények között képes a kettőskötésben levő szénatomot támadva epoxid képződést katalizálni (J. Bacteriol :382). Az oxiránok előállítására a kémiai módszerek ugyan előnyösebbek, mégis a reakció tanulmányozása a hidroxilezés biokémiai mechanizmusának jobb megértését segíti. Az epoxid-oxigén konfigurációja a hidroxil csoport konfigurációjával megegyezik. A kettőskötés π-elektronjai az axiális CH kötés elektronjaihoz hasonló orientációt mutatnak, miközban a szénatomot támadó, oxigént irányító hidroxilező enzimkomplex nem tud különbséget tenni a két helyzet között. A Curvularia lunata a 9,11-dehidro-Reichstein-S vegyületbe 9β,11β-epoxidot visz be, míg a 14,15-dehidro származékból 14α,15α-epoxid képződik. Más axiális hidroxilezők, így a 11-es helyen hidroxilező Cunninghamella blakesleana, vagy a 14-es helyzetben hidroxilező Helicostylum piriforme, valamint a Mucor griseocíanus ugyancsak megfelelő oxirán származékot alakít ki a dehidrogénezett szubsztrátumból. A Nocardia fajok aktivitása a 9α-hidroxilező képességükből következően a 9,11-dehidroszármazékból 9α,11α-epoxidot hoz létre (Gazz. Chim. Ital :260). Epoxidok egyszerű hidrolízisét Saccharomyces cerevisiae tenyészetében először Camerino írta le példaként a diaxiálisglikolkénti felnyitásra. A mechanizmus valójában egy savkatalizálta epoxidfelnyitáshoz hasonlít. A kombinált protonforrásként imidazolt tartalmazó enzim nukleofil támadóként jelenik meg. A víz nem tekinthető független résztvevőnek, inkább egy hidrogénkötésekkel összetákolt agglomerátumnak (Berichte :1863). Speciális esetben az epoxid felnyílás egy Wagner-Meervein átrendeződést von maga után, amit a szteroidszintézisben érdekelt vegyészek hasznosíthatnak. A folyamat teljesen megegyezik az enzimtől független, savkatalizálta felnyitáskor bekövetkező folyamattal. Ilyen speciális esetnek tekinthető a 16,17-oxidoprogeszteron konverziója Saccharomyces cerevisiae sejtjeivel, miközben a metil csoport a 13-as helyről a 17-es szénatomra vándorol. Az átrendeződés azonban egyszerű kémiai folyamat, ami spontán követi az oxirán enzimkatalizálta felnyitását. 25
26 A szteroidok C 11 helyzetben bekövetkező mikrobiális axiális hidroxilezése minden esetben a légköri oxigén beépülését jelenti, amit kísérletileg Hayano és Dorfman bizonyítottak. Jelölt vízzel /H 18 2 O, illetve D 2 O/ normál oxigén tenzióban végigvitt hidroxilezés esetén a termék jelöletlen maradt. Nehéz oxigén-atmoszférában viszont a 18 O beépülése bizonyítást nyert. Megállapították, hogy a 9/11/-telítetlen szteroidok nem prekurzorai a 11β-hidroxi származékoknak. Ezek a megállapítások a 6β-, 11α-, 11β-, 12β-, 15α-, 17α-, és 21-hidroxi csoportoknak a kiépítésére érvényesek. Hayano, Peterson, Corey, Gregorian, kísérletileg bizonyították, hogy a szénatom térhelyzete a szubsztitució alatt nem változik. Rhizopus nigricans-szal végzett kísérletekben 11αhidroxi csoportot vittek be pregnándionba, amely 11α- és 12α-helyzetben triciálva volt A hidroxilezés közben a 11α tricium elveszett, a 12α tricium viszont változatlanul maradt. Egy másik kísérletben 11β-Deuterium tartalmú pregnándiont hidroxileztek. Az eredmény a megfelelő 11α-hidroxipregán-3,20-dionban a 11β Deuterium változatlanul megmaradt. Ezek a kísérletek egyértelműen bizonyították, hogy a hidroxil bevitelben nem érintett hidrogén térhelyzetét változatlanul megtartja. Ringold elképzelése szerint a hidroxilezés a 2β-, 6β-, 10β-, 17α-hidroxilezés esetében egy enolizáció eredményeként létrejövő aktívált állapoton keresztül történhet. Az enolizáció is az enzim hatására következik be, amit az enzim által aktívált, nemdisszociált oxigén támad. Az első lépés egy hidroperoxid képződés, amit egy újabb enzimkatalizálta reakció tesz teljessé. A hidroperoxid NADPH segítségével hidroxillá alakul, egy molekula víz egyidejű képződésével. A hidroperoxid jelenlétét és az utána 10β- és 17α-helyzetben következő redukciós folyamatot Barton és munkatársai igazolták. A szteroid váz szinte minden szénatomja, a 3-as, 4-es és 13-as szénatom kivételével mikrobiális úton hidroxilezhető. A 3-as szénatomhoz a természetes szteroidokban minden esetben oxigén kapcsolódik, a 13-as kvaterner szénatomhoz viszont nem kapcsolódhat oxigén. Azokon a helyeken, ahol enolizációra nincs mód, ott más mechanizmus működését kell feltételezni. A mikróbák természetesen nem szűkítik tevékenységüket egyetlen reakció elvégzésére, hanem a térszerkezeti analógiák szerint a szteroid váz más pontjain is 26
27 végrehajthatnak hidroxilezést, sőt a mikróba enzimkészletének megfelelően egyéb reakciók végrehajtására is sor kerülhet. A SZTERIN VÁZ HIDROXILEZÉSE 1α Penicillium fajjal 1α-hidroxi-4-androsztén-3,17-dion dehidroepiandroszteronból Nocardia fajokkal pregnán-származékok 1α-hidroxilezése 1β Rhizoctonia ferrugene Reichstein féle S vegyület 1β- és 2β-hidroxilezése Absidia orchidis digitoxigenin 1β- és 5β-hidroxilezése 2α Nocardia corallina az ethiszteront 1α és 2α-helyzetben hidroxilezi Nocardia italica a noretiszteront 2α és 16α-helyzetben hidroxilezi 2β Rhizoctonia ferrugene 1β- és 2β-hidroxilezés 5β Absidia orchidis digitoxigenin 1β- és 5β-hidroxilezése 6β Mucor félék melléktermékként hidroxilez 6β helyzetben Rhizopus arrhizus a progesteront hidroxilezi 6β- és 11α -helyzetben Chaetomium a prednizont hidroxilezi 6β-helyzetben 7α Phycomyces blakesleeanus a progesteront hidroxilezi 7α-helyzetben 7β Proactinomyces a koleszterin 7β-hidroxilezése (Horváth-Krámli 1948) 8β Cercospora melonis Reichstein-S vegyület 8β-hidroxilezése 9α Helicostylum piriforme Reichstein féle S vegyület hidroxilezése Actinomycetales,Eubacteriales 1,2 dehidrogenáz hiány esetén különben bomlás követi 10β Rhizopus nigricans és egyéb gombák 19-nortesztoszteron 10β- és 11α-hidroxilezése 11α Rhizopus nigricans ATCC 6227 progeszteron 11α-hidroxilezése (14α) Aspergillus ochraceus NRRL 405 progeszteron 11α-hidroxilezése (14α) 11β Curvularia lunata hidrokortizon előállítása Cunninghamella blakesleeana hidrokortizon előállítása 12a Wojnowicia graminis testoszteronból 12α- és 14α-hidroxilezett terméket készít 12β Fusarium lini digitoxygenin digoxigeninné alakítása Streptomyces purpurascens digitoxin digoxinná hidroxilezése 14α Mucor félék Reichstein féle S vegyület 14α-hidroxilezés 15α Colletotrichum antirrhini progesteron 15α származéka Giberella baccata dezoxicorticosteron 15α-hidroxilezése 15β Phycomyces blacesleeanus progesteron 15β-hidroxilezése Lenzites abietina desoxicorticosteron 15β-hidroxilezése Bacillus megaterium Reichstein S vegyület 15β-hidroxilezése 16α Streptomyces roseochromogenus 9α-fluoro hidrokortizon 16α-hidroxilezése 16β Helicostylum piriforme digitoxigenin 16β-hidroxilezett származéka 17α Trichotecium roseum desoxicorticosteron 17α-hidroxilezése 18 Corinespora cassiicola corticosteron aldosteronná alakítása 19 Corticium sasakii a 19-metil hidroxilezése 21 Ophiobolus herpotrichus csak a természetes d-enantiomert hidroxilezi A gazdaságossági szempontok figyelembe vételével a technológia kialakításakor kell feloldani ezt a nemkívánatos helyzetet, és a mikróba kiválasztásával, a technológiai körülmények megválasztásával, esetleg a szubsztrátumon végzett kémiai átalakítással lehet biztosítani a célvegyület főtermékként való képződését. 27
28 A szteroid C 11 helyzetben történő hidroxilezésének folyamata valójában több lépésre különíthető: - az oxigén aktíválása, - a szubsztrátum aktíválása - az oxigén átvitel, végül -a redukált koenzim szerepe -és a regenerálása. Az nyilvánvaló, hogy a molekuláris oxigén reagál az elektrongazdag centrummal. A hidroxilező fehérje fajra jellemző szerkezetétől függően α- vagy β- oldalról közelítve a szubsztrát molekulát, aktíválva a CH kötést, kvázi ionos karakterüvé alakítja. A kötés nem szakad fel teljesen, de jelentős mértékben polarizálódik, ami lehetővé teszi a nemdisszociált, de fellazult kötésű oxigénmolekula beépülését. A hidroperoxid intermedier tovább alakításához Cooper, Estabrook és Rosenthal vizsgálatai szerint ekvivalens mennyiségű redukált kofaktor használódik fel. A regenerálás miatt az intermedier anyagcsere működőképessége nélkülözhetetlen feltétele a hidroxilezési folyamatnak. 28
29 KORTIKOIDOK ELŐÁLLÍTÁSÁT SZOLGÁLÓ EREDMÉNYEK A kortizon gyulladásgátló hatásának a felismerése a század közepén a gyógyszerkutatással foglalkozók figyelmét a kortikoidok előállítására irányította. Sarett 1946-ban megjelent dolgozatában közölte ugyan a kortizon kémiai szintézisét, ennek azonban csak elméleti jelentősége volt. A Searle cég anyagi támogatásával Hechter 1949-ben izolált mellékvese készítménnyel kortexonból (11β-hidroxilezéssel) kortikoszteront állított elő. A pénzt és fáradtságot nem kímélő tevékenység nyilvánvalóvá tette, hogy ez az út sem vezet piaci eredményre. Az irodalmi adatok szerint a szteroid vegyületek 11-helyzetben való hidroxilezése csak biológiai eljárással oldható meg. Gazdasági sikert remélve néhány gyógyszergyár (Upjohn, Squibb, Pfizer, Schering) anyagi támogatásával a mikrovilág biokonvereziós aktivitásának hasznosítása érdekében szisztematikus kutatómunkával feltérképezték a törzsgyüjteményekben fellelhető szervezetek biokémiai aktivitását. A rázott lombikban növekedő tenyészethez adott gazdaságosan előállítható kiindulási vegyület átalakulását papirkromatográfiás módszerrel követték. Az Upjohn támogatásával dolgozó Peterson és Murray 1952-ben megjelent szabadalma szerint a Rhizopus nigricans tenyészetéhez adott progesteronból 80%- os elméleti kihozatallal lehet 11αhidroxiprogesteront nyerni, amiből a gyógyszergyár kémiai eljárással üzemi méretben előállítható, piacképes gyulladásgátló kortikoidhoz jutott. A törzsgyüjteményben deponált 0.5% peptont és 5% malátakivonatot tartalmazó agar táptalajon fenntartott Rhizopus nigricans /ATTC 6227/ tenyészettel 3% kukoricalekvárt és 5% glükózt tartalmazó folyékony táptalajt oltva folyamatosan levegőztetve és keverve 24 C-on két nap alatt kifejődött tenyészethez acetonban oldva kerül az átalakítandó progeszteron, amely az oldószer kilevegőztetése után a gombafonalak felületére csapódik. A levegőztetést tovább folytatva a hidroxilezett termék a vizes fázisban dusul fel, ami a leszűrt vizes fázisból diklóretánnal extrahálható. A kiszűrt fonalas gombtenyészet napokig megtartva aktivitását hidroxilezésre ismételten felhasználható. Tökéletesítve az eljárást, a hidroxiprogeszteron elkülönítésére adszopciós gyantát alkalmazva, a hidroxilezett termék a szűrletből folyamatosan eltávolítható és így a reakcióterméktől megszabadított fermentlé levegő befúvásával visszajuttatható az erre a célra kialakított reaktorba. A folyamatos elkülönítés céljából, a sűlyesztett tenyészetben kinőtt mycélium olyan szűrőlappal ellátott reaktorban kerül felhasználásra, amelyben a hidroxilező aktivitással 29
30 rendelkező micélium tömegen oxigénnel telített fermentlé áramlik keresztül. Az átszűrt fermentlé visszajuttatását a reakciótérbe, a felszálló ágban elhelyezett fúvóka segíti. A befúvott levegő az oxigéntelítés mellett - a felszálló ágban levő frakció fajsúlyát, a légbuborékok segítségével csökkentve lehetővé teszi a fermentlé folyamatos visszatáplálását. A reaktorban elhelyezett micélium tőmeg hidroxilező aktivitása hat-nyolc nap alatt csökken a felére. Ez a módszer a rögzített enzimhordozó első gyakotlatban negvalósult alkalmazásának is tekinthető. Sikerrel alkalmazható a progesteron 11α-hidroxilezésére az Aspergillus ochraceus spóratömege amely ismételten felhasználható hidroxilazésre. A peptont és malátakivonatot tartalmazó szilárd táptalajon előállított spóratömeg 0.1%-os nátriumlauril szulfát oldatba szuszpendálva aktivitásveszteség nélkül 20 C-on hónapokig tárolható. A mikroszkopikus gombák átvizsgálásakor sok olyan törzset találtak, amely 11βhelyzetben hidroxilezte a szubsztrátumot. Colingswort és munkatársai Streptomyces fradiae törzset használva a Reichstein-féle S vegyületből hidrokortizont állítottak elő. Ez a felismerés nem csak azért volt jelentős, mert a termék az egyik legismertebb értékes glükokortikoid volt, hanem mert belőle Corynebacterium simplex dehidrogénező aktivitását használva igen értékes termék, a prednizolon nyerhető. Ez a vegyület 1955-ben jelent meg a gyógyszerpiacon mint hatásos gyulladáscsökkentő, a reumás izületi gyulladás ma is hatásos gyógyszere. Az egyik kiválasztott törzs, az Absidia orchidis, gyors növekedésű, homothallikusan zigosporát és szőrszerű légmicéliumot képző Mucor féle. A 2-4µ átmérőjű spórái kifejezett apofízissel rendelkező körte alakú sporangiumokban képződnek. Az üllepedő mikrokolonia formáját megtartó tenyészet szuszpenziója a Reichstein féle S vegyületből főtermékként hidrokortizont és az S vegyület 11α-hidroxilezett származékát állítja elő. A törzs C- 11-es hidroxilező képességét befolyásolni, a két termék arányát a körülmények változtatásával, illetve mutagén ágensek alkalmazásával nem lehet, ami azt igazolja, hogy csupán egyetlen hidroxilező enzim működik, amelynek aktiv centrumához különböző, a hidroxil csoport orientációját meghatározó helyzetben kötődhet a szubsztrátum. A kialakított technológia szerint malátakivonatot és peptont tartalmazó tápközegben, szigorúan meghatározott számú spórával oltva, készül a 24 C-on egy napig növekedő inokulum, amely a kukoricalekvárt, glükózt, és ammoniumszulfátot tartalmazó főfermentáció oltóanyaga. 40 óra növekedés után a mikrokoloniákból álló tenyészet, csapvizes mosás után, több napon keresztül változatlan aktivitással használható hidroxilező reagensként szolgál. A megfelelően higított, kimosott Absidia orchidis tenyészethez kalciumkloridos metanolban oldva adagolják a szubsztrátumot (Reichstein féle S vegyületet). A reakcióelegyet 38 órán keresztül megfelelő levegőztetés mellett 24 C-on keverik. A hidroxilezett szteroidok elegye a fermentlé szűrletéből etilacetáttal extrahálható, amelyből töményítés után kristályosodó nyers termék 55% hidrokortizont, 5% kortizont és 15% 11α-hidroxi származékot tartalmaz. A 30
31 kiszűrt micélium metanolos-csapvízzel mosva, változatlan aktivitással újra használható 6-8 napon keresztül. Gyakorlati szempontból előnyösebb a nyers szűrletet a hidrokortizon kinyerése nélkül tovább fermentálni a megfelelően higított Corynebacterium simplex tenyészettel, és csak az 1,2- dehidrogénezés befejeztével, a köztes termék kinyerésével együttjáró anyagveszteséget elkerülve nyerni ki a prednizolont. Ezzel a kombimált eljárással a kiindulási Reichstein féle S vegyületre számolva 48%-os hatásfokkal állítható elő a prednizolon. REICHSTEIN FÉLE S VEGYÜLET ELŐÁLLÍTÁSA A kombinált fermentációs eljárás szubsztrátumaként szereplő 17α,21-dihidroxi-4-pregnen-3,20- dion (Reichstein féle S vegyület) előállítására három kémiai lépés helyett biokonverziós eljárást használnak. A szteroid szapogeninek aglűkonjaiból, a dioszgeninből illetve a szolaszodinból induló kémiai átalakítási eljárás egyik intermedierje a 3β,17α,21-trihidroxi-5-pregnen-20-on.21- acetát, amelyből kémiai eljárással egy bromozást követő krómsavas oxidációval, majd cink jelenlétében végzett dehalogénezést követő alkalikus hidrolízissel nyerherő a 17α,21-dihidroxi- 4-pregnen-3,20-dion, az úgynevezett Reichstein féle S vegyület. Az antibiotikum termelésre, talajmintából izolált sugárgombák közül sikerült kiválasztani a fent ismertetett biokonverziós lépések bonyolítására használható mikroorganizmust. A később Streptomyces griseocarneus néven deponált törzs szójalisztet és glükózt tartalmazó táptalajon tenyésztve jelentős szteroid dehidrogenáz, és 21-aciláz aktivitással tünt ki. A törzs előnyös tulajdonsága, hogy a kívánt reakció vízzel nem elegyedő szerves oldószerek (toluol, diklóretán, butilacetát) jelenlétében is végbemegy. A szerves oldószer jelenléte nemcsak azért előnyös, mert a vízben alig oldódó szubsztrátumot nagy koncentrációban lehet a prokarióta tenyésztbe juttatni, hanem azért is hasznos, mert a mikroszervezet szteroidlebontó aktivitását, nem kívánt melléktermékek képződését a szerves oldószer megakadályozza. A Stretomyces griseocarneus-ban működö 3β-hidroxi csoportot dehidrogénező enzimet a légköri oxigén regenerálja. Szigorúan specifikus körülmények között működik, csak az A/B transz anellált szerkezetű 3β-hidroxycsoportot képes dehidrogénezni. Az oxidációt követő izomerizációt katalizáló enzimet nem sikerült elkülöníteni. Lehetséges, hogy a dehidrogénezést végző enzim szerkezete elősegíti az oxidációs lépés után a kötésvándorlást. Az iparilag is alkalmazható eljárásban a szójalisztet és glükózt tartalmazó táptalajon, 28 C-on, 48 óra alatt kinőtt tenyészethez literenként 65 ml diklóretánban oldva 20 g szubsztrátum kerül adagolásra. A reakcióelegyet ezután 37 C-on keverve, levegőztetve, és a ph-t lúgadagolással 8,2-8.4 értéken tartva, az oxidáció és a hidrolízis 3-5 nap alatt végbemegy. A terméket a szerves fázisból 96%-os hatásfokkal kristályos formában lehet kinyerni. 31
32 CURVULARIA FAJOK SZEREPE A KORRTIKOID SZINTÉZISBEN 1968-ban nyilvánosságra került irodalmi adatok alapján a prednizolon előállítására új lehetőség kínálkozott, mely azon a holland felismerésen alapult, hogy megfelelő Reichstein-S származék a hidroxilezésnél fellépő mellékreakciók lehetőségét csökkenti. A Curvulari lunata a hidrokortizon 11β-hidroxil csoportján kívül a 7α- és a 14α-helyzetű hidroxil csoportot is képes kialakítani. Ha azonban szubsztrátumként 17α-acetátot adagolunk a tenyészethez, akkor csupán egyetlen termék, a hidrokortizon.17α-acetát képződik, mivel az α oldalon elhelyezkedő nagy térfogatú szubsztituens akadályozza az enzim közeledését ebből az irányból. 17α-acetoxi származék előállítása Az új eljáráshoz Reichstein S 17α-acetátra volt szűkség, amiről kiderült, hogy kémiai eljárással gazdaságosan nem állítható elő. Járható útnak igérkezett az eddigi szubsztrátum triacetátjából induló fermentációs útvonal kidolgozása. Régóta ismert volt, hogy a primer, szekunder és tercier hidroxil csoportok észterei enzimatikusan eltérő sebességgel hidrolizálnak. Sajnos ebből a szempontból a Streptomyces griseocarneus teljesen alkalmatlan volt, mivel a hidroxiszteroid oxidázok szubsztrátumként nem fogadnak el 17α szubsztituált szteroidot. A 3β-hidroxiszteroid oxidázokkal ellentétben nem dehidrogénezik a dioxipregnenolon-17α-acetát származékot. Nagyszámú átvizsgált baktérium közül a már Arnaudi által 1949-ben izolált Flavobacterium lucecoloratum és a F. dehidrogenans tünt alkalmasnak, mivel az indukálható szteroidoxidáz mellett jelenlevő észteráza a tercier észtert 20-szor lassabban hidrolizálja mint a primer és szekunder alkohol észterét. Gyakorlatilag a szubsztrátumként bevitt triacetátból 6 ph-nál, szinte veszteség nélkül nyerhető a Reichstein féle S vegyület 17 acetát származéka. Az élesztőkivonatot tartalmazó táptalajon kinőtt 18 órás Flavobacterium tenyészethez 20mg/ml-es koncentrációban adagolható a 3β,17α,21-tracetoxi-5-pregnen-20-on, amely néhány nap alatt teljesen átalakulva kristályos formában 95%-os elmélei hatásfokkal kinyerhető, mint Reichstein-S 17.acetát. A 6.2 ph tartását a semleges ph-nál bekövetkező acilvándorlás (17-ről 21-re) akadályozása indokolja. Ez az új eljárás lehetőséget adott egy igen aktív hidroxilező törzs a Curvularia fallax alkalmazására, amely a Reichstein S-ből annak 14α-hidroxi és 14α,7α-dihidroxi származékát állítja elő. A 17-acetoxi csoport jelenléte (a hidroxilezés irányát az α-oldalról a β-oldalra terelve) egyetlen termék a 11βhidroxilezett származék képződését teszi lehetővé. A 3β,17α,21-trihidroxi-5-pregnen- 20-on.triacetátból hidrokortizon.17-acetát képződik főtermékként. Csekély mennyiségben melléktermékként a lánclebontást követő hidroxilezés eredményeként 14-hidroxi-4-androsztén- 3,17-dion képződhet. 32
33 A Curvularia lunata esetében a 21-acetoxi csoport akadályozza a hidroxilezési folyamatot, a gyenge észterhidrolizáló képessége miatt a diacetát illetve a triacetát számára alkalmatlan szubsztrátum. A Curvularia fallax kiemelkedő észterhidrolizáló képessége viszont néhány óra alatt eltávolítja az acilező csoportot a 21-es szénatomról és eredményesen hidroxilezi a 17 acetát származékot. Az átfordítás ténye Brannon elképzelését látszik igazolni. Szerinte a szteroid szubsztrátum a 3-as és 20-as oxigénfunkció által négy féle orientációban rögzülhet az enzim aktív centrumában. Ha a szubsztrátumon végrehajtott változtatás azt a helyet, ahova a hidroxilezés eredetileg irányul térbelileg gátolja, akkor az orientáció változása miatt a hidroxilezés a lehetséges engedély irányába módosul. A módosított szubsztrátum használat csak az esetben lehetséges, ha a nemkívánatos melléktermék az α oldalon axiálisan van hidroxilezve. Így csak a Curvularia esetében alkalmaztható, ahol a melléktermék 14α- illetve 7α-hidroxi származék. Ebből következően az Absidia orchidis esetében a módszer nem használható. Az eljárás ipari szempontból is igéretesnek látszik. Az átalakításhoz mindkét Curvularia törzs felhasználható, de a Curvularia fallax esetében a ph és a hőmérséklet beállítása nagyobb technológiai fegyelmet követel. Optimális körülmények között az inokulum táptalaj mogyorólisztet és glükózt, a főfermentáció, glükózt, szójalisztet, kukoricalekvárt és káliumfoszfátot tartalmaz. Hidroxilezés közben fontos a 6.2 ph érték tartása, mert lugosabb tartományban az észterhidrolízis megelőzheti a hidroxilezési folyamatot, ami elősegítheti a 7α,14α-hidroxilezett termékek megjelenését. A hidroxilező Curvularia sejttömeg biokémiai aktivitását megtartja. A kiszűrt micélium puffer oldatban szuszpendálva újra felhasználható. Aktivitása a harmadik alkalmazás után kezd csökkenni. A folyamat befejeztével, 8.5 ph-nál, 37 C o -on a hidrokortizon.17-acetátról az acilvándorlást követően az ecetsavat a gomba könnyen eltávolítja. Az Arthrobacter simplex a hidrokortizont és a hidrokortizon.17- acetátot azonos sebességgel dehidrogénezi. A 17-acetát csoport 33
34 gátolja a 9α-oxigenáz működését, azaz a szteroid váz lebomlását katalizáló terminális reakciót, a 9α-hidroxilezés folyamatát. Ezért az indukált tenyészet nagyobb koncentrációban, bomlási folyamat nélkül képes a hidrokortizon.17-acrtát dehidrogénezésével prednizolont előállítani. A fentiekből következően a Curvularia törzsek aktivitása hármas kombinált fermentációs rendszerbe építve, a 3β,17α,21-trihidroxi-5-pregnen-20-on.3,17,21-triacetátból közbenső tisztítási lépések nélkül, kristályos formában prednizolont szolgáltat a kiindulási anyagra számolva 45%-os kihozatallal. Az első lépésben a Flavobacterium lucecoloratum sejtszuszpenzió 28 C o -on 4 nap alatt 20mg/ml koncentrációban adagolt 3β,17α,21-trihidroxi-5-pregnen-20-on.3,17,21-triacetátból lehasítja a primer és szekunder acetát csoportot, és a szterinoxidáz hatására történik a 5-3-ol >> 4-3-on átrendeződés. A mikrokristályos formában kiülepedő végtermék, a 17α,21-dihidroxi-4-pregnen- 3,20-dion, az úgynevezett Reichstein féle S vegyület, ideális szubsztrátum a Curvularia törzsek által végzendő 11β-hidroxilezéshez, amely reakció 2 mg/ml koncentrációban folyik a már ismertetett körülmények között (6.2 ph, 28 C o ). A hidrokortizon.17-acetát 1,2-dehidrogénezését a szűrt fermentlébe juttatott a 18-ik órában andoszta-1,4-dien-3,17-dion adagolással indukált Arthrobacter simplex 24 órás tenyészetéből elkülönített, 20-szorosan higított baktérium tömeg végzi, amelytől az oxidációs folyamat végén centrifugálással különíthető el a prednizolon.17-acetát. Ennek a hidrolízise kémiailag végezhető. 34
35 A SZTEROID VÁZ OXIDATÍV LEBOMLÁSA A mikroorganizmusok csoportjai képesek a szteroid vegyületeket széndioxiddá és vízzé bontani. Ez a lebontó aktivitás különösen azokra a fajokra jellemző, amelyek a 3-keto-4-dehidroszteroidok 1,2-dehidrogénezésében kiemelkedő aktivitással vesznek részt. Ide sorolhatók a Corynebacterium, Mycobacterium, Nocardia, Pseudomonas, Arthrobacter genus fajai. Ebből következik, hogy a gyulladásgátló 1,2-dehidro-kortikoidok mikrobiológiai előállítása az anyagveszteség elkerülése érdekében nagy óvatosságot igényel. A lebontási utak biokémiai mechanizmusát fáradtságos munkával több kutatócsoport derítette fel. A megismert bomlási köztestermékekből az következtethető, hogy a lebontási útvonal a mikrovilágban általában azonos reakcióúton halad. A mellékelt ábra az oxidatív lebontási útba illeszthető reakciótermékekből összeállított vázlatot mutatja. A vizsgált mikroszervezet anyagcsere rendszerében uralkodó oxido-redukciós viszonyok határozzák meg a köztestermékek minőségét. A köztes anyagok mintegy biokémiai puffert képezve redukált formában is jelen lehetnek, ha valamilyen anyagcserezavar miatt a lebontásban szerepet játszó kofaktor regenerálása, visszaoxidálása átmenetileg késedelmet szenved. Dodson és Muir elsőként mutatták ki, hogy a szteroid váz bontása a 9α-hidroxilezést és az 1,2- dehidrogénezést követő retroaldol reakció eredményeként szekofenol köztes anyag képződéséhez vezet. A lépések sorredje a mikróba anyagcsere viszonyainak a függvénye. Az egyik vizsgált Nocardia törzsben a 9α-hidroxi csoport beépítése megelőzte a dehidrogénezést. A vizsgálatban szereplő Pseudomonas-nál viszont fordítva az 1,2-dehidrogénezés előzte meg a hidroxilezést. A két kulcsfontosságú enzim aktuális aktivitását a pillanatnyi anyagcsereviszonyok érzékenyen befolyásolhatják. Sok esetben a fiatal tenyészetben a hidroxilező aktivitás intenzívebb, az idősebb tenyészetben a stacioner fázis elején pedig éppen fordított a helyzet. A vizsgálatok igazolták, hogy a retroaldol reakció nem enzimes folyamat, másrészt, hogy az oxidáció légköri oxigén felhasználásával történik. Dodson és Muir megállapította, hogy a 19-hidroxi-4-androsztén-3,17-dion a Pseudomonas tenyészetben veszteség nélkül ösztronná alakul. Sih és Rahim Nocardia restrictus-t használva felismerte, hogy ha a 19-es anguláris metil oxidálva van, akkor a 9,10-szekofenol helyett minden esetben a retroaldol átrendeződés helyett az A gyürű aromatizációja következik be. A lebontási út következő lépése az A gyürű felnyitása, amit az aromás gyürű C 4 helyzetű hidroxilezése vezet be. A hidroxi származék képződése a gyürűfelnyitási folyamat előfeltétele. Ugyanez a hidroxilezés vezeti be Sih vizsgálatai szerint az ösztron oxidatív lebontásakor az A gyürű felnyitását. Az ábrán feltüntetett köztes terméket, a ketokarbonsav származékot csak ammoniával reagálva sikerült izolálni, mert a lebontási folyamat aktivitása a ketosavat maradéktalanul tovább alakítja. Sih Károly végül is propion-aldehid és piroszőlősav feldúsulását tapasztalta. 35
36 A szaggatott vonallal körülhatárolt átmeneti termék keletkezése pillanatában irreverzibilis retroaldol átrendeződés termékeként a szteroidváz szekofenol formájában jelenik meg. Ezért a szakma termináló oxigenáznak nevezi a szteroid-9α-hidroxilázt. A szteroid vázlebontás részleteinek feltárása, az elméleti alapokig hatoló vizsgálata gyakorlati értelmet nyer, mivel a hormonszintézishez szükséges alapanyagok előállítására szolgáló módszerek kialakitásához, fejlesztéséhez útmutatást ad. 36
37 A másik részletesen vizsgált terület a szterin oldallánc mikrobiológiai úton végrehajtott szelektív lebontása. Tatum már 1913-ban leírta, hogy a növényi, illetve állati eredetű szterolokat a mikroorganizmusok egy csoportja szénforrásként képes hasznosítani. Turfit 1948-ban közzétett dolgozatában arról számol be, hogy a Proactinomyces erithropolis a tápközegbe adott kolesztenont és az epesavat kismértékben etiosavvá alakítja. A szakmai közvélemény ennek a közlésnek nem tulajdonított jelentőséget. Annál nagyobb érdeklődést váltott ki 1964-ben Whitmarsh munkája, amely szerint koleszterinből androszt-4-én-3,17-diont, androszta-1,4-dién- 3,17-diont, 3-keto-bisnorkolénsavat és 3-keto-bisnor-1,4-koladién savat kapott. Ez a közlés nem jöhetett volna jobbkor a nyersanyaghiánnyal küzdő szteroidipar számára. A közlemény elérhető olcsó szteroidforrásra, az állati és növényi eredetű szterinekre irányította a kutatócsoportok figyelmét. Nem volt kétséges, hogy a gyógyszerpiac igényelte hormonhatású szteroidok, valamint gyógyászati jelentőségű származékaik ipari előállítása csak a teremészetben jelentős mennyiségben előforduló szterinekből történhet. A Mycobacterium katabolikus aktivitása a szterin molekula jól elkülöníthető két régióján, egyrészt a szteroid váz AB gyűrűjén érvényesül, másrészt az előbbitől teljesen függetlenül bontja az oldalláncot. Sih és Wang az általuk kidolgozott eljárással 3β,19-dihidroxi-5-kolesztén.3-acetátból 72%-os elméleti kinyeréssel jutottak ösztronhoz. Későbbi vizsgálataikban Sih és munkatársai koleszterint kívántak lebontani Nocardia, Mycobacterium, Corynebacterium, és Arthrobacter törzsekkel. Izotóppal jelzett szubsztrátumot használva megállapoították, hogy a lebontás folyamata az egyes törzseknél azonos, minden esetben kolánsav bisznorkolánsav közti terméken keresztül történik. A biokonverzió reakcióelegyéből izolált termékekből össze lehetett rakni az oldallánc-eltávolítás minden egyes lépését képviselő reakciósort, ami igazolta, hogy a lebomlás a zsírsav béta-lebomlás sémáját követi. Nyilvánvalóvá vált a retroaldol átrendeződés fontos szerepe. Kisérleteikben nehézoxigént tartalmazó vizet alkalmazva kiderült, hogy a végtermékben szereplő 17-es szénatomhoz kötődő oxigén nehéz vízből származik. Később megállapították, hogy az oldallánc lebontás baktériumokkal (Corynebacterium simplex) és gombákkal (Fusarium solani) egyaránt elvégezhető ben Dr. Wix Györgynek és munkatársainak sikerült a szterin-oldallánc szelektív lebontását elérni a Mycobacterium phlei tenyészethez adagolt 8-hidroxi-kinolinnal. Így szelektíven gátolva a 9αhidroxiláz enzim működését 1,4-androsztadién-3,17-dionhoz jutottak. Megkísérelték az A-gyűrű metabolizációját már az első lépésnél megakadályozni. A szterinek 3ß-hidroxil-csoportját kis térkitöltésű, rövid láncú alkil-éter, illetve karbamoil-védőcsoporttal látva el Ambrus Gábornak sikerült az A-gyűrű metabolizációját már az első lépésnél megakadályozni. E származékok oldalláncát a Mycobacterium phlei lebontotta. Ezt követően a védőcsoportot eltávolítva, az androgén hormonok, anabolikumok és aldoszteron-antagonisták szintézisének a kulcsintermedierjéhez a dehidroepiandroszteronhoz jutottak.a 6,19- ciklokoleszterinek, a szterin éterek, vagy a karbamoil szterinek oldallánca ugyanúgy lebontható, mint a szabad 3-oxigén funkcióval rendelkező szterinek oldallánca. Végül igazolták, hogy a diosgenin oldallánca, a spiroketál gyürűje is lebontható. A reakciótermék D gyürüjében az oxigén eredeti helyén maradva a 16-os szénatomhoz kötődik. A ketocsoport utólag bekövetkező redukciójára utal, hogy a reakcióelegy 16α- és 16β-hidroxi származékot is tartalmaz. A kisérleti eredmény valószinüsíti Kondo és Mitsugi elképzelését, amely egy hidrolitikus folyamatot követő dehidrogénezést, majd egy Baeyer-Villiger oxidációt követő újabb hidrolízist tételez fel, végül utolsó lépésként a retroaldol reakciót valószinüsíti. Eddig nem sikerült kimutatni a 20- hidroxi-pregnán származékot a reakciótermékek között. A Mycobacterium-nál igazolt β-lebontás is hasonló eredményre vezet. A két lebontási út egyidejű működése is lehetséges, ami a szterol-oldallánc szelektív lebontásakor Mycobacterium aureum esetében megmagyarázná a progeszteron megjelenését a reakcióelegyben. ** 37
38 A szteroid váz lebontása szempontjából döntő jelentőségű a vastartalmú 9α-hidroxiláz és a FAD kofaktort igénylő szteroid-1,2-dehidrogenáz. A két enzim aktivitásának hatására a szteroid váz szekofenol képződésen keresztül elbomlik. Az AD (androszténdion) gazdaságos előállításának feltétele a fent említett két enzim működésképtelensége. Xxxxxxxxxxx Az elmúlt évtizedekben különböző prokarióta szervezetekből számos enzimsérült mutánst lehetett izolálni, amelyeket a lebomlási út egyes közti termékeinek nagyobb mennyiségben való előállítására lehetett használni. A szteroid ipar nyersanyag ellátását a XXI században is ezeknek a mutánsoknak az ipari gyakorlatban való használata szolgálja. 38
39 A 9α-oxigenáz hiányos mutáns törzs csak az oldallánc lebontására képes. A szitoszterinből induló 4-androsztén-3,17-dion főtermék (AD) képződésére vezető biokonverziós eljárás során, az oldallánc lebontásában mindössze tíz enzim vesz részt 16 enzimkatalizálta lépésben. Néhány enzim többször is szerephez jut Az enzimek részben membránhoz kötve, illetve a membrán közelében levő mikroszómákban működnek. A membránon keresztül hatoló szterin molekula apoláros oldallámcára az oxigén funkciót az indukálható omega-oxigenáz katalizálja. A rekcióelegyben oldott oxigén beépítése a NADPH két elektronjának a felhasználásával történik egy molekula víz képződése mellett. Az alkohol aldehiddé oxidálását a citoplazmában működő alkohol dehidrogenáz végzi. Az eltávolított két elektront a NAD + kofaktor veszi fel. A következő két elektron eltávolítását ugyancsak NAD kofaktorral működő aldehid dehidrogenáz végzi. A következő működő enzim a tioforáz, amely acetil-coa felhasználásával az intermediert acil-s-coa származékká alakítja. Ez ideális szubsztrátum a mikroszómába rögzített acil-s-coa dehidrogenáz számára. Két hidrogént az enzimhez kötött FAD vesz fel. Ekkor következik be a szitoszterin C-24 etiloldalláncának karboxilezése. Ez az intermedier ideális szubsztrátum a metil-krotonil-coa karboxiláz számára, mert az etil oldallánc 1-szénatomja kitüntetetten reakcióképes A karboxilezést végző, indukálható enzim képződése a biotinil enzimet karboxilezett állapotban ígényli. A karboxilezés a kialakuló adenilsav szénsav vegyesanhidrid aktuális koncentrációjától (áttételesen a széndioxid parciális nyomásától) függ. A karboxilezett telítetlen intermedier vízfelvételét az enoil-hidratáz katalizálja. Ezt követi úgynevezett retroaldol folyamat eredményeként az első propionil-coa spontán leválása. A spontán folyamatot a CoA-tiolészter C 2 atom kitüntetett elektronvonzó képessége mozgatja. A következő lépésben újra a tioforáz lép működésbe, amely.acetil-coa igénybevételével katalizálja a tovább bontható származék képződését. Ezt követően a zsírsav ß-lebontásában is résztvevő ß-keto-tioláz CoA jelenlétében leválasztja a második propionil-coa-t. Ezt követően az acil-s-coa dehidrogenáz által előállított telítetlen intermedier vízfelvételét az enoil-hidratáz katalizálja. A hidroxilezett származékból a ß-hidroxiacil-S-CoA dehidrogenáz működését követően a keto-tioliáz hatására szabadul fel az acetil-coa. Az oldallánc lebontás utolsó szakasza a bisnorkolenil-coa dehidrogénezése. A két hidrogént ismét a FAD kofaktorral működő acil-s-coa dehidrogenáz távolítja el. Ezt követi a acil-coa-enoil-hidratáz által katalizált vízfelvétel, amelynek hatására a spontán bekövetkező retroaldol átrendeződés eredményeként megjelenik a hamadik propionil-s-coa és főtermékként AD képződik. Az utolsó enzimreakciók lefutását erősen befolyásolja a szteroid mag D gyűrüjének merev szerkezete, csökkenti a reakció teljesítményét (fluxusát). Az utolsó lépések szűk keresztmetszete miatt feltorlódó köztestermékek az aciltranszferáz hatására a kofaktor ellátottsággal összefüggő minőségben és mennyiségben megjelennek a biokonverziós elegyben. Ezek a köztesanyagok (például bisznorkolénsav, tesztoszteron), már nem kerülnek vissza a lebontási folyamatba, hanem a főterméket szennyező melléktermékként rontják az eljárás hatásfokát, a kinyerést. A lebontási folyamat lépései ugyan nem igénylik a baktérium sejt szaporodását, a kofaktorok (NADH illetve FADH 2 ) regenerálása szempontjából azonban a légző-rendszer működése, az életben tartás megkerülhetetlen. Ezért nem jöhet számításba szerves oldószer alkalmazása. Az oldallánc lebontás szük keresztmetszete a metilmalonil-coa szintetáz, más néven a propionil-coa karboxiláz nem kielégítő működése, mivel a nagyobb mennyiségben képződő propionil CoA eliminálására, hasznosítására a mikróba metabolizmusa csupán ezzel az egyetlen lehetőséggel rendelkezik. A propionil-coa metilmalonil-coa szintetáz hatására egy szén felvételével metilmalonil-coa-vá, ez pedig egy mutáz hatására szukcinil-coa-vá alakulva a Krebs ciklusban hasznosul. Mindkét enzim propionsavval, ill. propanollal indukálható. 39
40 Az oldallánc szelektív lebontásával egy időben a szteroid mag AB gyűrűjén is enzimek által katalizált átalakítások mehetnek végbe. A Mycobacterium törzsekben NAD-függő 3ßhidroxiszteroid dehidrogenáz alakítja ki a 3-keto-4-en szerkezetet. {β-enzim} A Nocardia törzsekben és több Actinomyces fajban előforduló szterinoxidáz működését az átvizsgált Mycobacterium törzsekben nem sikerült kimutatni. 40
41 19-NORSZTEROID TOTÁLSZINTÉZIS A születés szabályozás hormonális úton való megoldása gazdaságos norszteroid előállítási eljárás kidolgozását igényelte. Az egyik út az előbb bemutatott szelektív oldallánc lebontással nyert szterin 19 metil-csoportjának az eltávolítása. A Johnson nevéhez fűződő ösztronszintézis, a norszteroidok totálszintézissel történő előállítása a molekulában előforduló királis szénatomok miatt gazdaságosan nem oldható meg, bár a létrejövő racém termék szétválasztására az enzimek sztereospecifitásán alapuló módszerek jól alkalmazhatók. Hátránya, hogy ez az eljárás optimális esetben sem éri el a racém végtermékre számítva az 50 %-ot. Ebből következően nagy gazdasági jelentősége van minden olyan eljárásnak, amely egy adott szintézisút optikailag inaktív termékéből biológiai módszerrel alakítja ki az aktív intermediert. Ebben az esetben ugyanis nincs elvi akadálya a kvantitatív átalakítás tetszőleges megközelítésének. Döntő fordulatot jelentett Velluze, valamint a Smith-Torgov féle, eredetileg racém termék előállítását szolgáló totálszintézis kidolgozása. A 2-metil-2-aralkil-ciklopentán-dion viszonylag olcsó teremék. A C gyűrű sztereospecifikus zárása biztosíthatja a végtermék optikai tisztaságát. Ezt a feladatot legegyszerűbben az egyik szterikusan kitüntetett helyzetű ketocsoport reakcióképességének átmeneti felfüggesztésével lehet megoldani. 41
42 Kémiai redukcióval a szekodion redukálható keto csoportjai két helyzetben radukálódnak. A reakcióelegyből azonban a kivánt termék rezoválással való elkülönítése nem tekinthető gazdaságos megoldásnak. Izolálható viszont olyam élesztő, amelynek a NADH igényes 3ß,17ß-hidroxiszteroid dehidrogenáza képes a kívánt királis intermedier előállítására, miközben a kiválasztott törzsben jelenlevő 3α-hidroxiszteroid-dehidrogenáz csekély aktivitást teljesít. Inhibitor jelenlétében ez utóbbi aktivitás elnyomható. A két dehidrogenáz aktivitása az egyes törzsekben eltérő szintet képvisel, ami mutagén hatásra is változhat. A redukció végbemenetelének feltétele a redukált kofaktor (NADH) állandó jelenléte, ami azt jelenti, hogy nélkülözhetetlen a dehidrogénezhető vegyület (szénhidrát, esetleg alkohol) és az oxigén jelenléte. A prokirális szubsztrátum, a 3-metoxi-8,14-szeko-1,3,5(10),9(11)- ösztratetraén-14,17-dion mikrokristályos formában adagolva gyorsítja a reakció lefolyását. 42
43 Egy viszonylag gazdaságosan előállítható prokirális 3-metoxi-8,14-szeko-1,3,5(10),9(11)- ösztratetraén-14,17-dion köztestermékből (szekodion), a 17-ketocsoport sztereoszelektiv redukciójával nyerhető a kulcsintermedier (szekoolon). A szekodion 17 ß-hidroxi származékából nemkívánt melléktermék képződése nélkül jutunk el az optikailag tiszta természetes konfigurációjú ösztradiol (19-norszteroid) származékhoz. A 14-ß-hidroxi származékból előállítható az enantiomer ösztron. 43
44 44 A Pichia carsomii esetében a 3α-hidroxiszteroid oxidoreduktáz aktivitás a nagyobb, ezért főtermékként a szekodionból 14α származék képződik.
