Glia-neuron interakció aspektusai. Neuron-glia metabolikus együttműködés
|
|
|
- Krisztina Illés
- 5 évvel ezelőtt
- Látták:
Átírás
1 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása Neuron-glia metabolikus együttműködés glutamát-glutamin ciklus asztrocita neuron laktát sönt glutathione prekurzor szintézis Neurovaszkuláris kapcsoltság neurovaszkuláris unit (NvU) lokális véráramlás gliális szabályozása gliális BBB kontroll
2 Glia-neuron interakció aspektusai máshol tárgyalva Szinaptikus működés szabályozása tripartite synapse neuron glia szignalizáció neuron/glia szinapszisok glia neuron szignalizáció, gliotranszmitterek neuronális hálózatok kontrollja szinaptogenezis, szinapszis fenntartása, eliminációja Neurogenezis Neuronal guidence glia mint őssejt glia mint őssejt-niche tagja glia és az axon pathfinding Patofiziológiai aspektusok agyi ödéma akut léziók (trauma, stroke) krónikus neurodegeneratív kórképek neuroimmunológiai reakciók (mikroglia!) remodeling and recovery of neuronal function

3 Glia-neuron interakció Metabolikus együttműködés Agy metabolikusan legaktívabb szervünk: - a teljes testtömegnek csak 2 %-a, de - de a teljes vérellátás 15%-a - a teljes oxigén felhasználás ~20%-a - a teljes glükóz felhasználás ~25%-a az agyban történik. Bonvento 2002 Neuronális aktivitás metabolikus változások hemodinamikus változások kapcsolatának vizsgálata: Glükóz konzumpció: FDG-PET (FluoroDeoxyGlucose-Positron Emission Tomography (FDG-PET) [ 18 F]-2-FDG Oxigén konzumpció: oxyhemoglobin - deoxyhemoglobin (HbO 2 /Hb) szint változás mérése: Blood Oxygen Level Dependent-Magnetic Resonance Imaging (BOLD-MRI) [ 15 O]H 2 O Vérátáramlás mérése: H 2 O-PET.
4 Glia-neuron interakció Metabolikus együttműködés Neuronok energetikai problémái nincs direkt hozzáférésük vérben szállított tápanyagokhoz - vérben fő tápanyag: glükóz és albuminhoz kötött zsírsavak - pl. tüdő, szív mindkettőt hasznosíthatja, de a BBB az albumint kizárja: agyban glükóz monodependencia - glükóztól is 4 membrán (2 endotél, 2 asztro) választja el a neuront (kapilláris asztro végtalp borítottsága 99,7%!, lásd később) hosszú diffúziós útvonalak - pl. ATP a sejttestől 1 mm-re 1 óra alatt diffundálna pl. a denritben, miközben ATP turnover másodperces! (...axonal transport of mitochondria...) kis méretek (dendrittüske, axonátmérő, stb) okozta metabolikus kihívás (metabolic challenge) *(next page) - EPSP (a nagy energiafelhasználók) a felületen történnek: ATP pedig a mitok.-ban ill. a citoplazmában képződik. Minél vékonyabb egy neuronális nyúlvány, annál nagyobb a felület/térfogat arány: saját energiaellátás annál rosszabb. Asztro nyúlványban ezzel szemben egy nagyságrenddel gyorsabb a diffúzió, mint pl. transzporterek vagy metabolikus enzimek turnovere. fluktuáló energiaigény - mikor egy neuron nyugvó állapotból 10 Hz tüzelésbe kezd, ATP felhasználása 7x nő: tehát gyorsan reagáló ATP pótló rendszer kell: másodperceken belül kell új ATP! energiaraktározás hiánya! L. Felipe Barros 2013



5 * diffusionweighted MR spectroscopy (DW-MRS).. is a method to noninvasively quantitate the translational displacement of endogenous metabolites in intact mammalian tissues Nikolay 2001 ADC: apparent diffusion coefficient valós modell!!..we show how long-range morphology of neurons and astrocytes can affect the diffusion of cell-specific metabolites at ultra-long diffusion times and that adequate modeling strategy seems to allow extracting morphological parameters such as astrocytic and neuronal process length and complexity, opening the way to noninvasive histology of the brain Palombo 2016

6 Net gain ~36 ATP wiki
7 L. Felipe Barros 2013
8 ! Glycolysis and its branches acetyl CoA lasd kesobb 1 L. Felipe Barros 2013 PPP Neuronban van glycogen synthase, de inaktív. Ha inaktivitása mutálódik: aberráns neuronális glikogén felhalmozódás: Lafora disease. epilepszia, mental deterioration HEK : hexokinase PFK: phosphofructokinase PK: pyruvate kinase PDH: pyruvate dehydrogenase GP: glycogen phosphorilase G6PD: glucose-6-posphate dehydrogenase GS: glycogen synthase LDH: lactate dehydrogenase PC: pyruvate carboxylase 2 PPP
9 Glia-neuron interakció Metabolikus együttműködés Neuronok evolúciósan sok fajban neuronnak egyedül kellett megoldania e sok problémát - de aztán jött az asztro neuronok energiaigénye magasabb, mint asztroé, de lassabban metabolizálják a glükózt - tenyészetben - szeletben - FACS szerint 3 neuronokban a phosphofructokinase (PFK) működése funkcionálisan gátolt: pentose phosphate kör aktív: redukáló környezet (oxidatív stressz elleni védelem!) Ez nem jelenti azt, hogy ne lenne piruvát szintézis neuronban, csak azt, hogyx PPP itt inkább aktív, mint más sejtekben!) emiatt viszont neuronokban glikolízis nem effektív: bezzeg asztrocitákban: asztro ATP hiány esetén erőteljes glikolízisbe kezd!! Neuron viszont pyruvátot tud csinálni másból (pl. laktátból) is ami megy tovább az energiatermelő folyamatokba (Krebs, ox. foszf.) 4 asztrocitákban viszont pyruvate dehydrogenase foszforiláció által tónusosan gátolt: asztro a laktát termelés és export felé van eltolva 5 asztrociták glikogénből is inkább laktátot állítanak elő!
 FRET sensors are fusion proteins composed of a ligandbinding moiety, the recognition element, and a")
 causes a conformational change that affects the relative distance and/or orientation between the fluorescent proteins, causing an increase or a decrease in FRET")
10 módszer: genetically encoded FRET sensor Laconic in vivo imaging asztrociták és neuronok intravénásan adott L-laktátot akkumulálják az asztrocitáknak szignifikánsabb magasabb a laktát-szintje, mint a neuronoknak ez a laktát gradiens szükséges a carrier-mediálta laktát flux-hoz (asztroból neuronba) FRET sensors are fusion proteins composed of a ligandbinding moiety, the recognition element, and a fluorescent pair with overlapping emission and excitation spectra, typically CFP and YFP. Binding of the test molecule (lactate) causes a conformational change that affects the relative distance and/or orientation between the fluorescent proteins, causing an increase or a decrease in FRET efficiency. The nanosensor described here is based on LldR, a bacterial transcription regulator that consists of two modules, a lactate-binding/regulatory domain and a DNA-binding domain. FRET: fluorescence resonance energy transfer: a mechanism describing energy transfer between two light-sensitive molecules (chromophores).a donor chromophore, initially in its electronic excited state, may transfer energy to an acceptor chromophore through nonradiative dipole dipole coupling. The efficiency of this energy transfer is inversely proportional to the sixth power of the distance between donor and acceptor, making FRET extremely sensitive to small changes in distance.

11 FRET Förster-féle rezonáns energiaátadás
12 Glia-neuron interakció Neuronok Metabolikus együttműködés! Lényeg: neuronok a glikolízist kiszervezik asztrocitákba. Ennek előnyei neuron számára: - neuron antioxidáns státusza (reduktív környezete) érintetlen marad - asztro nyúlvány neuron körül sokkal nagyobb térfogatú: jóval több laktát tud itt keletkezni - gap junction hálózaton is tud glükóz/laktát terjedni (bár laktát GJ transzport függ a membrán-potenciáltól, ami az MCT transzporterekre nem igaz) - de ez jó is, mert neuronális aktivitásnál a laktát a közeli asztroban koncentrálódik! - laktát termelés térben szeparált és független a neuronális működéstől És még fontos: glikolízis oxigéntől független! Glia jól funkcionál ilyen körülmények között, jól túlél a glikolízis során termelt kevesebb ATP-vel. Lehet tulelo asztrocitat (asztrocitomat) izolalni akar a halal beallta utan 24 oraval is, neuron anoxias kornyezetben 3-5 percen belul elpusztul.
13 Luc Pellerin et al. 2007, 2010 Glia-neuron interakció Neuroenergetika (alap) GLUT3: neuronális glükóz transzporter GLUT1: gliális glükóz transzporter Asztro: glikogén raktár! MCT: laktát transzporter TCA cycle (monocarboxylate) Agyi glikogén: (humán): 0,5-1,5g (0,1% of total brain weight) GLUT3 has a higher affinity for glucose than GLUT1 Agyban: glükóz (majdnem) teljes oxidációja (6CO 2 +6H 2 0)!!

14 Glutamáterg aktiváció: korai fázis 1. Glu ürülés szinapszisban AMPA receptor aktiváció, EPSP, Na + belépés 2. depolarizációs hullám, fesz. függő Na + csatornák nyílnak, Na + /K + ATPáz visszaállítja iongradienst - ami sok energiát használ 3. oxidatív foszforiláció aktiválódik, NADH elhasználódik 5. De: AMPAR aktivációra a neuronális glükóz felvétel csökken (*see ref next slide) 4. Citromsavciklus felpörög - piruvát elhasználódik, nő a glükóz és laktát lebontás 6. főleg laktátot használ az idegsejt!

15 * Neuronalis GLUKOZ TRANSZPORTER hiany Neural Deletion of Glucose Transporter Isoform 3 Creates Distinct Postnatal and Adult Neurobehavioral Phenotypes Shin et al. J. Neurosci 2018 globalis GLUT3 hiany korai posztnat: fejlodesben visszamarad, csokkent agymeret, csokkent dendrittuske szam, megnovekedett neuronalis izgathatosag, rovid elettartam lokalis GLUT3 hiany felnottben - limited to the limbic system alone: csokkent felelem terbeli exploraciotol es szocalis helyzetektol, elettartam normalis v.o.: These findings have implications to humans who carry copy number variations pertinent to this gene and have been described to present with neurodevelopmental disorders.
16 Glutamáterg aktiváció: késői fázis GLAST: be: 1 glutamát, 3 Na+, 1 H+ ki: 1 K + 1. A szinaptikus résbe kerülő glutamátot az asztrocita felveszi (GLAST, GLT1 glutamát transzporterek) 2. ez nagy Na+ influxot is jelent, amit Na+/K+ ATPáz állít helyre 3. és aktiválódik a glükóz transzport 4. és a glükóz feldolgozás is a gliában 5. a glikolízis nagy citoplazmás NADH szint növekedést okoz ami segíti piruvát laktát átalakulást, és a laktát ürülést
17 Glutamáterg aktiváció: intenzív és hosszútávú stimuláció - asztro glikogénje lokálisan, saját környezetében hasznosul 1. intenzív glutamate reuptake asztrocitákban, és az ec. glükóz készlet hamar kimerül 2. ilyenkor a gliális glikogén mobilizálódik 3. glikolízis az elsődleges energiatermelési útvonal ilyenkor 4. és a laktát termelődés biztosítja a magas glikolitikus rátát - a raktározott glikogén 20 perccel növelheti meg az axon funkcióképességi idejét

18 Glia-neuron interakció Metabolikus együttműködés Amiről eddig beszéltünk : Asztrocita neuron laktát sönt hipotézis (ANLSH) Astrocyteneuron lactate shuttle Hipotézis!!! arról, hogy a neuronális aktivitás és glükóz felhasználás az asztrociták aerob glikolízisét és a neuronok laktát hasznosítását vonja maga után. Dienel GA, Cruz N Pellerin et al A neurometabolikus community megosztott: csak glükózt vagy laktátot is használ az agy energiaként!!