45 Üzemi körülmények között végzett biokonverziós folyamat adatait bemutató ábrán jól látható a répacukor adagolás hatása 45
46 Biológiai hatás szempontjából a 13-etil-19-norszteroidok (18-metil-19-norszteroid) előállítása céljából az etilszekodion redukcióját is meg kellett oldani. Megfelelő detergens alkalmazásával a biokonverziós aktivitás fokozható volt Redukálandó szubsztrátum 13-metil szekodion (18-metil) 13-etil szekodion androszténdion 17-ketoösztron (18-etil) 13-propil szekodion Az etilszekodionból biokonverzióval nyert 13-etil-szeko-17β ol-14-on ból előállítható 19- norszteroid ideális kiindulási termék a hatásos gestodén előállításához 46
47 Hatásos antikoncipiens 47
48 ENANTIOMER ÖSZTRON INTERMEDIER ELŐÁLLÍTÁSA Alkalmasan megválasztott mikroorganizmusok felhasználásával valamint a biotranszformációs lépéseket kémiai redukcióval kombinálva, az enantiomer ösztron előállítására szolgáló kulcsintermedier, a 14ß-hidroxi,17-keto-szekoolon is előállítható. Az első lépésben Saccharomyces uvarum-ot levegőztetett tenyészetben használva elkülönítjük a 17ß-szeko-olont, amit borohidrides redukcióval 14ß,17ß- és 14α,17ß-diolok elegyévé redukálunk. A borohidrides redukció a 14β,17β- diol képződésnek kedvez. {A 17βszekoolon ecetsavanhidriddel acetilezett származékának borohidrides redukciója előnyösen növeli a 14β,17β-acetil származék képződését.} A következő anaerob lépésben a 14α hidroxi csoportot a Flavobacterium dehidrogenans keto csoporttá redukálja. A képződő 17ßhidrozi származék recirkulálva újra redukálható borohidriddel. A katalitikus redukcióval nyert elegyben levő 14ß,17ß-diol 17ß-hidroxi csoportját anaerob reakciókörülmények között egy erre a célra kiválasztott Nocardia törzs dehidrogénezi. Az így előállított (cisz-helyzetű) 14ß-hidroxi-17- keto-szekoolonból jó hozammal előállítható az enantiomer ösztradiol. (Lásd az ábrát a 42. oldalon) {A prokarióta adott esetben az acetil csoportot is hidrolizálja.} 48
49 6-AMINOPENICILLANSAV (6-APS) ELŐÁLLÍTÁSA A szakmai körokben igen olvasott Nature című folyóirat 187-es kötetében a oldalakon 1960-ban két közlemény jelent meg. Az egyik angol szerzők tollából (Beecham Research Laboratories Ltd.), a másikat pedig egy amerikai gyógyszergyár (Bristol Co) kutatói írták. A két közlemény gazdaságos biokonverziós eljárás lehetőségét vázolta fel a penám alapmolekula, a 6-aminopenicillinsav nagyipari előállítására. Ennek a vegyületnek a jelenlétét prekurzor nélkül növekedő Penicillium chrysogenum tenyészetében már egy évvel előbb közölték Batchelor és munkatársai (Nature :257). Sőt a kép teljességéhez tartozik, hogy a japán Kato (J. Antibio :130, 6:184), valamint ugyancsak japán folyóiratban (J. Agric. Chem. Soc :411) Sakaguchi és Murao benzilpenicillinből való képződését már tíz évvel előbb közölték penicillin amidáznak nevezett enzim termékeként. Erről azonban, talán nyelvi nehézség miatt tíz éven keresztül nem vett tudomást az antibiotikum-ipar. Később ez az enzim penicillin acilázként (EC ) vonult be a szakirodalomba. A nagy áttörést Chain és munkatársainak az a felismerése jelentette, hogy a penicillin alapváz 6-aminocsoportja tetszőleges reagenssel acilezve új nagyhatású azetidinon származékok előállítását teszi lehetővé. A penám alapmolekula kémiai acilezésének a gondolatát valószínüleg a penicillin totálszintézisével foglalkozó kémikusok ezidőtájt nyilvánosságra került eredményei hívták elő. A penicillin szerkezeti képletének a megismerésekor, a királis szénatomok száma visszarettentette a kemikusokat a hatásos vegyület szintézisétől. A szerves szintézis gyors fejlődése, különösen a peptidszintézis módszereinek a fejlesztése az ötvenes években már királis szénatomot tartalmazó vegyületek gazdaságos előállítását tették lehetővé. A ftálimid védőcsoport irányító hatása, a tercier butilészter csoport könnyű eltávolíthatósága, és a diciklohexil-carbodiimid alkalmazása ugrásszerű fejlődést hozott. Sikerrel vállalkoztak a hatásos anyag totálszintézisére. Az első reakciólépésben penicillamid-hidrokloriddal nátriumacetáttal pufferolt vizes etanolban reagál a tercier-butil-ftálimido-malonaldehid. A ftalil csoport eltávolítását hidrazin komplex képzésen keresztül végezték ecetsavban szuszpendálva. A tecier-butiloxikarboniltiazolidin származék aminocsoportját egyenértéksúlynyi fenoxi-acetilkloriddal acilezték két 49
50 ekvivalens trietilamin jelenlétében. A tercier-butiloxi csoport eltávolítását metilén kloridban vízmentes sósavval végezték, amit átkristályosítás követett ekvivalens piridint tartalmazó vizes acetonból. A gyürűzárást ekvivalens mennyiségű N-diciklohexilkarbodiimiddel végezték dioxán vizes oldatában. A szintézis az enantiomerek előállítására is lehetőséget adott. Ezek azonban alig mutattak biológiai aktivitást (0.7% a teremészetes penicillinre vonatkoztatva). A tritil védőcsoport megadta a lehetőséget a penicillin mag a 6- aminopenicillinsav totálszintézisére, ami számos új penicillin származék előállítására adott lehetőséget. Ez esetben a ftalil védőcsoport eltávolítása előtt a karboxilcsoportot észterezték. A tercier butiloxivédelem eltávolítása után tritilezték az aminocsoportot dietilamin jelenlétében. Ezt követte a gyürűzárás, amit N,N -diizopropil-karbodiimiddel hajtottak végre dioxánban a D-α-4-karbometoxi-5,5-dimetil-α-tritil-amino-2-tiazolidin-ecetsavból. Al-oxidon végzett kromatografálás után kristályos formában nyerték a 6-tritil-aminopenicillinsavmetilésztert, amiből az észter hidrolízisével dietilamin sót nyertek, majd híg sósavval a tritil csoport eltávolítása következett. Nyilvánvaló, hogy Sheehan és munkatársainak 1955-ben a V-penicillin totálszintéziséről megjelent munkái katalizálták az antibiotikumokkal foglalkozó mikrobiológusok és vegyészek fantáziáját. Hamarosan számos mikrobiológiai eljárás született a 6-APS származékainak előállítására. A prekurzor nélkül végzett direkt-fermentációs eljáráson kívül az olcsó penicillint használva kiindulási anyagként sikerrel használtak biokonverziós módszereket. A prekurzor nemcsak hatásosabb termék képződését eredményezi, de a prekurzor beépülésével egyidejüleg a α-amino-adipinsav újra használhatóvá válik a szintézisben, ami a penám váz képződését segíti. Prekurzor jelenlétében több mint egy nagyságrenddel nagyobb koncentrációban képződik a penám váz, mint prekurzor nélkül. Az pedig, hogy a 6-APS előállításhoz G és a V penicillin egyaránt használható kiindulási anyagként a termék előállítási ára szempontjából nem elhanyagolható. Bizonyos baktériumok például az Escherichia coli a G penicillin dezacetilezését végzi hatásosan. A Streptomyces törzsek a V- penicillinről, a fenoxiecetsav eltávolításában jeleskednek. Lugos ph a hidrolízisnek, savas ph a szintézisnek kedvez. Az enzim erdetileg valamilyen transzacilezési folyamatot katalizált a membránban. Az eljárás ipari jelentősége miatt az enzimtermeló törzsek fejlesztése fontos feladat volt. Kezdetben a klasszikus mutagén ágenseket alkalmazva is sikerült nagyobb enzimszintet termelő törzseket izolálni. Később konstitutívan penicillin acilázt termelő mutánst izoláltunk. Az enzimes eljárás kihozatalát a szubsztrátum és a termék érzékeny volta befolyásolja. Semleges ph-nál 37 C-on 3 óra alatt a hasznos teremék 50 %-a inaktíválódik. A képződő 6-APS képes a vízben oldódó széndioxiddal, a szénsavval reagálni. A 6-APS karboniloxi származékából spontán képződő imidazolidon enolizált formája könnyen dekarboxileződik. A másik inaktiválódási lehetőség a polimerizáció, ami az enzim transzacilkező aktivitásának a következménye, és a reakció előrehaladtával a termékkoncentráció függvényében jelentős veszteséget okoz. A két folyamat egymást segítve rontja a kihozatalt. 50
51 A reakcióelegyben G-penicillinből képződő 6-aminopenicillansav sorsa 6-APS előállítása kémiai eljárással A félszintézishez használható alapanyag(6-aminopenicillansav) nagyvolumenű előállítására, illetve a fenoxiecetsav tetszőleges csoporttal való kicserélésére kémiai megoldás is ismeretes. Első lépésként a penicillin karboxil csoportjának a védelme alkil-szililészter képzésével oldható meg. A kapott reakcióterméket -30 o C alatt iminohalogenidet képző reagenssel, például (PCl 5 ) foszforpentakloriddal hozzák össze. Az átmeneti termékként képződő N-monoszubsztituált imino-kloridot alkohollal (esetünkben n-butanollal) reagáltatva imino-éterré alakítják, amelyből az iminoéter csoport és a védőcsoport hidrolízisével nyerhető a 6-APS. Ezek az eljárások gazdaságos lehetőséget jelentettek félszintézissel előállítható új penicillin származékok nagyipari előállítására. Általában így az oxacillin előállításakor is savkloridokat használnak acilező szerként. 51
52 Esetenként az enzimes szintézis is szerepet kap, kihasználva a penicillin-(aciláz)-amidáz reakció reverzibilis voltát. Különösen előnyös az enzim, ha ezzel elkerülhető a védőcsoport használata. Igy gazdaságosan nyerhető az ampicillin fenilglicintioglikolsav észterből és 6-APS-ből penicillin-aciláz enzimmel savanyú közegben végzett reakcióval. Ampicillin előállítása Escherichia coli tenyészetével A hatvanas években előállított sok ezer új vegyület között több száz hatásos termék akadt. Ezek az új származékok a fermentációs úton előállítható két (G- és V-penicillin) származéknál előnyösebb terápiás hatásúak. A savamid kötés közelében elhelyezett nagy térkitöltésű szubsztituens (methicillin, penbritin) védelmet jelentett a penicillináz inaktíváló hatásával szemben. A savállóságot növelte a savamid kötés közelében elhelyezkedő elektronszívó csoport (oxacillin, cloxacillin, flucloxacillin). A baktérium spektrum szélesítése céljából előnyös volt az amino csoportot tartalmazó arómás savvakkal (fenilglicin, p-hidroxi-fenilglicin) történő acilezés (ampicillin, amoxicillin). Szabad karboxil csoport jelenléte (fenilmalonsav) a Pseudomonas fajok leküzdésére is alkalmassá teszi az új származékot (carbenicillin). A penámváz karboxil csoportját érintő kémiai átalakítások (pl.g-penicillin acetoximetilésztere) az alkalmazhatóságot befolyásolhatják (maripen). A félszintézissel előállított új származékok antibakteriális hatása (MIC érték) is kedvezően megváltozhatott. Eltérést találtak a felszívódásban, a kiürülésben, a vérszint alakulásában. Félszintetikus azetidinon származékok előállítása A 6-aminopenicillansav acilezésével a penicillináznak ellenálló félszintetikus penicillin származékok nyerhetők, de a baktérium-spektrum szélesítésére is lehetőség adódott Oxacillin. (5-metil-3-fenil-4-izoxazolil-penicillin. nátrium monohidrát) A 3 fenil és az 5 metil csoport térbeli gátlása akadályozza a molekula kapcsolódását a penicillináz aktív központjához. A molekula elektronszerkezete savtűrést biztosít. Ezért per os adagolva is alkalmas a penicillin rezisztens fertőzések leküzdésére. A kapszulában forgalmazott (250 mg) hatóanyag a bélből felszívódva erősen kötődik a plazmafehérjékhez. Egy óra múlva hatásos vérszintet ad. A vesén keresztül változatlan formában ürül. Adagolása 4-6 óránként 2 kapszula egy órával az étkezés előtt. Ampicillin (Penbritin, Semicillin) D-α-aminobenzilpenicillin.trihidrát. Az aminocsoport az antibiotikum baktérium spektrumát kiterjeszti. Eredményesen alkalmazható a Gram negatív kórokzók, így az enterococcus okozta fertőzések esetében is. Különösen hatásos akut hugyuti fertőzések (Escherichia coli, Proteus mirabilis, etc.) leküzdésére. Az ampicillin amino csoportja savanyú körülmények között protonálva van, ami bizonyos mértékű savstabilitást biztosít. Kapszulában adagolva általában két óra alatt érhető el a maximális vérszint. A vesén keresztül változatlan formában ürül. Egy kapszula 250 mg hatóanyagot tartalmaz, amiből hat óránként kell egyet bevenni. Injekciós adagoláshoz nátrium sóját használják (porampullában). 52
53 Amoxicillin 6-D-α-amino-p-hidroxifenilacetamido-penicillansav, az ampicillin p- hidroxi analógja. Baktérium spektruma és kémiai stabilitása hasonló, de jobban szívódik fel a bélcsatornából, ezért magasabb vérszintet ad. A gyorsabb felszívódás miatt kevésbbé hatásos disenteria esetében. A vesén keresztül változatlan formában ürül, ezért hat óránként kell egy egy kapszulát (250 mg hatóanyag) adni. Clavulánsavval együtt adagolva a rezisztens törzsek ellen is hatásos széles spektrumú gyógyszerkészítményként kerül forgalomba. Ezek az eredmények félszázaddal ezelőtt a penicillináznak ellenálló, széles spektrumú, kevéssé toxikus β-laktám antibiotikum félszintézisének lehetőségét teremtették meg. A bakteriális fertőzések leküzdésében ezek a vegyületek ma is döntő szerepet töltenek be. Az eredmények azonban nem függetleníthetők a gazdaszervezet védekező mechanizmusának állapotától. A felhasznált hatóanyagok csak csökkentik a korokozók életképességét. A végső eredményt az ellenanyagképző mechanizmus, az immunológiai védelem teljesítménye határozza meg. Bacillus cereus esetében, a kezdetben membránhoz kötődő enzim a periplazmikus térben megjelenő proteáz hatására extracelluláris penicillinázként működik a környezetben. Limitált proteolízis Proteáz hatás apoláros szakasz **extracelluláris penicillináz***** H 2 N Met (Xxx) 15 Ser (Xxx) 22 Glu Ser Lys Thr Glu (Xxx) 258 Lys COOH O foszfatídsavval acilezett szerin-oh HO P=O HO CH CH CH 2 H O O O=C C=O R R R = páratlan szénatom számú alkil láncok (A foszfolipid zsírsavkomponense ) A membránhoz kötődő penicillináz extracelluláris enzimmé válása a citoplazmammbrán külső felületén tevékenykedő proteáz aktivitásának a függvénye. 53
54 A PENICILLIN FELFEDEZÉSE és a gyógyászatban való alkalmazásának történetét tanulságos röviden áttekintani. A Londonban működő St. Mary kórház bakteriológusa, a Hollandiából menekült családból származó Alexander Fleming ( ) doktori disszertációját a magasabb rendűek által védekező anyagként termelt lizozim tulajdonságait vizsgálva készítette. Ez az enzim folytonosan termelődve, az élőlényt károsító baktériumok sejtfalának szénhidrát építőelemei közötti β-glikozidos kötést hidrolizálva elpusztítja a korokozót. A szepszist okozó baktériumok tulajdonságait vizsgáló Fleming 1929-ben a tenyészetét fertőző penész hatására bekövetkező baktérium-pusztulást, valamint a hatásos anyag hő és ph érzékenységét tapasztalva, valami lizozimhoz hasonló hatóanyagra gondolt. A munkába kapcsolódó Dr. C. Thom mikológus rendszertanilag Penicillium notatum-mal, egy ritkán előforduló fajjal azonosította az antibakteriális hatást mutató anyagot termelő fonalas gombát. Az antibakteriális tulajdonságú hatásos anyag kinyerése céljából végzett eredménytelen kísérleti munka után, 1932-ben Clutterbuck, a vegyész munkatárs, csak annyit közölhetett, hogy a a termelő törzs fajnevéből következően penicillinnek elnevezett hatásos anyag labilis, ph és hő érzékeny gyenge sav. Az anyag hatásossága és elhanyagolhatóan csekély toxicitása azonban nem hagyta feledésbe merülni a felfedezést, bár a nehézségeket látva Fleming sem bízott a sikerben. Az 1930-as évek végén csupán a tenyészet szűrletét javasolhatta külsődleges alkalmazásra. Az Oxfordban összekovácsolódott munkacsoport kitartó munkája, Chain és Florey erőfeszítései végül eredményre vezettek. Felületi tenyészetből elkülönített tenyészléből fagyasztva szárítva sikerült kielégítő stabilitású barna port nyerni, amellyel Abraham és munkatársai már bizonyíthatták az anyagnak a szeptikus folyamatok leküzdésében elért fantasztikusnak tűnő gyógyító hatását. (Abraham közben észlelte a hatóanyagot lebontó rezisztens baktériumok megjelenését) Tiszta anyaggal nem rendelkezve, biológiai egységet használtak összehasonlítási alapként. Oxford-egységnek (NE) tekintették azt a legkisebb penicillin mennyiséget, amely egy bizonyos Staphylococcus törzs növekedését 50 ml táptalajban képes gátolni. (Ez ma o.6 mg kristályos benzilpenicillin Nasónak felel meg.) Fleming 1940 november 14.-én a Gyógyszerészeti Társaság tudományos űlésén számolt be az eredményekről. A háborúba keveredett Angliából ezekkel az eredményekkel indult 1941 júliusában Florey és Heatley az Egyesült Államokba az elnök anyagi támogatását kérni az eredmények továbbfejlesztéséhez. A kormányszervek felismerték a téma katonai jelentőségét. (Az első világháborúban megsebesült katonák több mint 50%-a szepszisben halt meg!) A Pentagon a Northern Regional Research Laboratory (NRRL Peoria, Illinois) fermentációs osztályán R. D. Coghill és A. N. Richards mikrobiológusokat bízta meg a téma vitelével. David Perlman, a Wisconsin egyetem munkatársa szerint a termelő törzsek kiválasztásában az USA lakosságának a segítségét is igénybe vették. Közhasznú hazafias tevékenységgé vált a különböző helyekről származó gombatenyészetek eljuttatása a Mezőgazdasági Minisztérium által fenntartott NRRL törzsgyűjteményébe, ahol szorgalmas munkával minden tenyészet biológiai aktivitását megvizsgálták. Az összegyűjtött törzsek közül egy háziasszony által beküldött grépfruitról származó Penicillium (notatum) chrysogenum néven leírt tenyészet a Fleming által izoláltnál 10-szer nagyobb biológiai aktivitást mutatott. Később (feltehetőleg szabadalmi okokból) a notatum előnév lemaradt. Ebből az izolátumból fejlesztették ki a gyógyszeripar fermentációs üzemeiben világszerte felhasználásra kerülő törzseket. Az első felhasználásra került barna por mindössze 1% hatóanyagot tartalmazott. A tisztításban való előrehaladás a kémiai szerkezet felderítésének lehetőségét igérte. Abban reménykedtek, hogy a kémiai szerkezet ismeretében a viszonylag kisméretű hatóanyag kémiai szintézisére lehetőséget találnak. A szerkezetkutatásban l944- ben már 59 kutatóhely vett részt teljes katonai titoktartás mellett. Ma már köztudott, hogy a kutató munkát az zavarta, hogy a különböző laboratóriumokban előállított gombatenyészetek szűrletei több mint 20 különféle penicillin keverékét tartalmazták, amelyeket az angol abc 54
55 betüivel jelöltek. Az USA laboratóriumaiban előállított penicillin készítményeknek a fizikaikémiai adatai jelentős mértékben eltértek az Angliában, szeszgyártási maradékon előállított készítmények vizsgálatakor nyert adatoktól. Az egyik esetben G-penicillin (benzil-penicillin), a másik esetben F-penicillin (pentenil-penicillin) volt a főtermék. Angliában a szeszgyártás maradékát (distillers solubl) használták táptalajként, az US kutatói viszont a táptalaj nitrogén forrásaként a keményítőgyártás melléktermékét, a fenilecetsavat is tartalmazó kukorica áztatólé tejsavas erjesztéssel dúsított koncentrátumát alkalmazták. A tenyésztő táptalaj kémiai összetételének meghatározó jelentőségét felismerve javasolták prekurzor alkalmazását. A tápközeghez adott fenilecetsav jelenlétében a kutató laboratóriumokban világszerte főleg benzil-penicillin képződött a tenyésztési ciklus második fázisában. A penicillin szerkezeti képletét megismerve, a királis szénatomok száma visszarettentette a kemikusokat a hatásos H-C == C-H H O H H H CH 3 : : H-C C C C N C C S C CH 3 H-C C-H H O=C N C H G-Penicillin szerkezeti képlete O=C OH vegyület kémiai szintézisétől. Így a biológusokra hárult a fáradtságos feladat, jobban termelő mutáns előállítása. Kezdetben négy gyógyszergyártó vállalat (Pfizer, Lederle, Merck, Squibb) és több kutató laboratórium, (Carnegie Institute, University of Minnesota, University of Wisconsin) kapcsolódott a munkába. A katonai titoktartás szabályait betartó együttműködés a különböző kutatóhelyek között tökéletes volt. A hadügyminisztérium szakemberei ezt az életmentőnek tűnő anyagot minél előbb a szövetséges hatalmak katonakórházaiba kívánták juttatni. Az anyagi és a szellemi erőfeszítés hamarosan eredményt hozott májusában az új gyógyszert már 300 sebesült esetében alkalmazhatták teljes sikerrel a szepszis kialakulásának akadályozására, illetve gyógyítására, sőt 1944-ben már indokolt esetben a polgári kórházak számára is engedélyezték a hatásos anyag, a G-penicillin alkalmazását júliusában a király - a V-bombák rendszertelen érkezése miatt a királyi palota alagsorában rendezett ceremonia keretében - tudományos érdemeit elismerve lovaggá ütötte Sir Alexander Fleminget. A Wisconsin egyetemen nyert mutáns a törzsgyüjteményben - Penicillium chrysogenum NRRL 1951 számon - ma is megtalálható. Ez a törzs szénforrásként laktózt tartalmazó, közel semlegesen tartott, jól levegőztetett táptalajon termeli a Gram-pozitív mikroorganizmusok növekedését akadályozó antibiotikumot. Az inokulum szénforrása azonban glükóz volt, mivel az antibiotikumot termelő gomba spórái csak glükóz szénforráson csírázva képesek növekedni, viszont represszálja az antibiotikum szintézisében szereplő enzimek képződését. Az azetidinon szerkezetű antibiotikumok használatbavételét követően Abraham sejtését igazolva, rezisztens kórokozók jelentek meg a kórházakban, ami indokolta az új antibiotikumok felfedezését célzó nem csekély anyagi hasznot hozó, eredményes kutatómunka szervezését. A katonai költségvetés a háború utolsó éveiben 30 millió dollárral támogatta a program sikerét. A munkában résztvevő kutatóhelyek száma közben 20-ra növekedett. A heti penicillin termelés 1943 végére elérte a 100 grammot (100 ezer NE penicillin akkor 20 dollárba került, ez ma 1 milliárd NE ára). A fejlődés ütemét mutatja, hogy 1944-ben a havi termelés meghaladta a 200 kg-ot, amely 1946-ban évi 32 tonnára emelkedett. Közel tíz év telik el a Penicillium notatum penész által termelt anyag, a penicillin hatásának felismerésétől, a parenterális gyógyászati alkalmazásáig. Ettől az időponttól ugrásszerű a fejlődés. Mi volt vajon a kezdeti sikertelenség oka? Nem más, mint az, hogy a kémiai tevékenység modern módszerei, a kis mennyiségű anyagok ma már megoldott tisztítási eljárásai az időben ismeretlenek voltak, illetve még gyermekcipőben jártak. Nem 55
56 utolsó sorban éppen a biológiailag aktív anyagok tisztítására való törekvés volt a serkentője e mikrokémiai módszerek fejlesztésének. Ezen időszakban a kutató kapacitás jelentős részét világszerte a gyógyszeripar által támogatott, szép eredményekkel dicsekvő (salvarzán, prontozil stb.) kemoterápia fejlesztése vonta el. Ebben az időszakban a kemoterápiai módszerek tökéletesedésével egy új gondolkodásmód terjedt el a fertőzéses megbetegedések leküzdése területén. Tüneti kezelés helyett, a gazdaszervezet károsítása nélkül a kórokozó elpusztítására törekedtek. A benzil-penicillin képződést katalizáló három enzim, [ACVs, IPNs, IAT] működése: L-α-aminoadipinsav L-cisztein L-valin ATP ATP ATP O H H SH CH 3 HOOC CΗ CΗ 2 CH 2 CH 2 C N C CH 2 H H C CH 3 NH 2 A három enzim képződését glükóz represszálja!!! O=C N C H δ (L-α-aminoadipil) - L-ciszteinil - D-valin COOH ACV tripeptid szintetáz (ACVs) Fe ++ O 2 Mg ++ ATP ditiotreitol Izopenicillin -N-szintetáz (IPNs) H H H H O H H H CH 3 : : HOOC C C C C C N C C S C CH 3 H 2 N H H H O=C N C H : izopenicillin-n COOH (δ)-l-α-aminoadipilsavval acilezett 6-amino-penicillansav Izopenicillin aciltranszferáz (IAT) COOH H 2 C CH 2 N H 6-oxopiperidin-2-karbonsav H 2 C CH 2 C = O NAD+ >>>> NADH H H CH 3 H H CH 3 : : : : HOOC-CH-CH 2-CH 2-CH 2-CO-NH-C---C S C CH 3 (BENZIL)--CO-NH-C---C S C CH 3 NH 2 O=C---N C H penicillinaciláz O=C---N C H : : COOH (BENZIL)-CO-S CoA COOH izopenicillin-n (δ)-l-α-aminoadipilsavval acilezett 6-amino-penicillansav G-penicillin (benzil-penicillin) Az aciláz által leválasztott aminoadipilsav átmenetileg gyűrűs formában (6-oxopiperidin-2- karbonsav) szabadul majd adenilsavval vegyesanhidridet képezve újra részt vehet az izopenicillin N bioszintézisben. A történelmi előzményekre gondolva már időszámításunk előtt évvel készült kínai feljegyzések említik, hogy szójababon nőtt gombák nedvével furunkulusok, gennyes fertőzések leküzdhetők. A tudománytörténet Roberts 1870-ben közzétett dolgozatát tekinti az első antibiózisra utaló híradásnak. Ebben a szerző leírja, hogy Penicillium glaucum tenyészetében nem növekednek a baktériumok. Ilyen jelenséget több klinikai laboratóriumban észleltek, sőt több esetben mint figyelemreméltó tapasztalat közlésre is került, mégsem lett 56
57 belőle piacképes termék. A második világháború után azonban a penicllin előállításával a gyógyítás történetében új korszak kezdődött. Az antibiotikumok megjelenése az orvosi gyakorlatban, a fertőző betegségek elleni küzdelemben addig nem tapasztalt forradalmi fejlődést jelentett. Széleskörű gyógyászati alkalmazásuk az átlagéletkor meghosszabbodását jelentette. A genetikailag megalapozott egyéni különbségek, az öröklött ellenálló képesség szintjében jelentkező eltérés szelekciós hatása csökkent. Az addig ritkábban előforduló halált okozó megbetegedések ennek következtében előtérbe kerültek (rák, keringési problémák). A gyermekhalandóság csökkenése, a szeptikus folyamatok kialakulásának megakadályozása, a fertőző betegségek leküzdése a lakosság életkori összetételét jelentősen megváltoztatta. Nem csekély mértékben az antibiotikumok használatának terjedése is hozzájárul a Föld népességének ugrásszerű növekedéséhez, ami áttételesen a társadalmi átalakulásokra is hatással van. Az antibiózis fogalma a biológiában régóta ismert, mint a szimbiózis ellentéte. A kifejezést először Vuillemin használta 1889-ben megjelent dolgozatában. Hagyománytiszteletből antibiotikumnak nevezzük azokat a mikroorganizmusok életműködését gátló vegyületeket, amelyeket a létért való küzdelem jegyében a mikroorganizmusok termelnek. E vegyületek utáni kutatás és a kiválasztott vegyületek nagyipari előállítása a gyógyszeripar dinamikusan fejlődő ágát teremtette meg. Miért nem toxikus a β-laktám antibiotikun? A sejtfal peptid vázának továbbépítését a membrán külső felületén működő transzglikozidáz [TGáz] és a transzpeptidáz [Tpáz] végzi. Ez az utóbbi enzim a muropeptid terminális D-alanin elemének eltávolításával felszabaduló karboxil csoportot illeszti a már meglevő murein váz szabad aminocsoportjához. A transzglikozidáz pedig a peptidoglükán építőelem diszacharid komponensét a glükán polimer terminális hexóz C 4 atomjához kapcsolja. Az újabb építőelem két ponton való kapcsolásával nő a térhálós szerkezetű sejtfal. A transzpeptidáz reakció végbemenetele közben a szubsztrátum átmenetileg acilezi az enzimet és az acilezett enzimről kerül az acilező ágens a muropeptid sejtfal N terminális amino csoportjára. 57
58 Az azetidinon molekula merevített szerkezete éppen a muropeptid építőelem D-alanin-Dalanin dipeptid szerkezetéhez hasonlít. Nem meglepő tehát, hogy nagyfokú szerkezeti analógia miatt a penicillin a sejtfalépítő transztpeptidáz (TPáz) aktív centrumába kötődve acilezi, inaktíválja az enzimet és ezzel a peptidoglükán sejtfal építését akadályozva végül is a baktérium pusztulását okozza A muropeptid D-Ala-D-Ala vége G-penicillin 58
59 Fleming felfedezését követően hosszabb ideig úgy vélték, hogy a penám váz szintézisére csak a Penicillium nemzetség néhány faja jöhet számításba. Az a tapasztalat, hogy a Penicillium tenyészhelyén baktériumok nem fordulnak elő, a létért folytatott küzdelemben való szerepe joggal kérdőjelezhető volt. A hatásos antibiotikum fekszaporodása csak mesterséges körülmények között, semleges körüli ph estén figyelhető meg. Közel húsz év telik el Fleming emlékezetes bejelentése után a második β-laktám típusú antibiotikum felfedezéséig. Brotzu 1948-ban közli, hogy egy Cephalosporium acremonium törzs tenyészlevében igen erős széles spektrumu antibakteriális hatás észlelhető (J. Gen. Microbiol. 47:1952). A fermentlé több biológiailag aktív terméket tartalmaz. A Cepalosporin-C hatóanyagon és a 6- aminopenicillansav D-α-aminoadipinsavval acilezett Gram-pozitív és Gram-negatív kórokozók ellen hatásos származékán, a penicillin-n antibiotikumon kívül egy savanyú természetű szteroid vázat tartalmazó kefalosporin-p-nek nevezett helvolinsav származék jelenlétét is bizonyították. A főtermékként megjelenő Cephalosporin-C-nek elnevezett azetidinon (β-laktám) származékot Abraham és Newton izolálta 1961-ben. Később a törzs rendszertani hovatartozását végző mikológusok az antibiotikumot termelő törzset helyesen Acremonium chrysogenum névvel különítették el a Cephalosporium fajoktól. A termelő törzs nevének változása nem befolyásolta az antibiotikum elnevezését. Megjegyzendő, hogy a Penicillium chrysogenum illetve az Acremonium chrysogenum élőhelyén a savanyú ph viszonyok miatt nem szaporodnak a baktériumok, a nyomokban termelődő β-laktám antibiotikum pedig savkatalizálta reakció keretében elbomlana.) NH 2 H H H O H H H H : : HOOC C C C C C N C C S C H O H H H H O=C N C== C CH 2 O C CH 3 Cephalosporin-C szerkezeti képlete O=C OH (δ)-d-α-aminoadipilsavval acilezett 7-amino-kefémsav.acetát H 2 N H H H O H H H CH 3 : : HOOC C C C C C N C C S C CH 3 H H H H O=C N C H : Penicillin-N (δ)-d-α-aminoadipilsavval acilezett 6-amino-penicillansav COOH A második azetidinon szerkezetű antibiotikum izolálása után szervezett kutatómunka indult újabb β-laktám szerkezetű hatóanyagok előállítása céljából, kiterjesztve a vizsgálatokat a prokariótákra. Hamarosan új vegyületcsoportot a carbapenemeket fedezték fel egymástól függetlenül a Merck és a Beecham kutatói, a Streptomyces fajok által termelt thienamycineket. Ez esetben a kén atom nem az anellált gyűrűben, hanem tioetanolamin származékként tioéter kötésben található. (Streptomyces clavuligerus, S olivaceus, Erwinia carotovora, Serratia sp.) CH 3 Tienamicinek szerkezeti képlete HO-C-H H O CH 3 H H : : : : H C C CH 2 HO S O C C C CH 2 H H O=C N---C== C S CH 2--CH 2--NH 2 O H O=C N C==C S C=C---N---C---CH 3 COOH COOH H O 59
60 Ugyancsak Streptomyces törzsek termelik a. Merck és az Eli Lilly kutatói által felfedezett cefamicineket. A Streptomyces clavuligerus, törzsek által termelt cefamicin származékok különlegessége a 7-α-metoxi csoport megjelenése és az oximetil csoport hiánya, illetve karbamoilezettsége. CH 3 NH 2 H H H O H O H : : HOOC C C C C C N C C S CH 2 O H H H H O= C N C===C CH 2 O C NH 2 Cephamycin szerkezete O=C OH A cefém váz penicillináznak ellenálló tulajdonsága, a nitrogén külső elektronpályáján levő elektronpár részvételéből adódó kvázi konjugált elektron eloszlásból következik Azetidinon gyűrűt tartalmaznak a monolactamok. Képviselőjüket a nocardicint Fujisava izolálta Nocardia uniformis tsuyamanensis tenyészlevéből. A molekula ciszteinből és homo-szerinből, a p-hidroxi-fenilglicin oximino származékából épül fel. A hetvenes évek végén végzett kutatómunka N-szulfonil azetidinon gyürűt tartalmazó Gramnegatív törzsekre ható β-laktám vegyületek felfedezéséhez vezetett. Monobaktám néven tárgyalja őket a irodalomm mert baktériumok (Pseudomonas acidophila, Gluconobacter, Acetobacter, Agrobacterium radiobacter) tenyészlevéből izolálták őket. A 3-aminobaktámsav alapváz savanyú természetét az N-szulfonsav csoport okozza. A β-helyzetű aminocsoportot különféle savak, illetve savszármazékok acilezik. Több esetben a 3-α-helyzetű hidrogént metoxi csoport helyettesíti. CH 3 H H H O H H : : O=C N C------CH 2 --CH C-----C-----N------C-----CH 2 O=C N NH O=C---- N SO 3 K + O=C CH 3 SO 3 K + Az Agrobacterium radiobacter törzs termékében az acilező sav aromás gyűrűt tartalmaz. Ezek az utóbbi azetidinon szerkezetű antibiotikumok gyógyászati felhasználásra nem kerültek, de a megszerzett biokémiai ismeretek termékenyítőleg hatottak az új hatóanyagok előállítását célzó kutató munkában. A század második felében végzett kiterjedt vizsgálatok megerősítették, hogy a bioszintetizáló rendszer a tömlősgombák számos nemzetségben előfordul. A molekuláris biológiai vizsgálatok egyértelműen a gén prokarióta eredetére utalnak, ahol a hatásos anyag megjelenése valóban előnyt jelent a létért való küzdelem szinpadán. A gombák kialakulásuk idején, egy milliárd évvel ezelőtt a prokariótákban addig kialakult örökítő anyagot változtatás nélkül átvették, függetlenül attól hogy az átvétel számukra előnyt jelentett vagy nem. 60
61 A Beecham Laboratórium kutatói által vizsgált Streptomyces clavuligerus tenyészlevéből egy clavulansavnak elnevezett vegyületet is izoláltak. Ez a vegyület a tiazolidin gyűrű helyett oxazolidin gyűrűt tartalmaz. A kén helyett oxigént tartalmazó alapváz acilező aktivitása fokozott mértékű. A C 6 szubsztituens hiánya miatt antibakteriális hatást ugyan nem mutat, de a penicillináz enzimet képes inaktíválni. A clavulánsav hatásmódja A β-laktam bontásáért felelős -SH csoportot a felnyíló laktám gyürű acilezi. Ezt követőleg az oxazol gyürű felnyílása után a reaktív oldallánc stabil tioéter kötésbe lép az enzim másik SH csoportjával. Széles spektrumú félszintetikus penicillin származékokkal együtt adagolva azóta is a penicillin rezisztens fertőzések leküzdésére használják. 61
62 A hatásos termék a fonalas gomba intenzív anyagcserét folytató csúcssejtjében képződik, a növekedési szakaszt követő ugynevezett idiofázisban, amikor jelentős mennyiségű fehérje képződése már nem észlelhető. Ezen ismeret birtokában olyan mutánsokat különítettek el, amelyek az induló sporaszám függvényében pellet képzésre hajlamot mutattak. Ez a tenyészlé egységnyi térfogatában nem csak növelte a csúcssejtek számát, de javult a tenyészet levegőztethetősége is, ami a metabolikus aktivitás szhintje szempontjából is előnyös volt. 62
63 KONVERZIÓRA ALKALMAS SEJTTÖMEG FELHASZNÁLÁSA. Az eljárás kifejlesztésének első lépése a fenilecetsav lehasítását végző mikroorganizmus kiválasztása, illetve teljesítményének fejlesztése. A mikroorganizmus teljesítőképességét agardiffúziós módszerrel célszerű vizsgálni. A kinőtt mikroorganizmus foszfátpufferben készült sejtszuszpenziójához foszfát pufferben készült G-.penicillin oldatot adva 28 C hőmérsékleten egy csepp toluol jelenlétében kerül a termosztátba. (A kontroll cső sejtszuszpenzió nélkül tartalmazza a penicillin oldatot.) Az inkubált csövekből vett minta centrifugált és megfelelően higított felülúszó 0.1 ml-ét Bacillus subtilis spóraszuszpenzióval fertőzött agarlemezen készített lyukakba cseppentjük. Ezek közül három lyukba fenilecetsav klorid került. Az agarlemezt 37 C-on inkubálva egy éjszakán keresztül, a gátlási zóna átmérője számszerüsíti a tenyészet teljesítményét. A kontroll a penicillint tartalmazó reakcióelegy gátló hatását mutatja. A sejtszuszpenziót tartalmazó minta viszont a penicillin gátló hatásának csökkenését jelzi az idő függvényében. A fenilecetsav-kloriddal kiegészített lyukak körül jelentkező gátló hatás a 6-aminopenicillinsav acilezésével nyert antibiotikus hatást számszerüsíti. A fenilecetsavklorid adagolás nem befolyásolja a gátló hatás csökkenését, ha valóban penicillináz inaktíválta a G- penicillin oldatot. Az egyik ipari gyakorlatban alkalmazott eljárás szerint a 2% kukoricalekvárt tartalmazó, enyhén lúgos (ph 8) körülmények között üledékmentesre szűrt táptalj induktorként 0.1% ammonium fenilacetátot tartalmazott. Az enzim katabolikus represszálhatósága miatt a tápközeg metabolizálható szénforrást, így glicerint sem tartalmazhat. Az 1 % rázott lombikban készült Escherichia coli tenyészettel oltott, enyhén kevert (300 fordulat/perc) tenyészet 26 C on mindössze 1 / 5 térfogat levegő átfúvása mellett, 26 óra alatt éri el a maximális enzimszintet, miközben a ph az ammonia termelés hatására az indulási értékről 8,5-8,9-re emelkedik. A kifejlődött tenyészet szupercentrifugával (15000 ford/perc) elkülönítve, 1 / 10 -ed térfogatra sürítve. 10% penicillin-sót tartalmazó csapvízben kerül felhasználásra. A sejtfal áteresztőképességét 0,01% butilacetát adagolása növeli. A reakcióelegy ph-ja a felszabaduló prekurzor hatását ellensúlyozva folyamatos lugadagolással ph-8 értéken tartandó. A reakció befejeztét a savanyodás mértékének csökkenése jelzi. Az újabb centrifugálással eltávolított sejttömeg ismételten felhasználható. A felülúszóból, a széteső sejtekből kikerült penicillin-acilázt tartalmazó fehérje eltávolítása után a reakciótermék, a 6-APS, hűtve, az izoelektromos pontján kristályosítható. A sejtből kiszabaduló aciláz azonban az ismételten felhasználásra kerülő sejttömeg összaktivitásából hiányzik. Az összegyüjtött sejtek csapvizes szuszpenziójához adott 0.1% glutárdialdehid a membránfehérjék és a sejtfal aminocsoportjaival reagálva az enzimet képződési helyére rögzíti. Ez a reagens azonban nem csak a sejtek ellenállóképességét növeli, de természetszerüleg az aciláz aminocsoportjaival is reagál, amit az aktivitás csökkenése jelez. Ezért fél óra kezelés után a feleslegben levő bifunkcionális reagenst ammonia adagolásával inaktíválni kell. Az így kezelt Escherichia coli sejtek mechanikai ellenállóképessége a sejtfeltáráshoz használt ultrahang készülék roncsoló hatását is elviseli. A korabeli hazai viszonyoket jellemzi, hogy azidőben a fermentációs ipar nem rendelkezett olyan szupercentrifugával, amely nagy mennyíségű (20-30 m 3 ) baktérium tenyészetből a sejteket képes lett volna elkülöníteni. Ilyen eszköz beszerzésére valuta sem állt rendelkezésre. A helyzetet felmérve, a hazai eszközállomány felhasználásával mégis sikerült elindítani egy évvel a Natur-ben megjelent közlemény után az ipari körülmények között is gazdaságos 6- APS termelést ( magyar szabadalmi bejelentés) mégpedig rögzített sejtekkel. A maximális enzimszintet elért tenyészethez adott nátrium foszfát oldódása után kálcium kloriddal gél nyerhető, amely megkötve a baktériumsejteket szűrőprés segítségével könnyen elkülöníthető, majd butilacetát jelenlétében a ph 8 folytonos ellenőrzése mellett penicillin hasítására használható. Az átalakítás befejeztével szűrőprés segítségével nyert baktérium mentes 6-APS tartalmú oldat félszintetikus készítmény alapanyagául szolgál. A kálciumfoszfát- 63
64 gél által megkötött sejtek pedig ismételt felhasználásra kerültek. A módszer előnye volt, hogy a gél a sejtlízis miatt kiszabadult enzimet is megkötötte, így a kiszűrt gél enzimaktivitása észrevehetően nem csökkent. 7-AMINO KEFÉMSAV (7-ACA) és szármaszékainak ELŐÁLLÍTÁSA A kefalosporin származékok penicillináz rezisztenciája a kutatók figyelmét a 7- aminokefém váz mikrobiális előállíthatóságára irányította. Amíg az L-α-aminosavval acilezett termék szelektív dezacetilezésre jól használható a Bacillus subtilis, vagy más baktérium aciláza, mindezideig nem sikerült mikróbát találni a kefalosporin-c D-αaminoadipinsav oldalláncának gazdaságos lehasítására. Dezacetil 7-aminokefémsav előállítása cephalosporin C antibiotikumból. Egyetlen meg nem erősített japán közlemény állítja a reakció végbemenetelét. Cseh szerzők két lépéssel oldották meg a feladatot. Az első lépésben a Trigonopsis variabilis D-aminosav oxidáza az oldalláncot ketoadipinsavvá oxidálja hidrogénperoxid és ammonia fekszabadulása mellett, amit már a baktériumokban, például akár a Pseudomonas fajokban is előforduló aciláz könnyen eltávolíthat. A kétlépéses biokonverzió, jelenleg nem versenyezhet a kémiai 7- ACA előállítási eljárással. A feladat géntechnológiai megoldása várható, amennyiben a kefalonsporin-c termelő törzsbe sikerül működöképesen a fenti reakciókat katakizáló enzimek génjeit beépíteni. Az Acremonium chrysogenum-ban természetes körülmények között végbemenő gyűrűtágítási reakció kémiai megfelelője iparilag is végrehajtható. A cefém váz a viszonylag olcsó V-penicillinből könnyen kivitelezhető reakcióval nyerhető. oxidáló reagensekkel előállítható szulfoxid köztiterméken keresztül. A tiazolidin gyűrű átrendeződése p-toluolszulfonsav jelenlétében refluxolva történik. 64
65 A végtermék penicillináz rezisztens, gyomorsav-stabil tehát szájon keresztül adható gyógyszerként forgalmazható.a fenoxiecetsav eltávolítására kémiai, illetve enzimológiai módszer egyaránt használható. A kémiai módszerrel kapott köztes termékről Bacillus subtilis tenyészetével távolítható el a fenoxiecetsav. Az így nyert dezacetoxi-7-aminokefalosporánsav hatásos félszintetikus antibiotikumok kiindulási anyagaként hasznosítható. A kefém alapváz nagyobb stabilitása miatt 7-amino-kefalosporánsav (7- ACA) előállítása jó hatásfokkal történhet kémiai úton a 6-APS előállítási eljárás analógiájára. A aminoadipinsav-n-en tercierbutiloxikarbonillal védett kefalosporin-c dibenzil észterét piridinben, foszfor-pentakloriddal alacsony hőmérsékleten,(cseppfolyós nitrogén) iminokloriddá alakítják. Ebből alkoholizissel imino-éter nyerhető. Az ezt követő hidrolízis, valamint a védőcsoportok eltávolítása ma már a peptid szintézis rutin feladata. A végtermék az első ábrán szereplő hatásos félszintetikus származékok előállításához használható. xxxxxxxxxxxxxxxxxxxxx H H H H : : H N C C S C H O=C N C==C CH 2 OH O=C OH Dezacetil 7-amino-kefémsav (7-amino-cephemsav) szerkezeti képlete (A védőcsoportok eltávolítása után) Az így nyert 7-amino-dezacetoxi cefalosporánsavból készül az előzőkben említett cephalexin (kefalexin), mégpedig tercier-butiloxi-karbonillal védett D-fenil-glicinnel való N-acilezéssel, amit a védőcsoport eltávolítása követ. 65
66 L- GLUTAMINSAV ELŐÁLLÍTÁSA A XX.ik század első felében, főleg konzervipari felhasználásra gabona csírából állították elő ipari méretben a glutaminsavat. A csírázó növényben meginduló fehérjeszintézishez szükséges aminosavak képződésekor ugyanis a glutaminsav amino donorként nagy mennyiségben kerül felhasználásra a transzaminációs folyamatokban. A szétroncsolt csíra vizes szuszpenziójából a glutaminsav - lévén amino dikarbonsav - könnyen kinyerhető. A nyomokban megtalálható természetes eredetű szennyezés nem zavarja az élelmiszeripari felhasználókat. Az acetilénből vagy akrilonitrilből kiinduló kémiai (Strecker) szintézissel nyert DL glutaminsav csak reszolválás után kerülhetne élelmiszeripari felhasználásra. Tudvalevő, hogy csak az L-enantiomer stimulálja az izlelőbimbókban elhelyezkedő receptorokat, a D enantiomer teljesen íztelen. Az antibotikum termelés céljára kifejlesztett bioreaktor az ötvenes években lehetőséget adott viszonylag elfogadható áron élelmiszeripari alapanyagok előállítására. Japán kutatók eredményei irányították a figyelmet az aminosavak mikrobiológiai előállíthatóságára. Szorgalmas munka eredményeként, mintegy 2000 átvizsgált prokariota és eukariota között több olyan törzs is akadt, amelyik glükózt hasznosítva nagy mennyiségben választott ki glutaminsavat a tápközegbe. A mikroszervezetekről szerzett élettani ismeretek alapján várható volt, hogy ammónia jelenlétében a képződő glutaminsav felszaporodik a környezetben, ha az α-ketoglutársav dekarboxilezésének a lehetősége csökken. Escherichia coli-ból mutagén kezelésssel előállítottak glutaminsavat termelő α-ketoglutarát-dehidrogenáz hiányos törzset. Az élelmiszeripar azonban az E. coli alkalmazásától tartózkodik. Kiemelkedő termelő képességet mutatott egy Gram-pozitívan festhető baktérium csoport néhány variánsa. Kinoshita és munkatársai által izolált baktérium törzset a termelőképességére utalva Micrococcus glutamicus néven írták le (J. Gen. Appl. Microbiol. 3:276) Megjegyzendő, hogy az ipari hasznosíthatóság céljából végzett kutatómunka közben nem rendszertani szempiontból vizsgálják a számukra elérhető mikroorganizmusokat. Ezért a fajnév általában szoros kapcsolatban van a törzs gyakorlati értékével. A nemzetség név pedig a mikroszkópi látvány alapján adatik. Ebből következik, hogy az első felfedezést követően a taxonomus a törzs rendszertani helyét a kornak megfelelő vizsgálatok alapján illeszti rendszertani helyére. Ezt a 30 g/liter glutaminsav termelésére képes törzset a Kiova & Hakko Kogyo Co. Ltd. hamarosan ipari termelő törzsként hasznosította Corynebacterium glutamicum néven. Mások Brevibacterum, Microbacterium, illetve Arthrobacter néven izoláltak glutaminsav termelésre alkalmas egymástól alig megkülönböztethető törzseket. Ezek a Gram-pozitív, nem spórázó, nem mozgó baktériumok a növekedésükhöz biotint igényelnek, amit a természetes élőhelyükön kellő mennyiségben megtalálnak. A törzsekre jellemző, hogy az α-ketoglutarátdehidrogenáz aktivitásuk nem mutatható ki, illetve elhanyagolhatóan csekély, viszont a glutamát-dehidrogenáz aktivitásuk feltűnően magas értéket ér el. A törzsek glutamát termelő képessége az izocitrát-liáz aktivitás genetikai úton történő csökkentésével fokozható. A glükóz szénforrást a Corynebacterium glutamicum törzs köztes anyagcseréje az Embden-Meyerhof-Parnas úton, valamint a pentózfoszfát cikluson keresztül, a trikarbonsav ciklusba táplálja, ahol a szénváz citromsavon, izocitráton keresztül oxidatív dekarboxilezéssel α-keto-glutársavvá alakul. Ebből a glutarát dehidrogenáz hiány miatt felszaporodó termékből reduktív aminálással képződik a glutaminsav. Egy önálló kis ciklusként, a NADP-függő glutamát dehidrogenáz működéséhez a NAPH szintet az izocitrát-dehidrogenáz ekvivalens mennyiségben szolgáltatja. Az oxidatív dekarboxilezéskor képződő redukált kofaktort használja fel a szervezet az ammonium ion 66
67 beépítéséhez. A vegyipar által előállított mikrobiális eredetű nátrium-glutamát évi mennyisége már meghaladja a 350 ezer tonnát. GLÜKÓZ Corinebacterium glutamicum tenyészetben a ATP G o GLUTAMINSAV BIOSZINTÉZIS VÁZLATA hexokináz -16 kj ADP NADP + NADPH G-6-P 6-PG G6P dehidrogenáz (-NADPH) G6P-izomeráz +1,6 kj NADP + 6-PG dehidrogenáz CO 2 F-6-P NADPH ( ATP, citrát) ATP Pi F6P-kináz bisz-foszfatáz Ru-5-P (+ADP,+AMP) epimeráz kj ADP Xu-5--P F-1,6-P 2 aldoláz +23,98 kj foszfoketoláz P i DHAP GAP +7.6 kj izomeráz Acetil-P NAD + P i GAP dehidrogenáz + 6,279 kj NADH 1,3-PG ADP PG kináz -18,84 kj ATP CoA-SH P i 3-PG PG mutáz + 4,43 kj foszfotranszacetiláz 2-PG PG enoláz >H 2O + 1,8 kj P-E-P ADP -31,39 kj PEP- piruvát kináz (+FDP citrát) karboxiláz ATP CoA-SH GDP TPP CO 2 NAD + NADH P i CO 2 PIRUVÁT >Acetil-S-CoA CO 2 PEP CO 2 ATP piruvát dehidrogenáz ( ATP ) karboxi piruvát karboxiláz (+acetil-coa) kináz GTP ADP citrát szintetáz ( ATP ) OXÁLACETÁT NADH CIT--RÁT almasav dehidogenáz 28,046 kj akonitát hidratáz NAD + ALMASAV IZOCITRÁT malát szintetáz Acetil-CoA NAD(P) + izocitrát- (+ADP) H 2O (gyenge) izocitrát liáz dehidrogenáz fumaráz NAD(P)H GLIOXILÁT CO 2 FUMARÁT SZUKCINÁT α-ketoglutarát FADH 2 FAD (+ADP) NADPH szukcinát dehidrogenáz glutamát dehidrogenáz NH 3 (-ATP) NADP + Net: 3 NADH + ATP (zárojelben) a szabályozó metabolitok L-GLUTAMINSAV A glutaminsavat termelő szervezetekben az izocitrát dehidrogenáz és a glutaminsav dehidrogenáz aktivitás lényegesen meghaladja az átlagot. Az enzim működését a felgyülemlő glutaminsav ugyan gátolja, de ha ennek a terméknek az eltávolítása megoldódik, akkor a glutaminsav termelés akadálytalanná válik. A ciklus működésének a feltétele a rendszer anaplerotikus feltöltése. Ezt a piruvát karboxiláz és a malát enzim végzi. A karboxiláz szerepét igazolja, hogy jelzett szén-dioxid jelenlétében a glutaminsav karboxil csoportja 67
68 mindig radioaktív. A piruvát karboxiláz egy nagy moltömegű, négy alegységből álló alloszterikusan szabályozott enzim komplex, amelynek működését az acetil-coa serkenti. Az enzim lizin oldalláncához biotin kapcsolódik. Az ATP segítségével aktíválódó szén-dioxid (foszforsav-szénsav vegyesanhidrid) kapcsolódik a biotin nitrogénatomjához. Ez az aktiv állapotban levő szénsav származék (karboxibiocitin) ideális karboxilező ágens, amely piroszőlősavval reagálva oxálecetsavat képez. Az oxálecetsav feldúsulása nem fenyeget, mert acetil-coa-val reagálva citráttá alakul, amelyből az igen aktív NADP függő izocitrát dehidrogenáz működésének eredményeként α-ketoglutarát képződik. Ammonium ion hiányában azonban a környezetben megjelenik az α ketoglutarát. A foszfoenol-piroszőlősav megjelenéséig a reakciósor önfenntartó az energiagazdag foszfátkötések szempontjából, sőt NADH és NADPH felesleg mutatkozik a G6P- és GAPdehidrogenáz valamint a piruvát-dehidrogenáz működésének az eredményeként. A glioxilát ciklus részvétele az oxálecetsav képződésben glükóz szénforrás használatakor jelentéktelen, különösen az izocitrát-liáz aktivitásban sérült nemesített törzs esetében. Elvileg 1 mol glükózból 1 mol glutaminsav nyerhető. C 6 H 12 O 6 + NH 3 + 1,5 O 2 = C 5 H 9 O 4 N + CO H 2 O Az ipari technológia kialakításakor az elméletileg leggazdaságosabb ideális állapot megközelítésére kell törekednünk annak ellenére, hogy ezt az elméleti értéket az ipari gyakorlatban nem lehet elérni. A 60 %-os átalakítás már elfogadható értéknek tekinthető. A termelés szintjét valójában a szénforrás koncentrációja és a glutaminsav környezetbe való kiválasztásának a sebessége határozza meg. A reakcióterméknek a környezetbe való megjelenése végeredményben természetellenes folyamat, hiszen a köztesanyagcsere értékes termékéről van szó. Ennek ellenére a glutaminsav kiválasztást a membrán áteresztőképességének a megváltozása teszi lehetővé. Tween 60 detergens adagolásával például fokozható az aminosav kiválasztása. A glutaminsavat termelő törzsek csekély szukcinát szükségletét egyrészt a viszonylag gyenge izocitrát-liáz aktivitás, másrészt az oxálecetsavból induló reduktív folyamatok elégítik ki. A glioxilsav pedig acetil-coa felhasználásával a ciklus feltöltését segíti. A kutatómunka eredményeként kiválasztott természetes biotin igényes mutáns tiszta tenyészete a kialakított üzemi körülmények között csak fellazult membrán szintézisére képes. A biotin-szint ugyanis az acetil-coa karboxiláz aktivitást befolyásolva a membrán minőségét befolyásoló zsírsav bioszintézisben okoz zavart. A foszfolipidszintézis zavara az L- glutaminsav spontán kiáramlását segíti elő. Természetes élőhelyén a mutáns biotin igénye nem jelent élettani problémát, mert ez a vitamin a környezetből felvehető. Ez a felismerés lehetővé tette a gazdaságos glutaminsav termelést biotinban gazdag olcsó szénforrás felhasználásával. Ennek felismerése a cukorgyártás melléktermékeként képződő biotinban gazdag melasz felhasználhatóvá vált az ipar számára. Az iparban használt termelő törzs limitált biotin jelenlétében (2,5-3 µg/liter) nagy mennyiségű glutaminsavat választ ki a környezetbe. A baktérium növekedése szempontjából ez nem előnyös. A termelés szempontjából viszont hasznos a visszafogott növekedés. A tápközeg biotin tartalmának növelése (5 µg felett) a prolinképződést serkenti. A biotin mennyiségét tizszeresre emelve glutaminsav helyett a glükózból képződő tejsav és szukcinát jelenik meg a tenyészlében. Oxigén hiány a redukált kofaktorok feldúsulása miatt a tejsav és borostyánkősav képződésnek kedvez, túllevegőztetés hatására viszont az ammonium ion kiűzése miatt a glutaminsav helyett α-ketoglutarát szaporodik fel a tenyészlében. Ipari méretben szénforrásként % szacharóztartalmú táptalajt, melaszt, vagy keményítő hidrolizátumot használnak. A glutaminsav szintézishez szűkséges ammóniát vagy gázalakban levegővel elegyítve vagy karbamid formájában adagolják. Az eljárást ph=8 körüli értéken vezetik. A szénforrás hasznosítás általában meghaladja az 50 %-ot 68
69 A termék kinyerése szempontjából ipari körülmények között 10 %-ot meghaladó glutaminsav szint elérésére törekednek. Ezt a termelési szakaszban adott glükózzal érik el. A tenyésztés hőmérsékletének emelése rontja az oxigén beoldódását, azaz a tenyészet oxigénellátását. Általában o C-on vezetett 5-6 napos fermentációs lépés végén, a beadott szénforrás felhasználása után nyert sejtmentes szűrletből történik a glutaminsav elkülönítése. A táptalajban maradó cukor zavarja a glutaminsav kristályosodását. A sejtmentes szűrletet hevítve besűrítik, a képződő csapadékot forrón kiszűrik. A nyert éles szűrletet hagyják lehűlni. 90 o C-ról induló kristályosodáskor apró, ß formának nevezett tűkristályokat nyerünk, amely sok esetben az anyalúg szennyezését is magába hordozza. 70 o C-on vezetett kristályosodás körülményei között az α-formát, prizma alakú kristályokat nyerhetünk. Egy 1975-ben bejelentett US szabadalom ( ) szerint a Brevibacterium divaricatum NRRL B-231 törzs inokulum táptalaja literenként 40 g glükózt, 1 g K 2 HPO 4 -ot, 0,5 g MgSO 4.7 H 2 O-t, 1 g élesztőkivonatot és 8 g karbamidot tartalmazott A tenyésztés 16 órán keresztül 35 o C-on folyt. A 6 % oltóanyaggal oltott főfermentáció tápközege literenként 121 g glükózt, 5 g ammónium acetátot, 6 g keményítőhidrolizátumot, 1,2 g kálium-foszfátot, 1,2 g kálium szulfátot, 6 g magnézium szulfátot, kevés vasat és mangánt tartalmazott. A tenyésztés megindításakor 0,65 ml olajsavat adagolnak literenként. A ph-t ammónia adagolásával 7,8 értéken tartják. A sejtszaporodási szakasz végén általában a 14-ik órában a tenyésztés hőfokát 32 o C-ról 38 o C-ra emelik. A glükóz szint 1 %-ra csökkenésekor kezdik a glükóz adagolást, ami általában literenként 160 g glükóz adagolását jelenti. A levegőztetést úgy szabályozzák, hogy az elmenő levegő széndioxid tartalma ne haladja meg a 4.5 térfogat % értéket. A tenyészlé glutaminsav tartalma általában 35 órás korban éri el a 100 g/liter értéket. Glutaminsav teremelés ecetsavból Nagyipari méretben kémiai úton előállítható ecetsavból is gazdaságosan nyerhető glutaminsav. Ecetsav szénforrásból természetesen a feltöltést a glioxilát ciklus végzi. 3 HOOC-CH 3 + NH 3 + 1,5 O 2 = HOOC-CH 2 -CH 2 -HCNH 2 -COOH + CO H 2 O 69
70 ESSZENCIÁLIS AMINOSAVAK IPARI ELŐÁLLÍTÁSA A növényi fehérje aminosav összetétele lényegesen különbözik az állati szervezetben előforduló fehérje aminosav tartalmától. (kukorica: 0,2%, zab:0,5%, árpa: 0,4%, búza: 0,6%, szója 2,3%, élesztő 3,4%, tejpor:2,5%, húsliszt: 2,6% lizint tartalmaz) Ezért a növényi fehérje jobb hasznosulása takarmányként elősegíthető megfelelő arányban adott esszenciális aminosavakkal. A nem teljes értékű növényi fehérjét az állati szervezet csak saját aminosav igényének megfelelő mértékben, a saját fehérjére jellemző aminosav arányban képes hasznosítani. A tenyészállatok a számukra fölösleges aminosavakat az anyagcsere enzimrendszerének segítségével lebontják, átalakítják, a fölös szénvázat pedig elégetik. A takarmány kukorica 0,11% metionint 0.21% lizint 0,36% treonint tartalmaz a takarmány búza 0,23% metionint 0,61% lizint 0,50% treonint tartalmaz a takarmány árpa 0,17% metionint 0,41% lizint 0,36% treonint tartalmaz a szójaliszt 0,53% metionint 2,31% lizint 1,41% treonint tartalmaz. Az ideális takarmányban a lizin hiány kielégítése után a treonin és a metionin igény jelentkezik. A mikrobiológiai eljárással nyerhető aminosavak közül általában a lizin, a triptofán és a treonin kerül felhasználásra tápanyagkiegészítőként. A világ évi lizintermelése 250 ezer tonna. A treoninból 10 ezer tonna kerül felhasználásra. Az évi triptofán termelés mindössze tonna. Az állattápszerben évenként felhasznált 300 ezer tonna DL metionin kémiai szintézissel készül, mert a sertések és a szárnyasok racemáza a DL-metionin 100%-os hasznosulását biztosítja. Ha a kukorica lisztet 0.2% lizinnel kiegészítjük, akkor a fehérje hasznosíthatósági hányadosa 0.85-ről 1.05-re emelkedik, de ha 0.2% lizinen kívül még 0.07% triptofánt is adagolunk akkor a hasznosíthatósági hányados 2.55-re emelkedik. A rizs fehérjetartalmának kiegészítése 0.2% lizinnel és ugyanennyi treoninnal a hasznosíthatósági hányadost 0.5-ről 2.61-re növeli. Még a szójaliszt is kiegészítésre szorul. Kémiai szintézissel nyerhető metioninnal válik teljes értékűvé. A szabályozási mechanizmus az élő világban azonos elvek szerint működik, de nemek, fajok sőt törzsek esetében, a szabályozás kivitelezésében eltérések lehetnek. A szakember éppen ezeket a kivitelben jelentkező különbségeket hasznosítja. Az esszenciális aminosavak fermentációs úton folyó előállítása nem más, mint a modern mikrobiológiai alapkutatás eredményeinek gyakorlati alkalmazása; nem kevesebb, mint az aminosavbioszintézis szabályozó mechanizmusainak megismerését követő tudatos genetikai munka eredményeinek ipari gyakorlatban való használata. A citoplazmában folyó nitrogén anyagcsere didaktikai szempontból anabolikus és katabolikus folyamatra különíthető. Valójában a két folyamat egymáshoz kötődve, egymást kiegészítve működik szoros kapcsolatban a szénhidrát anyagcserével. A mikrovilág ide sorolt anabolikus folyamatai életfontosságúak az egész élővilág fennmaradása szempontjából, mert az esszenciális aminosavak bioszintézisére, a légköri nitrogén megkötésére, bizonyos vitaminok, cobalamin származékok előállítására csak a mikrovilág, ezen belül a prokariota flóra képes. Az aminosavakat aktiváló enzimek csekély mérvű szubsztrát-specifitása csak meghatározott összetételű aminosav készlet [pool] esetén tudja biztosítani a trns állomány helyes feltöltését. A fölösleges aminosavak eltávolítása tehát létkérdés a mikróba számára. A koncentráció viszonyok, az arányok bármilyen okból bekövetkező eltolódása, hibás trns feltöltést és ennek következményeként hibás, működésképtelen fehérjék szintézisét okozhatja. Nem meglepő tehát, hogy a bioszintézis és a lebontás szabályozottságának bonyolultan, de eredményesen működő mechanizmusa alakult ki az evolúció során. A gazdaságos életvitel érdekében a citoplazmában található peptidázok lebontják a működésképtelen fehérjéket, építőelemeik más irányú felhasználásra kerülhetnek. 70