19 Egy újabb elem ebben a GLÜKÓZ vagy LAKTÁT vitában: Astrocyte-to-Neuron Lactate Shuttling May Occur at Rest, but Not during Neuronal Stimulation Impact
20



21 Glia-neuron interakció Metabolikus együttműködés G protein-coupled receptor GPR81, also known as hydroxycarboxylic acid receptor 1 (HCA1 or HCAR1) = lactate receptor GPR81 has been deorphanized recently: camp signaling
22 Morland et al 2017 Nat Comm mrfp- HCAR1

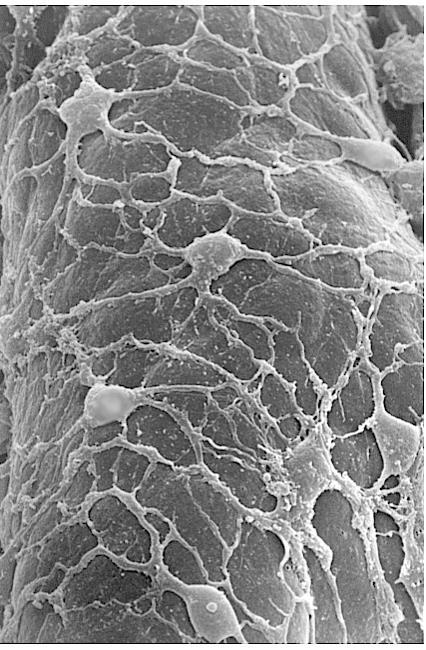
23 Periciták Winkler


24 Pericita
25 Nat Commun May 23;8: Exercise induces cerebral VEGF and angiogenesis via the lactate receptor HCAR1. Morland C 1,2,3, Andersson KA 1,2,3, Haugen ØP 1, Hadzic A 1,2,3, Kleppa L 1,3, Gille A 4, Rinholm JE 1,3, Palibrk V 5, Diget EH 1,6, Kennedy LH 1,3, Stølen T 7, Hennestad E 8, Moldestad O 9, Cai Y 1, Puchades M 3, Offermanns S 10, Vervaeke K 8, Bjørås M 5, Wisløff U 7, Storm- Mathisen J 3, Bergersen LH 1,3,6. Details of HCAR1-expressing cells in pia mater and pial and brain vessels.(a,b) Pial vessel with emerging capillaries labelled for basement membrane collagen IV (CoIV, green), decorated by leptomeningeal cells that contain mrfp-hcar1 (red) and co-localize fibroblast marker vimentin (magenta, b); confocal images from top frozen section tangential to the brain surface. Scale bar, 70 μm. (c) Pial cells (arrow) in confocal image of parasagittal frozen section perpendicular to the cerebral cortex of mrfp-hcar1 reporter mouse. Scale bar, 20 μm. (d) In the extension of pia in fissura hippocampi (*), which separates hippocampus stratum lacunosummoleculare (LM) from the molecular layer of area dentata (Mo), mrfp-hcar1 is in select blood vessels (arrowheads), including ones (small arrowhead) penetrating into the Mo. Scale bar, 50μm. (e h) Blood vessel penetrating into the cerebral cortex with a sheath of mrfp-hcar1-containing perivascular cells (long arrows), CD31 in endothelial cells (short arrows, green; f g) and DAPI-stained cell nuclei (blue; f); single optical section (1.38 μm). The picture is compatible with a slight mrfp-hcar1 signal also in endothelial cells. Subpial/paravascular/perivascular space (Pvs). Scale bar, 20 μm (e g). (h) Magnification of part of f. Scale bar, 10μm. (i) mrfp- HCAR1 in the hippocampus, magnified frame showing details of mrfp-hcar1-carrying cells (arrows, red), which extend processes (small arrows) around blood vessels (CoIV-labelled, green) that penetrate into the hippocampus through the extension of pia in the fissura hippocampi (indicated by white asterisks, brain surface to the left outside the picture). Hil, hilus of area dentata; Gr, granule layer of area dentata; Mi, Mm, Mo, inner, middle and outer zones of the molecular layer of area dentata; LM, Rad and Pyr, lacunosummoleculare, radiatum and pyramidal layers of hippocampus CA1. Dashes mark borders between Hil and Gr, and between LM and Rad. Scale bar, 50 μm. (j l) Small vessel in the cerebral neocortex, surrounded by cell co-expressing mrfp-hcar1 (j, red; l, yellow) and the pericyte-associated protein PDGFRβ (k, green; l, yellow). Staining of nuclei (arrow and arrowhead, DAPI, white) reveals that an endothelial cell (arrowhead) is located between the lumen and the mrfp-hcar1/pdgfrβ-co-expressing cell. Scale bar, 15 μm. mrfp-hcar1

26 Nat Commun May 23;8: Exercise induces cerebral VEGF and angiogenesis via the lactate receptor HCAR1. Morland C 1,2,3, Andersson KA 1,2,3, Haugen ØP 1, Hadzic A 1,2,3, Kleppa L 1,3, Gille A 4, Rinholm JE 1,3, Palibrk V 5, Diget EH 1,6, Kennedy LH 1,3, Stølen T 7, Hennestad E 8, Moldestad O 9, Cai Y 1, Puchades M 3, Offermanns S 10, Vervaeke K 8, Bjørås M 5, Wisløff U 7, Storm- Mathisen J 3, Bergersen LH 1,3,6. Organization of cells that carry HCAR1 and of the angiogenic action of lactate. Blood-borne lactate from exercising muscle penetrates the blood vessel wall (yellow) through monocarboxylate transporters located in the vascular endothelium (which represents the blood brain barrier). Extravascular lactate (from blood or generated in the brain parenchyma upon neural activation) is freely diffusible in the perivascular/subpial space, thereby bathing the leptomeningeal fibroblast-like cells carrying HCAR1 (red). Magnified inset indicates possible, yet unidentified (?), pathways leading from activation of HCAR1 in the cells in pia and perivascular sheaths to increased VEGFA and subsequent enhanced angiogenesis. The perivascular sheath extends as separate HCAR1-expressing pericyte-like cells at intracerebral microvessels, which may also contribute in the angiogenic process. Although apparently devoid of mrfp- HCAR1, other cells may possibly express low levels of HCAR1. HCAR1 may stimulate VEGFA in the same cells, or in other cells, through mediators. In addition to its angiogenic action, VEGFA has neurotrophic effects 11. Importantly, all blood to the brain parenchyma has to pass in close proximity to the perivascular sheath of HCAR1- carrying cells and therefore can convey products released from these cells upon activation of the receptor; blood to the hippocampus passes through vessels (such as the ones shown in Fig. 3i) entering in the hippocampal fissure, an extension of the pia mater that penetrates deep into the centre of the hippocampal formation.
27 Physical exercise can improve brain function and delay neurodegeneration; however, the initial signal from muscle to brain is unknown. Physical exercise Lactate production HCAR1 lactate receptor on leptomeningeal cells + pericytes cerebral vascular endothelial growth factor A and angiogenesis In contrast, skeletal muscle shows no vascular HCAR1 expression and no HCAR1-dependent change in vascularization induced by exercise or lactate. Morland et al 2017
28 at King Abdullah Univ - patkány HC: tanulás során glikogén eredetű laktát szint nő - gliális laktát transzporterek (MCT4 vagy MCT1) kilövése amnéziát okoz, amit L- laktát visszaállít (de a glükóz nem!!) - neuronális laktát transzporter hiánya szintén amnéziához vezet, amit sem a laktát, sem glükóz nem állít helyre
 sleep epochs are colored blue, and rapid eye movement (REM) sleep epochs are indicated in green.")
 glutamate")
29 Multiple sleep/wake cycles recorded using simultaneous electroencephalographic and (A) lactate/glutamate biosensors or (B) lactate/glucose biosensors plotted during the lights-on period. Epochs scored as wake are noted in red, Non-Rapid Eye Movement (NREM) sleep epochs are colored blue, and rapid eye movement (REM) sleep epochs are indicated in green. Concentration change for each analyte is indicated on the y-axis. The lower graphs (C, D) correspond to time periods on the upper graphs indicted by the solid box. In all expanded graphs, lactate concentration change is plotted in a manner similar to that of the large-scale graphs with colors indicating sleep/wake state and the secondary analyte (C) glutamate or (D) glucose plotted as on overlay in fuchsia or orange, respectively. From Naylor et al. (2012).

30 nice picture
31 Glia-neuron interakció Metabolikus együttműködés Asztrociták számos olyan metabolikus kulcsenzimet expresszálnak, melyek a neuronokban nem fordulnak elő (vagy nem jellemzőek/nem funkcionálisak):! glutamát-glutamin ciklus: GS glutamin synth(et)ase glikogén mobilizáció: glikogén foszforiláz 6 lásd korábbi dia glikolízisről oxálacetát anaplerotikus szintézise: pyruvát karboxiláz (azokat a reakciókat, amelyek a citromsavciklus köztitermékeit hozzák létre, anaplerotikus reakcióknak nevezzük) (citrát kör feltöltése ) lásd korábban lásd korábban taurine szintézis: cysteine sulfinate decarboxylase glycin metabolizmus: glycine cleavage system (glycine decarboxylase complex or GCS, több enzim), nagy glycin koncentrációk aktiválják neuronális glutathione szintézis prekurzorainak előállítása 2 lásd korábbi dia glikolízisről lásd nemsoká
32 neurotransmission unavoidably triggers increased mitochondrial reactive oxygen species in neurons Bolanos JP J Neurochem (Ca++ )

33 Glia-neuron interakció Metabolikus együttműködés ROS (reactive oxygen species) reaktív oxidatív szabadgyökök Hidroxilgyök OH Szuperoxidgyök O 2 Nitrogénoxid-gyök NO Lipidperoxil-gyök LOO Pl. stroke esetén: ROS káros hatásai: Hidrogén-hiperoxid H 2 O 2 Szuperoxid anion O 2 - Hipoklórossav HOCl Ózon O 3 Nitrogén monoxid NO Peroxynitrit ONOOoxidatív stressz - DNS károsítás - lipid peroxidáció - fehérje károsítás - enzimek oxidatív inaktiválása

34 Glia-neuron interakció Metabolikus együttműködés Neuronális glutathione szintézis prekurzorainak előállítása asztrocitákban Glutathione (g-glutamylcysteinylglycine, GSH): szulfhidril (-SH) antioxidáns, antitoxin, enzim kofaktor. GSH nagyon fontos a ROS detoxifikációban!! a GSH egy tripeptide C 10 H 17 N 3 O 6 S, mely egy-egy aminosav gyököt tartalmaz az alábbiakból: glutamát, cisztein, glicin A GSH szulfhidril csoportja alkalmas a ROS nem enzimatikus redukálására. A GSH keletkező, oxidált formája két molekulából áll, melyet diszulfid híd tart össze (GSSG). Glutathione hiány esetén: súlyos idegr. károsodások, neuronvesztés!!!!
35 Glycolysis and its branches acetyl CoA 1 L. Felipe Barros 2013 PPP Neuronban van glycogen synthase, de inaktív. Ha inaktivitása mutálódik: aberráns neuronális glikogén felhalmozódás: Lafora disease. epilepszia, mental deterioration HEK : hexokinase PFK: phosphofructokinase PK: pyruvate kinase PDH: pyruvate dehydrogenase GP: glycogen phosphorilase G6PD: glucose-6-posphate dehydrogenase GS: glycogen synthase LDH: lactate dehydrogenase PC: pyruvate carboxylase 2 PPP

36 relaxing picture
37 Glia-neuron interakció glutamát, cisztein, glicin Glutathione Metabolikus együttműködés xct Glycine Asztrociták glycint és glutamátot az EC térből akkumulálnak. A cystine-t cystine/glutamate exchanger (xct) révén veszik fel. Az ic. cystine cysteine né redukálódik. Ez a glutamáttal γ-glutamylcysteine t alkot (γ-glucys). Ez a dipeptid gylicin inkorporációval tripeptiddé, glutathionná (GSH) alakul. Hertz, Zielke 2004
 alakítja át: cysteinylglycine (CysGly) és glutamát keletkezik.")
38 Hertz, Zielke 2004 Glia-neuron interakció Metabolikus együttműködés Glutathione Neuron nem képesek a cystine cystein redukcióra, így gliális glutathione-tól függnek! xct A GSH-t a γ-glutamyl transpeptidase (γ-gt) alakítja át: cysteinylglycine (CysGly) és glutamát keletkezik. A cysteinylglycine-t a neuronális felszínen egy másik ektopeptidáz, az aminopeptidase N (ApN) hasítja: glycine és cysteine keletkezik. Mindkettőt a neuron akkumulálja. A neuronokban újra γ-glutamylcysteine keletkezik cysteine és glutamate felhasználásával (glutamát glutaminból vagy újrafelvételből származik). Végül glycine hozzáadásával elkészül a neuronális glutathione.
39 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása Neuron-glia metabolikus együttműködés glutamát-glutamin ciklus asztrocita neuron laktát sönt glutathione prekurzor szintézis Neurovaszkuláris kapcsoltság neurovaszkuláris unit (NvU) lokális véráramlás gliális szabályozása gliális BBB kontroll
40 Glia-neuron interakció aspektusai máshol tárgyalva Szinaptikus működés szabályozása tripartite synapse neuron glia szignalizáció neuron/glia szinapszisok glia neuron szignalizáció, gliotranszmitterek neuronális hálózatok kontrollja szinaptogenezis, szinapszis fenntartása, eliminációja Neurogenezis Neuronal guidence glia mint őssejt glia mint őssejt-niche tagja glia és az axon pathfinding Patofiziológiai aspektusok agyi ödéma akut léziók (trauma, stroke) krónikus neurodegeneratív kórképek neuroimmunológiai reakciók (mikroglia!) remodeling and recovery of neuronal function


41 Tripartite szinapszis In 1994, Vladimir Parpura and colleagues conducted a set of experiments making an intriguing discovery: cultured astrocytes release glutamate, which leads to Ca2+ elevation of nearby neurons. Later studies demonstrated that this process can be observed in acute brain slices and in vivo. Astrocytes were later shown to release a number of chemical transmitters, including ATP, D-serin, TNF-alpha, and ANP in a process that has recently been termed gliotransmission. Based on the early studies of gliotransmission, the concept of the tripartite synapse was proposed highlighting the role of the astrocyte as a third active element in information processing at the synapse. Although many aspects of this astrocytes-to-neuron communication are still to be elucidated the introduction of molecular genetic tools is shedding light on the neuromodulatory roles of astrocytes on brain function at the level of synapses, circuits, and behavior. Halassa 2007, 2010
42 Glia-neuron interakció Neuron glia Glia szignalizáció Glutamaterg szinapszis. 1. neuronok stimulációja Ca ++ szignalizációt indukál asztrocitákban 2. asztrociták érzékelik a neuronális aktivitás intenzitását: ha a szinaptikus aktivitás nő nő a glia Ca ++ oszcilláció frekvenciája is - Asztro is rendelkezik memóriával, mint neuron: intenzív neuronális aktivitás esetén neuronális EPSP amplitúdója nő (LTP!) ÉS a gliális Ca ++ válasz frekvenciája is nő! Verkhratsky Butt 2007
43 Glia-neuron interakció Neuron Glia szignalizáció Példa 1. Hippocampus Figure 1.5. Hippocampal pathways and their stimulation Signals from the entorhinal cortex (EC) enter the dentate gyrus (DG) via the perforant path (PP). From the DG granule cells information travels to the CA3 pyramidal neurons via the mossy fibers. From the CA3 neurons the signal leaves via the Schaffer collaterals and joins with the commissural fibers (Comm.) from the contralateral CA3 making connections with CA1 pyramidal neurons. Signals leaving CA1 then travel to neurons within the subiculum. A bipolar stimulating electrode was placed on the Schaffer collateral and commissural (comm.) fibers. Recording electrodes placed in the dendritic layer and/or the pyramidal layer of CA1 will record an Excitatory Postsynaptic Potential (EPSP) or a population spike (PS) following stimulation, respectively. EPSP represents the response at the CA3- CA1 synapse and the PS represents the number of pyramidal cells firing and the contribution of the EPSP at that location. The top portion of the figure demonstrates the four layers that the CA1 pyramidal neuron lies within (S. denotes Stratum). The small neuron with a letter I represents an inhibitory interneuron. The pathway diagramed in the top portion of the figure corresponds to the recurrent inhibitory loop in area CA1.
44 Glia-neuron interakció Neuron Glia szignalizáció Példa 1. Hippocampus Schaffer-kollaterálisok stimulációja [Ca ++ ] ic növekedést vált ki CA1 stratum radiatum asztrocitákban - kis szinaptikus aktivitás csak mglur-t aktivál asztron - nagy szinaptikus aktivitás mglur-t és AMPAR-t is aktivál - repetitív Ca ++ szignálokat csak a nagy-frekvenciájú stimuláció vált - az asztro nyúlványokban a Ca ++ válaszok aszinkron természetűek: az asztrociták különböző, relatíve izolált kompertmentumai egyedi vagy néhány szinapszis aktivitását követik Porter, Porter, McCarthy McCarthy
 Ca ++")

45 Glia-neuron interakció Neuron Glia szignalizáció a: stimuláció előtt [Ca ++ ] ic b,c: stimuláció után (200 ma, 50 Hz, 2 sec) Ca ++ szenzítív fluoreszcens festékkel töltött asztrociták d: stimuláció után 28 sec e: GFAP immunfestés azonosítás Porter, McCarthy 1996 Porter, McCarthy 1996


46 Glia-neuron interakció Példa 2. Cerebellum Hoogland 2010 Tanaka BGC: Bergmann glial cells PC: Purkinje cell ml: molecular layer AC: cerebellar astrocytes (velate protoplasmic astrocytes) gcl: granule cell layer wm: white matter

47 Glia-neuron interakció Példa 2. Cerebellum -parallel rost/purkinje neuron szinapszisokat Bergmann glia körbeveszi Neuron Glia szignalizáció -parallel rost stimulációra Bergmann gliában erősen lokalizált Ca ++ válaszok jelennek meg szignalizációs mikrodomének A: kísérleti felállás: parallel rost stimuálció Bergmann glia Ca ++ imaging B: Bergmann glia Oregon green 488 BAPTA-1 Ca ++ -szenzitív fluorfestékkel feltöltve. Ca ++ mérés több régióban C: Bergmann glia mikrodomének sematikus rajza. Több, közel futó parallel rost egy mikrodomént aktiválhat. Verkhratsky Butt 2007 függelék Bergmann glia főbb nyúlványa
48 Glia-neuron interakció Példa 3. Hippocampus Neuronális acetilkolin release asztrogliális Ca ++ tranzienseket generál Szeptumból hippocampusba érkező kolinerg afferensek repetitív stimulációja [Ca2] ic oszcillációkat vált ki a stratum oriensben található asztrocitákon, muszkarinikus AchRec-on keresztül. Tehát asztrociták távoli axonális inputra is reagálhatnak. Példa 4. Hippocampus GABAerg interneuronok aktiválják asztrocitákat Neuron Glia szignalizáció Araque 2002 CA1 piramissejteken szinaptizáló GABA-ergic interneuronok repetitív tüzelése [Ca2] ic szint növekedéshez vezet a környező asztrociták egy csoportjában (paired patch-clamp recordings), GABAB receptoron át, extracell Ca++-tól függő módon. Neuronális gátlás is Ca++ növekedést okoz. Példa 5. Hippocampus Neurotrophin kiváltotta Ca ++ szignalizáció asztrocitákban Kang 1998 BDNF (amely neuronokból aktivitás-függő módon ürülhet) a TrkB-T1 receptorokon keresztül késleltetett [Ca2] ic növekedést váltott ki CA1 asztrocitákon. Gliális Ca++ válasz késleltetése, időzítése! Rose 2003 Reviewed in Volterra, Steinhauser 2004
49 Glia-neuron interakció Neuron Glia szignalizáció Neuron/glia szinapszisok szinapszis-szerű kapcsolatok hipofízis pars intermedia pituicitái és a nucleus arcuatusból érkező axonok között norepinephrine terminálisok synaptoid kontaktusokkal végződnek szepto-hippocampalis asztrocitákon Christian Steinhauser szinapszisok végződnek GluR sejteken szinapszisok NG2+ sejteken!! lásd korábban, 04 előadás.,ng2 sejt

50 Neuron/glia szinapszisok Jabs 2005, Bergles 2010 Legalább kétféle preszinaptikus neuron (glutamaterg CA3 piramissejt és GABAerg interneuronok) innerválják az NG2 sejteket. Ezek posztszinaptikus áramai jól megkülönböztethetőek. Neuron-NG2 szinaptikus kapcsolat potenciális funkciója: NG2+ sejt myelináló oligodendrogliává való differenciálódásának szabályozása neuronális aktivitás függvényében! erre már kezd gyűlni némi bizonyíték:
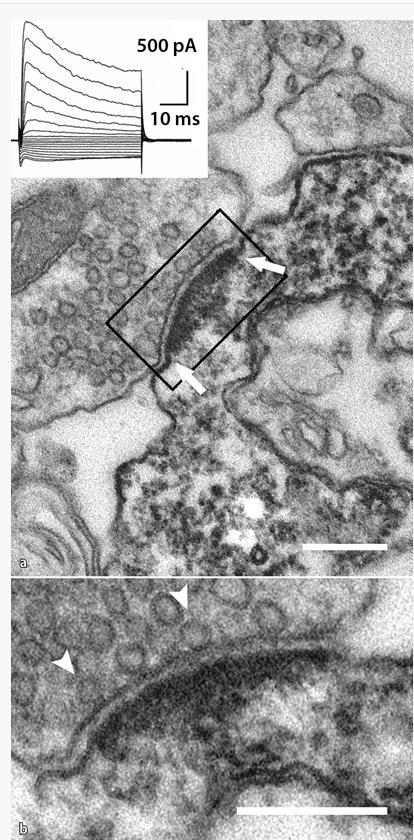

51 Neuron Glia szignalizáció Neuron/glia szinapszisok
52 Neuron Glia szignalizáció Neuron/glia szinapszisok

53
54 Glia-neuron interakció aspektusai Szinaptikus működés szabályozása tripartite synapse neuron glia szignalizáció neuron/glia szinapszisok neurotranszmitter konc szabályozása signaling by exosomes gliotranszmisszió, példák glia neuron szignalizáció, gliotranszmitterek neuronális hálózatok kontrollja szinaptogenezis, szinapszis fenntartása, eliminációja
55 Glia-neuron kapcsolat interakció Neurotranszmitter gyors eltávolítása, szinaptikus hatékonyság megőrzése: Glia Neuron szignalizáció 1. G N : neurotranszmitter konc. szab. A. Uptake Részleteket lásd korábban: [Glu] ec koncentráció szabályozása [GABA] ec koncentráció szabályozása illetve egyéb, szinapszis-típustól (pl. monoaminerg, etc.) függő uptake - amihez megfelelő gliális transzporterek társulnak B. Kötő-fehérje release: Központi kolinerg szinapszisokban: periszinaptikus asztrociták acetylcholine-kötő fehérjét (AChBP) szintetizálnak és ürítenek a szinaptikus résbe. E fehérje gliális szekréciója intenzív kolinerg stimulációra nő.