71 A fehérjeszintézishez szükséges aminosavak előállítására az úgynevezett vad törzsek általában képesek. Ez a bioszintézis a citoplazmában, a szénhidrátanyagcsere köztestermékeit hasznosítva, nem csekély energia befektetéssel illetve anyagfelhasználással szigorúan szabályozott körülmények között folyik. Az aminosav készlet optimális minőségi és mennyiségi összetételét a bioszintézis első reakcióját katalizáló enzimre ható visszacsatoló (feed-back) mechanizmus szabályozza, szükség szerint gátolva vagy serkentve működését. Egy-egy aminosav feldúsulása a túlélés érdekében a lebontására specifikus enzimrendszerek képződését indukálja. Esszenciális aminósavak szabályozott szintézise ASZPARTÁT aszparaginsavból (Escherichia coli) [HomTre] [Liz] {strukturgén NEVE} ATP [1] izoenzim képződését [Tre és Ile] gátolja {lysc} aszpartát-kináz [2] izoenzim képződését [Met] gátolja [1] [2] [3] [3] izoenzim képződését [Liz] gátolja ADP [1a] izoenzim képződését [Tre] gátolja ASZPARTIL FOSZFÁT [2a] izoenzim képződését [Met] gátolja NADPH feed-back gátlás jelzése: Tre] Liz] Hom] Ile] Val] Leu] {asd} aszpartátfélaldehid-dehidrogenáz NADP + ASZPARTÁT-ß-FÉLALDEHID = Liz] [Tre] [Met] NADH {dapa} dihidrodipikolinát-szintetáz {thra} {hom} homoszerin-dehidrogenáz piruvát [1a]- [2a] NAD + 2,3-DIHIDROXI-DIPIKOLINÁT HOMOSZERIN Tre]= NADPH ATP {dapb} dehidrogenáz {thrb} homoszerin-kináz NADP + ADP = Met] 1 -PIPERIDIN-2,6-DIKARBOXILÁT HOMOSZERIN-FOSZFÁT szukcinilcoa + szukcinil-coa H 2O aciltranszferáz {dapc} acil transzferáz {thrc}treonin-szintetáz CoA-SH Pi N-SZUKCINIL- TREONIN O-SZUKCINIL 2-AMINO-6-ΚΕΤΟ- Ile] = HOMOSZERIN L-PIMELÁT { ilva} treonin-dezamináz piruvát NH 3 α-ketoizovalerát α KETOBUTIRÁT cisztein Glutamát = Leu] /\ {dapd}transzamináz ipm.szintáz [Val]= { ilvbn} AHS szintetáz cisztation γ-szintetáz α-ketoglutarát acetil-coa < piruvát > szukcinát Ν-SZUKCINIL-L,L-2,6- α-izopropil- α-aceto- CΟ 2 α-aceto- DIAMINOPIMELÁT MALÁT LAKTÁT α-hidroxi- CISZTATION BUTIRÁT (dape)deszukciniláz NADPH NH 3 ipm.izomeráz {ilvc} reduktáz,mutáz cisztation ß-liáz szukcinát NADP + piruvát L,L -2,6- ß-IZOPROPIL- α,β DIHIDROXI- α,β DIHIDROXI- HOMOCISZTEIN DIAMINO- MALÁT IZOVALERÁT ß-METILVALERÁT PIMELÁT NAD {ilvd}dihidroxisav-dehidráz N 5 -metil-fh 4 {dapf} epimeráz ipm.dehidrogenáz metil-transzferáz H 2O FH 4 NADH mezo-2,6- α KETO- α KETO α KETO-ß- DIAMINO- IZOKAPROÁT IZOVALERÁT METILVALERÁT PIMELÁT glutamát glutamát ( lysa}dekarboxiláz transzamináz-b {ilve} transzamináz-b CO 2 α-ketoglutarát α-ketoglutarát LIZIN LEUCIN VALIN IZOLEUCIN METIONIN Az Escherichia coli-ban kialakult mechanizmus aszparaginsavból indulva nem csak a lizin finoman szabályozott képződését, de más esszenciális aminosavak treonin, metionin és az elágazó szénláncú valin, izoleucin és leucin bioszintézisét katalizálja. Az összetett rendszer 71
72 gazdaságos működtetését egyrészt izoenzimek működése biztosítja, másrészt a kritikus elágazási pontokon a végteremék okozta visszacsatolás (feed-back mechanizmus) hatása érvényesül. Megjegyzendő, hogy a diaminosav a peptidoglükán sejtfal felépítés nélkülözhetetlen eleme A sejtfal felépítés bifunkcionális építőelemként szerepel az L-lizin, az L,L-diaminopimelinsav illetve a mezo-diaminopimelinsav. Az igényeknek megfelelő mértékben oxálecetsavból képződő aszparaginsav foszforilezését első lépésként három represszálható izoenzim végzi. Az izoenzimek képződését szelektíven az összekapcsolódó bioszintézis út végtermékei (az egyiket lizin, a másikat metionin, a harmadikat pedig az izoleucin) represszálják. Ez indokolt, mert az aszpartilfoszfátból (aszparaginsav-foszforsav vegyesanhidrid) NADPH igényes reduktív hidrolízissel képződő aszparaginsav-ß-aldehid valójában olyan közös köztes termék, amely kiindulási anyag az említett aminosavak bioszintéziséhez. A szabályozás enzimszinten is érvényesül. A lizinnel represszálható izoenzim működését a lizin - mint végtermék - alloszterikusan is gátolja. Az izoleucinnal és treoninnal represszálható izoenzim működését pedig két végtermék, a treonin és a homoszerin befolyásolja. A metioninnal represszálható izoenzim biztosítja a rendszer működéséhez szükséges aszpartát-ß-félaldehid alapszintet. Az aszpartát β-félaldehid piruváttal reagálva dihidrodipikolinát-szintetáz (dapa) jelenlétében 2,3-dihidrodipikolináttá kondenzálódik. A kondenzációt végző enzimnek a specifikus aktivitása a végtermék képződését meghatározza, működését a lizin felszaporodása alloszterikusan szabályozza. Az aszparaginsav-ß-félaldehidből két NADPH függő homoszerindehidrogenáz izoenzim végzi a többi esszenciális aminosav képződéséhez szükséges homoszerin szintézisét. Az egyik képződését metionin represszálja, a másik dehidrogenáz képződését pedig a treonin és az izoleucin felszaporodása befolyásolja. Ez utóbbi működését a treonin, mint végtermék alloszterikusan is gátolja. A homoszerinből indul a cisztationon és homociszteinen keresztül vezető metionin szintézis. Az első enzim aktivitását a homoszerinszukciniltranszferáz működését a metionin, mint végtermék alloszterikusan gátolja. A homoszerinből induló treonin szintézis első enzimének, a homoszerin-kináznak a működését természetszerűleg a treonin szabályozza alloszterikusan. Az izoleucin a treoninból induló bioszintézis első enzimének a treonin dezamináznak a működését gátolja alloszterikusan, ugyanakkor represszálni képes a bioszintézisben résztvevő enzimek képződését. Az izoleucin szintézisút második enzimének, az acetohidroxisav szintetáznak a működését a valin gátolja alloszterikusan, mivel a valin szintézis szempontjából ez szerepel első enzimként. Az enzim közös használata miatt az ebből adódó veszélyt úgy oldja fel a rendszer, hogy a treonin dezamináz által szolgáltatott α -ketovajsav előnyt élvez a piruváttal összehasonlítva a kondenzációt katalizáló enzim aktív felületén. A leucin szintézishez a valin szintézis köztesterméke az α-ketoizovaleriánsav szolgál kiindulásul. A négy reakciólépést igénylő bioszintézis első enzimének az acetil-coa-t igénylő izopropilmalát szintetáznak a működését a leucin gátolja alloszterikusan. Az elágazó szénláncú aminosavak bioszintézisének bonyolult szabályozottsága külön figyelmet érdemel a közösen használt enzimekre való tekintettel. A zavartalan fehérjeszintézishez ugyanis a három aminosav optimális aránya szükséges. Erre való tekintettel bármelyik aminosav feldúsulása, vagy a környezetben való megjelenése esetén indukálódik a lebontást végző enzimrendszer, amely azután kiegyenlíti az eltérést. A körülményektől függő mértékben a mikroba a bioszintézisben érdekelt enzimek mennyiségét a represszor eltávolításával - azaz a struktúrgének átírásának engedélyezésével - a szükségletnek megfelelő mértékre növelheti (derepresszió). Mivel a fehérjeszintézis anyag és energiaigényes folyamat, a gazdaságos életvitelt szolgálja a szabályozást finomító második rendszer (attenuator) működése, amely végül is a feltöltött trns mennyiségének függvényében engedélyezi a bioszintézisben érdekelt gének átírását. 72
73 AZ AROMÁS AMINOSAVBIOSZINTÉZIS SZABÁLYOZOTTSÁGA (L-triptofán előállítása ) A reakciósor piacképes termékének tekinthető L-triptofán bioszintézisének és a szintézisút szabályozottságának felderítése a XX-ik század ötvenes éveiben folyó alapkutató munka eredménye. Az aromás aminosavak szintézisét végző enzimrendszer a három aminosav hiányában nagy mennyiségben képződik. Csak az általuk termelt aminosavak felszaporodása csökkenti ezen enzimek képződését. A fölös mennyiségben jelenlevő aminosav, esetünkben a triptofán a represszor fehérjéhez, az pedig a triptofán operon promoter régiójához kötődve megakadályozza az RNS-polimeráz működését. Ezzel a triptofán operonban kodolt enzimek szintézisét represszálja. A mikrobiális bioszintézis a pentóz-foszfát ciklusból leágazva, foszfo-2-keto-3-dezoxiheptonát-aldoláz által katalizált reakcióval indul. A rendszer enzimszinten és génszinten szabályozva izo-enzimek felhasználásával teljesíti feladatát. A bioszintézis köztes termékeinek a képződése is feed-back szabályozás alatt áll, mert nem csak az esszenciális aromás aminosavak szintézise, de egyes vitaminok (PAB, nikotinsav, etc.) képződése is innen ágazik el. A szabályozás finomítása céljával Escherichia coli esetében a foszfoenol-piroszőlősav és az eritróz-4-foszfát kondenzációját három izoenzim (foszfo-2-keto-3-dezoxi-heptonát aldoláz) katalizálja. Mindegyik aktivitását alloszterikusan valamelyik végtermék és egy köztes-termék befolyásolja. Az egyik fenilalaninra és prefénsavra, a másik triptofánra és korizminsavra érzékeny, a harmadik működését pedig tirozin és prefénsav gátolja. Az aromás bioszintézisút első végtermékek által szabályozott izoenzimek által katalizált reakciója (Escherichia coli) a 3-DEZOXI-D-ARABINO- HEPTULONSAV-7-FOSZFÁT képződését segíti A második polivalensen szabályozott alloszterikus enzim a shikiminsav-dehidrogenáz, amelynek működését alloszterikusan négy ligandum, a tirozin, a fenilalanin, a korizminsav és a prefénsav befolyásolja. Az aromás bioszintézisúton a shikiminsav képződését katalizáló, shikimát dehidrogenáz végtermék álzal történő szabályozása 73
74 Az aromás aminosavak pentózfoszfát ciklusból leágazó, szabályozott bioszintézisét katalizáló reakciósort bemutató vázlaton jól követhető a teljes folyamat FOSZFO-ENOLPIRUVÁT + ERITRÓZ-4-FOSZFÁT foszfo-2-keto-3-dezoxiheptonát-aldoláz [ Phe] [ Tir ] [ Trp ] [ pref] [ kor] [ pref] 3-DEZOXI-D-ARABINO-HEPTULONSAV-7-FOSZFÁT dehidrokvinát szintetáz Pi 5-DEHIDROKVINÁT 5-dehidrokvinát-dehidratáz H 2 O 3-DEHIDROSHIKIMINSAV NADPH shikimát dehidrogenáz [ Phe][ Tir][ pref][ kor] NADP + D-SHIKIMINSAV ATP shikimát-kináz ADP SHIKIMINSAV FOSZFÁT piruvil-shikimát-foszfát szintetáz P-E-P 5-ENOLPIRUVIL--SHIKIMINSAV-3-FOSZFÁT korizmát-szintetáz Pi KORIZMINSAV korizmát mutáz asszociátum{ Glutamin antranilát szintetáz trpe gén Glutamát [ Trp] Piruvát [ pref] }asszociátum ANTRANILÁT P-R-PP }asszociátum antranilát-foszforibozil-transzferáz trpd gén PREFENSAV PP [ Phe] N-(RIBOZIL-5'-FOSZFÁT-ANTRANILÁT NAD + [ Tir] foszforibozil-antranilát izomeráz H 2 O ENOL-1-o-KARBOXIFENILAMINO-1- pref-dehidratáz pref-dehidrogenáz -DEZOXIRIBULÓZ-FOSZFÁT CO 2 NADH H 2 O indol-3-glicerin-foszfát szintetáz trpc gén CO 2 CO 2 INDOL-3-GLICERIN-FOSZFÁT FENIL-PIRUVÁT p-ho-fenil-piruvát L-SZERIN <<<<<<Glutamát >>>> triptofán szintetáz (tetramer) transzamináz GAP 2 α alegység trpa gén >> α-ketoglutarát <<< 1 β 2 alegység trpb gén >> L-FENILALANIN L-TIROZIN L-TRIPTOFÁN 74
75 A szerkezeti képlet feltüntetése a tanulmányozó számára segítheti a folyamat követését PEP + eritrózfoszfát Deox.arabino.hept.7-P Dehidrokvinát COOH O=C-H DAHP CH 2 OPO 3 Dehidrokvinát szintetáz szintetáz C OPO 3 H-C-OH H-C-OH O COOH / CH 2 H-C-OH P I H-C OH CH + NAD + NADH CH 2-OPO 3 CH CH 2 OH [Phe, tir, triptofán ] (végtermék szabályozás) Enolpiruvinilsikimátszintetáz- 3-P Sikimát-kináz ATP COOH HC==C CH P I COOH O 3PO CH CH CH O C OH CH 2 Korizmát-szintetáz p I COOH PEP HC==C CH O 3PO CH CH CH OH OH COOH HC==C CH HO CH CH CH OH OH - Shikimát Dehid rogenáz HO COOH \ / H 2C C CH 2 O=CH CH CH OH Dehidrokvinát -dehidráz H 2O COOH HC==C CH O=CH CH CH OH OH KORIZMÁT ANTRANILÁT FOSZFORIBOZILANTRANILÁT COOH Antranil szintetáz COOH Antranilátfoszforibozil HOOC OH OH Gln Glu OH HC C==CH CH 2 HC C==C NH 2 transzferáz HC=CH C CH CH HC CH CH O C COOH HC CH=CH H 3C CO COOH PRPP PP I HC=CH C NH CH O CH OH [Phe ] [Tir ] CH 2OPO 3 [Tri ] (végtermékgátló) Korizmát-mutáz Foszforibozil-antranilát-izomeráz HOOC CH 2 CO COOH HOOC OH OH OH \ / HC C CH Prefenát-dehidrogenáz HC=CH C C CH CH HC CH CH HC=CH C NH CH CH 2OPO 3 OH NAD + Prefenát-dehidráz Indolglicerofoszfát szintetáz H 2O NADH CO 2 CO 2 CO 2 H 2O HC-CH=C-CH 2-CO-COOH OH OH HC CH=CH Fenilalanintranszamináz Glutamát α-ketoglutarát HC-CH=C-CH 2-CO-COOH HC CH=CH L-FENILALANIN HC CH=C CH 2 CO COOH C CH=CH OH Tirozin-transzamináz Glutamát α-ketoglutarát HC CH=C CH 2 CO COOH C CH=CH OH L-TIROZIN triptofánszintetáz HC=O H-C-OH H 2-C-OPO 3 HC CH C C CH CH HC=CH C NH CH CH 2OPO 3 NH 2 HOCH 2 CH-COOH SZERIN GAP L-TRIPTOFÁN NH 2 HC CH C C CH 2 CH HC=CH C NH CH COOH 75
76 Az összefoglaló vázlaton feltüntetett, és zavartalanul folyó bioszintézisút korizminsavnál elágazik. A két arómás aminosav, a fenilalanin és a tirozin szintézise irányába a korizmát szintetáz termékéből a prefénsav képződést két korizmát-mutáz izoenzim katalizálja. Az egyik izoenzim fenilalaninra, a másik tirozinra érzékeny. Mindegyik asszociátumot képez a megfelelő aminosav szintézisét katalizáló következő enzimmel: a fenilalaninra érzékeny izoenzim a prefenát-dehidratázzal, a tirozinra érzékeny pedig a prefenát-dehidrogenázzal alkot komplexet. Az asszociátumok működését triptofán serkenti. Ez a hatás a két aminosavnak a fehérjeszintézis szempontjából kedvező arányban való képződését segíti. Phe Tir Fenilalanin és tirozin bioszintézis szabályozása korizmáttól a ketosavképzodésig A fenilalanin és a tirozin szintézis utolsó lépését azonos enzim, egy-egy glitaminsavval működő transzamináz katalizálja. 76
77 A triptofán finoman szabályozott képződése két enzimből álló komplex, az antranilát szintetáz és az antranilát-foiszfo-ribozil-transzferáz együttműködésével indul. A két alegységből álló antranilát szintetáz, szoros asszociátumot képez az ugyancsak két alegységből álló antranilát-foszforibozil-transzferáz-zal. Az antranilsav szintézishez nélkülözhetetlen glutaminkötőhely ugyanis csak az ép asszociátumon alakul ki. A komplex azonban a képződött végtermék (triptofán) jelenlétében szétesik. Most kap feladatot a foszfo-ribozilantranilát izomeráz, amely a foszforibozilantranilsavból kialakítja az enol-1-okarboxifenil-amino-1-deoxi-ribulóz foszfát szerkezetet. Ez utóbbit CO 2 és víz eltávolításával az indol-3-glicerol-foszfát szintetáz indol-glicerol-foszfáttá alakítja. Amint látható létrejött az indolváz. 77
78 Az utolsó enzimkatalizálta történést a négy alegységből álló triptofán-szintetáz katalizálja, amely az indolt a szerin harmadik szénatomjához kapcsolja egy glicerinaldehid-3-foszfát felszabadításával. Az indol-3-glicerol-foszfátból két gén terméke a trpa gén kodolta 2 α alegység és a trpb kodolta β 2 (dimer) alegységekből álló triptofán szintetáz szerinre cseréli a glicerinaldehid 3-foszfátot és így létre hozza azt L-triptofánt A képződött termék érzékenysége a ß- szénatomhoz kapcsolódó delokalizált elektronszerkezetű indol jelenlétével magyarázható xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx 78
79 Yanofsky és munkatársai a triptofán szintézisért felelős enzimek képződésének szabályozási mechanizmusát molekuláris biológiai szinten vizsgálva egy olyan új általános jellegű felismerésre jutottak, amely az aminosavval töltött trns addig csak sejtett szabályozó szerepének módjára magyarázatot adott. A triptofán szintetáz mrns információ előtt egy 160 bázis terjedelmű szabályozási feladatot ellátó attenuátor régiónak elnevezett szakasz található. Ennek a szakasznak az a feladata, hogy a promoter régiónak a represszor fehérje távoztával bekövetkező felszabadulása után meginduló RNS-polimeráz működését megszakítsa az esetben, ha a triptofánnal töltött trns a fehérjeszintézis szempontjából még kielégítő koncentrációban jelen van a rendszerben. A mrns esetében a kódolt fehérjére vonatkozó információ előtt minden esetben a riboszómához kötődést biztosító szakasz található. A fehérje szintézise az eubaktériumoknál AUG kodonnal (formilmetioninnal) indul. A DNS-szálon akár egy génen belül is egyidejűleg több RNS-polimeráz is működhet, sőt az enzimről folyamatosan letekeredve képződő mrnsfonalakon körülbelül 40 bázisnyi távolságban már megindul a fehérjeszintézis. Ebből az következik, hogy meglehetősen kis területen egyidejűleg.változatos biokémiai reakciók (RNS ill. fehérje-szintézis) tömege folyik. Ezt a lehetőséget használja ki az élő szervezet az aminoacilezett trns szabályozó szerepének a megjelenítésére. Ennek keretében a promoter régión kötődő RNS-polimeráz, az operátorhoz kötődő represszor fehérje távoztával elindulhat a struktúrgén irányába. Az RNS-polimeráz működése azonban megszakad akkor, ha a megfelelő aminoaciltrns a fehérjeszintézis szempontjából szükséges koncentrációban jelen van a mikrobasejtben. Az első vázlat a triptofán represszáló hatását ábrázolja. A triptofánnal terhelt represszor az operátor régióhoz kötődve akadályozza az RNSpolimeráz működését. I promoter I operátor I attenuátor I trpe gén I trpd gén I trpc gén I trpb gén I trpa gén I represszor RNS > triptofán polimeráz (korepresszor) A második vázlat a derepresszió jelenségét ábrázolja. Triptofán hiányában a korepresszor (esetünkben triptofán) nélküli represszor fehérje lemozdul az operátorról. Ennek következtében elindulhat a transzlációs folyamat. Az RNS polimeráz elindul a struktúrgének felé. Közbe I promoter I operátor I attenuátor I trpe gén I trpd gén I trpc gén I trpb gén I trpa gén I RNS polimeráz ----> represszor truktúrgének: E = antranilát-szintetáz D = antranilát-transzferáz C = IGP-transzferáz B = triptofán-szintetáz B A = triptofán-szintetáz A találja azonban vezérpeptid információ tartalmát, aminek átírása után folytatja tevékenységét a struktúrgének irányába, Az egyre növekedő RNS-hez tapadva a riboszóma, a szükséges acilezett trns jelenlétében megkezdi a vezérpeptid szintézisét. Siker esetén a képződő RNS szálon történő komplememt képzés az RNS-polimeráz eltávolítását okozó termináló hurok képződését okozza. trip-trns hiányában a vezérpeptid szintézise leáll, az RNS-polimeráz pedig zavartalanul folytathatja útját a struktúrgének irányába. 79
80 A triptofán operon átírását befolyásoló attenuátor régió szabályozó szerepe Met Lys Ala Ile Phe Val Leu Lys pppaaguucacguaaaaaggguaucgacaaugaaagcaauuuucguacugaaa riboszómakötő szakasz G G G l U y AUAAGAGAUUAAACAAGUUUUUUUU - ACCCAUAGACUAACGAAAUGCGUA - U T A C - G C - G r C G - C C - G p A G - C A - U T A G - C C - G r U C - G U U G p G G - C A C A C G - C U G r A A U G - C g A U A A U - A T A ez a komplementer bázisok által stabilizált termináló G - C h C hurok leszorítja a σ faktor nélküli RNS-polimerázt a templátról A - U r A C U S CAAAAACCGACUCUCUCGAACUGCUG... G - C e ezért a triptofán-szintetáz struktúrfehérje génjei nem íródnak át G - C r G U C G A A A Met Lys Ala Ile Phe Val Leu Lys Gly pppaaguucacguaaaaaggguaucgacaaugaaagcaauuuucguacugaaaggu riboszómakötő szakasz U ACGAAAUGCGU G A A G UCAGA - UUCACC U U - A G Trp-tRNS hiány miatt a vezérpeptid képződése az UGG kódnál A - U G leáll, a polimeráz a komplementer kötések átrendeződése miatt C - G C zavartalanul tovább haladva megkezdheti a triptofán-szintetáz C U G struktúrgén átírását C - G C A A A G - C C C - G U C - G U C - G C AAUAAGAGAUUAAACAAGUUUUUUUUCGGGCGAGUAAUCCG C A 1 A A G U C C A Met Gln Thr... AAUGCAAACACAAAAACCGACUCUCUCGAACUGCUG... Elkezdődik a struktúrgének információja előtt elhelyezkedő attenuátor szakaszon kódolt rövid vezérfehérje szintézise. A vezérpeptid információja a promoter régiót követő rövid riboszómakötő szakasz után, a szokásos AUG kodonnal kezdődik és UGA kóddal végződik. Az RNS-polimeráz által szintetizált mrns-láncon, amelynek szekvenciája több komplementer szakaszt is tartalmaz, a polimerázt 40 bázisnyira követve folyik a vezérfehérje képződése. A két szintézis (mrns-képződés és peptidszintézis) közel azonos sebességgel folyik. A vezérpeptidtől 30 bázisnyi távolságra, jóval a struktúrgéneket iniciáló kodon elött található az a termináló hurok, amely képes lenyomni a σ-faktor nélküli, lazán kötődő polimerázt a templátjáról. A termináló hurok azonban, csak az esetben alakulhat ki, ha a vezérpeptid szintézise zavartalanul megtörténik. Az RNS-polimeráz tehát leválik az operonról és σ-faktorral kiegészülve újra kezdi működését a promoter szakasznál. Az anyagveszteség csekély, mert a jelenlevő proteázok a funkció nélküli vezérpeptidet lebontják, és az aminosavak visszakerülnek a bioszintetikus folyamatba. Triptofán esetében ez a vezérpeptid mindössze 14 aminosavból épül fel, de 80
81 szekvenciájában közvetlenül egymás után két triptofánt tartalmaz, amely százalékos arányban különösen nagy gyakoriságot jelent, mivel a prokariótákban általában 100 aminosavra jut egy triptofán. A vezérfehérje gén a triptofán szintézist katalizáló struktúr gének előtt található Ha azonban triptofánnal töltött trns hiányában megakad a vezérfehérje képződése, akkor a vezérszekvencia bázissorrendjében levő komplementer szakaszok által létrehívott új párkapcsolat kialakulása (73-84 és bázisok között) nem engedi kialakulni a termináló hurkot. Ennek következményeként a polimeráz a kritikus szakaszon tovább haladhat a struktúrgének AUG kezdő kodonja felé, és hamarosan megkezdi, ez esetben az aromás aminosav képződését katalizáló enzimek, tehát a triptofán-operonba szerveződött struktúrgének átírását. Elkezdődik az aromas aminosav bioszintézisét végző enzimek képződését irányitó mrns szintézise. ///Az ábra a strukturgének elhelyezkedését mutatja a bázispárok számának a feltüntetésével, az mrns képződését befejező termináló hurok szerkezetével kiegészítve.(lásd az előző ábrán a 4 jelű hurkot.)///// Amikor a végtermék által (feed-back) szabályozott, triptofánt szintetizáló rendszer aktív tevékenysége a vezérpeptid szintéziséhez elegendő triptofanil-trns szintet állít be, a struktúrgének további átírása megszűnik. 81
82 Az acilezett trptrns jelenlétében a termináló (4) hurok leválasztja az átírást végző enzimkomplexet a képződő RNS molekuláról, és ezzel befejeződik az RNS-polimeráz tevékenysége Triptofán hiány esetében a korepresszor (esetünkben triptofán) nélküli represszor fehérje lemozdul az operátorról. Így elindulhat az RNSpolimeráz a struktúrgének felé. Hamarosan megindul a transzlációs folyamat. Elkezdődik a struktúrgének információja előtt elhelyezkedő attenuátor szakaszon kódolt rövid vezérfehérje szintézise. A vezérpeptid információja a promoter régiót követő rövid riboszómakötő szakasz után, a szokásos AUG kodonnal kezdődik és UGA koddal végződik. Az RNS-polimeráz által szintetizált mrns- láncon, amelynek szekvenciája több komplementer szakaszt is tartalmaz, a polimerázt 40 bázisnyira követve folyik a vezérfehérje képződése. A két szintézis (a mrns- és a peptidszintézis) közel azonos sebességgel folyik. A vezérpeptidtől 30 bázis távolságra, jóval a struktúrgéneket iniciáló kodon előtt található az a termináló hurok, amely képes lenyomni a σ-faktor nélküli, lazán kötődő polimerázt a templátjáról. A termináló hurok azonban, csak az esetben alakulhat ki, ha a vezérpeptid szintézise zavartalanul megtörténik. Az RNS-polimeráz tehát leválik az operonról és σ-faktorral kiegészülve újra kezdi működését a promoter szakasznál. Az anyagveszteség csekély, mert a jelenlevő proteázok a funkció nélküli vezérpeptidet lebontják, és az aminosavak visszakerülnek a bioszintetikus folyamatba. Triptofán esetében ez a vezérpeptid mindössze 14 aminosavból épül fel, de szekvenciájában közvetlenül egymás után két triptofánt tartalmaz, amely százalékos arányban különösen nagy gyakoriságot jelent, mivel a prokariontákban általában 100 aminosavra esik egy triptofán. Ha azonban triptofánnal töltött trns hiányában megakad a vezérfehérje képződése, akkor a vezérszekvencia bázissorrendjében levő komplementer szakaszok által létrejött új párkapcsolat nem engedi kialakulni a termináló hurkot. Ennek következményeként a polimeráz a kritikus szakaszon tovább haladhat a struktúrgének kezdő kodonja felé. Hamarosan megkezdi az illetékes aminosav képződését katalizáló enzimek, ez esetben a triptofánoperonban lltező struktúrgének átírását, a mrns szintézisét. Amikor a végtermék által (feedback) szabályozott, triptofánt szintetizáló rendszer aktív tevékenysége a vezérpeptid szintéziséhez elegendő triptofanil-trns szintet állít be, a struktúrgének további átírása megszűnik. 82
83 A σ-faktor nélküli RNS-polimeráz lemozdul a DNS-ről a termináló hurok hatására Hasonló szabályozó rendszer irányítja a többi aminosav szintézisét katalizáló enzimek struktúrgénjeinek átírását is. Példaként vizsgáljuk az aminosavbioszintézist végző enzimek struktúrgénjeinek átírását befolyásoló úgynevezett VEZÉR-PEPTIDEK szerkezetét: Triptofán szintézis enzimeinek képződését akadályozó vezérpeptid szerkezete Met-Lys-Ala- Ile-Phe-Val-Leu--Lys-Gly-Trp-Trp-Arg-Thr-Ser Hisztidin szintézis enzimeinek képződését akadályozó_vezérpeptid szerkezete Met-Thr-Arg-Val-Gln-Phe-Lys-His-His-His-His-His-His-His-Pro-Asp Fenilalanin szintézis enzimeinek képződését akadályozó vezérpeptid szerkezete Met-Lys-His-Ile-Pro-Phe-Phe-Phe-Ala-Phe-Phe-Phe-Thr-Phe-Pro Treonin és Izoleucuin szintézis enzimeinek képződését akadályozó vezérpeptid szerkezete Met-Lys-Arg-Ile-Ser-Thr-Thr-Ile-Thr-Thr-Thr-Ile-Thr-Ile-Thr-Thr-Gly-Asn-Gly-Ala-Gly *(A vastagítás jelzi a szabályozást végző aminósavval acilezett trns ügydöntő szerepét) A vezérpeptidek kémiai szerkezetében az illetékes aminosav több példányban szerepel. Triptofán esetében ez a vezérpeptid mindössze 14 aminosavból épül fel, de szekvenciájában közvetlenül egymás után két triptofánt tartalmaz, amely százalékos arányban különösen nagy gyakoriságot jelent, mivel a prokariótákban általában 100 aminosavra esik egy triptofán. Hisztidin esetében 7 szomszédos kodon igényel feltöltött hisztidil-trns-t. A fenilalanin-szintézis enzimeinek képződését szabályozó vezérfehérje 7 fenilalanil-trns jelenlétét igényli az RNS-polimeráz leválasztásához. Figyelemre méltó a treonin és az izoleucin struktúrgénjeinek átírását szabályozó vezérpeptid szerkezete, amely alternáltan 8 treonin és 4 izoleucinal feltöltött trns-t igényel. Ez esetben tehát a treonin és az izoleucin szintézist katalizáló enzimek génjeinek átírását csak az izolucinnal, illetve treoninnal töltött trns jelenléte akadályozhatja ha mindkét aminosavval feltöltött trns jelen van a rendszerben. 83
84 Az ismertetett átírás mechanizmusát több antibiotikum képes gátolni. Az ansaláncú rifamicinszármazék, például rifampicin az RNS polimeráz ß-láncához kötődve akadályozza az RNSszintézis megindulását. A citosztatikus hatású antibiotikumok közé sorolt actinomycin-d a DNSbázispárok közé ékelődve akadályozza a kettős spirál fellazulását, a két szál szétválását, végeredményben az RNS szintézisét. Megjegyzendő, hogy ez az antibiotikum a DNSreplikációt csak nagyobb koncentrációban alkalmazva képes gátolni, mivel a helikáz nagyobb energiával lazítja fel a dupla spirál bázispárjait, mint az RNS-polimeráz. Mindezt felül szabályozza a szuperfeltekeredett állapot lazítását végző DNS-giráz működése, amit a novobiocin antibiotikum, illetve a magnézium-kötő kinolon származék, a széles spektrumu ciprofloxacin befolyásol. Az optimális aminosav szint fenntartására szolgáló komplex rendszer elemei az élő sejtben: 1, a környezetből történő aminosav felvétel transzportmechanizmusa, 2, attenuáltan szabályozott, mrns képződés által befolyásolt enzimállomány, 3, alloszterikusan ellenőrzött bioszintézis (feed-back), 4, katabolikusan derepresszálható lebontó rendszer (C és N hiány). A bioszintézisben nélkülözhetetlen enzimek sérülése vagy elvesztése a mikroba aminosav igényeként jelentkezik. Ezek a hiánymutánsok (auxotróf törzsek) jól használhatók a mikrobiális genetikai munkákban. A szabályozási mechanizmust érintő genetikai beavatkozással, bizonyos enzimek működésének akadályozásával vagy éppen bizonyos utak serkentésével egy-egy aminosav előállítására gazdaságosan használható mutánsok nyerhetők. Ilyen genetikailag hibás törzsek képességeit glutaminsav, lizin, fenilalanin, triptofán, treonin, izoleucin ipari előállítására hasznosítják. 84
85 ELJÁRÁSOK TRIPTOFÁN ELŐÁLLÍTÁSÁRA Triptofán szintézis L-szerinből és indolból. L-szerinből és indolból a triptofán szintetáz aktivitást hasznosítva gazdaságosan lehet nyerni ezt az esszenciális aminosavat. A triptofán-szintetáz (EC ) minden prototróf mikroorganizmusban előfordul a triptofán bioszintézist végző reakciósor utolsó tagjaként. Az Escherichia coli-ban ez az enzim móltömegű piridoxálfoszfát kofaktorral működő tetramer. Két α alegységből és egy β 2 dimerből épül fel. Az érintetlen tetramer komplex in vivo az indol-3-glicerol-foszfátból és az L-szerinből L-triptofánt és glicerinaldehid-3-foszfátot alakít. A tetramerből könnyen kiszakadó α alegység az indol-3-glicerol-foszfát szétesését katalizálja indolra és glicerinaldehid-3-foszfátra. A β 2 dimer az indolból és L szerinből történő triptofán szintézist katalizálja, de elősegíti a szerin bomlását piruvátra és ammóniára. A tetramer enzim előállítására alkalmas törzset a triptofánigényes mutánsok közül célszerű kiválasztani. Triptofán-limitáció körülményei között növekedő tenyészetben várható a bioszintézisben érdekelt enzimrendszer derepressziója. Miles Escherichia coli-b-ből előállított mutánsában az oldható fehérje 16 %-át tette ki ez a fehérje. A sejtekben képződött enzim nem csak triptofán előállítására, de különböző triptofán analógok szintézisére is sikerrel használható. Az enzim stabilitása és aktivitása nagyobb, mint a tisztított formában kinyert β 2 dimeré. Növeli az enzim aktivitását, piridoxál-5- foszfát adagolása a reakcióelegybe. A mellékelt ábrán jól követhető a P-5-P szerepe a kvinoidális köztes tereméken keresztül vezető reakcióban. A triptofán szintetázt nagy mértékben termelő, triptofán előállítására alkalmas törzs olyan fenilalanin és tirozin igényes mutáns, amely 5-metiltriptofán, 6-fluor triptofán, 4-metil-triptofán, p-fluor-fenilalanin, p-amino-fenilalanin triptofán-hidroxamát jelenlétében is képes nöni, tehát ezekre rezisztens. Az enzimkomplex az elölt sejtbe glutárdialdehiddel rögzíthető és bioreaktorban két hét felezési idővel használható triptofán előállítására. 85