56 Glia-neuron interakció 1. transzendocitózis, exoszómák Glia neuron szignalizáció

57 Volterra, Meldolesi Nature Rev Neurosci, 2005 Glia-neuron interakció Glia Neuron szignalizáció 2. Gliotranszmisszió
 akik megkérdőjelezik a gliotranszmisszió tényét/jelentőségét de valójában erre")
58 Glia-neuron interakció Glia Neuron szignalizáció 2. Gliotranszmisszió vannak (voltak) akik megkérdőjelezik a gliotranszmisszió tényét/jelentőségét de valójában erre számtalan bizonyíték van, ahogy korábban részletesen megbeszéltük
59 b) NMDAR c) AMPAR b) AMPAR/mGluR agonist L- quisqualate on astroctytes c) TFLLR agonist of the PAR- 1(protease activated) metabotropic receptors on astrocytes (ligand: pl thrombin) b) quantal events of vesicular glutamate release from the astrocyte c) glutamate release through the anion channel Best-1 froma astrocytes Two modes (vesicular and channel-mediated) of glutamate release detected by sniffer cells. (a) Schematic diagram of the sniffer cell experiments; astrocytes stimulated by receptor agonists; sniffer cells expressing transfected ionotropic glutamate receptors. (b) Astrocytic activation by the application of the AMPAR/mGluR agonist L-quisqualate (red arrowhead) triggers a rapid burst of excitatory post-synaptic currents (EPSCs) in an N-methyl-D-aspartate (NMDA) receptor-expressing HEK sniffer cell. These EPSCs are thought to reflect quantal events of vesicular glutamate release from the astrocyte (adapted with permission from [13], fig. 3b). (c) Application of a TFLLR agonist of the PAR-1 metabotropic receptors (arrowhead) to astrocytes triggers a slow current in HEK cells expressing non-desensitizing α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors. This current is thought to be mediated by glutamate release through anion channel Best-1 and disappears in Best-1 knockout or knock-down astrocytes (adapted with permission from [8], sniffer experiments on a cultured astrocyte transfected with TREK-1 shrna, fig. 2i) Sahlender et al 2014
 release asztroból ez visszahat a neuronműködésre: AMPA/NMDAR-on át direkt posztszinatikus neuronális depolarizációt okozhat ILLETVE preszinaptikus mglur-t is aktiválhat gliotranszmitter ATP is")
60 Glia-neuron interakció jelenlegi modell: neuronok által aktivált gliális Ca ++ szignalizáció következtében gliotranszmitter (pl. Glu) release asztroból ez visszahat a neuronműködésre: AMPA/NMDAR-on át direkt posztszinatikus neuronális depolarizációt okozhat ILLETVE preszinaptikus mglur-t is aktiválhat gliotranszmitter ATP is lehet: vagy P2 purinreceptorokon át direktben hat vagy adenozinná bomlik, és P1 receptorokon hat Glia Neuron szignalizáció 2. Gliotranszmisszió gliális [Ca ++ ]ic : bekövetkezhet még spontán vagy pl. elektromos vagy mechanikus stimulálásra vezikuláris nem vezikuláris
61 Glia-neuron interakció Glia Neuron szignalizáció Példa 1. Hippocampus Heteroszinaptikus depresszió Feed-forward szinaptikus moduláció 2. Gliotransz misszió 1. Glutamát ürül (1, piros folt) amikor a Schafferkollaterális (S) CA1 piramissejt (P) szinapszis nagy frekvenciájú tüzelése stimulál egy asztrocitát 2. Az asztrocita erre ATP kibocsátással reagál 3. Ez gyorsan adenozinná alakul (2; kék folt), ami egy másik S-P kapcsolatot a preszinaptikus adenozin A1 receptoron keresztül szupresszálni fog! tonic suppression of synaptic transmission Volterra, Meldolesi 2005

62 Glia-neuron interakció Glia Neuron szignalizáció Azt, hogy adenozin tonikusan aktiválja az A1 receptorokat a HCban, már régóta lehetett tudni ( , Cunha, Dunwiddie). Hogyan jöttek rá arra, hogy mi az ADENOZIN forrása? 2. Gliotransz misszió transzmembrán domén nélküli synaptobrevin 2 citoplazmatikus doménját kondicionálisan, asztrospecifikus expresszáltatták ez így domináns-negatív inhibitora a SNARE-függő membrán fúziónak: asztroban nincs vezikulárus gliotranszmisszió dnsnare asztrociták
63 Glia-neuron interakció Glia Neuron szignalizáció 2. Gliotransz misszió Schaffer kollaterális - CA1 szinapszisoknál dnsnare asztrociták mellett erősebb szinaptikus transzmisszió /aktiváció volt, mint a WT egerekben farmakológiai vizsgálatok szerint ennek oka az extracelluláris adenozin szint csökkenése: pl. A1 adenozin-receptor blokkoló (DPCPX) növelte a szinaptikus transzmisszót WT egérben, de transzgén egérben nem volt hatása azt is kimutatták, hogy dnsnare overexpresszió valójában nem is a gliális adenozin release-t hanem a gliális ATP release-t zavarja meg (ATPt ektonukleotidázok hidrolizálják) Vagyis: egyértelműen asztrociták mediálják a heteroszinaptikus depressziót itt * adenozin sztori izgalmas folytatása a *-jelölt dián később

64 Glia-neuron interakció Glia Neuron szignalizáció Példa 2. Hippocampus Szinaptikus transzmisszió asztrociták általi modulációja Szomszédos piramissejtek excitációja és szinkronizációja nem direkt módon kapcsolt neuronok asztrociták általi összeköttetése 1. asztrocita spontán [Ca2+]ic oszcillációt mutat 2. ez gliális glutamát-releaset triggerel (piros folt) 3. ezt két piramissejt is érzékeli egy időben 2. Gliotransz misszió 4. szinkronizált NMDAR-dependens excitatorikus áramok generálódnak Fiziológiás körülmények között spontán oszcillációk ritkák (szeletben): kb 2 percenként Volterra, Meldolesi 2005
 GABAerg interneuronpiramissejt szinapszisban, repetitív tüzelés során 2.")
65 Glia-neuron interakció Glia Neuron szignalizáció Példa 3. Hippocampus Szinaptikus transzmisszió asztrociták általi modulációja Feedback moduláció 2. Gliotransz misszió Gátló szinapszis potenciációja GABAerg interneuronok és piramissejtek között 1. GABA ürül (1; sárga folt) GABAerg interneuronpiramissejt szinapszisban, repetitív tüzelés során 2. a GABA a szomszédos asztrocitán GABAB receptorokat aktivál 3. ez az asztrocita glutamát ürítéssel válaszol (2; red spot) mely a GABAerg interneuront éri 4. ez tovább potencírozza (feedback potenciáció) a GABAerg interneuron általi gátlást (3) (decreases GABA mediated synaptic failures) Volterra, Meldolesi 2005


66 Glia Neuron szignalizáció N G is 2. Gliotranszmisszió Példa 4. Hipotalamusz Hogyan szabályozzák a neuroendokrin neuronok szekréciós aktivitását az asztrociták Sisk, Foster 2004 HT GnRH neuroendokrin neuronok eminencia mediana, portális keringés hipofízis, gonadotropin (LH, FSH) szekréció véráram, gonádokban gametogenezis szteroid hormonok ürülése... feedback... stb.. Neuroendokrin területeken asztro mglur és AMPAR sejtmembránban kapcsolatban van erbb receptorokkal (EGF rec. család, ligand pl. TGFa, neuregulin) Diedzic 2003

67 Glia-neuron interakció Glia Neuron szignalizáció Példa 4. GnRH neuronok szinaptikus aktivációja glutamát, gliális aktivációja PGE2 által. 2. Gliotransz misszió morphological rearrangement prosztaglandin 1. neuronális Glu release 2. Glu aktiválja asztron mglur-t, AMPAR-t 3. ez mátrix metalloproteinázokat (MMP/ADAM) aktivál melyek katalizálják a pro-tgfa és a pro- NRG (pro-neuregulin) ektodomén vedlését (shedding) 4. a szekretált érett TGFa and NRG aktiválja az erbb1/erbb2 and erbb4/erbb2 heterodimereket 5. erbb receptorok serkentik a gliális prosztanoidok (pl. PGE2) szekrécióját - ez posztszinaptikus neuronális EP2 recetoron át (PGE2 receptor alosztály) GnRH release-t indukálhat 5. * emelett TGFa és NRG a HT asztro morfológia változását is indukálják Prevot 2010

68
69 Glia-neuron interakció aspektusai Szinaptikus működés szabályozása tripartite synapse neuron glia szignalizáció neuron/glia szinapszisok neurotranszmitter konc szabályozása signaling by exosomes gliotranszmisszió, példák glia morfológiai átrendeződésének hatása a gliotranszmisszióra glia neuron szignalizáció, gliotranszmitterek neuronális hálózatok kontrollja szinaptogenezis, szinapszis fenntartása, eliminációja
70 Glia-neuron interakció Glia Neuron szignalizáció Szinaptikus borítottság változása 3. Gliotranszmisszió és asztrocita morfológiai remodelling Már régebben, GFAP festésekből is kiderült, a gliamorfológia változik: - hipotalamusz: nucleus supraopticus (SON), suprachiasmaticus (SCN), arcuatus, preopticus : dehidratáció, cirkadián ritmus, fluktuáló szteroid szintek hatasara - hippocampus CA1, vizuális cortex: gazdag szenzoros input esetén! De disztális glia nyúlványok motilitásának változása neuronális működéssel összhangban még egyértelműbb sok példa Lásd: review of Theodosis Hypothalamo-neurohypophysial system 2. Basal hypothalamus 6. Cerebellum 3. Suprachiasmatic nucleus 7. Barrel cortex 4. Brain stem 8. Neocortex 5. Hippocampus 9. Visual cortex


71 3. Gliotranszmisszió és asztrocita morfológiai remodelling asztro SON első alkalommal vemhes; stb SON nucleus supraopticus: vajúdás, tejelválasztás alatt OT neuronok glia borítottsága nagyon lecsökken neuronális felszínek egymás mellé kerülnek: asztro retrakció, OT neuron nagyobbodás Theodosis 2008 Gyors változás: neuroszekréció indukciója után 1 órával már észlelhető, in vivo mérések szerint
 következik be az axonokról és erekről a neuroszekretoros tevékenység aktivációjának hatására:")
72 Hipofízisben is vannak (pituicita!) morfológiai változások: 3. Gliotranszmisszió és asztrocita morfológiai remodelling Neurohipofízisben pituicita retrakció (szürke) következik be az axonokról és erekről a neuroszekretoros tevékenység aktivációjának hatására: nagyobb release, könyebb véráramba jutás Theodosis