86 A termelő törzs kiválasztásakor fontos szempont a szerin szintézis színvonala. Szénforrásként a megvalósított eljárásokban glükózon kívül glicerin, alkohol, növényi olaj azaz zsírsav is számításba jöhet. A különbség a regenerálandó kofaktorok mennyiségében szembetűnő. A technológiai paraméterek optimálása éppen ezt az élettani feladatot van hivatva megoldani. 1 / 2 glükóz +NH NAD + szerin + 2 NADH + 2 H + glicerin + NH NAD + szerin + 2 NADH + 2 H + zsírsav + NH FAD + 3 NAD + szerin + 3 FADH + 3 NADH + 3 H + 2etanol+NH 3 +ATP+FAD+3NAD + +2NADP + szerin+adp+fadh 2 +3NADH+2NADPH A szerin bioszintézis látszólag szabályozatlanul folyik, mivel élettani szempontból ez az aminosav központi szerepet játszik az anyagcserében. Bioszintézise a glikolízis köztes termékéből a glicerinsav-3-foszfátból indul 2-keto-3-foszfo-glicerinsavon és foszfo-szerinen keresztül, de képződhet C 1 töredék felhasználásával glycinből is. A glicerinsav-3-foszfát hexózból, pentózból, triózból származhat, de a Krebs-ciklusban fontos szerepet játszó oxálecetsavból a glükoneogenezis lépéseit hasznosítva is képződhet. Szerin szintézis glicerin <<<<<< ATP >>>>>>ADP NAD + glükóz <<<<<<<<< ATP >>>>>>>>ADP glükóz-6-foszfát fruktóz-6-foszfát <<<<<<<< ATP >>>>>>>>ADP fruktóz-1,6-biszfoszfát NADH α-glicerinfoszfát dihidroxi-aceton-foszfát Glicerinaldehid-3-foszfát NAD + >>>>>> P i NADH <<<<<< Glicerinsav-1,3-biszfoszfát glioxilát izocitrát citrát szukcinát <<<<<< ADP >>>>>>ATP <<<<< FAD H 2 C-O-PO 3 H 2 >>>>>FADH 2 H-C-OH 2-P-G COOH 3-P-G fumarát <<<<<< NAD + >>>>>>NADH H 2 O H 2 C-O-PO 3 H 2 C=O malát ac.coa 2-keto-3-P-G COOH glutamát <<<<< NAD + α-ketoglutarát >>>>>NADH H 2 C-O-PO 3 H 2 GTP GDP H-C-NH 2 ac.-coa oxálacetát P-E-P 3-foszfo-szerin COOH NADPH CO 2 Pi <<<<<NADP + acetil-coa NADH H 2 C-OH acetaldehid <<<<<<<< NAD + zsírsav β lebontás H-C-NH 2 NADH FADH 2 COOH <<<<<NAD + <<<<<<< FAD L-SZERIN etanol zsírsav Prekurzorok alkalmazásával vezetett triptofán szintézis esetében célszerű 5-metiltriptofánra, illetve 5-fluor-triptofánra rezisztens derepresszált mutánsokat használni. A derepresszált 86
87 törzsekben a bioszintézisben szerepet játszó enzimszint a vad törzsben mért érték 200- szorosát is elérheti. A magasabb enzimszint a céltermék túltermelését, főtermékként való megjelenését eredményezheti. A derepresszált mutáns előállítására egyszerű lehetőség adódik, mert a trns aciláz a szubsztrátspecifitásának függvényében a sejten belül kialakuló koncentrációviszonyoktól függő mértékben a novekedő tenyészethez adagolt aminosav analóggal tölti fel a triptofaniltrns-t. Ennek az a következménye, hogy a sejtfehérje olyan arányban tartalmazza az aminosav analógot, amilyen arányban a trns aciláz tévedett. Élettani következményként az elkészült hibás fehérjék arányában az egyed életképessége akár a teljes életképtelenségig csökkenhet. Nagyszámú egyed között - mutagénekkel való kezeléssel fokozható mértékben - előfordulhatnak olyan változatok (variánsok), amelyek nagyobb mennyiségben termelve a természetes aminosavat, kompetitiven kiszorítják az analógot az acilezési reakcióból, azaz a növekedést gátló aminosav analóg jelenlétében is képesek növekedni. Az aminosav túltermelésnek különböző oka lehet. Adott esetben, a bioszintézisben érdekelt enzimek képződnek nagyobb mennyiségben, tehát konstitutívan derepresszált mutánsokká váltak. Más esetben a szabályozási mechanizmus hibájával, a feed-back érzékenység elvesztésével találkozunk. A központi szerepet betöltő aminosavak szabályozás nélküli képződése transzamináz glicerinsav-3-foszfát G-2-P PEP piruvát glutamát oxálacetát <<<< NAD + foszfoglicerát-dehidrogenáz transzamináz >>>NADH α-ketoglutarát aszpartát hidroxi-piruvát-3-foszfát foszfatáz <<<<< Glutamát hidroxipiruvát transzamináz >>>>α-ketoglutarát glicin szerin-foszfát P i glioxilát transzamináz acetil-coa >>>> foszfatáz >>>>CoA-SH L-SZERIN glutamát α-ketoglutarát acetil-szerin piruvát alanin transzamináz H 2 S >>>>>>> >>>>>acetát H 2 O NH 3 + H 2 S dezamináz cisztein L-tripotofán szintézishez természetesen kémiai szintézissel előállított L-szerin is használható, ha például valamilyen piacképes hasznos termék előállítása közben mellék-termékként szaporodik fel. Erre példa a sejtfal bioszintézist gátló D-cikloszerin gyártás esete. 87
88 D-CIKLOSZERIN Ezt az egyszerű szerkezetű, (D-4-amino-3-izoxazolidon) Gram-pozitív és Gram-negatív baktériumok növekedését gátló, főleg antituberkulotikumként felhasználásra került antibiotikumot az ötvenes évek derekán, négy egymástól független kutatócsoport, négy különböző Streptomyces törzs fermentlevében találta (Harned: S. orchidaceus. Shull: S.lavendulae. Harris: S. garyphalus. Kuzikava: S. roseochromogenes ). Ez az amfoter tulajdonságú savstabilis vegyület ketoenol tautomériát mutat, vizes oldatban hosszabb idő alatt dimerizálódik. H + + NH 3 NH 3 NH 2 NH 2 N N / \ // \ H 2 C C H H 2 C C H H 2 C C H H 2 C C H H 2 C CH C O O C-OH O C-O O C=O O C-O O C HC CH 2 \ // \ // \ / \ // \ // \ / N N N N N N H H 4,5 ph tautomer alakok 7,4 ph dimer forma A cikloszerin gyűrűs formában merevített szerkezete a D-alanin térszerkezetéhez hasonlít. Kompetitív gátlással akadályozza azokat a reakciókat, ahol az enzim szubsztrátuma, illetve terméke D-alanin. Gátolja a D-Ala-D-Ala szintetáz működését, de nem akadályozza az aminosav aktíválását. Gátolja a D-alanin és a D glutaminsav-transzamináz aktivitását és gátolja az alanin permeáz működését. Az alanin racemáz működését az enzim kofaktorával a piridoxál-foszfáttal reagálva gátolja. A cikloszerin a D-Ala-D-Ala peptid szintézisét gátolva a sejtfal alkotórészeként nélkülözhetetlen muramilpentapeptid képződését akadályozza. Mikroszkoppal vizsgálva - cikloszerin jelenlétében - a baktériumokon a penicillin hatásához hasonló jelenség figyelhető meg. A sejtfalszintézis szelektív gátlása miatt először szferoplaszt képződik, majd a sejtek szétszakadása a baktériumtenyészet feltisztulásához vezet. Antibakteriálás hatását csökkenteni lehet in vivo D-alanin adagolásával. A D-cikloszerin például hatástalan a TBC-vel fertőzött egereken, illetve a tengerimalacon, mivel ezen két állatfaj magas endogén D-alanin szintje hatástalanítja az antibiotikumot. Humán TBC fertőzés leküzdésére izonikotinsav hidraziddal szokták alkalmazni. Egyszerű kémiai szerkezete miatt a szabadalmi bejelentésekben leírt bioreaktorban kivitelezhető módszereket ipari méretben csak átmenetileg használták. Előnyösebb és gazdaságosabb a kémiai szintézis D- szerin metilészterből. A szerin kémiai szintézisével nyert racemátból a D-forma elkülönítendő. A melléktermékként velszaporodó L-szerin adott esetben L-triptofán előállítására használható. 88
89 Triptofán szintézisre használható a reverzbilisen működő triptofanáz (EC ) amely természetes körülmények között egyes baktériumokra jellemzően a triptofán lebontására szolgál. (E. coli indol+, Klebsiella ndol, Salmonella indol, Edwardsiella indol+, Providencia indol+ viszont a Proteus és a Shigella lehet indol+, illetve ndol is) indol + piruvát + ammónia > triptofán + víz Ez a reakció végeredményben a baktériumokban előforduló jól ismert katabolikus α,β-eliminációs reakció visszafordítása. Nakazawa (1972) Proteus rettgeri tenyészetét használta enzimforrásként. A katabolikus enzim képződését nem csak a könnyen metabolizálható szénforrás, de a nitrogénforrás is represszálja. A promoter régióhoz kötődő aktív glutamin szintetáz és egy speciális fehérje komplexe serkenti az RNS polimeráz aktivitását az esetben, ha egyidejűleg a camp-cap komplex is jelen van. A glutamin-szintetáz 12 alegységből álló óriás enzimkomplex, amelynek működését glikokoll, alanin, triptofán, hisztidin, karbamoil-foszfát, glükózamin, citidil-trifoszfát, és adenilsav multivalensen gátolja. Teljesen inaktív állapotban minden alegységének tirozil oldallánca adenilsavval van acilezve. A fenti aminonitrogént tartalmazó vegyületek limitált mennyiségben való jelenléte a tápközegben előnyösen hat a nitrogénkatabolikus enzim képződésére. Szénhidrátban és egyéb nitrogénforrásban szegény táptalajon az enzim képződését triptofánnal indukálni lehet. A rekció közben felszabaduló indol azonban gátolja a baktérium növekedését. A gátló hatás polyoxietilén-alkil-fenoléter adagolásával védhető ki. Ez a detergens mikrocseppekbe zárva megköti a felszabaduló indolt. Ílyen körülmények között növekedő baktérium oldható fehérjetartalmának 6 %-a triptofanáz. A kifejlődött tenyészetből elkülönített sejtekhez literenként 80 g nátrium-piruvátot, 80 g ammónium-acetátot, 10 mg piridoxál-5-foszfátot, 1 g nátrium-szulfátot és 100 ml metanolban oldott 60 g indolt adva enyhén alkalikus (8,8 ph) körülmények között, 34 o C-on keverve, két nap alatt 75 g triptofán képződik. Az egyensúlyi reakciót teljessé lehet tenni, ha a reakcióelegyből a triptofánt kivonjuk, például inozinnal, ami a képződő triptofánnal vízoldhatatlan komplexet ad. Az eljárást sikerrel használják triptofán analógok, például 5- hidroxitriptofán előállításra. 89
90 L-TRIPTOFÁN NYERÉSE RACÉM HIDANTOIN SZÁRMAZÉKBÓL A teljességre törekedve nem hagyható figyelmen kívül egy bakteriális enzim a hidantoináz ipari alkalmazása L-triptofán előállítására. Ez az 5-hidroximetil-hidantoinnal indukálható enzim (Sano Agric. Biol.Chem. 41:819) a racém indolil-metil-hidantoinból első lépésben az L-triptofán N-karbamoil származékát állítja elő, majd második lépésként távolítja el a karbamoil csoportot. Az átalakítandó szubsztrátum gyakorlatilag teljes mértékben eladható termékké alakítható, mert az indolil-metil-hidantoin 8-9 ph tartományban spontán racemizálódik. Az indukált baktériumsejtek membránjának átjárhatóságát a reakcióelegyhez adott toluol fokozza, de az elegy baktériumos fertőződését is megakadályozza. H H O H H H C=C C C CH 2 C C N-H H C=C C C CH 2 C COOH H C=C C N C-H H-N C=O H C=C C N C H H N C=O H H Η 2 Ο H H NH 2 hidantoináz N-karbamoil-L-triptofán rac-indolilmetil-hidantoin formamid ph = 9 D-indolilmetil-hidantoin L-triptofán Bioszintézis prekurzorok felhasználásával. Az egyik jól ismert módszer antranilsav adagolással valósítható meg. Ez esetben az antranilsav adagolás a triptofánra érzékeny alloszterikusan feed-back szabályozott reakciót kerüli meg, ha a másik szubsztrátum az 5- foszforibozil-1-pirofoszfát kellő mennyiségben rendelkezésre áll. Ilyen eljárásokat dolgoztak ki Hansenula anomala, Candida utilis, Bacillus subtilis antranilsav auxotróf törzseit használva. A növekedő szakasz után kezdődik az antranilsavat tartalmazó tápoldat adagolás vigyázva arra, hogy az antranilsav szint 0,5 % alatt maradjon. sejt tömeg + o L-triptofán A táptalaj összetétele (H. anomala) o 5 % glükóz 0,3 % ammónium-nitrát 50g -- o 5g -- 0,05 % káliumdihidrogén-foszfát o 0,2 % dinátriumhidro-foszfát + o ,1 % nátriumdihidrogén-foszfát o % magnézium-szulfát o + 0,01 % nátrium-klorid o o + 0,001 % ferro-szulfát o + 0,1 % élesztőkivonat o + 2 % kalcium-karbonát o + 0,2 % antranilsav induláskor, o majd naponta adagolva h o 90
91 L-TRIPTOFÁN BIOSZINTÉZIS SZÉNHIDRÁTBÓL ÉS AMMÓNIÁBÓL. Leggazdaságosabb előállítási módja a mutánsok felhasználásával végzendő teljes bioszitézis. Az iparilag használható mutáns előállításához az eddigi tapasztalatok alalpján a Corynebacterium glutamicum vad törzsből indulva fenilalanint és tirozint igénylő, kétszeresen auxotróf korizmát-mutáz hiányos törzset izoláltak. Ez a törzs limitált fenilalanin és tirozin jelenlétében tenyésztve képes volt csekély mértékben triptofánt termelni, mivel a felszaporodó korizminsav továbbalakulását nem tudta megakadályozni a triptofánra érzékeny, alloszterikusan szabályozható antranilát szintetáz. A következő feladat éppen a feed-back mechanizmus hatástalanítása volt. Ennek a feladatnak a teljesítése érdekében egymás után végzett műveletekben 5-metil-triptofánnal, 5 fluor-triptofánnal, 4-metiltriptofánnal és triptofánhidroxamáttal szemben rezisztens mutánsokat izoláltak. Ezek közül választották ki a jobban termelőket és azokkal végezték tovább a klasszikus genetikai tevékenységet, az előállított mutánsok vizsgálatát. Ezek között a triptofán analógra rezisztens Tir Phe mutánsok legjobbjai már 0,5% triptofánt termeltek mélyfermentációs körülmények között. Logikailag várható, hogy fokozódik a triptofán termelés, ha a fenilalanin és tirozin érzékeny alloszterikus enzimek feed-back érzékenységét is sikerül megszüntetni. A további genetikai beavatkozásokban p-fluor-fenilalaninra, majd p-amino-fenilalaninra rezisztens mutánsokat állítottak elő. Végül tirozin-hidroxamátra, illetve fenilalaninhidroxamátra rezisztens módosulatokat választottak ki. Szisztematikusan mindíg a legjobbnak látszó mutánssal végezték a következő genetikai módosítást. A munka eredményeként olyan genetikailag stabilnak tekinthető törzshöz jutottak, amely 12 g triptofán/liter termelésére volt képes. A táptalaj 10 % redukáló cukrot, 2 % ammónium-szulfátot, 1 % kukoricalekvárt és 2 % kalcium-karbonátot tartalmazott. A mutáns triptofán termelőképessége ezen kezelések ellenére sem vesztette el teljesen a fenilalanin és tirozin érzékenységét. A technológia kialakításakor ezen két aminosav aktuális koncentrációját a táptalajban analitikai mérésekkel limitáló szintre kell beállítani. Aminosav forrásként előnyösen használható a szójafehérje hidrolízátuma fenilalanin kiegészítéssel. A két aromás aminosav egymáshoz viszonyított arányának meg kell egyezni a mutáns fehérje-összetételében mért aránnyal. A táptalaj szuboptimális aromás aminosav tartalma meghatározó jelentőségű. Ettől való eltérés lefelé elmaradást okoz a növekedésben, felfelé pedig a triptofán termelés csökkenésével jár. A beállított táptalaj összetétel csak a fermentációs paraméterek szigorú állandósága esetében hozhat eredményt. A hőmérsékletben, a levegőzésben és a keverésben, vagy az induló sejtszámban való változtatás az aminosavak felhasználását is változtatja, ami végül a triptofán termelésben való eltérés okozója lehet. Meghatározó jelentősége van a technológiai előírások betartásának és a fermentációs berendezés, műszaki színvonalának. A glükózból folyó triptofán szintézis oxigén igényéről fogalmat alkothatunk a vázlat alapján. A szintézis energia és anyagigénye független a törzstől, ezért az Escherichi coli adatai nem különböznek a C. glutamicum -étól. Egyetlen triptofán képződéséhez 3 glükóz és 2 ammónia szükséges, miközben 1 piruvát, 4 szén-dioxid képződik, miközben nyolc redukált kofaktor és öt ADP regenerálása terheli az anyagcserét. A piruvát elégetés is az anyagcsere feladata. Mindent összevetve triptofán molekulánként 11 NAD(P)H visszaoxidálását kell gazdaságosan megoldani. 91
92 L-ASCORBINSAV ELŐÁLLÍTÁSÁHOZ KAPCSOLÓDÓ TEVÉKENYSÉG A scorbut megszüntetésére alkalmas C-vitamint 1928-ban Szent-Györgyi Albert izolálta mellékveséből és narancsból (Biochem J :1387) Az aszkorbinsav kémiai szerkezetének felderítése több kémiai szintézisút kimunkálására késztette a vegyész kollegákat. Egymástól függetlenül 1933-ban Reichstein és Haworth kutatási eredményei kerültek ismertetésre (Nature :280; Soc ). Az akkor kidolgozott eljárások elemeit ma is használják az évenként tübb mint harmincötezer tonna C-vitamin előállítására. A Reichstein által kidolgozott eljárás egy ma is gazdaságosan végezhető mikrobiológiai lépést hasznosít. A glükózból katalitikus hidrogénezéssel nyert szorbitot mikrobiális eljárással dehidrogénezik. A képződött szorbóz diacetonidjából kémiai oxidációval állítják elő a diaceton-2-keto-gulonsavat, amiből további kémiai lépések (észterezés, enolizáció, laktonképzés) segítségével nyerik a 2-keto-gulonsav-metilésztert, amiből lugos hidrolízissel aszkorbinsav képződik. A dehidrogénezés Acetobacter xylunum, illetve Acetobacter suboxydans törzsekkel végezhető, amely baktériumok növekedésükhöz pantoténsavat, p-aminobenzoesavat és nikotinsavat igényelnek. Igényüket a tápközeghez adott élesztőfőzet elégíti ki. Az eljárás másik kényes pontja a katalitikus hidrogénezéssel előállított szorbit jelentős nikkel tartalma. A Raney-nikkel katalizátorból származó szennyezést nehéz maradéktalanul eltávolítani. Az ipari eljárásban az Acetobacter suboxydans olyan mutánsát alkalmazzák, amely literenként 24 mg nikkelt tartalmazó biokonverziós elegyben is működőképes. Az első időkben az átalakítást felületi tenyészetben végezve néhány hét alatt a szubsztrátum 80-85%-a szorbózzá alakult D-fruktóz és 5-keto-D-fruktóz képződése mellett. Az első sikeres sűlyesztett oxidációs eljárásról Wells és munkatársai 1937-ben számolnak be. Szorbitot és élesztőkivonatot tartalmazó táptalajon nőtt tenyészet óra alatt a szubsztrátum 60 %-át 92
93 átalakította. Mivel a szbsztrátumként szereplő szorbit nagyobb koncentrációban gátolja a baktérium növekedését, ezért 10%-ról indulva menetközben több részletben ráadagolva érik el a 35 %-os szorbit koncentrációt. A táptalaj 0.5% élesztőkivonatot tartalmaz. A tenyészközeg kémhatását 4-6 ph-n tartva 30 C-on indítják, majd a hőmérsékletet 35 C-ra emelve 30 óra alatt fejeződik be a biokonverziós folyamat. Ezt követőleg az elegy alakos elemeit, a sejtroncsokat szűréssel eltávolítják, majd ionmentesítés után enyhe körülmények között töményítve, a hűtött anyalugból a szorbózt kristályos formában elkülönítik. Általában a kiindulási szorbitra számolva %-os hozammal nyerhető a szorbóz C-vitamin gyártáshoz szűkséges minőségben. A teljes folyamatot tekintve 2 kg glükózból 1 kg C-vitamint lehet előállítani. A mikrovilág enzimkészlete a szintézisút további kémiai lépéseinek helyettesítésére is képes. David Perlman vezetésével dolgozó kutatócsoport Gluconobacter melanogenus biokémiai aktivitását hasznosítva a szorbózból szorbozont, L-xilohexogulozt állított elő, amit ugyanez az organizmus tovább oxidált 2-keto-L-gulonsavvá. Ez utóbbi átalakítás azonban meglehetősen gyenge eredménnyel ment, ezért különböző mikroorganizmusok aldehidoxidáló képességét hasonlították össze, sőt az eredeti vad törzsből mutagén kezeléssel igyekeztek alkalmasabb enzimforráshoz jutni. Végül is egy Pseudomonas putida ATCC törzs felhasználásával 80%-os hatásfokkal lehetett szorbozonból indulva 2-keto-Lgulonsavhoz jutni. A szorbozon koncentrációja azonban alig haladta meg az 5 mg/ml értéket, ami az eredmény gyakorlati használhatóságát kérdőjelezi. Kezdettől felmerült a vágy a katalitikus hidrogénezési lépés kiváltására. A szorbit oxidációban szereplő baktériumok biokémiai aktivitásának a vizsgálata egy alternatív út kimunkálásának lehetőségével kecsegtetett. Közismert volt, hogy az Acetobacter suboxidans glükózból egyetlen fermentációs ciklusban 5-keto-D-glükonsavat állít elő, mégpedig 10 %-nyi glükózt tartalmazó reakóelegyben 90%-os elméleti hozammal. A biokonverziós folyamat részletes vizsgálata két egymást követő lépés szerepét tisztázta. Az első lépés egy FAD enzim által katalizált glukonsav képzés. A folyamat közben képződő FADH 2 regenerálása a légköri oxigén felhasználásával hidrogén-peroxid képződés közben történik. A glukonsav képződést katalizáló hexóz-oxidáz enyhén savanyú körülmények között (ph 5) a membránhoz kötve glükózból δ-glukonolaktont állít elő. Ez a gyürüs szerkezetű, elektromos töltéssel nem rendelkező vegyület könnyen jut a periplazmikus térbe, ahol a membrán külső felületéhez kötött laktonáz glükonsavvá hidrolizálja. A glukonsav tovább alakulását savanyú körülmények között (ph 4.5) a membránhoz kötött 5 ketoreduktáz katalizálja, eltávolítva az 5-ös szénatomhoz kötött hidrogént. Az enzim szerkezete az egyensúlyi reakciót a savképződés irányába tólja. A reakcióelegybe adagolt kalciummal lehet a ketosavképződést fokozni. A reakcióelegybe juttatott kalciumkarbonát elősegíti a vízben rosszul oldódó kálcium-5-keto-glukonát képződést. A reakcióelegybe juttatott kálcium mennyisége kritikus a folyamat kedvező irányba terelése szempontjából. Ekvivalens mennyiségben adva rontja a ketonizáció esélyét, főtermékként glukonsav képződik. A szaporodás és a termékképződés ph optimuma eltér. A semlegesítés csökkenti a baktérium növekedését. A reakcióelegy hőmérséklete is eltérő hatást vált ki. A növekedés 93
94 hőoptimuma C. A glukonolakton képződés sebessége növelhető a hőmérséklet emeléssel, a ketonizáció folyamatát viszont gátolja ez a beavatkozás. Megjegyzendő, hogy az azonos rendszertani egységbe sorolt mikróbák ketonizációs aktivitása jelentősen eltérhet. Ebből a szempontból az Acetobacter suboxidans ATCC 621 törzs a legaktívabb. A teljes biokonverziós folyamatban azonban nem csak a membránhoz kötött enzimek vesznek részt, szerepet kapnak a citoplazmában működő enzimek is. A glükóz a permeáz segítségével glükóz-6-foszfát formájában jelenik meg a citoplazmában, ahonnan egyrészt a NADP + igényes G-6-P-dehidrogenáz és 6-P-G-dehidrogenáz által katalizált uton dekarboxileződve, a pentózfoszfát ciklus igényét, másrészt a citrát ciklust támogató glükolízis által a mikróba energiaigényét elégíti ki. Az Acetobacter suboxidans membránhoz kötött FAD igényes dehidrogenáza Ca++ jelenlétében a glükózból kálcium-glükonátot képez, amit a NAD + igényes 5-ketoreduktáz alakít 5-ketoglükonsavvá. 10 %-os glükóz oldatból 33 óra alatt 25 C-on 90%-os hozammal nyert 5- ketoglükonát-kálciumból katalitikus hidrogénezéssel kalcium L-gulonát és D-glukonát elegyét nyerték. A könnyen elválasztható L-gulonát mikrobiológiai úton alakítható 2-keto-L-gulonsavvá (US.Pat: ). A D-glukonát pedig ismételten visszavezethető a folyamatba. Mások a 2,5-diketo-glükonsav utat előnyösebbnek tartják. Pseudomonas albosesamae ATCC 21998; Acetobacter melanogenus ATCC 9937; illetve az Acetobacter cerinum segítségével kéthárom nap alatt 16.5%-os glükóz oldatból 95%-os kihozatallal nyerhető a 2,5- diketoglükonsav. Tovább lépve az 5- ketocsoport Corynebacterium és Brevibacterium törzsekkel szelektíven redukálható. Sonoyama és munkatársai szerint Acetobacter melanogenus ATCC 9937 és Brevibacteriun ketosoreductum ATCC törzsek kevert tenyészetével glükózból egy lépésben 2-keto-gulonsav nyerhető (US Pat: ). A kevert tenyészet bizomytalan reprodukálhatósága miatt nem került ipari gyakorlatba. 94
95 GLÜKÓZ-FRUKTÓZ CUKORSZÖRP NAGYIPARI ELŐÁLLÍTÁSA Európa népessége minden bizonnyal a történelmi idők elött is ismerte az édesség varázsát. A gyümölcsből főzéssel készített lekvár, a méhek által gyüjtött méz piacképes termék volt. Dél- Amerika felfedezése után, a cukornádból kipréselt sejtnedvből kristályosodó termék, a nádcukor megjelenése Európa élelmiszerpiacán az édesipar fejlődését hozta. A napoloni háborúk idején meghirdetett tengeri blokád következményének felszámolása céljából, a kontinens cukorigényének a tengerhajózástól független megoldása vezetett a répacukorgyártás kifejlesztéséhez. Az így előállított kristálycukor azonban a fejlesztések ellenére sem volt képes a piaci versenyben hátrányos helyzetén javítani. Az enzimek nagyipari hasznosíthatósága a huszadik század 70-es éveiben azonban a mérsékelt égőv édesipara számára is előnyös helyzetet teremtett, ami a répacukor gyártás visszaszorulását hozta. Marshall és Kooi 1957-ben közölték, hogy a Pseudomonas hydrophila a glükózt képes fruktózzá izomerizálni. A felismerés 20 éven keresztül csipkerózsika álmát aludta. A hetvenes években jelentkező cukorhiány, a nádcukor árának hirtelen emelkedése előhívta a feledésből az addig csak irodalmi adatnak tekintett közlést. Kiderült, hogy a felfedezés ipari gyakorlatba vétele jelentős gazdasági hasznot hoz a felhasználóinak. Mikrobiális izomeráz alkalmazásával lehetőség adódott keményítő alapanyagból a nádcukornál gazdaságosabban, 10-15%-kal olcsóbban előállítani az édesipar, konzervipar és a szörp üzemek számára a glükóz-fruktóz keveréket, az invertcukrot. (Az élelmiszeripar a kristályosodó szacharózt minden esetben invertált formában hsználta.) Ennek az ipari eljárásnak az elterjedésével a cukorgyárak elvesztették monopol helyzetüket. Az első üzemek az Egyesült Államokban kezdtek termelni főleg kukoricakeményítőből ban már Japánban is 17 üzem működött. Európa sem akart elmaradni a versenyben. Hazánkban Szabadegyházán épült ilyen szörpüzem. Az eljárás elterjedésében nagy szerep jutott az enzimrögzítési technológia alkalmazásának, mivel így az előálllított biokatalizátort hosszú ideig lehetett folyamatos üzemben alkalmazni. A modern géntechnológiai módszerekkel előállított, és szántóföldi termelésbe fogott Zea mays törzs biológiai tulajdonsága (gyomirtó rezisztencia) a kukoricakeményítő előállítási árát is kedvezően befolyásolja. A piaci helyzet stabilizálásán kívül az eljárás az egészséges táplálkozás szermpontjából is nagy jelentőségű fejlesztésnek tekinthető, mivel a glükóz-fruktóz keverékből a fruktóz kromatográfiás módszerrel könnyen elkülöníthető. A visszamaradó glükózszörp többszöri visszaforgatásával a keményítő a glükóznál kétszer édesebb fruktóz sziruppá alakítható (Starch 1980,32:11; :55). Fruktózt a cukorbetegek is fogyaszthatnak, mivel a glükóznál lassabban szivódik fel és az anyagcserében is lassabban hasznosul. Ez az izomeráz természetes életkörülmények között a mikroorganizmus életbenmaradását biztosítja xilózon, mint egyedüli szénforráson. Ezzel a szénforrással a mikrovilág nap mind nap találkozik, mivel a xilózból álló polimer - a xilán - a hemicellulóz alkotórészeként a növényvilág fontos építőeleme. Ennek a polimernek a lebontását egy indukálható enzim, a xilanáz végzi. A hidrolízis hatására megjelenő pentóz - a xilóz - azonban nem bontható, mert a mikrobiális enzimek között nen akad olyan, amely a xilózból foszforsavésztert alakítana, márpedig ez feltétel a szénhidrátanyagcserébe való bekapcsolódásnak. A mikroorganizmusok egy részében előforduló izomeráz azonban képes az aldopentózt ketopentózzá alakítani, a hatására képződő xilulóz foszforilezésére szolgáló enzim viszont minden mikroorganizmusban működőképes. (A baktériumok egy csoportja xiliten keresztül vezető kerülő úton képes hasznosítani a pentózokat, ennek jelentősége 95
96 azonban csekély.) A xilóz-ketoizomeráz /EC / a szubsztrátum hasonlósága miatt az egy szénatommal hosszabb szénhidrátot, a glükózt is elfogadja szubsztrátumként, és közel azonos sebességgel fruktózt állít elő belőle. Ennek a lehetőségnek a felismerése után nagy számú törzs (Streptomyces, Nocardia, Arthrobacter és Bacillus) enzimkészletét vizsgálták meg, keresve azt a törzset, amelyik ezt az enzimet az ipari elvárásoknak megfelelő szinten termeli. Könnyíti a munkát, hogy egyedüli szénforrásként xilánt tartalmazó táptalajon csak azok a szervezetek növekednek, amelyek egyrészt extracelluláris xilanázt termelnek, másrészt xilóz-ketoizomerázzal is rendelkeznek. Az utóbbi enzim, általában intracelluláris. A tápközegben ez az enzim csak a sejtmembrán károsodása esetén jelenik meg. Mindkét enzim képződését katabolikus represszió szabályozza. Képződésük előfeltétele a könnyen metabolizálódó szénforrás, például glükóz hiánya. A hiány bizonyos kulcsenzim működésének szelektív gátlásával is előidézhető. A xilózketoizomeráz működése felfüggeszthető egy glükóz-analóggal, a szorbittal. Ez a hexit elfoglalja az enzimfehérjén a glükóz-, illetve a xilóz-kötőhelyet és csak fölös mennyiségben adott glükóz vagy xilóz képes kiszorítani onnan. Ezzel a módszerrel a katabolikus represszió hosszabb ideig felfüggeszthető. Az enzimes eljárás gazdaságos kivitelezése szempontjából az izomeráz enzim néhány tulajdonságát is célszerű vizsgálni. Az enzimműködés általában kobaltot igényel. Ezt az iont egészségi okokból el kell távolítani a szirupból. Gyakorlati szempontból ezért fontos, hogy az enzim nagy cukorkoncentráció (40%) esetén is működőképes maradjon, továbbá, hogy. magashőmérsékleten (70-80 C-on), közel fiziológiás ph-nál az enzimet tartalmazó sejtek ne lizáljanak. Egyes eljárások enzimkészítménye 100 C-on is használható nyomásálló reaktorban. Az enzimtermelés céljára azt a törzset választják, amelyik mindezen feltételeknek megfelelve az enzimszint szempontjából is kiemelkedő teljesítményre képes. A törzs szabályozó rendszerét klasszikus genetikai mdszerekkel módosítva sikerült az enzimszintet 60-80%-kal növelni. Géntechnológiai módszerekkel az Upjohn kutatói az enzimképződést négyszeresre fokozták (Appl. Environ. Microbiol :1402). A GYIKI kutatói által kiválasztott törzs a Streptomyces matensis (MNG magyar szabadalom), amelynek a fenntartására egyedüli szénforrásként xilánt tartalmazó táptalaj a legalkalmasabb. Gazdasági meggondolásból az enzimtermelő táptalaj kezdetben glükózt tartalmaz, mégpedig olyan mennyiségben, hogy az a logaritmikus fázis elejéig elfogyjon. A glükóz elfogyásakor a kifejlődött sejttömeghez adott xilóz indukálja az izomeráz képződést. A xilóz mellé adott szorbit a mesterségesen fenntartott látszólagos xilulóz hiányt okozva az izomeráz túltermelését eredményezi. A maximális enzimszint elérésekor, mivel az izomeráz 80 C-on sem inaktíválódik, célszerű a tenyészet felmelegítésével elölni a sejteket és egyben inaktíválni a hőérzékeny proteolitikus enzimeket, megakadályozandó a fonalas sejtek lízisét. 96