73 Glia-neuron interakció Glia Neuron szignalizáció Szinaptikus borítottság változása Nem stimulált: pl. szűz vagy elválasztás utáni patkány Stimulált: laktáló és dehidrált patkányok 3. Gliotranszmisszió és asztrocita morfológiai remodelling A hipotalamikus szupraoptikus magban az asztrociták morfológiai plaszticitása a tejelválasztással hozható összefüggésbe 1. oxytocin-termelő idegsejteken levő szinapszisokról asztrociták visszahúzzák nyúlványaikat laktáció alatt 2. így a glia kevésbé tudja visszavenni a szinapszisba ömlő glutamátot, és diffúzió is jobb (fekete nyilak) így a Glu képes távolabb elhelyezkedő mglur receptorokat aktiválni preszinaptikusan vagy szomszédos GABAerg terminálisokon 3. ez a neurotranszmitter-ürülés homoés heteroszinaptikus modulációját okozza: más glutamaterg és GABAerg szinapszisok működésének modulációja Theodosis 2008
 : (a)")
 Gyors")
74 Glia-neuron interakció Glia Neuron szignalizáció 3. Gliotranszmisszió és asztrocita morfológiai remodelling D-serine függő metaplaszticitás Szupraoptikus magban (SON) : (a) asztrocita-eredetű D-serine el tudja érni az NMDA receptorokat, és segíti az NMDAR mediálta transzmissziót és az LTP kialakulását (b) Ha a glianyúlványok visszahúzódnak, a D-serine nem tud ko-agonistaként működni az NMDA receptorokon, LTD alakul ki inkább (c) Gyors asztrocita (zöld) nyúlvány mozgás a dendrittüske (piros) körül (sec) Theodosis 2008 Halassa and Haydon rev. 2009
75 Glia-neuron interakció aspektusai Szinaptikus működés szabályozása tripartite synapse neuron glia szignalizáció neuron/glia szinapszisok neurotranszmitter konc szabályozása signaling by exosomes gliotranszmisszió, példák glia morfológiai átrendeződésének hatása a gliotranszmisszióra glia neuron szignalizáció, gliotranszmitterek neuronális hálózatok kontrollja szinaptogenezis, szinapszis fenntartása, eliminációja
 dendritből álló csoportot")
76 Glia-neuron interakció Glia Neuron szignalizáció 4. Neuronális Hálózatok kontrollja Philip Haydon Asztro network??? Halassa 2007 Reichenbach 2010 A, 3 asztrocita. Különböző neuronális kompartmentumokat más-más asztro kontrollálhat. B, egyetlen asztro is koordinálhat pl. egy több ( ezer!!) dendritből álló csoportot


77 glutamaterg neuron tüzel glia mglur aktiválódik glia Ca++ szint nő ez kiváltható így is: photolysis of caged Ca++ glia erre Glutamátot ürít! gliából kiszabaduló Glutamát neuronokat aktivál neuronális epileptikus aktivitás
 gliotranszmisszió csökkent lassú oszcillációk a szomatoszenzoros kéregben Representative LFP (local field")
78 Glia-neuron interakció Glia Neuron szignalizáció 4. Neuronális Hálózatok kontrollja dnsnare transzgén egér: asztrocitában nincs (vezikuláris) gliotranszmisszió csökkent lassú oszcillációk a szomatoszenzoros kéregben Representative LFP (local field potential) recording showing slow oscillation activity in the somatosensory cortex In vivo current-clamp recordings from representative neurons showing slow oscillations in the cortex of WT asztro ezt a folyamatot két ponton modulálja: 1. adenozin A1 receptor aktiváció gátlódik (tehát heteroszinaptikus gátlás megszűnik) 2. neuronális NMDAR funkció is csökken (itt a gliális D-serine ürülése csökken)! Tehát asztrocita eredetű gliotranszmisszió valóban befolyásolja agyi hálózatok működését in vivo
79 NREM Glia-neuron interakció Glia Neuron szignalizáció Alvás korai kutatása: vér, CSF transzfer alvó állatból éberbe: sikertelenek de kiderül, hogy agyban lokálisan termelődő faktorokról van szó: sleep factors. Adenozin is egy endogén sleep factor (2002, 2003 Porkka-Heiskannen et al): ébrenlét adenozin szint progresszívan nő alvás alatt leapad CSF: cerebrospinal fluid 4. Neuronális Hálózatok kontrollja NREM (Non-Rapid-Eye-Movement) fázis (vagy lassú hullámú alvás) : lassú oszcillációk

80 sok Adenozin: - akkumulálódva álmossá tesz, lelassítja a neuronális működést - ér-dilatációt okoz (alvás alatti jobb oxigénellátás) Koffein: - adenozin-receptorhoz köt, melyhez ezáltal az adenozin nem fér hozzá - nagyobb neuronális aktivitás, éberség - a hipofízis a nagyobb neuronális aktivitás hatására adrenalint ürítésre utasítja a mellékvesét - emiatt: pupilla kitágul, gyorsabb szívverés, légutak megnyílnak (asztmások adrenalint kapnak), vérnyomás nő, vércukorszint nő, izmok vérellátása fokozódik, - adenozin-receptor gátlás agyi érkontrakciót is okoz (néhány fejfájás elleni orvosság koffein-tartalmú) KV addiktív! (heroin is a dopamin szintet emeli ) koffein 6 óra alatt tűnik el a szervezetből és ne feledd: koffein RyR agonista! - és még valami: Migraine pain is caused by vasodilation in the cranial blood vessels! Szemben a fejfájással, ahol érkonstrikció jellemző.
 akkor ezekben az egerekben romlik a memória a")
81 Glia-neuron interakció dnsnare egereken krónikus EEG mérések (beültetett elektródák) miközben alszanak csökkent lassú hullámú aktivitás (slow wave activity, SWA) 5. Viselkedés kontrollja * itt van az adenozin sztori izgalmas folytatása ha A1 receptorokat kiütik előagyi excitatorikus neuronokon (Bjorness 2009) akkor ezekben az egerekben romlik a memória a krónikus alváshiány miatt dnsnare egerek rezisztensek a rövidtávú alvásdepriváció okozta kognitív zavarokra!! Vagyis: asztro eredetű adenozin viselkedési mintázatokba is beleszól! Purinergic Gliotransmission Contributes to Memory Impairment Following Sleep Loss (A) Novel object recognition (NOR) paradigm: mice are trained to recognize two identical objects and are either left undisturbed or sleep deprived for 6 hr following training. At hour 24, mice are tested for the ability to recognize a novel object replacing one of the familiar objects. (B) SD impairs NOR in wild-type mice. (C) dnsnare mice are unaffected by the effects of SD on NOR memory Halassa 2009
82 dnsnare : 2005 óta min. 12 cikk, jó újságokban stb.. de!!! Vita...válasz is volt, azóta 3 új cikk dnsnare egérrel ( ).

83 Glia-neuron interakció 5. Viselkedés kontrollja...accumulating evidence points to an involvement of astrocytes in higher cognitive processes...

84 Glia-neuron interakció 5. Viselkedés kontrollja


: sejt-sejt és sejt-mátrix adhézió (fibrinogen,")
85 Glia-neuron interakció Szinaptogenezis Néhány hetes tiszta RGC (retinal ganglion cell) vagy gerincvelői motoneuron tenyészet: - alacsony szinaptikus aktivitás, - szinaptikus fehérjék alacsony szintje Asztroglia + RGC kokultúra: - nagy szinaptikus aktivitás (100 X); - szinapszisszám nő (7X) 1 héttel asztrociták eltávolítása után a kokultúrákból: - a legtöbb szinapszis eltűnik ACM: asztro conditioned medium, asztro tenyészet felülúszója Ullian EM. Christopherson KS, Barres BA 2004 Thrombospondin (TSP): sejt-sejt és sejt-mátrix adhézió (fibrinogen, fibronectin, laminin, collagen és integrin kötő glikoprotein) In vitro szinapszis OK Tenyésztett neuronok közti szinapszisok száma

86
: makromolekulák paracelluláris transzportja és vérsejtek")
")
87 Neurovaszkuláris kapcsoltság Agyi kapilláris Perifériás kapilláris Agyi endotél: TJ kapcsoltság (BBB): makromolekulák paracelluláris transzportja és vérsejtek inváziója gátolt. (de pl. choroid plexusban: fenesztrált endotélum, igaz itt az epitélsejtek között van TJ) Perifériás kapillárisokban jellemzőek a pinocitotikus granulumok

 http://www.netterimages.com/image/13431.")
88 Neurovaszkuláris kapcsoltság endotélsejtek közötti tight junction Agyi kapilláris. endothelial cell (EC) tight junctions (arrows) part of a pericyte (P) astrocytes end-feet (EF)
 This sheath surrounds the capillary and pericyte. A perivascular astrocyte with nucleus (N) and cell body (ascb) embraces (aspve II) the endothelial tube.")
89 Neurovaszkuláris kapcsoltság Serial Section Electron Microscopy and 3D Reconstruction The capillary surface is completely covered by a perivascular glial sheath. A) This sheath surrounds the capillary and pericyte. A perivascular astrocyte with nucleus (N) and cell body (ascb) embraces (aspve II) the endothelial tube. The pericyte (pe) is seen through discontinuities in the endfoot indexed aspve III. Astrocytic processes (asp) from the endfeet stretch out in different directions. B) Structure in A viewed from the right. All four endfeet (pve I-IV) in the reconstruction is represented together with the pericyte (pe). Part of a probable microglial cell process, indexed as a perivascular cell process (pvcp), is seen peripheral to the glial sheath. C,D) Removal of the pericyte, pve III, and pve IV (structures reconstructed in C) allows the remaining perivascular sheath (pve I and II) to be viewed from the inside (D). The most important feature in C is the six linearly arranged discontinuities of the third endfoot (pveiii). E) Elements reconstructed in D are shown in color and entered into a drawing adapted from Ramon y Cajal (1911).
 endotél sejtek közötti TJ kapcsolatot indukálják és - fenntartják az endotél luminális/bazális polarizáltságát (ioncsatorna, AQP, receptorkészlet) - prostaglandin (PGE), nitrogén monoxid")
90 Neurovaszkuláris kapcsoltság Asztro végtalpak: teljesen körbeölelik kapillárisokat Asztro eredetű faktorok: - (pl. TGFa, GDNF) endotél sejtek közötti TJ kapcsolatot indukálják és - fenntartják az endotél luminális/bazális polarizáltságát (ioncsatorna, AQP, receptorkészlet) - prostaglandin (PGE), nitrogén monoxid (NO) vazodilatáció - endothelin, ATP - vazokonstrikció - arachidonsav (AA) - vazokonstrikció és/vagy dilatáció Endotélsejtek is jeleznek asztro felé: - LIF (leukemia inhibitory factor) az asztrociták érését segíti Agyi endotél szelektív permeabilitása: (1) adenine-nucleotide binding (ABC) cassette transzporterek (energiaigényesek) melyek xenobiotikumokat exkretálnak (drogok, antibiotikumok, citosztatikumok stb) cytostatics, opioids etc. nem jutnak át BBB-n) (2) aminosav transzporterek (L1) (3) glükóz transzporterek (GLUT1 type) (4) ion exchangerek, stb.

91 Neurovaszkuláris kapcsoltság Transzporter rendszerek (protein szinten ismertek) BCRP, breast cancer resistance protein (also known as ABC transporter G family member 2); GLUT, solute carrier family 2, facilitated glucose transporter member; LRP, low-density lipoprotein receptor-related protein family member; MCT, monocarboxylic acid transporter family member; MRP, multidrug resistanceassociated protein family member; OAT, organic anion transporter family member; OATP, organic anion transporter polypeptide family member; Pgp, P-glycoprotein; RAGe, advanced glycosylation end productspecific receptor; RLiP76, Ral-binding protein 1 Neuwelt 2011
 NVU extended NvU: neuronok,")


92 Neurovaszkuláris kapcsoltság Neurovaszkuláris unit (NvU) NVU extended NvU: neuronok, asztrociták, mikroglia, endotél, pericita, simaizom sejt, vérsejtek: polymorfonukleáris (PMN) sejtek, limfociták, monociták, + perivaszkuláris makrofágok BBB modell + asztro és pericita Cardoso 2010 Neuwelt 2011

93 Neurovaszkuláris kapcsoltság Agyi mikrocirkuláció szabályozása Funkcionális hyperaemia: lokális neuronális aktivitás esetén gyorsan nő a lokális agyi vérellátás 1890, Sherrington - vazodilatáció az aktív neuronokhoz közel, um távolságban következik be - sokféle elmélet próbálta magyarázni: lokális neuronális vazoaktív faktor release, lokális innerváció, NO termelés - ma: asztro szerepe egyértelmű! neuronális aktivitás asztro [Ca ++ ] ic arachidonsav (AA) kibocsátása glia végtalpból asztro Ca++ szignalizáció gátlása szétkapcsolja a neuronális aktivitást az értónus szabályozástól!! e végtalp által körbefogott kis arteriolák tónusának változása vazodilatáció (blood flow nő): ha AA prosztaglandin-származékká alakul cyclooxygenáz segítségével (ezt aszpirin gátolja) vazokonstrikció: ha AA átalakulás útja 20- hydroxyeicosatetraenoic acid (2-HETE), cytochrome p450 enzim segítségével

94 Agyi mikrocirkuláció szabályozása David Attwell et al Nature EET: epoxyeicosatrienoic acids Pathways from astrocytes and neurons that regulate blood flow by sending messengers to influence the smooth muscle around the arterioles that supply oxygen and glucose to the cells (shown as the vessel lumen surrounded by endothelial cells and smooth muscle). In neurons, synaptically released glutamate acts on N-methyl-D-aspartate receptors (NMDAR) to raise [Ca 2+ ] i, causing neuronal nitric oxide synthase (nnos) to release NO, which activates smooth muscle guanylate cyclase. This generates cgmp to dilate vessels. Raised [Ca 2+ ] i may also (dashed line) generate arachidonic acid (AA) from phospholipase A 2 (PLA 2 ), which is converted by COX2 to prostaglandins (PG) that dilate vessels. Glutamate raises [Ca 2+ ] i in astrocytes by activating metabotropic glutamate receptors (mglur), generating arachidonic acid and thus three types of metabolite: prostaglandins (by COX1/3, and COX2 in pathological situations) and EETs (by P450 epoxygenase) in astrocytes, which dilate vessels, and 20-HETE (by ω-hydroxylase) in smooth muscle, which constricts vessels. A rise of [Ca 2+ ] i in astrocyte endfeet may activate Ca 2+ -gated K + channels (g K(Ca) ), releasing K +, which also dilates vessels.
95 Neurovaszkuláris kapcsoltság Agyi mikrocirkuláció szabályozása AA membrán foszfolipidekből keletkezik Ca 2+ -függő és Ca 2+ -independens lipázok segítségével. Zöld: vazodilátor, Piros: vazokonstriktor, Kék: a megfelelő enzimek lokalizációja. COX, cyclooxygenase; CYP, cytochrome P450 superfamily of enzymes; EET, epoxyeicosatrienoic acid; 20-HETE, 20-hydroxyeicosatetraenoic acid; HPETE, hydroperoxy-eicosatetraenoic acid. David Attwell et al Nature