97 FRUKTÓZ-SZÖRP ELŐÁLLÍTÁSA AZ ELJÁRÁS KIFEJLESZTÉSE Természetes forrásból származó mikróba szuszpenzió Egyedüli szénforrásként xilánt tartalmazó minimál táptalajra szélesztés, majd egy hét inkubálás 30 C-on Szélesztés xilánt tartalmazó minimál táptalajra Tiszta tenyészet izolálása ferde agar táptalajra A törzs tisztaságának ellenőrzése komplett táptalajra szélesztéssel, ismételt vissza-oltogatás xilánt tartalmazó táptalajra Ellenőrzött tiszta tenyészettel xilózt tartalmazó folyékony táptalaj oltása 48 órán keresztül levegőztetve kerve A kinőtt tenyészetek sejtjeinek elkülönítése centrifugálással A sejtek inkubálása 30 %-os glükóz oldatban 60 C-on A glükóz-fruktóz arány változásának vizsgálata A legaktívabb tenyészetekkel xilózos cső oltása Különböző hőfokon inkubálva napos korban A legaktívabb tenyészetek sejtjeit ismételten 30%-os glükóz oldatba szuszpendálni A legaktívabb törzsekkel optimális hőmérsékleten ideális korú tenyészetet készítünk A tenyészetet elkülönítve glükóz oldatban inkubálni különböző hőfokon 72 óra inkubálás után a sejteket ismételten szuszpendáljuk, legalább hat alkalommal és 72 órában mérjük az G/F arányt és a felezési időt A mérés ismétlése a legelőnyösebbnek talált 3 törzs-zsel Kiegészítve a fehérjetartalom változás vizsgálatával. A törzs fenntartása egyedüli szénforrásként xilánt tartalmazó táptalajon történik. Fokozható az izomeráz kötődése a sejt alkotórészeihez glutárdialdehiddel, amely vegyület az enzim és a közelében levő sejtalkotórész ideális távolságban levő aminocsoportjai között létesít kovalens kötéseket. Az így nyert enzimkészítmény tartós használatra alkalmas. 40 nap folytonos használat után feleződik az aktivitása. 97
98 A glükóz-fruktóz izomeráz az az enzim, amelynek az előállítására folytonos fermentációs technológiát dolgoztak ki. Az eljárást évtizedek óta ipari méretben használja a Novo Industry A/S. Az intracelluláris enzimet egy atipikus termofil Bacillus coagulans konstitutív módon termeli. A tenyésztés hőmérséklete 50 C, ami a közel semleges optimális ph mellett a tápanyagban gazdag táptalajon, a környezetből származó idegen mikróbák elszaporodását nehezítve a tenyészet homogenitását elősegíti. A táptalaj előkészítésére, sterilezésére, az egész apparátus bakteriológiai tisztaságát szolgáló módszerek kidolgozására különös gondot fordítottak. A termelő törzs genetikai stabilitása akár 1000 órás folytonos tenyésztést is elbír a biológiai teljesítőképesség észrevehető változása nélkül. A törzs eredetileg endospora képző volt, ezért a folytonos fermentációs technológia kidolgozása céljából klasszikus genetikai módszerek alkalmazásával olyan stabil nemspórázó mutánsát állították elő, asmelyben a litikus hatású enzimek képződésének a szabályozása nem sérült meg A folytonos eljárás termelékenysége 3-4-szerese a szokásos eljárásnál kapott értéknek, ami a viszonylag nagy higítási rátával magyarázható. (µ = óra 1 ). A nagyobb termelékenység kompenzálja a drágább táptalaj és a költségesebb technológia alkalmazásának hátrányait. Az évtizedek óta használatban levő eljárás a lehető legalacsonyabb glükózkoncentráció mellett, oxigénlimitált körülmények között, a higítási sebesség változtatásával biztosítja a maximális specifikus aktivitású sejtek nagy mennyiségének a képződését. Az élelmiszeripari felhasználáson kívül újabban eredményesen alkalmazzák a glükóz-fruktóz izomerázt a növényekben nagy mennyiségben előforduló xilánból folyó alkoholgyártáshoz (bioetanol). A xilán hidrolízisekor felszabaduló xilóz hasznosítására két lehetőség terjedt el az élővilágban. Az egyik a xilóz redukciója xilitollá, amit egy NADPH függő oxidoreduktáz katalizál. A képződő xilitolt egy NAD függő dehidrogenáz alakítja xilulózzá. Ezt a terméket foszforilezi a xilulokináz, és ezzel a reakcióval indulhat a pentózok alkoholos erjedése. Két xilózból 3 etanol mellett 3 ATP, 4 szén-dioxid és 2 NADH képződik. A NADH REGENERÁLÁSA aerob körülményeket igényel. Anaerob körülmények nem javítják a helyzetet 98
99 A másik megoldás, ha az erjesztendő xilózhoz izomerázt adunk. Ez esetben a szubsztrátum hasznosítása jobb. Három xilózból 5 etanol, 5 ATP, és 5 szén-dioxid képződik. Ez az út lehetőséget ad az élesztők használatára. 99
100 BIOETANOL A szeszgyártás alapanyaga kezdetben a cukorgyártás melléktermékeként visszamaradó melasz volt. A piaci igényekhez alkalmazkodva, adott esetben a cukorkihozatal növelése a melasz mennyiségét csökkentette, ellenkező esetben növelte, ami a szeszgyártásban bizonytalanságot okozott. A szesz iránt növekedő kereslet hatására a keményítő gyárak mellé is szeszgyárak települtek. Először a keményítő és glükóz gyártás hulladékát hasznosították. Később az igényeknek megfelelően adott esetben a keményítő jelentős része alkoholként került a piacra. Az alkohol-gyártás a mezőgazdasági termelésben jelentkező időjárás okozta hullámzásokat pufferként tudta felfogni. Az esetenként bekövetkező túltermelés nem vezetett az eladhatatlan termékek felszaporodására, azaz nem csökkentette a termelési kedvet. A szénhidrát tartalmú termékeket túltermelés esetén (burgonya, kukorica) alkohollá alakítva értékállóan lehetett tárolni. Az alkoholgyártás gazdaságosságát vizsgálva össze kell vetnünk a cukorgyártásban hasznosított mezőgazdasági termékek előállítási költségeit és a belőlük nyerhető hasznos termékek mennyiségét. Egy tonna 73% víztartalmú cukornád átlag 769 kg cukortartalmú nádszár mellett 231 kg levél és szárcsúcsi részt tartalmaz. A nádszárból felaprítás után 174 kg szárazanyagot tartalmazó vizes oldatot lehet kipréselni. A visszamaradt növényi maradék száraz súlya 97,4 kg, amelyből elégetve 3,3 kg hamu marad vissza. A víz-oldható frakcióban 122,5 kg fermentálható szénhidrát és 51,9 kg nem fermentálható anyag (19,5 kg nitrogén tartalmú vegyület, 5-6 kg lipid, 13 kg hamu) található. Egy tonna cukorrépából 515 kg szénhidrátot tartalmazó gyökér és 485 takarmányként hasznosítható répafej és levél különíthető el. A répa több mint 81% vizet tartalmaz. A lefejezett gyökérből 84,8 kg szárazanyag tartalmú víz-oldható anyag és 26,2 kg szárazmaradékot tartalmazó, takarmányként hasznosítható extrahált gyökér maradvány képződik. A víz-oldható anyag 66,4 kg fermentálható szénhidrátot és 18,4 kg nem fermentálható maradékot tartalmaz. Ha az adatokat az egy hektáron termelhető ipari növény mennyiségével összevetjük, akkor nyilvánvalóvá válik a cukornád termelésére alkalmas klimatikus viszonyokkal rendelkező államok gazdasági előnye. ( foton /m 2 ) Ez az előny fokozódik, ha a két alapanyagból nyert melléktermék a nádmelasz és a répamelasz fermentálhatóságát összehasonlítjuk. A cukornád melasz nagyobb cukor, biotin és foszfor tartama előnyösen befolyásolja az alkohol termelését. Üzemanyagként való használatát az árviszonyok, a gazdasági meggondolások egyértelműen meghatározzák. A 70-es években bekövetkező olajár-robbanás a gazdasági szakértők figyelmét az etanolra fordította. Ez a környezetvédők igényeit kielégítő energiahordozó vegyipari alapanyagként petrokémiai termékek előállítására is használható. A mezőgazdasági termelésbe fogható területek limitált volta és az előállítási költségek azonban óvatosságra intenek. Nem véletlen, hogy csak azokban az államokban került gyakorlati megvalósításra ez a program, ahol a mezőgazdasági termelés túltermelési válsággal küzd, illetve jelentős mennyiségű nyersolajat importál. Braziliában a proalcool program keretében az 1976-ban 700 millió literes etanol termelést négy év alatt 4 milliárd literre emelték. Ez 450 ezer autó átállítását jelentette etanol üzemre. Az olajimport csökkentése miatt növelték az etanol termelést Dél-Afrikában is. Az USA Gasohol programja évi 400 millió liter etanol üzemanyagként való hasznosítását teszi lehetővé. Az etanol nagyipari előállításának energia igénye: cukornádból 18 MJ/kg etanol, kukoricából 19,4 MJ/kg etanol. Az etanol energia tartalma (26,6 MJ.kg -1 ) alacsonyabb a motorbenzinénél (43,8 MJ kg -1 ), oktánszáma viszont között van. A cukornádból nyert etanol 20%-kal, a kukoricából nyert 50%-kal drágább, mint a cukoripari melléktermékből, a melaszból nyerhető etanol. A melasz mennyiségét viszont a cukorgyárak kapacitása egyértelműen meghatározza. 100
101 A répacukor gyártás technológiai áttekintésekor ipari gyakorlati példaként vizsgáljuk meg a Lehrter Zucker AG (D-3160 Lehrte) szenny-víztisztító rendszerének a működési adatait. Ez az üzem naponta 2000 m 3 szennyvizet termel, amely átlagban 7000 mg/l (BOI) biológiailag oxidálható szerves anyagot tartalmaz. A nagy szervesanyagtartalmú szennyvízet (1) két hőcserélőn (2) átvezetve a metántermelő szakasz optimális hőmérsékletére melegítik fel részben az üzemből származó ipari vizzel (3), illetve a bepárlóból származó (4) kondenzátummal. A vasbetonból készült 3 kamrás összesen több mint 5000 m 3 hasznos térfogatú anaerob reaktor (5) némi denitrifikáció mellett naponta 7500 m 3 gázt termel (80% metán tartalommal), miközben az oxidálható szerves-anyag tartalom 1500 BOI-értékre csökken. A nitrtifikációt is végző aerob szakasz (11) céljára szolgáló, 3500 m 3 térfogatú, levegőztetett reaktorban 1000 m 3 /óra levegő átfúvásával a tisztított szennyvíz oxidálható szervesanyag tartalmát BOI-egységre csökkentik. Az aerob szakasz után egy végső denitrifikáló térben (10) találkozik az anaerob köztesülepítőből származó előtisztított szennyvíz és az utóülepítőből származó (8) eleveniszap, valamint a nyerscukorsűrítőből származó, a hőcserélőn átvezetett még meleg kondenzátum (4). A metánt fermentáló reaktorban a biomassza leülepedését a gáztartályból visszavezetett biogázzal (16) akadályozzák meg. A metánreaktorból egy köztes gázmentesítő kamrán (6) átvezetve kerül a részben tisztított szennyvíz a köztes ülepítőbe (7). Az itt felgyűlő eleven iszap nagy része (8) visszakerül a metántermelő (5) reaktorba. A fölösleg (9) szárításra kerül. Az anaerob fázisban képződő biogáz egy részét a gáztartály (15) gyűjti, a gáz nagyobbik hányada (18) teljes mértékben fedezi a 850 m 3 térfogatú utótisztítóban (13) felhalmozódó, illetve a köztes ülepítőből származó mikroszervezetek (9) szárításához szükséges energiát. A biogáz rendszert az elmaradhatatlan biztonsági fáklya (17) zárja. A tisztított szennyvíz, mely a kiindulási szerves anyag tartalom alig két százalékát tartalmazza, végül az utóülepítőből kerül (14) a környezetbe. D betü jelzi a rendszer azon szakaszait, ahol denitrifikációt igénylő szennyezés még előfordulhat. 101
102 Tanulságos megismerni az 1 tonna alapanyagból előállítható etanol mennyiségét. Cukornád 70 liter Nyárfa 160 liter Répagyökér 95 liter Burgonya 100 liter Melasz 280 liter Kukorica 370 liter Az 1 hektáron termelhető alapanyag és a belőle nyerhető alkohol mennyiségét Átlag Kiemelkedő Átlag Kiemelkedő Cukornád 60 tonna 12,5 tonna 4200 liter 8750 liter Cukorrépa 30 tonna 56 tonna 2850 liter 5320 liter Nyárfa 15 tonna 30 tonna 2700 liter 5400 liter Kukorica 5 tonna 7 tonna 1850 liter 2590 liter A teljesség kedvéért az alapanyag előállítás energia igényét is számításba kell venni Cukornád Brazilia 16 GJ/ha 4,0 MJ/kg etanol Cukorrépa Új-Zéland 16 GJ/ha 5,3 MJ/kg etanol Kukorica USA 21,8 GJ/ha 12,3 MJ/kg etanol Az ipari szesztermelés jelentős hányada ma a gabonafélék szénhidrát tartalmának a hasznosításával készül. A keményítő savas hidrolízise ugyan könnyen kivitelezhető, azonban toxikus melléktermékek (levulinsav, 5-hidroxmetilfurfurol, hangyasav) képződnek. A sav koncentráció optimálásával és a hőmérséklet megválasztásával javítani lehet a helyzeten, mégis az utóbbi időben az enzimes bontást alkalmazzák világszerte. α-amiláz (α-1,4-glukan-4-glukano hidroláz) random bontja a keményítő α-1,4 kötéseit. A mikroorganizmusokban előforduló 60 kd méretű enzim nem bontja az α-1,6 kötést, de az α-1,6 kötés szomszédságában levő α-1,4 kötést sem. Ezért a reakció termékben jelentős mennyiségben fordulnak elő oligomerek. Az Aspergillus oryzae által termelt enzim például keményítőből 4 % glükózt, 56 % maltózt, 28 % maltotriózt és egyéb oligoszacharidot tartalmazó maltóz szirupot készít: β-amiláz (α-1,4-glukán maltohidroláz) a Bacillus genusban előforduló enzim a keményítő redukáló végén kezdve maltóz egységeket hasít le. Nem bontja az 1,6 kötéseket. GLUKOAMILÁZ. Széles spektrumú, a poliszacharid nem redukáló végétől indulva az 1,3, 1,4, és 1,6 kötéseket bontja. Főleg gombákban Aspergillus awamori, Aspergillus niger, Rhizopus fajokban fordul elő. Keményítőből 97 % glükózt, 1,5 % maltozt és valamennyi egyéb oligoszacharidot tartalmazó elegyet készít. Az α-amiláz jelenléte gyorsítja a reakciót, fokozza az eljárás teljesítményét. A fenti enzimek keveréke jó hatásfokkal, de az egyes növényfajokból származó keményítőt eltérő sebességgel glükózig bontja. A gabonakeményítő például könnyebben bontható, mint a burgonyakeményítő. Sok helyen a kukorica teljes feldolgozására törekedve az egyes termékek, például csíraolaj, áztatólé, keményítő, glükózszörp, izomeróz stb. árában jelentkező gazdasági haszon javítja az alkohol termelői árának alakulását. Jelentős javulás várható az alapanyag keményítő, cellulóz, hemicellulóz, fehérje, lipid, nukleinsav és hamutartalmának ismeretében, a növényi maradék fermentálhatóvá alakításától. <Cellulózt bontó mikroszervezetek természetes élőhelyükről különleges dúsító eljárással izolálhatók. Ilyen dúsító táptalaj összetétele lehet 100 ml desztillált vízben oldva 0.1 g NaNO 3 ; 0.1g K 2 HPO 4 ; 0.03g KCl; 0.05g MgSO 4.7H 2 O; 0.05 g élesztőkivonat; 0.05g pepton; 1 g Macherey Nagel 300-as kristályos cellulózpor; 2 g agar-agar A mikroelem igény kiegészítése céljából 0.1 ml baktérium mentesre szűrt nyomelem oldat adagolása (1000 ml desztillált vízben oldva 100mg MnCl 2.4H 2 O; 100mg CoCl 2 ; 10mg CuSO 4 ; 10mg Na 2 MoO 4.2H 2 O; 20mg ZnCl 2 ; 5mg LiCl; 5mg SnCl 2.2H 2 O; 10mg H 3 BO 3 ; 20mg KBr; 8mg NaFe 3+ -trihidrát) szükséges. Ha a tenyésztést C tartományban végezzük, akkor a cellulózt hasznosító törzsek kiemelésére van lehetőség. A 8-10 nap után izolálható telepek közül hatásos törzsek izolálhatók. Ezek 102
103 tisztasága többszöri hígítás után karboximetil-cellulózt tartalmazó táptalajra szélesztve igazolható. A telep körül képződő kráter a CMC bontó aktivitásról tudósít.> Nem hagyható figyelmen kívül, hogy az oligomerből illetve a polimerből felszabaduló monomer az egyensúlyi állapot eléréséig változtatja szerkezetét Az anomerek átalakulása spontán folyamat, amiről a forgatóképesség változása informál [a] 20 D= , α-d-glükopiranóz β-d-glükopiranóz [a] 20 D= A vízben oldott monomer forgatóképessége folyamatosan változik, amíg el nem éri az egyensúlyi állapot ( 1 / 3 α-d-glükóz <> 2 / 3 β-d-glükóz) adta értéket, a ot a dextróz (jobbra) forgató képességét. A hidrolízist követő anomerizáció határozza meg a reakcióképességet. Az enzim az egyik anomert kedveli. 103
104 A CIKLODEXTRIN BIOLÓGIÁJA A piacképes β-ciklodextrin apoláros belső teret kialakító hét glükopiranóz egységet tartalmazó gyürűs vegyület, amelynek vizoldhatósága a külső felületén elhelyezkedő hidroxil csoportok következménye. Ez a termék, különböző apoláros vegyületek, gyógyszerek vizes rendszerbe juttatására alkalmas. Ebből következően, mint metabolizálódó csomagoló anyag hasznosítható. A vegyület a 70 kd méretű ciklodextrinogenáz aktivitását hasznosítva állítható elő keményítőből. Schardinger a XX. század első éveiben észlelte, hogy egy Bacillus macerans törzs, keményítőt tartalmazó táptalajon nem-redukáló kristályos anyagot állít elő. A szakirodalom a felfedező iránti tiszteletből Schardinger dextrinként említette. (Zentr. Bact. Parasiten :172-81; :188-97) E vegyületek komplexképző tulajdonságára Pringsheim hivta fel a figyelmet (Ber. Deut. Chem. Ges : ; : ; : ) A harmincas évek közepén Freudenberg és munkatársai vizsgálták részletesen a fenti komplexképző vegyületek szerkezetét. Kiderült, hogy hat, hét és nyolc glükopiranoz egységet tartalmazó gyürük, amelyet a görög α, β, γ, betüivel jelöltek. (Ann. Chemie : ). A ciklodextrinek képződését katalizáló enzimek vizsgálatával Tilden és Hudson a negyvenes évek elején foglalkoztak (J. Bact :527-44). A hatvanas években DePinto 104
 A hetvenes években japán kutatók Horikoshi és munkatársai ciklodextrin-glükoziltranszferázt termelő alkalofil baktériumokat izoláltak (Agr. Biol. Chem 1971. 35:1783-91).")
105 részletesen vizsgálva a Bacillus macerans enzimkészletét a ciklodextrinogenázon kívül egy intracelluláris ciklodextrináz aktivitását is kimutatta. (Science : ) A hetvenes években japán kutatók Horikoshi és munkatársai ciklodextrin-glükoziltranszferázt termelő alkalofil baktériumokat izoláltak (Agr. Biol. Chem : ). Kitahata és munkatársai pedig részletesen összehasonlították a Bacillus macerans és a Bacillus megaterium enzimkészletét (Agr. Biol. Chem :387-93). A ciklodextrin- (CD) alapú, nem kovalens molekulakomplexek a 70-es évek közepétől indultak el a tudományos érdekességek világából a gyakorlati alkalmazás, a termékfejlesztés irányába. A CD-technológia nemzetközileg elismert úttörője és irányitója Szejtli József volt, így a téma igazi hungarikumnak számít. Ennek bizonyítéka az is, hogy a ciklodextrineket 100 kg-os léptékben, a világpiacot megelőzve, már a 80-as években gyártották a Chinoinban. A CD-molekulák különleges sajátossága, hogy hossztengelyük mentén mindkét végén nyitott üreggel rendelkeznek. Ezekbe az üregekbe alifás molekulák, két-három aromás gyűrűből álló szerkezetek részben vagy egészben beleférnek. A glükózegységek konformációja következtében a CD-molekula amfifil jelleget mutat: a CD-k üregét bélelő glikozidos oxigénatomok és hidrogének miatt a molekula belső felszíne apoláris sajátságú, ám a külső felszínen lévő hidroxilcsoportok kívülről polárissá teszik (J. Szejtli, Cyclodextrin Technology, Kluwer Acad. Publ., 1988.). A ciklodextrinek funkcionális jellemző tulajdonsága a zárványkomplexképzés. A CD-k képesek magukba zárni a víznél kevésbé poláris, geometriailag az adott üregnek megfelelő méretű ún. vendégmolekulákat, melyek lehetnek gyógyszerek, aromák, színezékek, növényvédő szerek, illatanyagok stb. A CD-zárvány-komplexek képződésekor az üregnek megfelelő méretű, apoláris vendégmolekulák és a CD-gazdamolekula között nem kovalens másodlagos kölcsönhatás lép fel. A képződött komplexben a bezárt vendégmolekula időlegesen, a körülményektől függő mértékben, átmenetileg megváltoztatja fizikai és kémiai sajátságait között a fejlesztés iránya a CD-k komplexképző segédanyagfunkcióinak feltárása volt. Ekkor derítették ki, hogy alkalmazhatók oldékonyság-fokozó, hatóanyagstabilizáló diszpergálószerként. A kezdeti 30 év alatt a gyógyszeripari alkalmazások eredménye: 40 piacra vitt sikeres készitmény (K. H. Frömming, J. Szejtli, Cyclodextrins in Pharmacy, Kluwer Acad. Publ., 1994.). Az üres alfa-ciklodextrinek kölcsönhatásba lépnek a zsírsavakkal, mint.zsírsav komplexáló szerek. A bezárt zsírsav nem disszociál, szinte vizoldhatatlanná válik. Egy-egy αcd üreg 5-6 darab CH 2 - csoportot képes fedni. Folyamatos fogyasztása csökkenti a zsírfelszívódást. Az ábrán bemutatott esetben 180 napon keresztül napi 3g alfa- 105
106 ciklodextrin fogyasztása mellett a szérumtriglicerid tartalom 46%-kal, a testttömeg 10%-kal cskkent. Letális toxint termelő mikroorganizmus elleni küzdelem Tankönyvi adatként ismerve a Bacillus anthracis, a lépfene kórokozójának támadási mechanizmusát jól tudott, hogy a protektív antigén heptamerje ül a sejtmembránban pórusként. A lépfene kórokozója egy letális (LF) faktort és egy edema (EF) faktort választ ki, amelyek ezen heptamer póruson jutnak a sejtbe. Újszerű védekezés lehetősége nyílik meg a bakteriális toxinok ellen, mivel egy kémiailag pontosan hangolt CD, a heptakis per-6-[3- aminopropilthio]-β--cd 0.2 nm koncentrációban, az Anthrax porusba illeszkedve gátolja az LF sejtbe jutását. (Karginov et al. PNAS ) A kémiailag módosított ciklodextrin a giráz működését gátló ciprofloxacin hatását potencírozva javítja a gazdaszervezet túlélési lehetőségét. 106
107 A giráz működáse nem csak a sejt osztódása, azaz a DNS állomány megkettőződése szempontjából életfontosságú, de a sejt életműködése mrns képződése szempontjából is nélkülözhetetlen, nem helyettesíthető tevékenység. A kettős szálú DNS a helykihasználás céljából nyugvó állapotban szuperhélix formában, kettősen feltekeredve alkotja a maganyagot. Ebből következően a szuperhelix a DNS replikáció illetve az mrns képződés előtt az információ kezdőpontjánál fellazítandó, majd átmenetilag felhasítandó, végül újra összekapcsolandó az információt hordozó kettős szálú nukleinsav. Ezt a bonyolult folyamatot katalizálja a négy alegységből szerveződött DNS-giráz enzim-komplex, amely az igénynek megfelelően, a szuperfeltekeredett DNS-re kapcsolódva indítja a folyamatot. Mivel az összekapcsoláshoz szükséges energiát a magnézium jelenlétét igénylő ATP hidrolízis szolgáltatja, nem kétséges, hogy a Mg ++ hiánya a folyamatot markánsan gátolja ben észlelték a nalidixsav giráz-enzim működését gátló hatását. Az alapvegyület sokoldalúan fejlesztett változatai közül a ciprofloxacinnak nevezett széles spektrumú magnéziumot kötő, fluórtartalmú kinolonkarbonsav származék bizonyult a leghatásosabbnak. A giráz enzimkomplex működésének gátlása mint látjuk a mikroszervezetek szaporodását akadályozza. Bonyolitja a helyzetet, ha a kórokzó letális toxint is termel. Jó példa erre a Bacillus anthracis okozta kóreset. 107
108 A CIKLODEXTRIN ELŐÁLLÍTÁS TECHNOLÓGIAI KÖRÜLMÉNYEI A Schardinger által vizsgált Bacillus macerans törzs erősen fénytörő ellipszoid endospórát képez. Húslevest és keményítőt tartalmazó táptalajon 38 C -on már a második napon megindul a spórázás, ami a negyedik napra a sejtek teljes lízisével fejeződik be. Alacsonyabb hőmérsékleten, például 28 C -on tenyésztve nem következik be a spórásodás. A tenyészet spórásodásával együttjáró lízis bekövetkeztekor jelenik meg a tenyészet szűrletében a ciklodextrin képződést katalizáló ciklodextrinogenáz, miközben amiláz aktivitást a sejtmentes felüluszóban ekkor sem lehet kimutatni. Optimális tápközeg: 5% zabpehely, 0,4% ammoniumszulfát, 0,2% káliumfoszfát, 1% kálciumkarbonát, 6,8 ph A kezdetben viszkózus állományú tápközeg a tenyészet növekedése közben felhigul. A membránhoz kötött amilolitikus aktivitás hatására - redukáló termék megjelenése nélkül - a keményítőből felszabaduló szénhidrát a sejt belső terében foszforilezve hasznosul. A sűlyesztett fermentáció lefutását bemutató ábrán jól követhető, hogy a növekedés befejeztével jelenik meg a ciklodextrinogenáz aktivitás hasznos terméke, a ciklodextrin, a CD. Az élősejtszám nem csökken, mert a vegetatív sejtek életképes spórává alakulnak, amit igazol a légzési aktivitás csökkenése. A fejlődés korai szakaszában a tápközeg glükóz tartalma katabolikusan represszálja a sejtmembránhoz kötött amilolitikus aktivitást hordozó fehérjék kiszakadását. A növekedési szakaszban a glükóz akadályozza a ciklodextrinogenáz aktivitás megjelenését. Ezzel szemben a spórásodási szakaszban adagolt glükózzal a ciklodextrinogenáz képződését nem lehet represszálni. A spórázó tenyészethez adagolt fehérjeszintézist gátló kloromycetin (klóramfenikol) sem gátolja a ciklodextrinogenáz megjelenését. Ez azt sugallja, hogy az enzimaktivitás megjelenése nem de novo fehérjeszintézis eredménye, hanem a növekedési szakaszban képződő és a sejtmembránhoz kötve működő amilolitikus enzim válik szabaddá a sejtlízis következményeként. A membránból kiszakadó enzim szerkezete általában megváltozik a membránba rögzítő hidrofob szakasz lehasadása miatt. Esetünkben ez a hidrofob, apoláros szakasz nem hasad le, hanem a transzglikozilezés irányába érvényesíti a hidrolitikus hatást. Ezt az aktivitást ciklodextrin-glükozil-transzferáz néven írták le japán kutatók. A keményítő molekula natív állapotban egy balmenetű csavarra emlékeztető csőnek fogható fel, amelynek belső ürege apoláros természetű. Ez abból következik, hogy a 4-es szénatomhoz kapcsolódó ekvatoriális hidroxil és a következő glükóz molekula egyes szénatomjának axiális hidroxilja között alakul ki a glikozidos kötés. Amit a kettes és hármas szénatomokhoz kapcsolódó hidrogénhíd tovább merevít. Ez a szerkezet marad meg a ciklodextrin képződéskor. A gyürű külső oldalán elhelyezkedő hatos szénatom hidroxil csoportja jelenti a hidrofil jelleget a belső apoláros üreggel szemben. A reverzibilisen működő ciklodextrinogenáz apoláros karakterű fehérjelánca a keményítő-csőben elhelyezkedve úgy orientálja az aktív centrumot, hogy a glikozidos kötés bontásakor a helikális szerkezet miatt térközelben levő láncvégi glükóz molekulával kapcsolva az eredetihez hasonló természetű glikozidos kötést alakít ki. A ciklodextrin feldúsulása esetén az enzim apoláros szakaszán nagy gyakorisággal a reakciótermék helyezkedik el, megakadályozva ezzel az enzim kapcsolatát természetes szubsztrátumával, a keményítővel. A biotechnológiai folyamatok irányításához megfelelő analitikai módszerek kidolgozása szűkséges, ami ez esetben nehezen teljesíthető. A Kitahata által használt módszer 108
109 azon a feltételezésen alapul, hogy a ciklodextrinogenázt termelő mikróbák tenyészlevében amiláz aktivitás nem jelenik meg. A reakcióelegybe juttatott keményítő fogyása, a keményítőjód komplex színelnyelésének csökkenése a képződő ciklodextrinnel arányos. Az eredmény értékelését megkönnyíti a Tilden és Hudson módszerként ismert eljárás alkalmazása. Eszerint 0.5 ml enzimoldatot 1 ml 3%-os keményítő oldattal inkubálnak, majd az időnként kivett 3 csepp mintához 1 csepp káliumjodidos-jód oldatot /0,1 M KJ oldatban 0.1 n J 2 / adva mikroszkóppal megfigyelhető a ciklodextrin-jódkomplex hexagonális kristályainak megjelenése. Négy-öt napos, spóratömeggé alakult tenyészet szűrletéből a ciklodextrinogenáz aktivitás jelentős részét hődenaturált /120 C-on 20 percig kezelve/ keményítőszemcsékre adszorbeálva, szelektíven lehet kinyerni. A tenyészlé szűrletéhez 5% denaturált keményítőt adva 4 C-on 20 percig keverve a felülúszóban aktivitás nem mutatható ki. A kiszűrt keményítő szemcsékről 40 C-on desztillált vízzel leoldható az enzim, amely fagyasztva szárítva aktivitásvesztés nélkül tárolható. A 70 kd méretű enzim 5.5 ph-n 40 C-on műküdik maximális aktivitással. xxxxxxxxxxxxx 2005-től napjainkig a ciklodextrinek újabb érdekes funkcionális sajátságait fedezték fel. Felismerték a membrán-aktív CD-származékok alkalmazásának kedvező gyógyászati hatását. A membrán-cd kölcsönhatások molekuláris mechanizmusának felderítése új utat nyitott a multidrug-rezisztencia feloldása felé. Kiderült, hogy egyes ciklodextrinek bejutnak az élő sejtbe, tehát az intracelluláris anyagtranszportban is alkalmazhatók, sőt a vér-agy gáton történő anyagtranszport lehetőségét is igazolták. Oldékonyságfokozó ciklodextrinszármazékok A lipofil anyagok vízoldékonyságának fokozására ez idáig két általánosan engedélyezett CD-származékot használunk [J. Pitha, J. Pharm. Sci. (1985) 74, 9, ]. A 2-hidroxipropil-béta-CD (HPBCD) az Egyesült Államok és az Európai Unió gyógyszerkönyveiben is hivatalos. Az anyag kompozit izomerkeverék, mely így nem kristályosodó, amorf alakban kiváló oldószer. Az anionos szintén izomerkeverék szulfobutiléter-béta-ciklodextrin (SBEBCD) az Egyesült Államokban forgalmazott injektábilis formulációk általános szolubilizálószere. (R. Rajewski et al., J. Pharm. Sci. (1995) 84, 8, ) Kozmetikai, élelmiszeripari és háztartásvegyipari alkalmazások A CD-k legnagyobb mennyiségben történő alkalmazása éppen ezen a területen valósult meg. (2009-ben 7 ezer tonnát használtak fel.) A cél itt is a CD-k molekuláris kapszulaként történő alkalmazása (parfüm-, aromastabilizálás, lipidek, instant italok szolubilizálása) (L. Szente, J. Szejtli, Trends in Food Sci. Technol. (2004) 15, 3 4, ) 109
110 Analitikai alkalmazások A CD-k az analitikai kémia szinte minden területén alkalmazhatók, a legelterjedtebben mégis az enantiomerek elválasztásában használják őket. A királis CDüregek segítségével mind álló, mind mozgó fázisú elválasztások végezhetők. Az eddig ismert ciklodextrin tárgyú közleményből 2800 cikk szól a CD-alapú királis elválasztásokról. Ma a világon legalább húszféle CD-alapú terméket forgalmaznak királis elválasztásokra (Zs. Bikadi et al., Current Drug Disc. Technol. (2007) 4, ) Az elmúlt 35 év meglepetései: Meglepő módon a koleszterinaffinitást mutató CD-k megváltoztatják a lipidöszszetételt az élő sejtek lipiddús membránszakaszain, befolyásolják a membrán fluiditását, a membránfehérjék működését (K. Kamau et al., In vitro Cell. and Dev. Biol. Animal , 7, ) További meglepetés az a felismerés, hogy a HPBCD és a metil-cd-k hatékonyan gátolják a HIV-vírus fertőzőképességét. Ma klinikai fázisban van egy ilyen, HPBCD-t tartalmazó antivirális szer fejlesztése (Z. Liao et al., AIDS Res. Human Retroviruses , 11, ) Váratlan siker lett egy kémiailag pontosan módositott CD önmagában mutatott erős farmakológiai hatása, mely forradalmasította a klinikai aneszteziológiát. A Sugammadex/Bridion felfedezése a tökéletes gazda vendég komplexképzés gyakorlati alkalmazhatóságának szép példája és a CD-kémia legjelentősebb felfedezése (M. Naguib, Anesthesia and Analgesia (2007) 104, 3, ) Szintén az üres CD-nanoüreg érzékeny felismerőképességén alapul egy meglepő DNS-diagnosztikai lehetőség. (J. Clarke et al., Nature Nanotechnology , 4, ) Automatizálható, specifikus, érzékeny DNS-szenzor fejlesztése folyik az Oxfordi Egyetemen pórusképző fehérjébe ültetett CD-származékkal. ÖSSZEGEZVE elmondható, hogy az elmúlt 35 év alatt a ciklodextrin technológia a modern ipar szinte minden területén talált gyakorlati alkalmazást. Eddig 45 gyógyszer, kb. ezer élelmiszeripari- kozmetikai- és háztartásvegyipari termék szolgálja az emberiség egészségét, kényelmét. Az 1975-ben, Szejtli József által kezdeményezett és évtizedekig vezényelt CD technológia napjaink tankönyvi tétele. A munkásságunk kezdetekor néhány grammos szintű, kétes jövőjű anyagokból ma évi ezer tonnát használnak fel. A CD-k tudományos publikációs dinamikája is töretlen, jelenleg CD-tárgyú közlést tartunk nyilván, melyben a magyar kutatók hozzájárulása igen jelentős. Tán emiatt tartják még 35 év után is a magyar CD kutatást és a Szente Lajos vezette CycloLab-ot a világ ciklodextrin technológiai központjának. [Ciklodextrin Kutató Fejlesztő Laboratórium Kft ] 110