96 Ide tartozik a Glimfatikus rendszerről szóló kiselőadás!! korábban megbeszéltük! tessék átnézni
+(b) a és b szignálok integrációja Piros nyíl: véráramlás Kék nyíl: neuronális aktivitás szignáljainak elvezetődése a kapillárisoktól a proximális artériákig (1) Asztro")
97 Neurovaszkuláris kapcsoltság Proximális integráció modellje Agyi mikrocirkuláció szabályozása A piális artéria B penetráló artéria C prekapilláris artéria D valódi kapilláris (a, b) neuronális aktiváció helyszínei (a)+(b) a és b szignálok integrációja Piros nyíl: véráramlás Kék nyíl: neuronális aktivitás szignáljainak elvezetődése a kapillárisoktól a proximális artériákig (1) Asztro végtalp szignáljai szabályozzák lokális arteriolát a simaizomsejtek tónusának változtatásával. A fehér nyilak a perifériáról jövő info integrációját jelzik. (2) A kapilláris véráramlását Mindig is kérdéses volt, hogy a lokális neuronális kisrészt lokális pericitakonstrikció szabályozza, tüzelés infoja hogyan jut el a proximális (kezdeti) artériákig, nagyobb erekig a kapillárisoktól: asztro nagyrészt a prekapilláris illetve pericita GJ mediálta retrográd info-terjedés területről érkező véráram erek mentén!! egyelőre ez csak egy hipotézis!!! Itoh 2012
Az adenozin Adenozin receptorok:
 Az adenozin Nukleinsavak és energiaraktározó vegyületek építőeleme Jelenléte ATP hidrolízisére utal -> extracelluláris szintje utal a korábbi neuronális és gliális aktivitásra Adenozin receptorok: 1-es
Az adenozin Nukleinsavak és energiaraktározó vegyületek építőeleme Jelenléte ATP hidrolízisére utal -> extracelluláris szintje utal a korábbi neuronális és gliális aktivitásra Adenozin receptorok: 1-es
Glia-neuron interakció aspektusai. Neurovaszkuláris. kapcsoltság
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
AsztroGlia - neuron interakció
 2011.04. 06. AsztroGlia - neuron interakció protoplazmás asztroglia (szürkeállomány); rostos asztroglia (fehérállomány); oligodendroglia (CNS); Schwann sejt (PNS); radiális glia (cortex); Bergmann glia
2011.04. 06. AsztroGlia - neuron interakció protoplazmás asztroglia (szürkeállomány); rostos asztroglia (fehérállomány); oligodendroglia (CNS); Schwann sejt (PNS); radiális glia (cortex); Bergmann glia
Glia-neuron interakció aspektusai. Neurovaszkuláris. kapcsoltság
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai. Neuron-glia metabolikus együttműködés
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai. Neuron-glia metabolikus együttműködés
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai. Neuron-glia metabolikus együttműködés
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása Neuron-glia metabolikus együttműködés glutamát-glutamin
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása Neuron-glia metabolikus együttműködés glutamát-glutamin
Glia-neuron interakció aspektusai. Neurovaszkuláris. kapcsoltság
 aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása neurovaszkuláris
aspektusai Agyi mikrokörnyezet szabályozása Neurovaszkuláris kapcsoltság Neuron-glia metabolikus együttműködés ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása neurovaszkuláris
Glia - neuron interakció
 Glia - neuron interakció 2017.05.05. kornyei@koki.hu Gliális sejttípusok az idegrendszerben neuroektodermális eredet (kivéve mikroglia) NEUROGLIA glia glia (görög): ragadós 1858, Rudolf Virchow agyi kötőszövet
Glia - neuron interakció 2017.05.05. kornyei@koki.hu Gliális sejttípusok az idegrendszerben neuroektodermális eredet (kivéve mikroglia) NEUROGLIA glia glia (görög): ragadós 1858, Rudolf Virchow agyi kötőszövet
Computational Neuroscience
 Computational Neuroscience Zoltán Somogyvári senior research fellow KFKI Research Institute for Particle and Nuclear Physics Supporting materials: http://www.kfki.hu/~soma/bscs/ BSCS 2010 Lengyel Máté:
Computational Neuroscience Zoltán Somogyvári senior research fellow KFKI Research Institute for Particle and Nuclear Physics Supporting materials: http://www.kfki.hu/~soma/bscs/ BSCS 2010 Lengyel Máté:
Glia - neuron interakció
 Glia - neuron interakció 2016.04.27. kornyei@koki.hu Neuronális heterogenitás: Cajal rajzai alapján Gliasejtek morfológiai diverzitása Gustaf Retzius hgfap-gfp Emsley 2006 Gliális sejttípusok az idegrendszerben
Glia - neuron interakció 2016.04.27. kornyei@koki.hu Neuronális heterogenitás: Cajal rajzai alapján Gliasejtek morfológiai diverzitása Gustaf Retzius hgfap-gfp Emsley 2006 Gliális sejttípusok az idegrendszerben
Asztrociták: a központi idegrendszer sokoldalú sejtjei. 2009.11.04. Dr Környei Zsuzsanna
 Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV
 KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus
 Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Humán asztrociták. Nagyobb és komplexebb. idegrendszeri fejlődésben jelentős szerepű
 Humán asztrociták Nagyobb és komplexebb idegrendszeri fejlődésben jelentős szerepű Forrás: Human vs Rodent astrocytes. (Courtesy Alexi Verkhratsky (Chapter 3), Neuroglia by Kettenmann) Glial Progenitor
Humán asztrociták Nagyobb és komplexebb idegrendszeri fejlődésben jelentős szerepű Forrás: Human vs Rodent astrocytes. (Courtesy Alexi Verkhratsky (Chapter 3), Neuroglia by Kettenmann) Glial Progenitor
Az idegrendszer határfelszínei és a neurovaszkuláris egység
 Az idegrendszer határfelszínei és a neurovaszkuláris egység Határfelszínek az idegrendszerben vér-agy gát [blood-brain barrier (BBB)] vér-liquor gát [bloodcerebrospinal fluid barrier (BCSFB)] arachnoid
Az idegrendszer határfelszínei és a neurovaszkuláris egység Határfelszínek az idegrendszerben vér-agy gát [blood-brain barrier (BBB)] vér-liquor gát [bloodcerebrospinal fluid barrier (BCSFB)] arachnoid
Membrántranszport. Gyógyszerész előadás Dr. Barkó Szilvia
 Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA
 A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA 2017.11.14. AZ ASZTROGLIA SEJTEK FONTOSABB TULAJDONSÁGAI AZ EPILEPTIKUS AKTIVITÁS SZEMPONTJÁBÓL (Devinsky és mtsai.,
A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA 2017.11.14. AZ ASZTROGLIA SEJTEK FONTOSABB TULAJDONSÁGAI AZ EPILEPTIKUS AKTIVITÁS SZEMPONTJÁBÓL (Devinsky és mtsai.,
A nemi különbségek vizsgálatáról lévén szó, elsődleges volt a nemi hormonok, mint belső környezetbeli különbségeket létrehozó tényezők szerepének
 Kutatási beszámoló Pályázatunk célja annak kiderítése volt, hogy az agyi asztrociták mutatnak-e nemi különbségeket, akár struktura, akár területi megoszlás, akár reaktivitás tekintetében. Alkalmazott megközelítésünk
Kutatási beszámoló Pályázatunk célja annak kiderítése volt, hogy az agyi asztrociták mutatnak-e nemi különbségeket, akár struktura, akár területi megoszlás, akár reaktivitás tekintetében. Alkalmazott megközelítésünk
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az fmri alapjai BOLD fiziológia. Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika
 Az fmri alapjai BOLD fiziológia Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika T2* Az obszervált transzverzális relaxáció (T2*) több különböző komponens összege Many physical effects result
Az fmri alapjai BOLD fiziológia Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika T2* Az obszervált transzverzális relaxáció (T2*) több különböző komponens összege Many physical effects result
Új szignalizációs utak a prodromális fázisban. Oláh Zita
 Új szignalizációs utak a prodromális fázisban Oláh Zita 2015.10.07 Prodromális fázis Prodromalis fázis: De mi történik?? Beta-amiloid: OK vagy OKOZAT? Beta-amiloid hogyan okozhat neurodegenerációt? Tau
Új szignalizációs utak a prodromális fázisban Oláh Zita 2015.10.07 Prodromális fázis Prodromalis fázis: De mi történik?? Beta-amiloid: OK vagy OKOZAT? Beta-amiloid hogyan okozhat neurodegenerációt? Tau
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Az agyi metabolizmus, és a vérkeringés metabolikus szabályozása. Dr. Domoki Ferenc
 Az agyi metabolizmus, és a vérkeringés metabolikus szabályozása Dr. Domoki Ferenc Klasszikus agyi metabolizmus Glükóz központi (egyedüli) szerepet játszik a neuronok anyagcseréjében Aerob oxidáció agyi
Az agyi metabolizmus, és a vérkeringés metabolikus szabályozása Dr. Domoki Ferenc Klasszikus agyi metabolizmus Glükóz központi (egyedüli) szerepet játszik a neuronok anyagcseréjében Aerob oxidáció agyi
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Construction of a cube given with its centre and a sideline
 Transformation of a plane of projection Construction of a cube given with its centre and a sideline Exercise. Given the center O and a sideline e of a cube, where e is a vertical line. Construct the projections
Transformation of a plane of projection Construction of a cube given with its centre and a sideline Exercise. Given the center O and a sideline e of a cube, where e is a vertical line. Construct the projections
Az agyi értónust befolyásoló tényezők
 2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
2006 1. Nemszinaptikus receptorok és szubmikronos Ca2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra.
 2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Supplementary materials to: Whole-mount single molecule FISH method for zebrafish embryo
 Supplementary materials to: Whole-mount single molecule FISH method for zebrafish embryo Yuma Oka and Thomas N. Sato Supplementary Figure S1. Whole-mount smfish with and without the methanol pretreatment.
Supplementary materials to: Whole-mount single molecule FISH method for zebrafish embryo Yuma Oka and Thomas N. Sato Supplementary Figure S1. Whole-mount smfish with and without the methanol pretreatment.
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN
 AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN Kormann Eszter Idegi sejtdifferenciáció 2. 2012.12.10. AZ EPILEPSZIÁRÓL RÖVIDEN Definíció: az agyban kialakuló betegség, melyet legalább két alkalommal
AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN Kormann Eszter Idegi sejtdifferenciáció 2. 2012.12.10. AZ EPILEPSZIÁRÓL RÖVIDEN Definíció: az agyban kialakuló betegség, melyet legalább két alkalommal
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
A tanulási és emlékezési zavarok pathofiziológiája. Szeged,
 A tanulási és emlékezési zavarok pathofiziológiája Szeged, 2015.09.09 Szerkezet, működés, információáramlás, memória, tanulás: 1. Neokortex 2. Limbikus rendszer Limbikus rendszer és a memória Paul Broca
A tanulási és emlékezési zavarok pathofiziológiája Szeged, 2015.09.09 Szerkezet, működés, információáramlás, memória, tanulás: 1. Neokortex 2. Limbikus rendszer Limbikus rendszer és a memória Paul Broca
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Neurovaszkuláris csatolás
 Neurovaszkuláris csatolás Farkas Eszter 2016. október 20. Az első bizonyíték a neurovaszkuláris csatolásra Kognitív feladat végzése az agytérfogat változásával jár (Mosso, 1881) A Roy-Sherrington elv Neurovaszkuláris
Neurovaszkuláris csatolás Farkas Eszter 2016. október 20. Az első bizonyíték a neurovaszkuláris csatolásra Kognitív feladat végzése az agytérfogat változásával jár (Mosso, 1881) A Roy-Sherrington elv Neurovaszkuláris
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika
 Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Receptor Tyrosine-Kinases
 Receptor Tyrosine-Kinases MAPkinase pathway PI3Kinase Protein Kinase B pathway PI3K/PK-B pathway Phosphatidyl-inositol-bisphosphate...(PI(4,5)P 2...) Phosphatidyl-inositol-3-kinase (PI3K) Protein kinase
Receptor Tyrosine-Kinases MAPkinase pathway PI3Kinase Protein Kinase B pathway PI3K/PK-B pathway Phosphatidyl-inositol-bisphosphate...(PI(4,5)P 2...) Phosphatidyl-inositol-3-kinase (PI3K) Protein kinase
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
Cerebrovaszkuláris elváltozások öregedésben és Alzheimer-kórban
 Cerebrovaszkuláris elváltozások öregedésben és Alzheimer-kórban Farkas Eszter 2016. november 17. Mi történik az agyunkkal, ahogy öregszünk? ( Luke, én aaah a fenébe is, valami fontosat akartam mondani,
Cerebrovaszkuláris elváltozások öregedésben és Alzheimer-kórban Farkas Eszter 2016. november 17. Mi történik az agyunkkal, ahogy öregszünk? ( Luke, én aaah a fenébe is, valami fontosat akartam mondani,
Supporting Information
 Supporting Information Cell-free GFP simulations Cell-free simulations of degfp production were consistent with experimental measurements (Fig. S1). Dual emmission GFP was produced under a P70a promoter
Supporting Information Cell-free GFP simulations Cell-free simulations of degfp production were consistent with experimental measurements (Fig. S1). Dual emmission GFP was produced under a P70a promoter
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Komplementrendszer szerepe
 Komplementrendszer szerepe Veerhuis et al., 2011 Készítette: Udvari Edina Vérben és testnedvekben Nagy része a májban termelődik, de makrofágok, endotélsejtek is termelnek Klasszikus, alternatív és lektinindukált
Komplementrendszer szerepe Veerhuis et al., 2011 Készítette: Udvari Edina Vérben és testnedvekben Nagy része a májban termelődik, de makrofágok, endotélsejtek is termelnek Klasszikus, alternatív és lektinindukált
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA NOVEMBER
 Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
Szinapszis, szinaptogenezis
 Szinapszis, szinaptogenezis en passant és terminális szinapszisok parakrin szinapszis Kémiai szinapszis Jena, B.J., J. Cell. Mol. Med. Vol 8, No 1, 2004 SNARE proteins ("SNAP and NSF a>achment receptors")
Szinapszis, szinaptogenezis en passant és terminális szinapszisok parakrin szinapszis Kémiai szinapszis Jena, B.J., J. Cell. Mol. Med. Vol 8, No 1, 2004 SNARE proteins ("SNAP and NSF a>achment receptors")
Interneurális kommunikáció
 Interneurális kommunikáció 2010/2011 Sejtélettan II. Szinapszisok osztályozása Na channel Transmitter vesicle Local circuit current Na 2+ Ca channel PRE- SYNAPTIC Ca++ PRE- SYNAPTIC Ca-induced exocytosis
Interneurális kommunikáció 2010/2011 Sejtélettan II. Szinapszisok osztályozása Na channel Transmitter vesicle Local circuit current Na 2+ Ca channel PRE- SYNAPTIC Ca++ PRE- SYNAPTIC Ca-induced exocytosis
Supplementary Table 1. Cystometric parameters in sham-operated wild type and Trpv4 -/- rats during saline infusion and
 WT sham Trpv4 -/- sham Saline 10µM GSK1016709A P value Saline 10µM GSK1016709A P value Number 10 10 8 8 Intercontractile interval (sec) 143 (102 155) 98.4 (71.4 148) 0.01 96 (92 121) 109 (95 123) 0.3 Voided
WT sham Trpv4 -/- sham Saline 10µM GSK1016709A P value Saline 10µM GSK1016709A P value Number 10 10 8 8 Intercontractile interval (sec) 143 (102 155) 98.4 (71.4 148) 0.01 96 (92 121) 109 (95 123) 0.3 Voided
-Két fő korlát: - asztrogliák rendkívüli morfológiája -Ca szignálok értelmezési nehézségei
 Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
A neurogliaform sejtek szerepe az agykéregben
 A neurogliaform sejtek szerepe az agykéregben Ph.D. értekezés tézisei Oláh Szabolcs Témavezetõ: Tamás Gábor, Ph.D., D.Sc. SZEGEDI TUDOMÁNYEGYETEM Természettudományi és Informatikai Kar Élettani, Szervezettani
A neurogliaform sejtek szerepe az agykéregben Ph.D. értekezés tézisei Oláh Szabolcs Témavezetõ: Tamás Gábor, Ph.D., D.Sc. SZEGEDI TUDOMÁNYEGYETEM Természettudományi és Informatikai Kar Élettani, Szervezettani
GLUTAMINSAV-GABA CSEREFOLYAMAT A KÖZPONTI IDEGRENDSZERBEN
 GLUTAMINSAV-GABA CSEREFOLYAMAT A KÖZPONTI IDEGRENDSZERBEN (Doktori Értekezés Tézisei) Héja László ELTE TTK, Kémia Doktori Iskola (Dr. Inzelt György, D.Sc.) Szintetikus Kémia, Anyagtudomány, Biomolekuláris
GLUTAMINSAV-GABA CSEREFOLYAMAT A KÖZPONTI IDEGRENDSZERBEN (Doktori Értekezés Tézisei) Héja László ELTE TTK, Kémia Doktori Iskola (Dr. Inzelt György, D.Sc.) Szintetikus Kémia, Anyagtudomány, Biomolekuláris
IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója
 IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója A Golgi-impregnáció kulcsfontosságú módszer a struktúra megismerésében rer: tigroid vs Nissl rögök Tigroid: Lenhossék
IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója A Golgi-impregnáció kulcsfontosságú módszer a struktúra megismerésében rer: tigroid vs Nissl rögök Tigroid: Lenhossék
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Stressz és neurogenezis
 Stressz és neurogenezis Charlotte A. Oomen et al. J. Neurosci. 2010; Mirescu et al., Nature Neuroscience 2004 I.M. Abraham, et al., J. Neuroendocrinology, 2001 Neurogenesis and neuronal regeneration in
Stressz és neurogenezis Charlotte A. Oomen et al. J. Neurosci. 2010; Mirescu et al., Nature Neuroscience 2004 I.M. Abraham, et al., J. Neuroendocrinology, 2001 Neurogenesis and neuronal regeneration in
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
Nevezze meg a jelölt csontot latinul! Name the bone marked! Nevezze meg a jelölt csont típusát! What is the type of the bone marked?
 1 Nevezze meg a jelölt csontot latinul! Name the bone marked! 2 Nevezze meg a jelölt csont típusát! What is the type of the bone marked? 3 Milyen csontállomány található a jelölt csont belsejében? What
1 Nevezze meg a jelölt csontot latinul! Name the bone marked! 2 Nevezze meg a jelölt csont típusát! What is the type of the bone marked? 3 Milyen csontállomány található a jelölt csont belsejében? What
Az alvás biológiája. Lőrincz Magor
 Az alvás biológiája Lőrincz Magor mlorincz@gmail.com Alvás a. Szerepe b. cirkadián ritmusok, mechanizmusai b. elektromos jelenségek, mechanizmusai c. szabályzás d. alvászavarok Alvás a. Szerepe b. cirkadián
Az alvás biológiája Lőrincz Magor mlorincz@gmail.com Alvás a. Szerepe b. cirkadián ritmusok, mechanizmusai b. elektromos jelenségek, mechanizmusai c. szabályzás d. alvászavarok Alvás a. Szerepe b. cirkadián
1. Mi jellemző a connexin fehérjékre?
 Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
A sejtek közöti kommunikáció formái. BsC II. Sejtélettani alapok Dr. Fodor János
 A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
Tanulás és memória. A tanulás és a memória formái, agyi lokalizációjuk és celluláris mechanizmusok. Pszichofiziológia ea.
 Tanulás és memória A tanulás és a memória formái, agyi lokalizációjuk és celluláris mechanizmusok Pszichofiziológia ea., 2016, Bali Zsolt Mi a tanulás és a memória? A tanulás tartós belső reprezentációk
Tanulás és memória A tanulás és a memória formái, agyi lokalizációjuk és celluláris mechanizmusok Pszichofiziológia ea., 2016, Bali Zsolt Mi a tanulás és a memória? A tanulás tartós belső reprezentációk
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Synchronization of cluster-firing cells in the medial septum
 Synchronization of cluster-firing cells in the medial septum Balázs Ujfalussy and Tamás Kiss 25. december 9. Tartalom Miért burstöl a Wang-féle sejt? - bifurkációk Xpp-vel. Az ANDREW-project második félideje
Synchronization of cluster-firing cells in the medial septum Balázs Ujfalussy and Tamás Kiss 25. december 9. Tartalom Miért burstöl a Wang-féle sejt? - bifurkációk Xpp-vel. Az ANDREW-project második félideje
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Neurofiziológia I. Schlett Katalin Élettani és Neurobiológiai Tanszék. tel: 8380 mellék
 Neurofiziológia I. Schlett Katalin Élettani és Neurobiológiai Tanszék schlettk@ludens.elte.hu tel: 8380 mellék ajánlott irodalom: From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience
Neurofiziológia I. Schlett Katalin Élettani és Neurobiológiai Tanszék schlettk@ludens.elte.hu tel: 8380 mellék ajánlott irodalom: From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
A hízósejtek szerepe az immunológiai folyamatokban
 A hízósejtek szerepe az immunológiai folyamatokban Berki Timea Boldizsár F, Bartis D, Talabér G, Szabó M, Németh P, University of Pécs, Department of Immunology & Biotechnology, Pécs, Hungary Additon of
A hízósejtek szerepe az immunológiai folyamatokban Berki Timea Boldizsár F, Bartis D, Talabér G, Szabó M, Németh P, University of Pécs, Department of Immunology & Biotechnology, Pécs, Hungary Additon of
Degeneráció és regeneráció az idegrendszerben
 Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
A felszíni ektoderma differenciációja
 A felszíni ektoderma differenciációja - elkülönülése - kialakulása az egyedfejlődés során - szerkezete és a felnőttkori őssejtek -interfollikuláris epidermis -szőrtüsző -faggyúmirigy - a bőrpótlás (klinikai)
A felszíni ektoderma differenciációja - elkülönülése - kialakulása az egyedfejlődés során - szerkezete és a felnőttkori őssejtek -interfollikuláris epidermis -szőrtüsző -faggyúmirigy - a bőrpótlás (klinikai)
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
2012.11.27. Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai. Neuronok izolálása I
 Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai Sejtszintű elektrofiziológia 1.: csatornák funkcionális Sejtszintű elektrofiziológia 2.: izolált/sejtkultúrában
Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai Sejtszintű elektrofiziológia 1.: csatornák funkcionális Sejtszintű elektrofiziológia 2.: izolált/sejtkultúrában
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül.
 7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
A feladat: A glükóz aerob oxidációja. Az oxigén alternatív felhasználása. A glükóz alternatív felhasználása
 A feladat: Az agyi metabolizmus, mitokondriumok az agyban A neuronok intermedier anyagcseréjének és energiaháztartásának jellegzetességei. A neuronális energiaháztartás zavarai, a mitochondriumok szerepe
A feladat: Az agyi metabolizmus, mitokondriumok az agyban A neuronok intermedier anyagcseréjének és energiaháztartásának jellegzetességei. A neuronális energiaháztartás zavarai, a mitochondriumok szerepe
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
FAMILY STRUCTURES THROUGH THE LIFE CYCLE
 FAMILY STRUCTURES THROUGH THE LIFE CYCLE István Harcsa Judit Monostori A magyar társadalom 2012-ben: trendek és perspektívák EU összehasonlításban Budapest, 2012 november 22-23 Introduction Factors which
FAMILY STRUCTURES THROUGH THE LIFE CYCLE István Harcsa Judit Monostori A magyar társadalom 2012-ben: trendek és perspektívák EU összehasonlításban Budapest, 2012 november 22-23 Introduction Factors which
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
A posztszinapszis és a PSD (posztszinaptikus. szinaptikus plaszticitásban játszott szerepük
 A posztszinapszis és a PSD (posztszinaptikus denzitás), valamint a szinaptikus plaszticitásban játszott szerepük A posztszinapszis szimmetrikus (Gray II): variábilis, nagy vezikulák; ált. gátló aszimmetrikus
A posztszinapszis és a PSD (posztszinaptikus denzitás), valamint a szinaptikus plaszticitásban játszott szerepük A posztszinapszis szimmetrikus (Gray II): variábilis, nagy vezikulák; ált. gátló aszimmetrikus
CT/MRI képalkotás alapjai. Prof. Bogner Péter
 CT/MRI képalkotás alapjai Prof. Bogner Péter CT - computed tomography Godfrey N. Hounsfield Allan M. Cormack The Nobel Prize in Physiology or Medicine 1979 MRI - magnetic resonance imaging Sir Peter Mansfield
CT/MRI képalkotás alapjai Prof. Bogner Péter CT - computed tomography Godfrey N. Hounsfield Allan M. Cormack The Nobel Prize in Physiology or Medicine 1979 MRI - magnetic resonance imaging Sir Peter Mansfield
Szabályozásbiológia EA
 Szabályozásbiológia EA Biológia MSc, 1. félév előadók: Schlett Katalin, Dobolyi Árpád, Kiss János, Világi Ildikó, Détári László Élettani és Neurobiológiai Tanszék csütörtök, 10:15 12:45 Szabályozásbiológia
Szabályozásbiológia EA Biológia MSc, 1. félév előadók: Schlett Katalin, Dobolyi Árpád, Kiss János, Világi Ildikó, Détári László Élettani és Neurobiológiai Tanszék csütörtök, 10:15 12:45 Szabályozásbiológia
Hiperlipidémia okozta neurodegeneratív és vér-agy gát-elváltozások ApoB-100 transzgenikus egerekben
 Hiperlipidémia okozta neurodegeneratív és vér-agy gát-elváltozások ApoB-100 transzgenikus egerekben Lénárt Nikolett Doktori (Ph. D.) értekezés tézisei Témavezető: Dr. Sántha Miklós tudományos főmunkatárs
Hiperlipidémia okozta neurodegeneratív és vér-agy gát-elváltozások ApoB-100 transzgenikus egerekben Lénárt Nikolett Doktori (Ph. D.) értekezés tézisei Témavezető: Dr. Sántha Miklós tudományos főmunkatárs
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Nem vezikuláris release. Kapcsoltság
 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Suppl. Materials. Polyhydroxyalkanoate (PHA) Granules Have no Phospholipids. Germany
 Granules Have no Phospholipids. Germany") Suppl. Materials Polyhydroxyalkanoate (PHA) Granules Have no Phospholipids Stephanie Bresan 1, Anna Sznajder 1, Waldemar Hauf 2, Karl Forchhammer 2, Daniel Pfeiffer 3 and Dieter Jendrossek 1 1 Institute
Suppl. Materials Polyhydroxyalkanoate (PHA) Granules Have no Phospholipids Stephanie Bresan 1, Anna Sznajder 1, Waldemar Hauf 2, Karl Forchhammer 2, Daniel Pfeiffer 3 and Dieter Jendrossek 1 1 Institute
A citokin egyensúly. Gyulladásgátló cytokinek. Gyulladáskeltő citokinek. Védelem és sejttúlélés. Gyulladás, sejtpusztulás NA DA.
 Apoptózis; Asztrocita: fagocitózis; PGD4; Növekedési faktorok: TGFβ1; neurosteroidok Gyulladásgátló cytokinek A citokin egyensúly NEUROTRANSZMITTEREK Reaktív asztroglia, aktivált mikroglia; Perifériás
Apoptózis; Asztrocita: fagocitózis; PGD4; Növekedési faktorok: TGFβ1; neurosteroidok Gyulladásgátló cytokinek A citokin egyensúly NEUROTRANSZMITTEREK Reaktív asztroglia, aktivált mikroglia; Perifériás
Endokrinológia. Közös jellemzők: nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások. váladékuk a hormon
 Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
Közös jellemzők: Endokrinológia nincs kivezetőcső, nincs végkamra - hámsejt csoportosulások váladékuk a hormon váladékukat a vér szállítja el - bő vérellátás távoli szervekre fejtik ki hatásukat (legtöbbször)
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Mikrogliák eredete és differenciációja
 Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Mikrogliák eredete és differenciációja 2017. 10. 24. Jordán Viktória F. Ginhoux et al. Origin and differentiation of microglia, 2013 F. Ginhoux et al. Fate mapping anaylsis reveals that adult microglia
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák. Nem vezikuláris release
 I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz mitter/neuro
I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz mitter/neuro
A rosszindulatú daganatos halálozás változása 1975 és 2001 között Magyarországon
 A rosszindulatú daganatos halálozás változása és között Eredeti közlemény Gaudi István 1,2, Kásler Miklós 2 1 MTA Számítástechnikai és Automatizálási Kutató Intézete, Budapest 2 Országos Onkológiai Intézet,
A rosszindulatú daganatos halálozás változása és között Eredeti közlemény Gaudi István 1,2, Kásler Miklós 2 1 MTA Számítástechnikai és Automatizálási Kutató Intézete, Budapest 2 Országos Onkológiai Intézet,
Az idegi működés strukturális és sejtes alapjai
 Az idegi működés strukturális és sejtes alapjai Élettani és Neurobiológiai Tanszék MTA-ELTE NAP B Idegi Sejtbiológiai Kutatócsoport Schlett Katalin a kurzus anyaga elérhető: http://physiology.elte.hu/agykutatas.html
Az idegi működés strukturális és sejtes alapjai Élettani és Neurobiológiai Tanszék MTA-ELTE NAP B Idegi Sejtbiológiai Kutatócsoport Schlett Katalin a kurzus anyaga elérhető: http://physiology.elte.hu/agykutatas.html
Azobezitás és a sejtek metabolizmusának összefüggései, a diabetes és táplálkozás viszonya
 Azobezitás és a sejtek metabolizmusának összefüggései, a diabetes és táplálkozás viszonya Máthé Endre, Sipos Péter, Remenyik Judit, Vígh Szabolcs, Horváth Brigitta Debreceni Egyetem, Mezőgazdaság, Élelmiszertudományi
Azobezitás és a sejtek metabolizmusának összefüggései, a diabetes és táplálkozás viszonya Máthé Endre, Sipos Péter, Remenyik Judit, Vígh Szabolcs, Horváth Brigitta Debreceni Egyetem, Mezőgazdaság, Élelmiszertudományi
A posztszinapszis és a PSD (posztszinaptikus. szinaptikus plaszticitásban játszott szerepük
 A posztszinapszis és a PSD (posztszinaptikus denzitás), valamint a szinaptikus plaszticitásban játszott szerepük A posztszinapszis szimmetrikus (Gray II): variábilis, nagy vezikulák; ált. gátló aszimmetrikus
A posztszinapszis és a PSD (posztszinaptikus denzitás), valamint a szinaptikus plaszticitásban játszott szerepük A posztszinapszis szimmetrikus (Gray II): variábilis, nagy vezikulák; ált. gátló aszimmetrikus
EEG Alvás Napszaki ritmusok
 EEG Alvás Napszaki ritmusok ALVÁS ÉS ÁLOM Az alvás a fiziológiás tudatvesztés periodikusan és reverzibilisen fellépő állapota, melyet jellemző testi, vegetatív és pszichés jelek kísérnek. Az álom az
EEG Alvás Napszaki ritmusok ALVÁS ÉS ÁLOM Az alvás a fiziológiás tudatvesztés periodikusan és reverzibilisen fellépő állapota, melyet jellemző testi, vegetatív és pszichés jelek kísérnek. Az álom az