111 KÖRNYEZETET SZENNYEZŐ ANYAGOK BIOLÓGIAI HASZNOSÍTÁSA A szerves anyagok anaerob lebontása évmilliárdok óta nélkülözhetetlen részfolyamata a Föld felszín anyagforgalmának. A tudomány ezzel az érdekes és még ma is sok nyitott kérdést tartalmazó problémakörrel a tavak fenekén felszaporodó üledék rothadási folyamatainak vizsgálatakor, a tőzegmocsarak különleges biológiai jelenségeinek vagy a kérődzők bendőjében végbemenő átalakulásoknak a magyarázatakor találkozott. A szemétgödrökben, illetve trágyatárolókban folyó anaerob folyamatok alapjelenségei hosszú ideig senkit sem érdekeltek, annak ellenére, hogy ugyanakkor a jelenség gyakorlati hasznát közösen élvezte az emberiség. Amikor Franciaországban Louis H. Mouras de Vesoul 1860-ban zárt emésztőgödröt épített a birtokán, valószínűleg csak a kellemetlen illatanyagok kiszabadulását akarta megakadályozni. Később az emésztőgödörben végbemenő folyamatot figyelve megállapította, hogy a szilárd növényi hulladékok egy idő után teljesen elfolyósodnak. A képződő gáz (biogáz) hasznosítására azonban nem gondolt. Az irodalmi adatok szerint először 1895-ben Donald Cameron használta a kiáramló gázelegyet fűtőanyagként. Az Exeter-ben épített, nagy emésztőtartályai körül ezzel a gázzal világítottak. A század első felében a szennyvíztisztítást energiaigényes aerob módszerrel próbálták megoldani. Az eljárás valójában a biológiailag eloxidálható szervesanyag-tartalom (AOX) lebontásához szükséges levegő mennyiségének a meghatározására szolgáló analitikai módszer megnövelt méretű változatának tekinthető. A szennyvizet ugyanis a szervesanyag-tartalmának a biológiai lebontásához szükséges oxigén mennyiségével lehet minősíteni. Az aerob bomlást elősegítő levegőztetés azonban nagy felületet és nem kevés energiát igényelt. A levegőztetett szakasz után elhelyezett ülepítő medencék biológiai folyamatait nem vizsgálták. A felhasznált felület csökkentése és a levegőbuborékok oxigén tartalmának jobb kihasználása érdekében 30 méternél magasabb toronyreaktorokat építve igyekeztek gazdaságosabbá tenni az 111
112 eljárást. Megfelelő fúvókák alkalmazásával a 8-10%-os oxigén kihasználás 40-60%-ra emelhető A túlfolyó tölcsérben felgyűlő biomassza egy részét szükség esetén visszavezetik a kezelőtérbe. Az aerob szennyvíz tisztítónak a beruházási költségen túl jelentős az energiaigénye, a közösséget terhelő költségigénye. Tovább fokozható a tisztítás hatásfoka tiszta oxigén bevitelével. Ugrásszerű fejlődést hozott a hetvenes években jelentkező energiakrízis. A nyersolaj árának emelkedése. a szakemberek figyelmét az energetikailag kedvezőbb anaerob módszerekre irányította. Az aerob eljárással történő tisztításkor a szennyvíz eloxidálható széntartalmának 45 %-a széndioxid alakjában távozik; a bevitt anyag fele a szennyvíziszapban dúsul fel (ez később elégethető); 5 % pedig az elfolyó tisztított vízben marad oldva. Anaerob módszerrel viszont ugyanebből a szennyvízből a metabolizálható széntartalom 80 %-a metán és széndioxid formájában gázfázisba vihető, miközben a szennyvíziszapban l0 %, az elfolyó tisztított vízben ugyancsak l0 % széntartalom van jelen. Ez utóbbi érték közel felére csökkenthető, ha az anaerob rendszer után egy kisméretű aerob reaktort működtetünk. Az anaerob reaktorból távozó biogáz összetételét a szennyvíz kémiai összetétele jelentős mértékben befolyásolja: Szénhidrátból 50 % metán és ugyanennyi széndioxid képződik. A fehérje tartalmú szennyvízből képződő széndioxid és metán aránya 3:7, de jelentős az ammóniaképződés. Lipidekből 67 % metántartalom mellett 33 % széndioxid képződik. Természetesen különböző technológiai fogásokkal - hidrogén bevitellel vagy a széndioxid és az ammónia eltávolításával - a gázelegy (biogáz) metántartalma lényegesen növelhető. A gazdasági haszon számottevő, hiszen a hexóz aerob lebontásakor 2803 kj/mol energiát szolgáltat, anaerob körülmények között viszont mindössze l32 kj/mol szabadul fel. A különbség jelentős hányadát a fűtőanyagként használható metán tartalmazza. A szeméttelepeken folyó metánképződésről Jones és Owen már 1934-ben beszámolt. Az energiabőség miatt azonban erre a felfedezésre senki sem figyelt fel. Csupán a 60-as években kezdtek német és amerikai mérnökök e felismerés gyakorlati hasznosításával foglalkozni. Az első gáztermelő üzem a Palos Verdes Landfillnél 1975-ben indult. A szeméttelepről nyert gázt elektromos energia termelésére hasznosították. Ezt követőleg egyre több hasonló üzem épült szerte a világon. Egy1988-ban megjelent statisztikai adat szerint addig 15 országban 146 üzem kezdte meg a működését. Az így nyert gáz általában 55 % metánt tartalmazott. A termelt gáz mennyisége évenként tonna szénnel egyenértékű fűtő teljesítményt jelent. A metánképződést mint általános jelenséget a Föld különböző pontjain végzett vizsgálatok igazolják. A mérhető csekély koncentráció ellenére a Föld évi metántermelése meghaladja a gigatonna mértéket. Ez a metánmennyiség különböző mikroorganizmusok egymást segítő és kiegészítő tevékenységének eredményeként kerül a légkörbe. A szerves anyagból folyó biológiai metánképződés négy, biokémiailag jól megkülönböztethető szakaszra osztható, mivel az egyes szakaszok mikroflórája számára az optimális fizikai-kémiai körülmények jelentős mértékben eltérnek. Az első lépésben nagyméretű szerves molekulák, polimerek, fehérjék, poliszacharidok lebontása folyik. Ezt a feladatot különböző mikroaerofil és fakultatív anaerob szervezetek extracelluláris enzimei végzik nagyon hatásosan. A második lépésben nagyobb molekulák lebontása folyik, miközben C-1 töredékek, szerves savak, alkoholok, széndioxid és hidrogén képződik. Ezt a szakaszt a reakcióelegy savanyodása miatt acidogén fázisnak nevezik. Az itt működő gyors növekedésű mikrobák ezt a savanyodást jól tűrik. 112
113 A harmadik lépésben a fenti vegyületekből a szerves savak és alkoholok továbbalakulása közben főleg ecetsav képződik. Acetogén fázisnek nevezik ezt a szakaszt (Acetil-S-CoA), mert mikrobiális úton ecetsav képződhet közvetlenül széndioxidból és hidrogénből is. A befejező szakasz a metánképződés, amely molekulánként legalább egy ATP képződésével jár. Ebben a fázisban az ecetsav, valamint a C-1 töredékek: metanol, formaldehid, formiát, metilamin szolgáltatják az alapanyagot a metánképződéshez, de a jelenlevő széndioxid és hidrogén is metánná alakul. Innen származik a metanogén fázis elnevezés. A négy szakasz természetes körülmények között térbelileg nem különül el, ezért a metánképződés sohasem éri el a célüzemekben, illetve laboratóriumokban megvalósítható teljesítményt. Itt ugyanis a négy szakaszt térben elkülönítve, két reaktorban folytatják le, ami az optimális rekciókörülmények biztosítását lehetővé teszi. Az ősbaktériumok és az eubaktériumok biokémiai és élettani különbségeit ismerve nem csodálkozhatunk azon, hogy az anaerob szennyvíztisztítás technológiájának kialakításakor a két szakasz térbeli szétválasztására törekedtek. Ezzel a lebontandó szerves anyag összetételétől többé-kevésbbé függetlenné vált a metánképződés. A rendszert tovább fejlesztve a lassan szaporodó metanogének porózus üveggyűrűre rögzítésével olyan sejttömeget lehet a biogázt termelő reaktorban felhalmozni, amely a térfogategységre vonatkoztatott aktivitást a gazdaságosság szintjére emeli. A képződő metán a rendszer működtetéséhez szükséges energiaigényt bőven fedezi. Az első reaktorban egymás tevékenységét segítő mikroaerofil, fakultatív anaerob és szigorúan anaerob baktériumk konzorciuma működik. Ezek széles ph tartományban képesek osztódni, mégpedig a baktériumokra jellemző 30 perces generációs idővel. Zömmel mezofilek, optimális hőmérsékletük 35 o C. A jelenlevő oxigéntűrő szervezetek rövid idő alatt felhasználják a kezdetben jelenlevő oxigén nyomait is. Az első reaktor konzorciuma a nagy molekulák (polimerek) hidrolízisét, szerves savak, alkoholok akár C-1 töredékekig való lebomlását, széndioxid és hidrogén képződését teszi lehetővé. Ezzel előkészítik a fermentációs körülményeket az acetogén és metanogén fázis számára. A hidrogén az anaerob, illetve fakultatív anaerob szervezetekben folyó fermentáció végtermékeként jelenik meg. Fontos szerepet kap a bélbaktériumokban működő hangyasav dehidrogenáz és a piruvát-formiát liáz H 3 C-CO-COOH + CoA-SH > acetil-s-coa + HCOOH HCOOH > H 2 + CO 2 A Clostridium-okban a foszforoklasztikus reakciót segíti a ferredoxin hidrogenáz ferredoxin-hidrogenáz piruvát + szervetlen-foszfát > acetilfoszfát + H 2 + CO 2 red ferredoxin ox A Clostridiumon kívül hidrogént termelnek még többek között az Eubacterium, a Peptococcus és az Aerobacter fajok. Az itt tevékenykedő mikroorganizmusok gyorsan szaporodva hatásosan képesek alkalmazkodni a szubsztrátum minőségében bekövetkező változáshoz. Ezt az alkalmazkodóképességet a technológiai paraméterek szabályozásával segíteni lehet. Nyilvánvaló, hogy a szennyvíz kémiai összetételének állandó volta nagy mértékben képes fokozni a rendszer 113
114 működésének biztonságát és gazdaságosságát. Ezért ezeket a szennyező anyagra optimalizált technológia szerint működő reaktorokat közvetlenül a szennyvizet termelő üzem mellé telepítik, és ezzel a kommunális szennyvíztisztító berendezéseket tehermentesítik. A vázlatrajzon követhetően az Uhde/Schwarting anaerob szennyvíztisztító eljárás egy fakultatív anaerob mikroorganizmusok hatásos savanyító tevékenységét biztosító BO1 jelű fermentorban bontja le a szennyvíz szervesanyagtartalmát. A fermentlé hőmérsékletét 35 Con tartja a beépített WTO1 hőcserélő, amelynek hatásfokát a TIC szabályzó ellenőrzése alatt működő VO2 szelep befolyásolja. Az optimális ph (4,5-5,5) beállítását az LI szabályzó végzi. A szennyvíz betáplálása a PO1 szivattyú feladata. A fermentlé a VO1 szelep állásától függő ütembe kerül át az obligát anaerob ősbaktériumokat foglalkoztató BO2 fermentorba, amelynek optimális hőmérsékletét (55 C) a VO3 szelep által ellenőrzött WTO3 hőcserélő állítja. A második reaktorban az ősbaktériumok az előbbiből származó elegyet ecetsavvá, végül pedig metánná és széndioxiddá alakítják. Az acetogén és a metanogén funkciót végző szigorúan anaerob, termofil baktériumok közössége különleges tulajdonságú, lassú növekedésű fajokból áll össze. A generációs idő harminc órától harminc napig változhat, amit az itt működő (Archaeobacteria) baktériumfajok biológiai tulajdonságai határoznak meg. Legismertebb rendszertani csoportként a Methanobacteriales rend említhető. A rendszer optimalizálásával és egy kisméretű aerob utóreaktor működtetésével, a tisztított szennyvíz az üzemen belül újra felhasználhatóvá válik. A tisztítás hatásfokának látványos bizonyítékaként Japánban aranyhalakat tenyésztenek a tisztított szennyvízben. A két fermentor között szűkség esetén a PO2 szivattyú teremt kapcsolatot, amely a perforált lemezzel szétválasztott BO3 fermentorból nyert sejtmentes lével képes higítani a BO1 fermentor tartalmát. A BO2 fermentorban képződő tisztított szennyvíz a WTO2 hőcserélőn keresztül hagyja el a rendszert, amely a BO1-ből a BO2 fermentorba átfolyó fermentlevet előmelegíti. Mindkét fermentorból a komposztálásra kerülő leülepedő mikroorganizmusokat egy-egy csavarszivattyú távolítja el. A fermentorokban képződő biogáz metántartalma gázmotor működtetésével megtermeli a rendszer működtetéséhez szűkséges elektromos energiát. Mindkét fermentorból a kiülepedő, komposztálandó mikroorganizmusokat egy-egy csavarszivattyú távolítja el. 114
115 A tápanyagokként hasznosuló nitrogéntartalmú vegyületek lebontása - a sejt anyagainak felépítéséhez szükséges szerves nitrogénigényen felül - jelentős ammóniaképződéssel jár. Ezért a biogázban a lebontandó szerves anyag összetételétől függően mindig találunk több-kevesebb ammóniát. Ammóniában dús szennyvíz tisztítása erre a célra kialakított nirtrifikációt és denitrifikációt végző sorba kapcsolt egységek működtetését igényli. A nitrifikáció során aerob körülmények között a Nitrosomonas törzsek az ammóniát nitritig, a Nitrobacter nemzetség pedig tovább nitrátig oxidálja. NH ,5 O 2 2 H + + H 2 O + NO 2 H = 301-től 352 kj NO ,5 O 2 NO 3 H = 65 től 88 kj Anaerob körülmények között a két faj asszociátuma egymást segítve a nitrátlégzés terhére végzi az ammónia oxidációját 5 NH NO 3 4 N 2 +9 H 2 O + 2 H + G = 1483 kj Ammóniában gazdag szennyvíz denirifikálását végzõ eljárás folyamatábrája A denitrifikáció szigorúan anaerob körülményeket igényel. Elektronforrásként a szennyvíz dehidrogénezhető szénforrása szolgál. A folyamatábrán metanol szerepel oxidálható szubsztrátumként NO 3 + 6H e 0,5N H 2 O NO 3 + 0,33 CH 3 OH NO 2 + 0,67H 2 O NO 2 + 0,5 CH 3 OH 0,5 N 2 + 0,5 CO 2 + 0,5 H 2 O + OH némi lugosodással. A denitrifikáló egységben Thiobacillus denitrificans, Micrococcus denitrificans, Pseudomonas, Serratia, Achromobacter nemzetség elektron akceptorként nitrátot használó törzsei végzik. 2 NO 3 > 4 e - > 2 NO 2 > 4 e - > 2 NO > 2 e - > N 2 O > 2 e - > N 2 Az anaerob folyamat közben jelentős energia szabadul fel, amely a műveletet végző szervezetek számára a szénforrás gazdaságos hasznosítását jelenti. 115
116 nitrifikáló egységek vázlatrajzai A nitrifikáló egységekben a levegőztetés hatékonyságát toronyreaktorok kifejlesztésével és megfelelő levegőztető fúvókák alkalmazásával lehet fokozni. A levegőztetett reaktor általában méteres toronyként magasodik a felszín fölé, de gyakran süllyesztik a felszín alá. A Ilyen berendezésben a finom eloszlásban bejuttatott levegő oxigéntartalmának %-a a légbuborék úthosszával arányosan felhasználásra kerül A levegőztetett reaktor után elhelyezett ülepítőből távozik a kezelt szennyvíz. A kiülepedett aktív biomassza egy része a levegőztetett reaktorba visszatáplálva növeli annak a teljesítményét. Az ülepítő hatásfokát mechanikai úton (végtelenített szalaggal) lehet javítani. A tápközeg kéntartalma is redukált formában van jelen a reakciótérben. A sejttömeg kifejlődéséhez szükséges mennyiségen felül a kénhidrogén egyrészt a gázfázisban a biogáztelepek jól ismert illataként jelentkezik, másrészt a jelenlevő fémekkel reagálva fémszulfidok alakjában kicsapódik, ami a szennyvíziszap (zagy) fekete színét okozza. A kénhidrogén toxikus hatása zavarhatja a rendszer működését, amit mésztej adagolásával lehet kivédeni Megoldandó kérdés még, mi történjék a feleslegben képződő sejttömeggel? Az első reaktor eubaktérium sejtjei - inaktiválás után - állattápszerként jól hasznosíthatók. A második reaktor baktériumtömege azonban zömmel az ősbaktériumok csoportját képviseli. A bennük nagy mennyiségben előforduló fitanil-éter származékok miatt inkább talajerő pótlóként célszerű hasznosítani. Ez a vegyületcsoport ugyanis a talaj mikrobaközössége számára jól használható, a magasabb rendűek viszont szerény enzimkészletükkel nem képesek hasznosítani. Az actinomycesek közé sorolt mikroszervezetek pótolhatatlan szerepet játszanak a komposztálás folyamatában a növényi poliszacharidok (cellulóz, hemicellulóz, lignocellulóz) lebontásával. A növényi lignocellulóz felépítettségéből következően nehezen lebontható. A szorosan kötegekbe rendezett cellulóz láncokhoz rövidebb hemicellulóz egységek tapadnak. Ehhez kapcsolódnak extenzinek közvetítésével a savas karakterű pektin láncok. Az egész képződményt a lignin (fenilpropán egységekből álló polimer) szilárdítja az egyes növénycsoportokra jellemző lignocellulózzá. A városi (kommunális) szennyvíz korszerű tisztítására példaként szerepeltethető a FARE segélyből Debrecenben épített tisztítómű, amely a város teljes szennyvíz mennyiségének (50000 m 3 /nap) tisztítását a szigorú EU szabványnak megfelelően végzi. A kialakított rendszer a levegőztetett aerob és fakultatív anaerob tisztítási műveletet, valamint a metanogéneket foglalkoztató szigorúan anaerob reaktorok tevékenységét egységes rendszerben önfenntartó módon működteti. 116


117 A Debrecenben megépített szennyvíztisztító vázlatos rajza és helikopterről készült felvétele. 117
MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben
 Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben Tartalék energiaforrás, membránstruktúra alkotása, mechanikai
Modul cím: MEDICINÁLIS ALAPISMERETEK AZ ÉLŐ SZERVEZETEK KÉMIAI ÉPÍTŐKÖVEI A LIPIDEK 1. kulcsszó cím: A lipidek szerepe az emberi szervezetben Tartalék energiaforrás, membránstruktúra alkotása, mechanikai
A szteroidok. A szteroidok általános előállítása:
 A szteroidok A szteroidok általános előállítása: A szteroidok kémiai vegyületcsalád, de gyártásukban sok biokémiai folyamat van. Előállításuk általában soklépéses folyamat, amelyben a biokémiai és szintetikus
A szteroidok A szteroidok általános előállítása: A szteroidok kémiai vegyületcsalád, de gyártásukban sok biokémiai folyamat van. Előállításuk általában soklépéses folyamat, amelyben a biokémiai és szintetikus
SZTEROIDKONVERZIÓK. BME Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1. Szteroidkonverziók
 SZTEROIDKONVERZIÓK A szterán váz planáris, merev szerkezet, pl. a 3-as és 17- es C-ek távolsága ill. a rajtuk levő szubsztituensek távolsága pontosan meghatározott. A szteránvázas vegyületek bioszintézise
SZTEROIDKONVERZIÓK A szterán váz planáris, merev szerkezet, pl. a 3-as és 17- es C-ek távolsága ill. a rajtuk levő szubsztituensek távolsága pontosan meghatározott. A szteránvázas vegyületek bioszintézise
09. A citromsav ciklus
 09. A citromsav ciklus 1 Alternatív nevek: Citromsav ciklus Citrát kör Trikarbonsav ciklus Szent-Györgyi Albert Krebs ciklus Szent-Györgyi Krebs ciklus Hans Adolf Krebs 2 Áttekintés 1 + 8 lépés 0: piruvát
09. A citromsav ciklus 1 Alternatív nevek: Citromsav ciklus Citrát kör Trikarbonsav ciklus Szent-Györgyi Albert Krebs ciklus Szent-Györgyi Krebs ciklus Hans Adolf Krebs 2 Áttekintés 1 + 8 lépés 0: piruvát
A KOLESZTERIN SZERKEZETE. (koleszterin v. koleszterol)
") 19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
neutrális zsírok, foszfolipidek, szteroidok karotinoidok.
 Lipidek A lipidek/zsírszerű anyagok az élőlényekben előforduló, változatos szerkezetű szerves vegyületek. Közös sajátságuk, hogy apoláris oldószerekben oldódnak. A lipidek csoportjába tartoznak: neutrális
Lipidek A lipidek/zsírszerű anyagok az élőlényekben előforduló, változatos szerkezetű szerves vegyületek. Közös sajátságuk, hogy apoláris oldószerekben oldódnak. A lipidek csoportjába tartoznak: neutrális
Bevezetés a biokémiába fogorvostan hallgatóknak
 Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 4. hét
 Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 4. hét Szerves kémia ismétlése, a szerves kémiai ismeretek gyakorlása a biokémiához Írták: Agócs Attila, Berente Zoltán, Gulyás Gergely, Jakus
Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 4. hét Szerves kémia ismétlése, a szerves kémiai ismeretek gyakorlása a biokémiához Írták: Agócs Attila, Berente Zoltán, Gulyás Gergely, Jakus
R R C X C X R R X + C H R CH CH R H + BH 2 + Eliminációs reakciók
 Eliminációs reakciók Amennyiben egy szénatomhoz távozó csoport kapcsolódik és ugyanazon a szénatomon egy (az ábrákon vel jelölt) bázis által protonként leszakítható hidrogén is található, a nukleofil szubsztitúció
Eliminációs reakciók Amennyiben egy szénatomhoz távozó csoport kapcsolódik és ugyanazon a szénatomon egy (az ábrákon vel jelölt) bázis által protonként leszakítható hidrogén is található, a nukleofil szubsztitúció
A bioenergetika a biokémiai folyamatok során lezajló energiaváltozásokkal foglalkozik.
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
A koleszterin és az epesavak bioszintézise
 A koleszterin és az epesavak bioszintézise Koleszterin A koleszterin a biológia legkitüntetettebb kis molekulája. Tizenhárom Nobel-díjat ítéltek oda azon tudósoknak, aki karrierjük legnagyobb részét a
A koleszterin és az epesavak bioszintézise Koleszterin A koleszterin a biológia legkitüntetettebb kis molekulája. Tizenhárom Nobel-díjat ítéltek oda azon tudósoknak, aki karrierjük legnagyobb részét a
Biotechnológiai alapismeretek tantárgy
 Biotechnológiai alapismeretek tantárgy A biotechnológiai alapismeretek tantárgy magába foglalja a kémia, fizikai kémia és a biológia tantárgyak témaköreit. 1. A) Ismertesse az atomok elektronszerkezetét!
Biotechnológiai alapismeretek tantárgy A biotechnológiai alapismeretek tantárgy magába foglalja a kémia, fizikai kémia és a biológia tantárgyak témaköreit. 1. A) Ismertesse az atomok elektronszerkezetét!
Izoprénvázas vegyületek
 Izoprénvázas vegyületek Izoprénvázas vegyületek (terpének, karotinoidok) Természetes anyagok, amelyek izoprén molekulákból épülnek fel Izoprén C 2 C C C 2 -C 2-2 C C 2 C C 2 C 2 C 5 8 mevalonsav az izoprenoidok
Izoprénvázas vegyületek Izoprénvázas vegyületek (terpének, karotinoidok) Természetes anyagok, amelyek izoprén molekulákból épülnek fel Izoprén C 2 C C C 2 -C 2-2 C C 2 C C 2 C 2 C 5 8 mevalonsav az izoprenoidok
szabad bázis a szerves fázisban oldódik
 1. feladat Oldhatóság 1 2 vízben tel. Na 2 CO 3 oldatban EtOAc/víz elegyben O-védett protonált sóként oldódik a sóból felszabadult a nem oldódó O-védett szabad bázis a felszabadult O-védett szabad bázis
1. feladat Oldhatóság 1 2 vízben tel. Na 2 CO 3 oldatban EtOAc/víz elegyben O-védett protonált sóként oldódik a sóból felszabadult a nem oldódó O-védett szabad bázis a felszabadult O-védett szabad bázis
1.A konverziók általános lefolyása:
 Szteroidkonverziók Az emberi szervezetben a szteránvázas hormonoknak a víz- és sóháztartás szabályozásában (mineralo-kortikoidok), az anyagcserében (pl. glükokortikoidok - glükoneogenezis), a nemi működésekben
Szteroidkonverziók Az emberi szervezetben a szteránvázas hormonoknak a víz- és sóháztartás szabályozásában (mineralo-kortikoidok), az anyagcserében (pl. glükokortikoidok - glükoneogenezis), a nemi működésekben
A cukrok szerkezetkémiája
 A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
A cukrok szerkezetkémiája A cukrokról,szénhidrátokról általánosan o o o Kémiailag a cukrok a szénhidrátok,vagy szacharidok csoportjába tartozó vegyületek. A szacharid arab eredetű szó,jelentése: édes.
Zsírsav szintézis. Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P. 2 i
 Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
2.Enzimes és mikrobiális biokonverziók (alapfolyamatok)
") Lord, I fall upon my knees and pray that all my syntheses may no longer be inferior to those conducted by Bacteria (ZENEA Bio Products and Fine hemicals, Billingham, UK) 2.Enzimes és mikrobiális biokonverziók
Lord, I fall upon my knees and pray that all my syntheses may no longer be inferior to those conducted by Bacteria (ZENEA Bio Products and Fine hemicals, Billingham, UK) 2.Enzimes és mikrobiális biokonverziók
Glikolízis. Csala Miklós
 Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A piruvát-dehidrogenáz komplex. Csala Miklós
 A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
A piruvát-dehidrogenáz komplex Csala Miklós szénhidrátok fehérjék lipidek glikolízis glukóz aminosavak zsírsavak acil-koa szintetáz e - piruvát acil-koa légz. lánc H + H + H + O 2 ATP szint. piruvát H
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL
 TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
TRIPSZIN TISZTÍTÁSA AFFINITÁS KROMATOGRÁFIA SEGÍTSÉGÉVEL Az egyes biomolekulák izolálása kulcsfontosságú a biológiai szerepük tisztázásához. Az affinitás kromatográfia egyszerűsége, reprodukálhatósága
Intra- és intermolekuláris reakciók összehasonlítása
 Intra- és intermolekuláris reakciók összehasonlítása Intr a- és inter molekulár is r eakciok összehasonlítása molekulán belüli reakciók molekulák közötti reakciók 5- és 6-tagú gyűrűk könnyen kialakulnak.
Intra- és intermolekuláris reakciók összehasonlítása Intr a- és inter molekulár is r eakciok összehasonlítása molekulán belüli reakciók molekulák közötti reakciók 5- és 6-tagú gyűrűk könnyen kialakulnak.
Szerves Kémiai Problémamegoldó Verseny
 Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Energiatermelés a sejtekben, katabolizmus. Az energiaközvetítő molekula: ATP
 Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Szerves Kémia II. Dr. Patonay Tamás egyetemi tanár E 405 Tel:
 Szerves Kémia II. TKBE0312 Előfeltétel: TKBE03 1 Szerves kémia I. Előadás: 2 óra/hét Dr. Patonay Tamás egyetemi tanár E 405 Tel: 22464 tpatonay@puma.unideb.hu A 2010/11. tanév tavaszi félévében az előadás
Szerves Kémia II. TKBE0312 Előfeltétel: TKBE03 1 Szerves kémia I. Előadás: 2 óra/hét Dr. Patonay Tamás egyetemi tanár E 405 Tel: 22464 tpatonay@puma.unideb.hu A 2010/11. tanév tavaszi félévében az előadás
EGYSEJTŰ REAKTOROK BIOKATALÍZIS:
 EGYSEJTŰ REAKTOROK BIOKATALÍZIS: A GÉNMÓDOSÍTÁSTÓL AZ IPARI FERMENTÁCIÓIG SZAMECZ BÉLA BIOKATALÍZIS - DEFINÍCIÓ szerves vegyületek átalakítása biológiai rendszer a katalizátor Enzim: élő sejt vagy tisztított
EGYSEJTŰ REAKTOROK BIOKATALÍZIS: A GÉNMÓDOSÍTÁSTÓL AZ IPARI FERMENTÁCIÓIG SZAMECZ BÉLA BIOKATALÍZIS - DEFINÍCIÓ szerves vegyületek átalakítása biológiai rendszer a katalizátor Enzim: élő sejt vagy tisztított
VEBI BIOMÉRÖKI MŰVELETEK KÖVETELMÉNYEK. Pécs Miklós: Vebi Biomérnöki műveletek. 1. előadás: Bevezetés és enzimkinetika
 VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
Helyettesített karbonsavak
 elyettesített karbonsavak 1 elyettesített savak alogénezett savak idroxisavak xosavak Dikarbonsavak Aminosavak (és fehérjék, l. Természetes szerves vegyületek) 2 alogénezett savak R az R halogént tartalmaz
elyettesített karbonsavak 1 elyettesített savak alogénezett savak idroxisavak xosavak Dikarbonsavak Aminosavak (és fehérjék, l. Természetes szerves vegyületek) 2 alogénezett savak R az R halogént tartalmaz
VEBI BIOMÉRÖKI MŰVELETEK
 VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
VEB BOMÉRÖK MŰVELETEK Műszaki menedzser BSc hallgatók számára 3 + 1 + 0 óra, részvizsga Előadó: dr. Pécs Miklós egyetemi docens Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
4. SZERVES SAVAK. Az ecetsav biológiai előállítása SZERVES SAVAK. Ecetsav baktériumok. Az ecetsav baktériumok osztályozása ECETSAV. 04.
 Az ecetsav biológiai előállítása 4. SZERVES SAVAK A bor után legősibb (bio)technológia: a bor megecetesedik borecet keletkezik A folyamat bruttó leírása: C 2 H 5 OH + O 2 CH 3 COOH + H 2 O Az ecetsav baktériumok
Az ecetsav biológiai előállítása 4. SZERVES SAVAK A bor után legősibb (bio)technológia: a bor megecetesedik borecet keletkezik A folyamat bruttó leírása: C 2 H 5 OH + O 2 CH 3 COOH + H 2 O Az ecetsav baktériumok
szerotonin idegi mûködésben szerpet játszó vegyület
 3 2 2 3 2 3 2 3 2 2 3 3 1 amin 1 amin 2 amin 3 amin 2 3 3 2 3 1-aminobután butánamin n-butilamin 2-amino-2-metil-propán 2-metil-2-propánamin tercier-butilamin 1-metilamino-propán -metil-propánamin metil-propilamin
3 2 2 3 2 3 2 3 2 2 3 3 1 amin 1 amin 2 amin 3 amin 2 3 3 2 3 1-aminobután butánamin n-butilamin 2-amino-2-metil-propán 2-metil-2-propánamin tercier-butilamin 1-metilamino-propán -metil-propánamin metil-propilamin
IV. Elektrofil addíció
 IV. Elektrofil addíció Szerves molekulákban a kettős kötés kimutatására ismert analitikai módszer a 2 -os vagy a KMnO 4 -os reakció. 2 2 Mi történik tehát a brómmolekula addíciója során? 2 2 ciklusos bromónium
IV. Elektrofil addíció Szerves molekulákban a kettős kötés kimutatására ismert analitikai módszer a 2 -os vagy a KMnO 4 -os reakció. 2 2 Mi történik tehát a brómmolekula addíciója során? 2 2 ciklusos bromónium
Szerves Kémiai Problémamegoldó Verseny
 Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Szerves Kémiai Problémamegoldó Verseny 2015. április 24. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Egy C 4 H 10 O 3 összegképletű vegyület 0,1776
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
A nitrogén körforgalma. A környezetvédelem alapjai május 3.
 A nitrogén körforgalma A környezetvédelem alapjai 2017. május 3. A biológiai nitrogén körforgalom A nitrogén minden élő szervezet számára nélkülözhetetlen, ún. biogén elem Részt vesz a nukleinsavak, a
A nitrogén körforgalma A környezetvédelem alapjai 2017. május 3. A biológiai nitrogén körforgalom A nitrogén minden élő szervezet számára nélkülözhetetlen, ún. biogén elem Részt vesz a nukleinsavak, a
BIOTERMÉK TECHNOLÓGIA-2
 BIOTERMÉK TECHNOLÓGIA-2 MSc Biomérnök hallgatók számára Előadó: 3 + 0 + 0 óra, 4 kredit szóbeli vizsga Pécs Miklós, Ballagi András Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
BIOTERMÉK TECHNOLÓGIA-2 MSc Biomérnök hallgatók számára Előadó: 3 + 0 + 0 óra, 4 kredit szóbeli vizsga Pécs Miklós, Ballagi András Elérhetőség: F épület, FE lépcsőház földszint 1 (463-) 40-31 pecs@eik.bme.hu
KARBONIL-VEGY. aldehidek. ketonok O C O. muszkon (pézsmaszarvas)
") KABNIL-VEGY VEGYÜLETEK (XVEGYÜLETEK) aldehidek ketonok ' muszkon (pézsmaszarvas) oxocsoport: karbonilcsoport: Elnevezés Aldehidek szénhidrogén neve + al funkciós csoport neve: formil + triviális nevek
KABNIL-VEGY VEGYÜLETEK (XVEGYÜLETEK) aldehidek ketonok ' muszkon (pézsmaszarvas) oxocsoport: karbonilcsoport: Elnevezés Aldehidek szénhidrogén neve + al funkciós csoport neve: formil + triviális nevek
A tananyag felépítése: A BIOLÓGIA ALAPJAI. I. Prokarióták és eukarióták. Az eukarióta sejt. Pécs Miklós: A biológia alapjai
 A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
A BIOLÓGIA ALAPJAI A tananyag felépítése: Környezetmérnök és műszaki menedzser hallgatók számára Előadó: 2 + 0 + 0 óra, félévközi számonkérés 3 ZH: október 3, november 5, december 5 dr. Pécs Miklós egyetemi
Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015
 2015") Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015 A kérdés 1. A sejtről általában, a szervetlen alkotórészeiről, a vízről részletesen. 2. A sejtről általában, a szervetlen alkotórészeiről,
Kollokviumi vizsgakérdések biokémiából humánkineziológia levelező (BSc) 2015 A kérdés 1. A sejtről általában, a szervetlen alkotórészeiről, a vízről részletesen. 2. A sejtről általában, a szervetlen alkotórészeiről,
Xilit fermentáció Candida boidinii segítségével. Kutatási beszámoló
 Xilit fermentáció Candida boidinii segítségével Kutatási beszámoló Dr. Kálmán Gergely A xilit méltán tart számot nagy érdeklődésre sokrétű alkalmazhatóságának köszönhetően kezdve az élelmiszeripartól,
Xilit fermentáció Candida boidinii segítségével Kutatási beszámoló Dr. Kálmán Gergely A xilit méltán tart számot nagy érdeklődésre sokrétű alkalmazhatóságának köszönhetően kezdve az élelmiszeripartól,
Az enzimműködés termodinamikai és szerkezeti alapjai
 2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
2017. 02. 23. Dr. Tretter László, Dr. Kolev Kraszimir Az enzimműködés termodinamikai és szerkezeti alapjai 2017. február 27., március 2. 1 Mit kell(ene) tudni az előadás után: 1. Az enzimműködés termodinamikai
Mikrobák táplálkozása, anyagcseréje
 Mikrobák táplálkozása, anyagcseréje Táplálkozási módok Autotróf Heterotróf Paratróf Fotoautotrófok Kénbaktériumok A fényenergia 1 szakaszban hasznosul Ciklikus és nem ciklikus foszforilálás Ciklikus: csak
Mikrobák táplálkozása, anyagcseréje Táplálkozási módok Autotróf Heterotróf Paratróf Fotoautotrófok Kénbaktériumok A fényenergia 1 szakaszban hasznosul Ciklikus és nem ciklikus foszforilálás Ciklikus: csak
kiindulási vegyületből egy vagy több enzim specifikus a célvegyületnek tekintett piacképes hasznos termékhez juthatunk.
 MSc-BIOTERMÉK TECHNOLÓGIA frissítve:2012.12.02 Az ismertetendő eljárásokban különféle szerves vagy szervetlen kiindulási vegyületből egy vagy több enzim specifikus katalitikus aktivitását hasznosítva a
MSc-BIOTERMÉK TECHNOLÓGIA frissítve:2012.12.02 Az ismertetendő eljárásokban különféle szerves vagy szervetlen kiindulási vegyületből egy vagy több enzim specifikus katalitikus aktivitását hasznosítva a
Kulcsfogalmak és dokumentációkészítés, III. rész
 Kulcsfogalmak és dokumentációkészítés, III. rész UVCB anyagok Gabriele CHRIST http://echa.europa.eu 1 UVCB anyagok Ismeretlen vagy Változó összetétel Komplex reakciótermék vagy Biológiai eredet http://echa.europa.eu
Kulcsfogalmak és dokumentációkészítés, III. rész UVCB anyagok Gabriele CHRIST http://echa.europa.eu 1 UVCB anyagok Ismeretlen vagy Változó összetétel Komplex reakciótermék vagy Biológiai eredet http://echa.europa.eu
Véralvadásgátló hatású pentaszacharidszulfonsav származék szintézise
 Véralvadásgátló hatású pentaszacharidszulfonsav származék szintézise Varga Eszter IV. éves gyógyszerészhallgató DE-GYTK GYÓGYSZERÉSZI KÉMIAI TANSZÉK Témavezető: Dr. Borbás Anikó tanszékvezető, egyetemi
Véralvadásgátló hatású pentaszacharidszulfonsav származék szintézise Varga Eszter IV. éves gyógyszerészhallgató DE-GYTK GYÓGYSZERÉSZI KÉMIAI TANSZÉK Témavezető: Dr. Borbás Anikó tanszékvezető, egyetemi
Vizeletszteroid-profilok meghatározása GC-MS technikával
 Vizeletszteroid-profilok meghatározása GC-MS technikával Bufa Anita PTE ÁOK Bioanalitikai Intézet 2009 Bevezetés Magyarországon orvosi diagnosztikai célokra egyedül a PTE ÁOK Bionalitikai Intézetben alkalmazzuk
Vizeletszteroid-profilok meghatározása GC-MS technikával Bufa Anita PTE ÁOK Bioanalitikai Intézet 2009 Bevezetés Magyarországon orvosi diagnosztikai célokra egyedül a PTE ÁOK Bionalitikai Intézetben alkalmazzuk
1. feladat. Versenyző rajtszáma:
 1. feladat / 4 pont Válassza ki, hogy az 1 és 2 anyagok közül melyik az 1,3,4,6-tetra-O-acetil-α-D-glükózamin hidroklorid! Rajzolja fel a kérdésben szereplő molekula szerkezetét, és értelmezze részletesen
1. feladat / 4 pont Válassza ki, hogy az 1 és 2 anyagok közül melyik az 1,3,4,6-tetra-O-acetil-α-D-glükózamin hidroklorid! Rajzolja fel a kérdésben szereplő molekula szerkezetét, és értelmezze részletesen
A felvétel és a leadás közötti átalakító folyamatok összességét intermedier - köztes anyagcserének nevezzük.
 1 Az anyagcsere Szerk.: Vizkievicz András Általános bevezető Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az élő sejtek nyílt anyagi rendszerek, azaz környezetükkel állandó
1 Az anyagcsere Szerk.: Vizkievicz András Általános bevezető Az élő sejtekben zajló biokémiai folyamatok összességét anyagcserének nevezzük. Az élő sejtek nyílt anyagi rendszerek, azaz környezetükkel állandó
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek
 Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Több oxigéntartalmú funkciós csoportot tartalmazó vegyületek Hidroxikarbonsavak α-hidroxi karbonsavak -Glikolsav (kézkrémek) - Tejsav (tejtermékek, izomláz, fogszuvasodás) - Citromsav (citrusfélékben,
Név: Pontszám: / 3 pont. 1. feladat Adja meg a hiányzó vegyületek szerkezeti képletét!
 Név: Pontszám: / 3 pont 1. feladat Adja meg a hiányzó vegyületek szerkezeti képletét! Név: Pontszám: / 4 pont 2. feladat Az ábrán látható vegyületnek a) hány sztereoizomerje, b) hány enantiomerje van?
Név: Pontszám: / 3 pont 1. feladat Adja meg a hiányzó vegyületek szerkezeti képletét! Név: Pontszám: / 4 pont 2. feladat Az ábrán látható vegyületnek a) hány sztereoizomerje, b) hány enantiomerje van?
Fémorganikus vegyületek
 Fémorganikus vegyületek A fémorganikus vegyületek fém-szén kötést tartalmaznak. Ennek polaritása a fém elektropozitivitásának mértékétől függ: az alkálifém-szén kötések erősen polárosak, jelentős százalékban
Fémorganikus vegyületek A fémorganikus vegyületek fém-szén kötést tartalmaznak. Ennek polaritása a fém elektropozitivitásának mértékétől függ: az alkálifém-szén kötések erősen polárosak, jelentős százalékban
Az élelmiszerek mikrobiális ökológiája. Mohácsiné dr. Farkas Csilla
 Az élelmiszerek mikrobiális ökológiája Mohácsiné dr. Farkas Csilla Az élelmiszerek mikroökológiai tényezői Szennyeződés forrásai és közvetítői A mikroorganizmusok belső tulajdosnágai Belső tényezők (az
Az élelmiszerek mikrobiális ökológiája Mohácsiné dr. Farkas Csilla Az élelmiszerek mikroökológiai tényezői Szennyeződés forrásai és közvetítői A mikroorganizmusok belső tulajdosnágai Belső tényezők (az
A felépítő és lebontó folyamatok. Biológiai alapismeretek
 A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
A felépítő és lebontó folyamatok Biológiai alapismeretek Anyagforgalom: Lebontó Felépítő Lebontó folyamatok csoportosítása: Biológiai oxidáció Erjedés Lebontó folyamatok összehasonlítása Szénhidrátok
Osztályozó vizsgatételek. Kémia - 9. évfolyam - I. félév
 Kémia - 9. évfolyam - I. félév 1. Atom felépítése (elemi részecskék), alaptörvények (elektronszerkezet kiépülésének szabályai). 2. A periódusos rendszer felépítése, periódusok és csoportok jellemzése.
Kémia - 9. évfolyam - I. félév 1. Atom felépítése (elemi részecskék), alaptörvények (elektronszerkezet kiépülésének szabályai). 2. A periódusos rendszer felépítése, periódusok és csoportok jellemzése.
A koenzim Q10 fél évszázados története
 A koenzim Q10 fél évszázados története A koenzim Q10 a sejtek optimális működéséhez nélkülözhetetle A koenzim Q10 (KoQ10) vitaminszerű vegyület. Az ubikinonok k 1 / 8 A Nobel-díjas Dr. Peter Mitchell Kémiailag
A koenzim Q10 fél évszázados története A koenzim Q10 a sejtek optimális működéséhez nélkülözhetetle A koenzim Q10 (KoQ10) vitaminszerű vegyület. Az ubikinonok k 1 / 8 A Nobel-díjas Dr. Peter Mitchell Kémiailag
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak
 BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
Anaerob fermentált szennyvíziszap jellemzése enzimaktivitás-mérésekkel
 Eötvös Loránd Tudományegyetem Természettudományi Kar Környezettudományi Centrum Anaerob fermentált szennyvíziszap jellemzése enzimaktivitás-mérésekkel készítette: Felföldi Edit környezettudomány szakos
Eötvös Loránd Tudományegyetem Természettudományi Kar Környezettudományi Centrum Anaerob fermentált szennyvíziszap jellemzése enzimaktivitás-mérésekkel készítette: Felföldi Edit környezettudomány szakos
Versenyző rajtszáma: 1. feladat
 1. feladat / 5 pont Jelölje meg az alábbi vegyület valamennyi királis szénatomját, és adja meg ezek konfigurációját a Cahn Ingold Prelog (CIP) konvenció szerint! 2. feladat / 6 pont 1887-ben egy orosz
1. feladat / 5 pont Jelölje meg az alábbi vegyület valamennyi királis szénatomját, és adja meg ezek konfigurációját a Cahn Ingold Prelog (CIP) konvenció szerint! 2. feladat / 6 pont 1887-ben egy orosz
ZÁRÓJELENTÉS. OAc. COOMe. N Br
 ZÁRÓJELETÉS A kutatás előzményeként az L-treoninból kiindulva előállított metil-[(2s,3r, R)-3-( acetoxi)etil-1-(3-bróm-4-metoxifenil)-4-oxoazetidin-2-karboxilát] 1a röntgendiffrakciós vizsgálatával bizonyítottuk,
ZÁRÓJELETÉS A kutatás előzményeként az L-treoninból kiindulva előállított metil-[(2s,3r, R)-3-( acetoxi)etil-1-(3-bróm-4-metoxifenil)-4-oxoazetidin-2-karboxilát] 1a röntgendiffrakciós vizsgálatával bizonyítottuk,
Szerves Kémiai Problémamegoldó Verseny
 Szerves Kémiai Problémamegoldó Verseny 2014. április 25. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Adja meg a hiányzó vegyületek szerkezeti képletét!
Szerves Kémiai Problémamegoldó Verseny 2014. április 25. Név: E-mail cím: Egyetem: Szak: Képzési szint: Évfolyam: Pontszám: Név: Pontszám: / 3 pont 1. feladat Adja meg a hiányzó vegyületek szerkezeti képletét!
SZERVES KÉMIAI REAKCIÓEGYENLETEK
 SZERVES KÉMIAI REAKCIÓEGYENLETEK Budapesti Reáltanoda Fontos! Sok reakcióegyenlet több témakörhöz is hozzátartozik. Szögletes zárójel jelzi a reakciót, ami más témakörnél található meg. Alkánok, cikloalkánok
SZERVES KÉMIAI REAKCIÓEGYENLETEK Budapesti Reáltanoda Fontos! Sok reakcióegyenlet több témakörhöz is hozzátartozik. Szögletes zárójel jelzi a reakciót, ami más témakörnél található meg. Alkánok, cikloalkánok
A citoszolikus NADH mitokondriumba jutása
 A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
KARBONSAVAK. A) Nyílt láncú telített monokarbonsavak (zsírsavak) O OH. karboxilcsoport. Példák. pl. metánsav, etánsav, propánsav...
 Nyílt láncú telített monokarbonsavak (zsírsavak) O OH. karboxilcsoport. Példák. pl. metánsav, etánsav, propánsav...") KABNSAVAK karboxilcsoport Példák A) Nyílt láncú telített monokarbonsavak (zsírsavak) "alkánsav" pl. metánsav, etánsav, propánsav... (nem használjuk) omológ sor hangyasav 3 2 2 2 valeriánsav 3 ecetsav 3
KABNSAVAK karboxilcsoport Példák A) Nyílt láncú telített monokarbonsavak (zsírsavak) "alkánsav" pl. metánsav, etánsav, propánsav... (nem használjuk) omológ sor hangyasav 3 2 2 2 valeriánsav 3 ecetsav 3
A biotechnológia alapjai A biotechnológia régen és ma. Pomázi Andrea
 A biotechnológia alapjai A biotechnológia régen és ma Pomázi Andrea A biotechnológia fogalma Alkalmazott biológia A fogalom állandó változásban van A biológia és a biotechnológia közötti különbség a méretekben
A biotechnológia alapjai A biotechnológia régen és ma Pomázi Andrea A biotechnológia fogalma Alkalmazott biológia A fogalom állandó változásban van A biológia és a biotechnológia közötti különbség a méretekben
BIOGÉN ELEMEK MÁSODLAGOS BIOGÉN ELEMEK (> 0,005 %)
") BIOGÉN ELEMEK ELSŐDLEGES BIOGÉN ELEMEK(kb. 95%) ÁLLANDÓ BIOGÉN ELEMEK MAKROELEMEK MÁSODLAGOS BIOGÉN ELEMEK (> 0,005 %) C, H, O, N P, S, Cl, Na, K, Ca, Mg MIKROELEMEK (NYOMELEMEK) (< 0,005%) I, Fe, Cu,
BIOGÉN ELEMEK ELSŐDLEGES BIOGÉN ELEMEK(kb. 95%) ÁLLANDÓ BIOGÉN ELEMEK MAKROELEMEK MÁSODLAGOS BIOGÉN ELEMEK (> 0,005 %) C, H, O, N P, S, Cl, Na, K, Ca, Mg MIKROELEMEK (NYOMELEMEK) (< 0,005%) I, Fe, Cu,
Nitrogéntartalmú szerves vegyületek. 6. előadás
 Nitrogéntartalmú szerves vegyületek 6. előadás Aminok Funkciós csoport: NH 2 (amino csoport) Az ammónia (NH 3 ) származékai Attól függően, hogy hány H-t cserélünk le, kapunk primer, szekundner és tercier
Nitrogéntartalmú szerves vegyületek 6. előadás Aminok Funkciós csoport: NH 2 (amino csoport) Az ammónia (NH 3 ) származékai Attól függően, hogy hány H-t cserélünk le, kapunk primer, szekundner és tercier
H H 2. ábra: A diazometán kötésszerkezete σ-kötések: fekete; π z -kötés: kék, π y -kötés: piros sp-hibrid magányos elektronpár: rózsaszín
 3. DIAZ- ÉS DIAZÓIUMSPRTT TARTALMAZÓ VEGYÜLETEK 3.1. A diazometán A diazometán ( 2 2 ) egy erősen mérgező (rákkeltő), robbanékony gázhalmazállapotú anyag. 1. ábra: A diazometán határszerkezetei A diazometán
3. DIAZ- ÉS DIAZÓIUMSPRTT TARTALMAZÓ VEGYÜLETEK 3.1. A diazometán A diazometán ( 2 2 ) egy erősen mérgező (rákkeltő), robbanékony gázhalmazállapotú anyag. 1. ábra: A diazometán határszerkezetei A diazometán
Szénhidrogének III: Alkinok. 3. előadás
 Szénhidrogének III: Alkinok 3. előadás Általános jellemzők Általános képlet C n H 2n 2 Kevesebb C H kötés van bennük, mint a megfelelő tagszámú alkánokban : telítetlen vegyületek Legalább egy C C kötést
Szénhidrogének III: Alkinok 3. előadás Általános jellemzők Általános képlet C n H 2n 2 Kevesebb C H kötés van bennük, mint a megfelelő tagszámú alkánokban : telítetlen vegyületek Legalább egy C C kötést
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
HALOGÉNEZETT SZÉNHIDROGÉNEK
 ALOGÉNEZETT SZÉNIDOGÉNEK Elnevezés Nyíltláncú, telített általános név: halogénalkán alkilhalogenid l 2 l 2 2 l klórmetán klóretán 1klórpropán l metilklorid etilklorid propilklorid 2klórpropán izopropilklorid
ALOGÉNEZETT SZÉNIDOGÉNEK Elnevezés Nyíltláncú, telített általános név: halogénalkán alkilhalogenid l 2 l 2 2 l klórmetán klóretán 1klórpropán l metilklorid etilklorid propilklorid 2klórpropán izopropilklorid
Zárójelentés a Sonogashira reakció vizsgálata című 48657sz. OTKA Posztdoktori pályázathoz. Novák Zoltán, PhD.
 Zárójelentés a Sonogashira reakció vizsgálata című 48657sz. OTKA Posztdoktori pályázathoz Novák Zoltán, PhD. A Sonogashira reakciót széles körben alkalmazzák szerves szintézisekben acetilénszármazékok
Zárójelentés a Sonogashira reakció vizsgálata című 48657sz. OTKA Posztdoktori pályázathoz Novák Zoltán, PhD. A Sonogashira reakciót széles körben alkalmazzák szerves szintézisekben acetilénszármazékok
Fémorganikus kémia 1
 Fémorganikus kémia 1 A fémorganikus kémia tárgya a szerves fémvegyületek előállítása, szerkezetvizsgálata és kémiai reakcióik tanulmányozása A fémorganikus kémia fejlődése 1760 Cadet bisz(dimetil-arzén(iii))-oxid
Fémorganikus kémia 1 A fémorganikus kémia tárgya a szerves fémvegyületek előállítása, szerkezetvizsgálata és kémiai reakcióik tanulmányozása A fémorganikus kémia fejlődése 1760 Cadet bisz(dimetil-arzén(iii))-oxid
Szerves Kémia II. 2016/17
 Szerves Kémia II. 2016/17 TKBE0301 és TKBE0312 4 kr Előfeltétel: TKBE0301 Szerves kémia I. Előadás: 2 óra/hét Dr. Juhász László egyetemi docens E 409 Tel: 22464 juhasz.laszlo@science.unideb.hu A 2016/17.
Szerves Kémia II. 2016/17 TKBE0301 és TKBE0312 4 kr Előfeltétel: TKBE0301 Szerves kémia I. Előadás: 2 óra/hét Dr. Juhász László egyetemi docens E 409 Tel: 22464 juhasz.laszlo@science.unideb.hu A 2016/17.
ENZIMSZINTŰ SZABÁLYOZÁS
 ENZIMEK 1833.: Sörfőzés kapcsán kezdtek el vele foglalkozni (csírázó árpa vizsgálata) valamilyen anyag katalizátorként működik (Berzelius, 1835.) 1850. körül: ez valamilyen N-tartalmú szervesanyag 1874.:
ENZIMEK 1833.: Sörfőzés kapcsán kezdtek el vele foglalkozni (csírázó árpa vizsgálata) valamilyen anyag katalizátorként működik (Berzelius, 1835.) 1850. körül: ez valamilyen N-tartalmú szervesanyag 1874.:
Heterociklusos vegyületek
 Szerves kémia A gyűrű felépítésében más atom (szénatomon kívül!), ún. HETEROATOM is részt vesz. A gyűrűt alkotó heteroatomként leggyakrabban a nitrogén, oxigén, kén szerepel, (de ismerünk arzént, szilíciumot,
Szerves kémia A gyűrű felépítésében más atom (szénatomon kívül!), ún. HETEROATOM is részt vesz. A gyűrűt alkotó heteroatomként leggyakrabban a nitrogén, oxigén, kén szerepel, (de ismerünk arzént, szilíciumot,
Mire költi a szervezet energiáját?
 Glükóz lebontás Lebontó folyamatok A szénhidrátok és zsírok lebontása során széndioxid és víz keletkezése közben energia keletkezik (a széndioxidot kilélegezzük, a vizet pedig szervezetünkben felhasználjuk).
Glükóz lebontás Lebontó folyamatok A szénhidrátok és zsírok lebontása során széndioxid és víz keletkezése közben energia keletkezik (a széndioxidot kilélegezzük, a vizet pedig szervezetünkben felhasználjuk).
KARBONSAV-SZÁRMAZÉKOK
 KABNSAV-SZÁMAZÉKK Karbonsavszármazékok Karbonsavak H X Karbonsavszármazékok X Halogén Savhalogenid l Alkoxi Észter ' Amino Amid N '' ' Karboxilát Anhidrid Karbonsavhalogenidek Tulajdonságok: - színtelen,
KABNSAV-SZÁMAZÉKK Karbonsavszármazékok Karbonsavak H X Karbonsavszármazékok X Halogén Savhalogenid l Alkoxi Észter ' Amino Amid N '' ' Karboxilát Anhidrid Karbonsavhalogenidek Tulajdonságok: - színtelen,
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
SZTEROIDKONVERZIÓK A SZTEROIDOK BIOSZINTÉZISE. A szteroidok bioszintézise. A szteroidok bioszintézise. A szteroidok bioszintézise
 SZTEROIDKONVERZIÓK A szterán váz planáris, merev szerkezet, pl. a 3-as és 17- es C-ek távolsága ill. a rajtuk levő szubsztituensek távolsága pontosan meghatározott. A szteránvázas vegyületek bioszintézise
SZTEROIDKONVERZIÓK A szterán váz planáris, merev szerkezet, pl. a 3-as és 17- es C-ek távolsága ill. a rajtuk levő szubsztituensek távolsága pontosan meghatározott. A szteránvázas vegyületek bioszintézise
A mikrobaszaporodás alapösszefüggései TÁPOLDATOK, TÁPTALAJOK HOZAMKIFEJEZÉS ÁLTALÁNOSITÁSA. Fermentációs tápoldatok MIKROORGANIZMUSOK TÁPANYAG IGÉNYE
 A mikrobaszaporodás alapösszefüggései BIM SB 2001 TÁPOLDATOK, TÁPTALAJOK HOZAMKIFEJEZÉS ÁLTALÁNOSITÁSA dx ds i = x S = dx dt ds dt µ = µ x S µ = Q x S Y x/s vagy = -Y i i Fermentációs tápoldatok MIKOOGANIZMUSOK
A mikrobaszaporodás alapösszefüggései BIM SB 2001 TÁPOLDATOK, TÁPTALAJOK HOZAMKIFEJEZÉS ÁLTALÁNOSITÁSA dx ds i = x S = dx dt ds dt µ = µ x S µ = Q x S Y x/s vagy = -Y i i Fermentációs tápoldatok MIKOOGANIZMUSOK
Porfin vázas vegyületek (PORFIRINEK)
") Porfin vázas vegyületek (PRFIRIEK) Az élő szervezetekre jellemző kémiai átalakulások két kiemelkedő fontosságú folyamata a porfinvázas vegyületekhez kapcsolódik. Az egyik a zöld növények széndioxid-asszimilációja,
Porfin vázas vegyületek (PRFIRIEK) Az élő szervezetekre jellemző kémiai átalakulások két kiemelkedő fontosságú folyamata a porfinvázas vegyületekhez kapcsolódik. Az egyik a zöld növények széndioxid-asszimilációja,
Aromás vegyületek II. 4. előadás
 Aromás vegyületek II. 4. előadás Szubsztituensek irányító hatása Egy következő elektrofil hova épül be orto, meta, para pozíció CH 3 CH 3 CH 3 CH 3 E E E orto (1,2) meta (1,3) para (1,4) Szubsztituensek
Aromás vegyületek II. 4. előadás Szubsztituensek irányító hatása Egy következő elektrofil hova épül be orto, meta, para pozíció CH 3 CH 3 CH 3 CH 3 E E E orto (1,2) meta (1,3) para (1,4) Szubsztituensek
Talaj mikrobiális biomasszatartalom. meghatározásának néhány lehetősége és a módszerek komparatív áttekintése
 Talaj mikrobiális biomasszatartalom mennyiségi meghatározásának néhány lehetősége és a módszerek komparatív áttekintése A talajminőség és a mikrobiális biomassza kapcsolata A klasszikus talajdefiníciók
Talaj mikrobiális biomasszatartalom mennyiségi meghatározásának néhány lehetősége és a módszerek komparatív áttekintése A talajminőség és a mikrobiális biomassza kapcsolata A klasszikus talajdefiníciók
4. változat. 2. Jelöld meg azt a részecskét, amely megőrzi az anyag összes kémiai tulajdonságait! A molekula; Б atom; В gyök; Г ion.
 4. változat z 1-től 16-ig terjedő feladatokban négy válaszlehetőség van, amelyek közül csak egy helyes. Válaszd ki a helyes választ és jelöld be a válaszlapon! 1. Melyik sor fejezi be helyesen az állítást:
4. változat z 1-től 16-ig terjedő feladatokban négy válaszlehetőség van, amelyek közül csak egy helyes. Válaszd ki a helyes választ és jelöld be a válaszlapon! 1. Melyik sor fejezi be helyesen az állítást:
Összefoglalók Kémia BSc 2012/2013 I. félév
 Összefoglalók Kémia BSc 2012/2013 I. félév Készült: Eötvös Loránd Tudományegyetem Kémiai Intézet Szerves Kémiai Tanszékén 2012.12.17. Összeállította Szilvágyi Gábor PhD hallgató Tartalomjegyzék Orgován
Összefoglalók Kémia BSc 2012/2013 I. félév Készült: Eötvös Loránd Tudományegyetem Kémiai Intézet Szerves Kémiai Tanszékén 2012.12.17. Összeállította Szilvágyi Gábor PhD hallgató Tartalomjegyzék Orgován
SZABADALMI IGÉNYPONTOK. képlettel rendelkezik:
 SZABADALMI IGÉNYPONTOK l. Izolált atorvasztatin epoxi dihidroxi (AED), amely az alábbi képlettel rendelkezik: 13 2. Az l. igénypont szerinti AED, amely az alábbiak közül választott adatokkal jellemezhető:
SZABADALMI IGÉNYPONTOK l. Izolált atorvasztatin epoxi dihidroxi (AED), amely az alábbi képlettel rendelkezik: 13 2. Az l. igénypont szerinti AED, amely az alábbiak közül választott adatokkal jellemezhető:
A mérgek eloszlása a szervezetben. Toxikológia. Szervek méreg megkötő képessége. A mérgek átalakítása a szervezetben - Biotranszformáció
 A mérgek eloszlása a szervezetben Toxikológia V. előadás A mérgek eloszlása a szervezetben Biotranszformáció Akkumuláció A mérgek kiválasztása A mérgek általában azokban a szervekben halmozódnak fel, amelyek
A mérgek eloszlása a szervezetben Toxikológia V. előadás A mérgek eloszlása a szervezetben Biotranszformáció Akkumuláció A mérgek kiválasztása A mérgek általában azokban a szervekben halmozódnak fel, amelyek
(11) Lajstromszám: E (13) T2 EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA
 Lajstromszám: E (13) T2 EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA") !HU00000634T2! (19) HU (11) Lajstromszám: E 006 34 (13) T2 MAGYAR KÖZTÁRSASÁG Magyar Szabadalmi Hivatal EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA (21) Magyar ügyszám: E 0 743972 (22) A bejelentés napja:
!HU00000634T2! (19) HU (11) Lajstromszám: E 006 34 (13) T2 MAGYAR KÖZTÁRSASÁG Magyar Szabadalmi Hivatal EURÓPAI SZABADALOM SZÖVEGÉNEK FORDÍTÁSA (21) Magyar ügyszám: E 0 743972 (22) A bejelentés napja:
Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus. Szülő- és betegtájékoztató
 Bevezetés Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus Szülő- és betegtájékoztató Minden szülő azt várja, hogy gyermeke egészséges lesz. Így azután mélyen megrázza őket, ha megtudják, hogy
Bevezetés Congenitalis adrenalis hyperplasia, 21-hidroxiláz defektus Szülő- és betegtájékoztató Minden szülő azt várja, hogy gyermeke egészséges lesz. Így azután mélyen megrázza őket, ha megtudják, hogy
Szteránvázas vegyületek csoportosítása
 Nemi hormonok Szteránvázas vegyületek csoportosítása a, szterinek b, epesavak c, szívre ható glikozidok d, kortikoidok e, nemi hormonok 1, ösztrogének 2, androgének 3, gesztagének 4, antiösztrogének 5,
Nemi hormonok Szteránvázas vegyületek csoportosítása a, szterinek b, epesavak c, szívre ható glikozidok d, kortikoidok e, nemi hormonok 1, ösztrogének 2, androgének 3, gesztagének 4, antiösztrogének 5,
ALKOHOLOK ÉS SZÁRMAZÉKAIK
 ALKLK ÉS SZÁRMAZÉKAIK Levezetés R R alkohol R R R éter Elnevezés Nyíltláncú, telített alkoholok általános név: alkanol alkil-alkohol 2 2 2 metanol etanol propán-1-ol metil-alkohol etil-alkohol propil-alkohol
ALKLK ÉS SZÁRMAZÉKAIK Levezetés R R alkohol R R R éter Elnevezés Nyíltláncú, telített alkoholok általános név: alkanol alkil-alkohol 2 2 2 metanol etanol propán-1-ol metil-alkohol etil-alkohol propil-alkohol
Badari Andrea Cecília
 Nagy nitrogéntartalmú bio-olajokra jellemző modellvegyületek katalitikus hidrodenitrogénezése Badari Andrea Cecília MTA Természettudományi Kutatóközpont, Anyag- és Környezetkémiai Intézet, Környezetkémiai
Nagy nitrogéntartalmú bio-olajokra jellemző modellvegyületek katalitikus hidrodenitrogénezése Badari Andrea Cecília MTA Természettudományi Kutatóközpont, Anyag- és Környezetkémiai Intézet, Környezetkémiai
Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások
 Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Oktatási Hivatal Országos Középiskolai Tanulmányi Verseny 2010/2011. tanév Kémia I. kategória 2. forduló Megoldások I. FELADATSOR 1. C 6. C 11. E 16. C 2. D 7. B 12. E 17. C 3. B 8. C 13. D 18. C 4. D
Szabó Andrea. Ph.D. értekezés tézisei. Témavezető: Dr. Petneházy Imre Konzulens: Dr. Jászay M. Zsuzsa
 Budapesti Műszaki és Gazdaságtudományi Egyetem Szerves Kémiai Technológia Tanszék α-aminofoszfinsavak és származékaik sztereoszelektív szintézise Szabó Andrea h.d. értekezés tézisei Témavezető: Dr. etneházy
Budapesti Műszaki és Gazdaságtudományi Egyetem Szerves Kémiai Technológia Tanszék α-aminofoszfinsavak és származékaik sztereoszelektív szintézise Szabó Andrea h.d. értekezés tézisei Témavezető: Dr. etneházy
Tartalmi követelmények kémia tantárgyból az érettségin K Ö Z É P S Z I N T
 1. Általános kémia Atomok és a belőlük származtatható ionok Molekulák és összetett ionok Halmazok A kémiai reakciók A kémiai reakciók jelölése Termokémia Reakciókinetika Kémiai egyensúly Reakciótípusok
1. Általános kémia Atomok és a belőlük származtatható ionok Molekulák és összetett ionok Halmazok A kémiai reakciók A kémiai reakciók jelölése Termokémia Reakciókinetika Kémiai egyensúly Reakciótípusok
Részletes tematika: I. Félév: 1. Hét (4 óra): 2. hét (4 óra): 3. hét (4 óra): 4. hét (4 óra):
: 2. hét (4 óra): 3. hét (4 óra): 4. hét (4 óra):") Részletes tematika: I. Félév: 1. Hét (4 óra): Szerves Vegyületek Szerkezete. Kötéselmélet Lewis kötéselmélet; atompálya, molekulapálya; molekulapálya elmélet; átlapolódás, orbitálok hibridizációja; molekulák
Részletes tematika: I. Félév: 1. Hét (4 óra): Szerves Vegyületek Szerkezete. Kötéselmélet Lewis kötéselmélet; atompálya, molekulapálya; molekulapálya elmélet; átlapolódás, orbitálok hibridizációja; molekulák
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek