A CHRONICUS LYMPHOCYTÁS LEUKAEMIA ÉS A DIFFÚZ NAGY B-SEJTES LYMPHOMA IMMUNGLOBULIN NEHÉZLÁNC GÉNJEINEK MUTÁCIÓS STÁTUSZA RICHTER SZINDRÓMÁBAN
|
|
|
- Ildikó Boros
- 7 évvel ezelőtt
- Látták:
Átírás

1 A CHRONICUS LYMPHOCYTÁS LEUKAEMIA ÉS A DIFFÚZ NAGY B-SEJTES LYMPHOMA IMMUNGLOBULIN NEHÉZLÁNC GÉNJEINEK MUTÁCIÓS STÁTUSZA RICHTER SZINDRÓMÁBAN Doktori értekezés Dr. Timár Botond Semmelweis Egyetem Patológiai Tudományok Doktori Iskola Témavezető: Dr. Matolcsy András egyetemi tanár, MTA doktora Hivatalos bírálók: Dr. Magyarosy Edina egyetemi docens, Ph.D. Dr. Sápi Zoltán főorvos, orvostud. kandidátusa Szigorlati bizottság elnöke: Szigorlati bizottság tagjai: Dr. Demeter Judit egyetemi tanár, MTA doktora Dr. Simon Károly osztályvezető főorvos, orvostud. kandidátusa Dr. Várkonyi Judit egyetemi adjunktus, Ph.D. Budapest 2006.
2 TARTALOMJEGYZÉK I. Összefoglalás 4 II. Summary 5 III. Bevezetés 6 III.1. A chronicus lymphocytás leukaemia 6 III.1.1 A B-CLL morfológiája 7 III.1.2 A B-CLL immunfenotípusa és genetikája 10 III.1.3. A B-CLL sejteredete 11 III Az immunglobulin (Ig) gének szerveződése 11 III Az IgH génátrendeződés 12 III A B-sejtek fejlődési fázisai 14 III A B-CLL sejteredete 16 III.2 A Richter szindróma 19 III.2.1 A lymphoma progresszió 19 III.2.2 A Richter szindróma 19 III.2.3 A diffúz nagy B-sejtes lymphoma 20 III.2.4 A Richter szindróma gyakorisága, klinikuma 21 III.2.5 A Richter szindróma morfológiája és immunfenotípusa 21 III.2.6 A Richter szindrómában előforduló genetikai változások 22 III.2.7 A B-CLL és a Richter szindróma klonális kapcsolata 24 IV. Célkitűzés 28 V. Anyag és módszer 29 V.1 Szövetminták 29 V.2 DNS izolálás 29 V.3 Az IgV H CDR3 régiójának PCR vizsgálata 31 V.4 Az IgV H gének PCR amplifikációja, klónozása és szekvenálása 32 VI. Eredmények 36 VI.1 Klonális kapcsolat a CLL és a kialakuló DLBCL minták között 36 VI.2 A mintákban előforduló V H géncsaládok 37 VI.3 Az IgV H gének szomatikus mutációi 38 VI.4 A szomatikus mutációk elemzése
3 VII. Megbeszélés 44 VIII. Eredeti megállapítások 49 IX. Rövidítések jegyzéke 50 X. Irodalomjegyzék 53 XI. Saját közlemények jegyzéke 66 XII. Köszönetnyilvánítás
4 I. ÖSSZEFOGLALÁS A chronicus lymphocytás leukaemia (CLL) indolens lefolyású lymphoproliferatív kórkép. Az esetek egy részében a megbetegedés klinikai és morfológiai progressziót mutat diffúz nagy B-sejtes lymphomába (DLBCL), amit Richter szindrómaként ismerünk. Az immunglobulin nehézlánc gének (IgV H ) mutációs státuszának prognosztikai szerepe van a CLL-ben. Azoknál a betegeknél, akik mutált IgV H gént hordoznak, a betegség jobb prognózisú, míg azon betegek esetében akiknek az IgV H génjük nem mutált, a betegség sokkal agresszívebb lefolyású. Azon CLL-ek mutációs státusza, amelyek DLBCL-be transzformálódnak, nem ismert. Tanulmányunkban ezért azt vizsgáltuk, hogy a Richter szindróma a mutált vagy a nem mutált IgV H génekkel rendelkező CLL-ből alakul-e ki. Nyolc eset mintapárjain végeztük el az IgV H gének mutációs státuszának elemzését. A CLL-es és a DLBCL-es tumorklónok öt esetben azonosnak és három esetben különbözőnek bizonyultak. Hat CLL-es minta nem mutált, két minta mutált IgV H gént hordozott. A hat nem mutált CLL esetből ötben a DLBCL-ek IgV H génje sem tartalmazott szomatikus hipermutációt és a DLBCL klónok a CLL-es tumorklónokból származtak. Egy nem mutált és két mutált CLL-es esetben a DLBCL mutált IgV H -t hordozott, de ebben a három esetben a tumorklónok nem egyeztek meg egymással, ami független, második tumor létrejöttét jelenti. Ezek alapján kijelenthetjük, hogy a klinikai lefolyás szerint egységes Richter szindróma keletkezési mechanizmusa alapján két úton, transzformációval és de novo második tumorként is kialakulhat. A CLL-ek DLBCL-be való transzformációja azonban csak a nem mutált altípusba tartozó CLL-ekben lép fel
5 II. SUMMARY Patients with chronic lymphocytic leukemia (CLL) may develop diffuse large B- cell lymphoma (DLBCL), also known as Richter s syndrome. Mutational status of immunoglobulin heavy chain variable region (IgV H ) genes have prognostic impact in CLL. Patients with mutated IgV H genes have stable disease, whereas patients with unmutated IgV H gene have more aggressive disease. The mutational status of CLLs that transform to DLBCL is unknown. To reveal whether Richter s syndrome occurs in CLLs with mutated or unmutated IgV H genes, we have performed mutational analysis on serial specimens from eight patients. CLL and DLBCL tumorclones were identical in five cases and they were different in three cases. Six CLL expressed unmutated and two cases expressed mutated IgV H genes. In five of the six unmutated CLLs, the DLBCL clones evolved from CLL tumorclones and the IgV H genes expressed by DLBCLs were also unmutated. In one unmutated and two mutated CLLs, the DLBCL expressed mutated IgV H genes, but in these three cases the DLBCL tumorclones developed as independent secondary neoplasm. These result suggest that Richter s syndrome may develop in both mutated or unmutated CLLs, but clonal transformation of CLL to DLBCL occur only in the unmutated subgroup of CLL
6 III. BEVEZETÉS III.1. A CHRONICUS LYMPHOCYTÁS LEUKAEMIA A chronicus lymphocytás leukaemia (CLL)/ kis lymphocytás lymphoma (SLL) klonális lymphoproliferatív megbetegedés, amelynek az incidenciája 3/ , és amely az összes non-hodgkin lymphoma (NHL) 6,7 %-át alkotja. 2 Megjelenésekor az átlagos életkor év, a betegség férfiaknál kétszer gyakoribb, mint nőknél. A 65 év feletti felnőtt lakosságban előforduló leukaemiák 40%-át teszi ki, a betegség az esetek 20-30%-ában az 55 év alatti populációban jelenik meg, ugyanakkor a 30 év alattiakban nagyon ritka. 1 A betegség klinikai lefolyása heterogén. A betegek többsége hosszabb-rövidebb ideig is tünetmentes lehet, és csak később jelentkeznek az általában aspecifikus tünetek, mint például gyengeség, fáradékonyság, súlyvesztés, jellemző lehet a splenomegalia, hepatomegalia, lymphadenopathia is. 3, 4 Vannak esetek, ahol a hosszú tünetmentes túléléshez sem kell terápiás beavatkozás, de vannak olyanok is, ahol az agresszív kezelés ellenére sem sikerül biztosítani a betegek túlélését. 5 A CLL diagnózisakor a betegekben a csontvelő lymphocytás infiltrációja és a perifériás vér abszolút lymphocytosisa (több mint 5000 érettnek tűnő lymphocyta/μl) figyelhető meg (B-CLL), az esetek kisebb részében csak a nyirokcsomók érintettek (B-SLL). A betegség ezen két megjelenési formája sem klinikailag, sem prognosztikailag nem különbözik egymástól



 (B) A perifériás kenetben azonosítható károsodott lymphocyták (Grumprecht-rögök, nyíl) (Giemsa, 400X).")
7 III.1.1 A B-CLL morfológiája A B-CLL/SLL az érett B-sejtek daganatos betegsége. A B-CLL-es betegek vérében és csontvelejében is kis, heterokromatinizált magvú, keskeny, enyhén basophil cytoplasmával rendelkező lymphocyták szaporodnak fel (1A. ábra). A vérből készült kenetben a daganatos sejtek mellett gyakran megfigyelhetők a szétesett sejteknek megfelelő felhők, az úgynevezett Grumprecht rögök, amelyek a kenetkészítés során széteső, fizikai behatásra érzékeny tumorsejtek maradványai (1B. ábra). A B 1. ábra CLL megjelenése perifériás kenetben. (A) Heterokromatinizált magvú, keskeny, enyhén basophil cytoplasmával rendelkező lymphocyták a beteg vérében. (Giemsa, 1000X) (B) A perifériás kenetben azonosítható károsodott lymphocyták (Grumprecht-rögök, nyíl) (Giemsa, 400X). CLL-ben a prolymphocyták száma kevesebb mint 3%. A 10% feletti prolymphocyta-arány a kórkép agresszívebb klinikai lefolyású variánsát jelzi, amelyet krónikus lymphocytás leukémia / prolymphocytás lymphoma (CLL/PLL) diagnózissal jelölnek. 7 A csontvelői infiltráció általában noduláris, intersticiális vagy diffúz jellegű, de a csontvelő peritrabecularis tereit nem érinti (2. ábra)


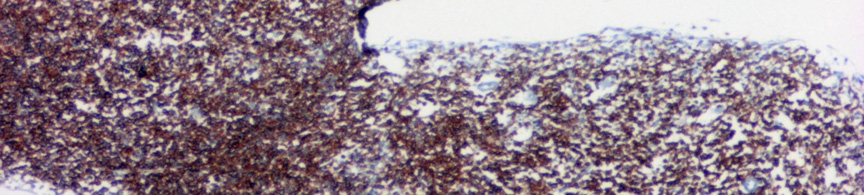
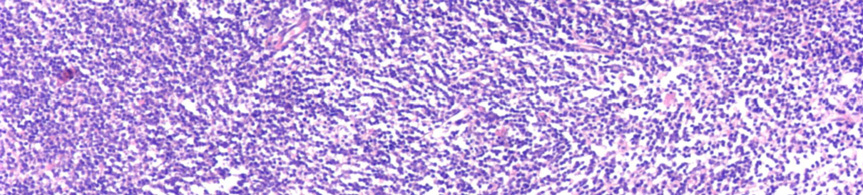

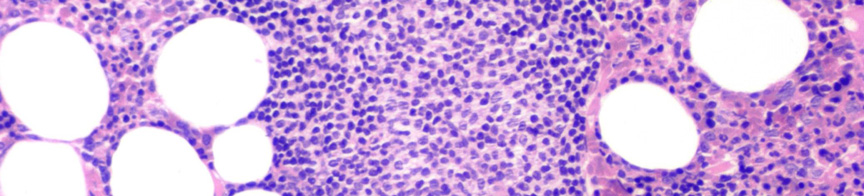

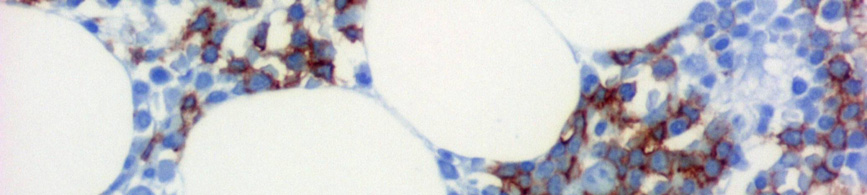
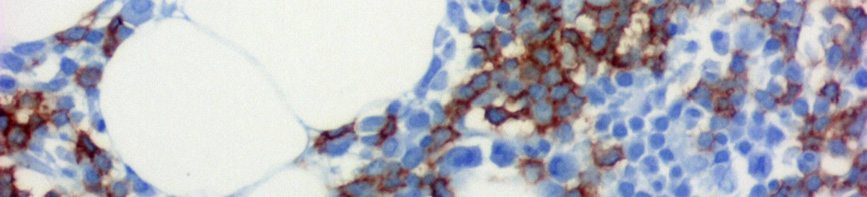



8 A B C D E F 2. ábra CLL szövettani megjelenése csontvelőben. (A) Kis, kerek magvú, keskeny cytoplasmájú sejtek diffúzan infiltrálják a csontvelőt (HE, 40X). (B) Az immuhisztokémiával láthatóvá válnak CD20+ infiltráló sejtek (CD20 immunhisztokémia, 40X). (C-D) Hasonló módon tehető láthatóvá a noduláris (HE, 40X; CD20, 40X) és az (E-F) intersticiális (HE 100X; CD20 100X) típusú infiltráció is. -8-

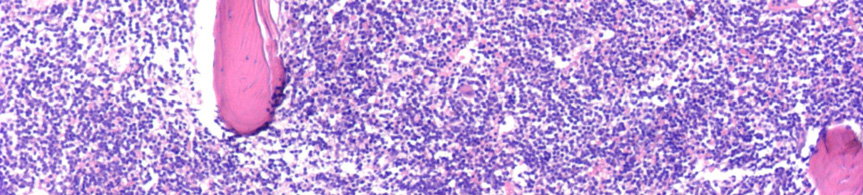
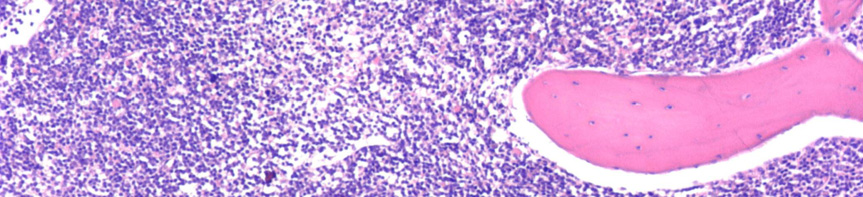
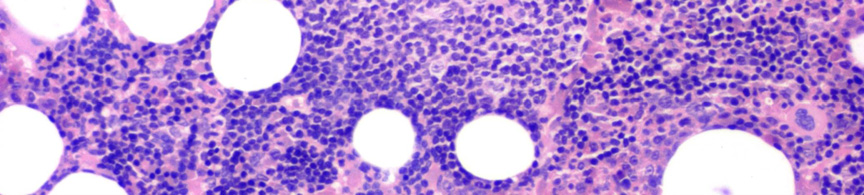
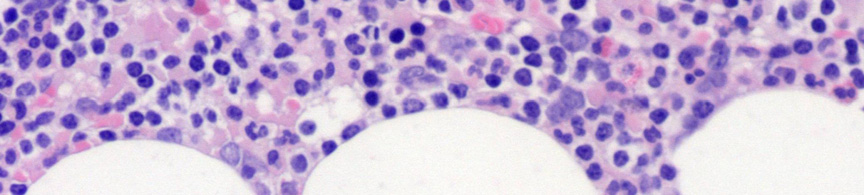
9 Nyirokcsomó érintettség esetén annak follicularis alapszerkezete felbomlik. Állományában a sötétebb, kisebb sejteket tartalmazó háttér mellett nagyobb sejteket tartalmazó világosabb területek, úgynevezett pseudofolliculusok vannak jelen. 6 Az infiltrátumban a kis 6-12 μm átmérőjű keskeny cytoplasmájú lymphocyták dominálnak, amelyeknek kerek, heterokromatinizált magjuk van 7 (3A. ábra). A mitotikus aktivitás általában nagyon alacsony. A pseudofolliculusok, vagy más néven proliferációs centrumok, kis, középnagy és nagy sejtekből állnak. A közepes méretű sejtek a prolymphocyták, melyek laza kromatinnal és kis nucleolusszal rendelkeznek. A paraimmunoblastok középnagy-nagy sejtek, amelyeknek kerek vagy ovális sejtmagjuk, centrálisan elhelyezkedő eosinophil nucleolusuk és enyhén basophil cytoplasmájuk van 8, 9 (3B. ábra). A lépben a betegség a fehér pulpában van jelen, de a folyamat előrehaladtával az infiltrátum diffúzzá válhat. 10 A B 3. ábra CLL szövettani megjelenése nyirokcsomóban és kenetben. (A) A nagyobb sejteket tartalmazó világosabb területek (pseudofolliculusok) a nyirokcsomó felbomlott állományában (HE, 40X). (B) A pseudofolliculuson belül nagyobb prolymphocyták, immunoblastok és paraimmunoblastok láthatóak a kis, kerek, tömött kromatinnal rendelkező sejtek mellett. (HE, 200X) - 9 -
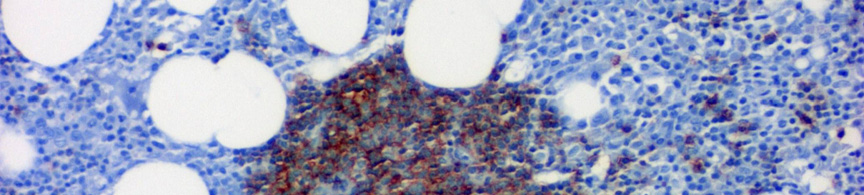
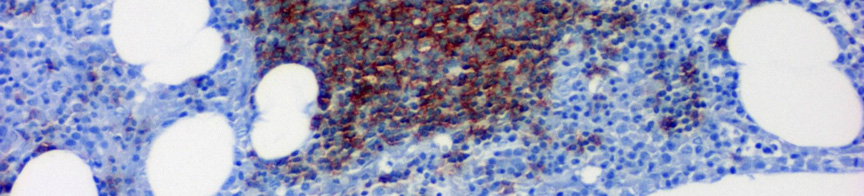
10 III.1.2 A B-CLL immunfenotípusa és genetikája A tumorsejtek a B-sejtes markerek közül a CD19, CD20, CD23-at expresszálják a T-sejtekre jellemző CD5-tel együtt. Alacsony intenzitással fejezik ki a felszíni IgM-et és/vagy IgD-t. A tumorsejtek CD10, Cyclin D1, CD22 és FMC7 negatívak (4. ábra) ábra CLL kimutatása áramláscitometria segítségével. A lymphocyták CD19-CD23-CD5 koexpressziót mutatnak lambda könnyűlánc izotípusexklúzió mellett, amely a CLL jellemzője. Az immunglobulin könnyű- és nehézláncok klonális átrendeződést mutatnak. Az immunglobulin nehézlánc variábilis régiójának (IgV H ) génjei 40-50%-ban nem hordoznak szomatikus mutációkat, 50-60%-ban azonban az IgV H gének szomatikus hipermutációja figyelhető meg, amivel jól korrelál a ZAP70 és a CD38 markerek expressziója (nem mutált IgV H esetén a CD38 és a ZAP70 emelkedett). Fluoreszcens in situ hibridizációval (FISH) az esetek 80%-ában mutatható ki valamilyen abnormális kariotípus 15 (5.ábra). A 12-es kromoszóma triszómiája az esetek 20%-ában, a 13q14 deléció az esetek 50%-ában fordul elő. 16, 17 Míg a 12-es triszómiás esetek nagy részében

 A 12-es kromoszóma triszómiája (három zöld centroméra-specifikus jel látható a normális kettő helyett). (D) A 17p13 régió deléciója. III.1.3. A B-CLL sejteredete III.")


11 az IgV H nem mutált, addig a 13q14 deléciót hordozó esetekben gyakrabban írtak le mutációt 18. A 11q22-23 deléció 20%-ban, a 6q21 5%-ban és a 17p13 az esetek 10%-ában jelenik meg. 19 A B C D ábra CLL citogenetikai eltérései. (A) A 11q22 régió deléciója FISH (2 piros jel helyett csak 1 látható) és metafázis kromoszóma vizsgálattal. (B) 13q14 deléció. (C) A 12-es kromoszóma triszómiája (három zöld centroméra-specifikus jel látható a normális kettő helyett). (D) A 17p13 régió deléciója. III.1.3. A B-CLL sejteredete III Az immunglobulin (Ig) gének szerveződése Az Ig nehézlánc-gének (IgH) a 14-es kromoszómán, a könnyűlánc-gének (IgL) közül a κ könnyűlánc a 2-es, a λ könnyűlánc a 22-es kromoszómán található. A konstans κ-lánc domént (C κ ) egyetlen exon kódolja, míg a variábilis domént a nagy számban jelenlévő V (variábilis) és J ( joining ) szegmentumok. A mintegy 40 V κ gén egyike a V-domén N-terminális szekvenciájának nagyobbik részét, míg a fennmaradó részt az 5 J κ szegmentum valamelyike kódolja (6A. ábra). A λ-láncokat kódoló gének 3 szegmentumba rendeződnek: 30 V λ, 4 funkcionális C λ, és mindegyik C λ mellett egy
 sorolható (6C. ábra). 6. ábra Az immunglobulin könnyű- (A, B) és nehézlánc-gének (C) szerveződése.")
 található elrendeződésben tartalmazza az Ig-géneket. A B-sejt érés során megindul az Ig-gének átrendeződése.")


12 egy J λ szegmentum található (6B. ábra). A nehézlánc variábilis doménjét kódoló ~40 V H, 25 D (diverzitás) és 6 J H szegmentum egy csoportban helyezkedik el. 20 A V H szegmentumot alkotó kb. 40 funkcionális gén a szekvenciáik homológiája alapján 6 géncsaládba (V H 1, V H 2,... V H 6) sorolható (6C. ábra). 6. ábra Az immunglobulin könnyű- (A, B) és nehézlánc-gének (C) szerveződése. III Az IgH génátrendeződés A B-sejtek fejlődése során az Ig-gének előbb említett sorrendje a gének jellegzetes átrendeződése miatt megváltozik. A B-sejtek kivételével valamennyi sejt a csíravonalban (germline) található elrendeződésben tartalmazza az Ig-géneket. A B-sejt érés során megindul az Ig-gének átrendeződése. A folyamat első szakaszában egy véletlenszerűen kiválasztott D és egy J H gén kerül egymás mellé, majd az így kialakult D-J H szakaszhoz egy V H gén kapcsolódik. E szomatikus rekombináció során az egymás mellé kerülő szegmentumok közötti DNS-szakaszok kiesnek és létrejön a funkcionális V H DJ H génszegmentum. Az átrendeződött IgH génkonfiguráció az adott B-sejtre jellemző és végigkíséri a B-sejt fejlődés folyamatát. A V H -szegmentum kódolja a variábilis domén N-terminálisának nagy részét. A variábilis doménen belül, a vázszekvenciák között ( framework régió FR) hipervariábilis régiók helyezkednek el (komplementaritást meghatározó régiók CDR), amelyek az antigénkötő hely



 enzim által")
.")
13 kialakításában vesznek részt. Ezeket CDR1-, CDR2 és CDR3-nak nevezzük, amelyek közül legutóbbi mutatja a legnagyobb variabilitást. A CDR3 variabilitását az átrendeződő génszakaszok pontatlan kapcsolódása (kapcsolódási diverzitás) és az ún. N-szekvenciáknak nevezett, a terminális deoxiribonukleotidil-transzferáz (TdT) enzim által véletlenszerűen beépített addicionális nukleotidok fokozzák (N-régió-diverzitás). A variábilis domént konstans domének követik (C H ) 20 (7. ábra). 7. ábra Az IgH génátrendeződés sematikus ábrája
14 III A B-sejtek fejlődési fázisai Az a fejlődési folyamat, amelynek során a haemopoeticus őssejtek a B-lymphocyta-érés különböző fázisain keresztül ellenanyagokat termelő plazmasejtekké vagy memória sejtekké alakulnak, jellegzetes és jól karakterizálható szakaszokban zajlik (8. ábra). A szakaszokat az immunglobulin génátrendeződés és az immunglobulin nehéz- és könnyűláncok expressziója határozza meg. A legkorábbi B-sejt irányba elkötelezett sejt a progenitor tulajdonságokkal rendelkező pro-b-sejt. Ezekben a sejtekben történik az IgH gének átrendeződése. A korai pro-b-sejtekben a D-J H szakaszok, majd a késői pro-b-sejtekben a V H -DJ H szakaszok rendeződnek át. A produktív V H DJ H átrendeződés következménye a sejt μ-lánc termelése, ami már a következő fejlődési stádiumban levő pre-b-sejtekre jellemző. A μ-lánc a nagy pre-b-sejtekben termelődik és nagy része intracytoplasmatikusan található meg, egy része azonban a felszínen is megjelenik és a pót -L-lánccal (surrogate light-chain) együtt a pre-b-sejt receptort (pre-bcr) hozzák létre. A pre-bcr-en keresztül közvetített hatások eredményeképp a B-sejt befejezi az IgH-lókusz génjeinek további átrendeződését és néhány osztódás után megindítja az IgL-génátrendeződést, ami immár a kis pre-b-sejtekben zajlik. Amint létrejön egy működő könnyűlánc, a sejt teljes értékű IgM (BcR) komplexet expresszál a felszínén. A teljes értékű receptor viszont olyan jeleket közvetít, amelyek az átrendeződést vezérlő RAG-1, RAG-2 gének represszióját, következésképp a VDJ rekombináz aktivitás teljes megszűnését eredményezik. Ebben a stádiumban a sejteket éretlen B-sejteknek nevezzük. A fejlődés eddig a pontig a csontvelőben, antigéntől függetlenül zajlik. Ennél a pontnál történik a B-sejtek szelekciója is, amelynek eredményeképpen létrejön a nem-saját antigének felismerésére alkalmas, felszíni IgM és IgD molekulát hordozó érett B-sejt készlet 20 (8.ábra)
15 8.ábra A B-sejtek fejlődési fázisai. SHM: szomatikus hipermutáció, GC: centrum germinatívum, H-lánc: nehézlánc, L-lánc: könnyűlánc
16 Az érett B-sejtek elhagyják a csontvelőt és a perifériás nyirokszervekbe jutnak, ahol immár antigéntől függő módon tovább differenciálódnak. Az antigénnel való kapcsolat megfelelő specificitású B-sejtek klonális szelekcióját indukálja. Ezek a sejtek (centroblastok) jellegzetes morfológiai és funkcionális változáson mennek keresztül, amelynek módjai a következők: - Nehézlánc-izotípusváltás. Megfelelő hatásokra a felszíni IgM és IgD expressziót más izotípusba tartozó nehézláncokat tartalmazó Ig molekulák termelése váltja fel. - Affinitásérés. Az immunglobulin variábilis régiókban bekövetkező szomatikus hipermutációk (SHM), amelyeket az aktiváció-indukált citidin deamináz (AID) fehérje vezérel és aminek következtében az érett B-sejt receptorának antigén iránti affinitása fokozódik. A centroblastokból ezek után centrocyták lesznek, amelyek, ha megfelelő antigénnel találkoznak proliferálni kezdenek, míg antigénstimulus hiányában néhány napon belül elpusztulnak. Ez leggyakrabban a T-sejtektől függő folyamat és a nyirokcsomók centrum germinatívumaiban következik be. 21 Ebben az esetben mindig létrejön a variábilis régió szomatikus hipermutációja. Az érési folyamat azonban T-sejt független módon a centrum germinatívumokon kívül a marginális zónában is létrejöhet. Ilyenkor a további érésnek nem feltétele a variábilis régió szomatikus hipermutációja A B-sejt-érés utolsó fázisaként mindkét folyamat eredményeképpen memória B-sejt és ellenanyag termelő plazmasejt alakul ki (9. ábra). III A B-CLL sejteredete A B-sejtes lymphomák csoportjába olyan betegség típusok tartoznak, amelyek a különböző mértékben differenciált B-sejtek daganatos megbetegedései. A lymphomás transzformáció a progenitor B-sejtektől a plazmasejtes differenciációt elérő B-sejtekig terjedő érési stádiumok közül bárhol előfordulhat. Az esetek többségében a tumorsejtek morfológiai megjelenésükben, fenotípus és genotípus jellegzetességeikben hasonlítanak a B-sejt-differenciáció egyes érési stádiumaira, illetve a normális nyirokszövetet felépítő B-sejt-populációkra
17 BN: Nyugvó naiv BA: Aktivált naiv CB: Centroblast CC: Centrocyta PC: Plazmasejt BM: Memória sejt T: T-sejt FDC: Follicularis dendriticus sejt Perifériás vér és T-sejt zóna BN BN T BN BA BA CB CB BA CC Sötét zóna Köpenyzóna Bazális világos zóna Apikális világos zóna Marginális zóna 9. ábra A nyirokcsomó folliculusban történő B-sejt érés sematikus ábrája. A folliculus több zónára osztható, amelyekben az érési folyamatok zajlanak. A B-sejtek antigéntől függő érése a másodlagos nyirokszervekben zajlik, ahol a T-helper-sejtektől kapják meg a további fejlődéshez szükséges ingereket. A folyamat azonban T-sejt független módon is létrejöhet. PC Klonális expanzió Apoptosis Differenciáció Szomatikus hipermutáció Izotípusváltás T CC FD PC BM BM Korábban azt tartották, hogy a CLL-es sejtek a follicularis köpenyzóna B1-sejtjeiből keletkeznek, amelyekre jellemző, hogy a sejtek CD5, CD23 pozitívak, CD38 negatívak, valamint felszíni IgM-et és IgD-t expresszálnak és nem mutált IgV H génekkel rendelkeznek. Ezt azzal magyarázták, hogy a sejtek a centrum germinatívumban (GC) még nem találkoztak antigénnel A korai vizsgálatok a V H géneket csíravonal konfigurációjúnak találták 30-34, ami úgy tűnt, hogy alátámasztja a naiv B-sejtes eredetet. Ez az elképzelés sokáig tarotta magát, azonban később az irodalomban megjelentek olyan munkák, ahol azt bizonyították, hogy az esetek 50-60%-ában az IgV H gének szomatikusan hipermutáltak, ami a centrum germinatívumon már áthaladt B-sejtekre jellemző. Az, hogy GC vagy post-gc stádiumot képvisel az adott tumorsejt, az ún. intraklonális diverzitás, vagyis a mutációk kontinuus ( ongoing ) jellegének vizsgálatával állapítható meg. Ez azt jelenti, hogy
18 a neoplasticussá vált sejtek megőrzik hipermutációs aktivitásukat és folyamatosan újabb mutációkat szereznek, ami a tumorklónok genetikailag instabil állapotát jelenti. Az adatok szerint a CLL-ek biológiailag és klinikailag két különböző csoportra oszthatóak, az egyikre az IgV H gének csíravonal expressziója jellemző és rosszabb klinikai prognózist jelent, a másikra pedig az IgV H gének szomatikus hipermutációja jellemző és a klinikailag kedvezőbb csoportba tartozik. 18, Frissebb tanulmányok szerint a CLL-re egy közös génexpressziós mintázat jellemző, amely leginkább a memória B-sejtekére hasonlít és az IgV H genotípustól független, jóllehet az IgV H régióban mutált és nem mutált esetek egy 30 génből álló expressziós panellel 40, 41 elkülöníthetőek. Fenotípus tekintetében az aktivációs markerek (CD23, CD69, CD71), valamint a memória sejtekre jellemző CD27 szintén a posztgerminális eredetet valószínűsíti. Ez ugyan nehezen egyeztethető össze azokkal az esetekkel, amelyeknél az IgV H szomatikus mutációja hiányzik. Ez az ellentmondás feloldható, ha feltételezzük, hogy a naiv B-sejtek a nyiroktüszőkön kívül T-independens antigénekkel, vagy szuperantigénekkel találkoznak, amelyek a sejtfelszíni immunglobulinhoz (BcR) nem az antigén-felismerő régiónál, hanem a framework 39, 42 régiónál kötődnek; így elmarad a IgV H szomatikus mutációin alapuló affinitás-érés. A CLL-ben leggyakrabban átrendeződő IgV H gének (V H 1-69, V H 3-7 és a V H 4-34) előfordulása is lényegesen magasabb, mint ami véletlenszerűen várható lenne. Tehát sejteredet szempontjából a CLL sejtek a legújabb kutatások alapján egységesek, 43, 44 mégpedig érett, antigénszelekción átesett B-sejtekből alakulnak ki
19 III.2. A RICHTER SZINDRÓMA III.2.1 A lymphoma progresszió A szolid tumorok kialakulásához hasonlóan a lymphomák létrejöttét is többlépcsős folyamatnak tartják, ahol a genetikai változások a lymphoid sejtek autonóm növekedését okozzák az érési folyamat különböző fázisaiban. Klinikai megnyilvánulása ennek a folyamatnak lehet lymphoma és/vagy leukaemia. Az új lymphoma klasszifikációk, amelyek egy-egy lymphoma entitás morfológiai, genetikai és klinikai tulajdonságait is figyelembe veszik, teret nyújtottak azoknak a szemléleteknek, amelyek dinamikus biológiai és morfológiai heterogenitásról beszélnek egyetlen lymphoma entitáson belül is. A lymphomagenezis többlépcsős modelljének megfelelően a lymphomák gyakran válnak agresszív fenotípusúvá a lefolyásuk során, amit lymphoma progressziónak nevezünk Habár a lymphoma progresszió morfológiai, klinikai és biológiai aspektusai nincsenek mindig átfedésben, a lymphoma morfológiájában történő változások gyakran együtt járnak a klinikai és biológiai viselkedésben történő változással. Egy progressziót mutató lymphoma eredeti és progrediált komponense közötti klonális kapcsolat esetén lymphoma transzformációról beszélünk. A progresszió azonban egy de novo másodlagosan kialakuló lymphomát is jelenthet, amely klonálisan független az azt megelőző komponenstől. Ezekben az esetekben kompozit lymphomáról beszélünk. III.2.2 A Richter szindróma A klinikailag indolens lefolyású alacsony malignitású B-CLL/SLL magas malignitású non-hodgkin lymphomába való progresszióját Richter szindrómának (RS) nevezzük. A szindrómát először Maurice N. Richter írta le 1928-ban, mint egy CLL-hez társuló, gyorsan progrediáló agresszív lymphomát, amely generalizált lymphadenopathiával és hepatosplenomegaliával jár ben Lortholary és munkatársai vezették be a "Richter szindróma" megjelölést azokra az esetekre, ahol szövettanilag a CLL-ből "malignus reticulopathia" alakul ki. 50 A későbbiekben kiterjesztették a Richter szindróma fogalmát olyan más
20 lymphoid malignitásokra is, amelyek a CLL-es páciensekben alakulnak ki. Ezekben az esetekben prolymphocytás leukaemia (PLL), kis nem hasadt sejtes lymphoma 51, lymphoblastos lymphoma 52, hajas sejtes leukaemia 53 és Hodgkin lymphoma (HL), amely a Richter transzformáció Hodgkin variánsaként ismert 54, alakulhat ki. Leggyakrabban azonban a progresszió során DLBCL alakul ki. III.2.3 A diffúz nagy B-sejtes lymphoma A diffúz nagy B-sejtes lymphoma (DLBCL) az érett B-sejtek tumoros megbetegedése. A felnőttkori NHL-ák 30-40%-át teszi ki. A DLBCL-ek mind morfológiailag, fenotípus és molekuláris tulajdonságok tekintetében valamint a klinikai viselkedés szempontjából is heterogén csoportot alkotnak. A tumor az esetek többségében a nyirokcsomóból vagy az extranodalis szövetekből indul ki és csak másodlagosan érinti a csontvelőt. Morfológiai megjelenésük alapján centroblastos, immunoblastos, multilobulált, T-sejt/histiocyta gazdag, plasmoblastos és anaplasticus variánsokra oszthatóak. 8 A génexpressziós vizsgálatok alapján két fő csoportot különítettek el az alapján, hogy a tumorsejtek az aktivált B-sejtek expressziós mintázatához (ABC-DLBCL), vagy a centrum germinatívum B-sejtjeinek (GCB-DLBCL) mintázatához hasonlítanak. Tanulmányok szerint klinikailag a GCB-DLBCL a betegség sokkal kedvezőbb lefolyását jelenti, míg az ABC-DLBCL sokkal rosszabb prognózissal jár. 55 A DLBCL kialakulhat de novo vagy megelőző alacsony malignitású lymphoma talaján (pl. CLL). A DLBCL tumorsejtjei az IgV H régióiban szomatikus mutációkat hordoznak, ami alapján két csoportra lehet őket osztani aszerint, hogy az ún. intraklonális divergencia kimutatható-e vagy sem. Tanulmányok szerint az esetek egy részében kimutatható a kontinuus ( ongoing ) jellegű mutáció, ami ezen sejtek GC eredetére utal. A többi esetben antigén szelektált B-sejtes eredetre (post-gc) jellemző mutációkat találtak Azonban egyik tanulmány sem említi, hogy a Richter szindróma DLBCL-jeiben milyen az IgV H gének mutációs profilja
21 III.2.4 A Richter szindróma gyakorisága, klinikuma A CLL-es betegekben a kialakuló RS gyakoriságát 3-10%-ra teszik, azonban a valós incidencia ennél nagyobb lehet, mivel post mortem ritkán történnek ez irányban vizsgálatok. 60 Egy 1011 B-CLL-es betegen végzett vizsgálat szerint a betegek 2,2%-ában alakult ki RS, ebből 1,8%-ban DLBCL és 0,4%-ban HL. Felfigyeltek arra is, hogy a fiatalabb ( 55 év) és az idősebb (>55év) betegek közül a fiatalabbakban az RS előfordulása magasabb (5,9% ill. 1,2%), aminek legvalószínűbb oka az, hogy az idősebb, kezelésre nem reagáló betegeknél kevesebbszer végeznek biopsziát. 61 Egy másik retrospektív vizsgálat során, ahol 1374 CLL-el diagnosztizált beteget kezeltek 1972 és 1992 között, 39 betegben (2,8%) észlelték a RS kialakulását. Egy harmadik tanulmány szerint között 2147 beteg követése során 105-ben (4,9%) figyeltek meg RS-t. A transzformáció incidenciája a sorozatos kezelések következtében nőtt meg. 62 Földrajzilag a B-CLL Európában és az Egyesült Államokban a leggyakoribb haematológiai megbetegedés, ezért a kialakuló RS-át is ezekben az országokban észlelik a legtöbbször. Más földrajzi területeken a pontos előfordulás nem ismert, de nagyon ritka, bár Japánban és Tajvanon is írtak már le eseteket. 63 A Richter szindrómát általános tünetek (láz, súlycsökkenés, éjszakai izzadás) megjelenése és az általános állapot gyors romlása jellemzi. Emellett nyirokcsomó megnagyobbodás, valamint hasi panaszokat okozó hepatomegalia és/vagy splenomegalia esetleg központi idegrendszeri érintettségre utaló jelek kísérik A leggyakoribb közös tulajdonság a RS-ás eseteknél az emelkedett LDH szint, amely a tumornövekedés egyik markere. Egy tanulmány szerint az RS esetek 82%-ában az LDH szint a normálisnak legalább kétszerese volt, míg ugyanezt a CLL-es esetek csupán 8%-ában találták. Paraproteinemiát a betegek 44%-ában figyeltek meg. 62 Természetesen a RS diagnózisának megállapításához nyirokcsomó biopszia szükséges. III.2.5 A Richter szindróma morfológiája és immunfenotípusa 1928-ban Richter a leírt eset szövettani vizsgálata során a nyirokcsomókban, a májban és a lépben leukaemiás és tumorsejteket egyaránt leírt. A leukaemiás sejteket kis méretű lymphocytaként, a tumorsejteket pedig számos, a lymphocytánál néhányszor
22 nagyobb, bő basophil cytoplasmával, határozott maggal és néhány prominens nucleolusszal rendelkező polymorph endotheloid sejtként azonosította. 49 Ez a szövettani kép a jelenlegi WHO osztályozás szerint a DLBCL immunoblastos variánsának (DLBCL-IB) felel meg 8 (10. ábra). Irodalmi adatok szerint a DLBCL centroblastos variánsa (DLBCL-CB) is előfordulhat. 68 Ebben az esetben a centroblastok diffúz, monoton proliferációját lehet látni, amibe esetenként immunoblastok is keveredhetnek. Az összes DLBCL közül az IB esetek aránya kb. 10% 69, de RS-ban ez az arány jóval magasabb. A RS immunfenotípusáról igen kevés irodalmi adat áll rendelkezésre. Bizonyos esetekben a RS megőrzi a megelőző B-CLL immunfenotípusát (IgM +, IgD +/-, CD5 +, CD19 +, CD23 + ) 70, 71, egyéb esetekben viszont a RS immunhisztokémiai megjelenése eltér a B-CLL-től (CD5 -, IgD - ) A proliferáló sejtek aránya a CLL-hez képest jelentősen megemelkedik. III.2.6 A Richter szindrómában előforduló genetikai változások A legtöbb indolens B-sejtes NHL a lymphomagenezis jelenleg elfogadott elvei szerint leginkább az apoptotikus folyamatok zavarával jellemezhető. 75 A lymphocyták hiperproliferációja az agresszív lymphomák velejárója, ami számos esetben a másodlagosan bekövetkező genetikai zavaroknak tudható be. 76 Ezek a másodlagos genetikai zavarok különböző alacsony malignitású B-sejtes NHL progresszióját egyaránt jellemezhetik, függetlenül azok típusától A jelenség független 68, 80 a kemoterápiától, kemoterápiában nem részesült betegekben is előfordulhat. A megfigyelt genetikai eltérések gyakran komplex módon fordulnak elő. A leírt eltérések közé tartozik a 12-es triszómia, 14q expanzió, a 11q23, 13q és a 17p régió deléciója. Citogenetikai vizsgálatok eredményei megerősítették azt a feltételezést, miszerint a B-CLL-ben detektálható komplex kariotípus-eltérések nagyobb valószínűséggel vezetnek RS-hoz, mint a kromoszomális eltérések hiánya, 62, vagy a 12. triszómia önmagában
 A CLL sejtjei keverednek a DLBCL nagyobb sejtjeivel (nyíl) (HE, 200X). (B) DLBCL-IB.")
.")
. (Giemsa, 1000X). A p53 gén mutációját először B-sejtes RS-ában írták le.")
23 A B C * * ** 10. ábra A Richter szindróma szövettani megjelenése nyirokcsomóban és perifériás kenetben. (A) A CLL sejtjei keverednek a DLBCL nagyobb sejtjeivel (nyíl) (HE, 200X). (B) DLBCL-IB. A csontvelőt infiltráló immunoblastok nagy, mérsékelten pleiomorph sejtek és centrálisan elhelyezkedő, erősen bazofil nagy nucleolusszal rendelkeznek (HE, 200X). (C) A kenetben a CLL lymphocytái (*) mellett nagy, közepesen széles cytoplasmájú, laza kromatin szerkezetű multiplex nucleolust tartalmazó centroblast sejtek láthatók (**). (Giemsa, 1000X). A p53 gén mutációját először B-sejtes RS-ában írták le. 84 Akkoriban azt gondolták, hogy a p53 gén mutációja lehet a kulcs az alacsony malignitású lymphoma magas malignitású lymphomába való progressziójához. Kutatási eredmények viszont azt mutatták, hogy a RS előfordulhat p53 mutációval illetve anélkül is, és hogy a p53 inaktiválódása már a B-CLL stádiumban is bekövetkezhet, így nem feltétlen velejárója
24 a RS-nak sem A p16 INK4A/ARF gén inaktiválódása is kapcsolatba hozható a RS kialakulásával. 87, 88 A p16 INK4A/ARF gátló hatással van a sejtciklusra azáltal, hogy a CDK4 és CDK6 enzimek regulatorikus alegységeihez kapcsolódva inaktiválja a ciklind/ CDK4/6 komplex hatását a G1/S fordulóponton. 89 A p16 INK4A/ARF inaktivációját a progrediált CLL, follicularis lymphoma (FL) és a köpenysejtes lymphoma blastos variánsának 33%-ában, azonban ezek indolens megfelelőjében csak 5%-ában észlelték. 90 p53 mutáció és/vagy p16 INK4A/ARF homozigóta deléció a RS-ás esetek 60%-ában fordul elő. 84, 85, 90, 91 A többszörös genetikai léziók a CLL transzformált stádiumában arra utalhatnak, hogy szerzett genetikai instabilitás van jelen genomszerte, ami a Richter szindróma kialakulását segíti. 92, 93 Munkacsoportunk egy korábbi tanulmányában azt a kérdést vizsgálta, hogy a B-CLL Richter transzformációja során szerepet tölt-e be a mikroszatellita instabilitás (MSI), és az instabilitás kialakulásának hátterében állhat-e a hmlh1 és hmsh2 mismatch repair gének esetleges genetikai alterációja. 19 CLL-es betegből 9 beteg volt RS-ás. Eredményeink arra utaltak, hogy a B-CLL Richter transzformációjának egyes eseteiben a hmlh1 gén epigenetikai inaktiválódása által kiváltott genetikai instabilitás hozzájárulhat a B-CLL progressziójához, ugyanakkor a hmlh1 és hmsh2 gének a folyamat során strukturálisan nem érintettek 92 III.2.7 A B-CLL és a Richter szindróma klonális kapcsolata A megelőző B-CLL-ből kialakuló DLBCL nem mindig az eredeti tumorklónból származik, de novo is kialakulhat. A molekuláris genetikai vizsgálómódszerek elterjedése előtt a klonalitást a B-sejtek sejtfelszíni immunglobulin könnyűlánc (IgL) immunfenotípusa alapján állapították meg. Amennyiben a RS és a B-CLL neoplasztikus B-sejtjei ugyanazon IgL-t hordozták, akkor a RS feltételezhetően a megelőző B-CLL klónból alakult ki, különböző IgL-k detektálása de novo megjelenő RS-ra utalt A molekuláris biológia fejlődése lehetővé tette a molekuláris technikák alkalmazását a klonális kapcsolat bizonyítására. A korai vizsgálatok során a Southern hibridizáció bizonyult a legalkalmasabbnak ennek eldöntésére. Ehhez az immunglobulin nehézlánchoz (IgH) kötődő próbát alkalmaztak, amivel ki lehetett mutatni, a csíravonal konfigurációt és a génátrendeződést is. Az átrendeződés mérete alapján lehetett

25 a klonális kapcsolatra következtetni Amennyiben a Southern blottal a génátrendeződés azonos méretű sávot eredményez mind a B-CLL, mind a másodlagosan kialakuló DLBCL mintában, akkor ez a tumoros sejtek közös eredetére utalhat. Azonban eltérő migrációs kép esetén egy új malignitás meglétét támasztja alá (11. ábra) G 23 kb 9 kb 6.6 kb 11. ábra Southern-blot analízis klonalitás meghatározása céljából. EcoRI restrikciós endonukleáz emésztést követően a J H régióra tervezett próba segítségével mutatható ki a klonális génátrendeződés. Az 1-2 minták migrációs mintázata azonos, ami közös klonális eredetre utal. A 3-4-es minták esetén a migrációs mintázat különbözik, vagyis a két minta között klonális kapcsolat nem mutatható ki. A lymphomák klonális kapcsolatának modern módszerekkel történő igazolása szükségessé teszi az IgH harmadik komplementaritást determináló régiójának (CDR3) polimeráz láncreakcióval (PCR) történő vizsgálatát. A CDR3 régió tartalmazza a B-sejtekben átrendeződő variábilis (V H ), diverzitás (D) és J H génszakaszok egyedi, adott sejtre specifikus kombinációját, 105, 106 amely végigkíséri az adott B-sejt differenciálódásának folyamatát. 107 A CDR3 régió PCR-amplifikációja a V H és J H -génekre specifikus primerpárokkal érhető el. A megegyező hosszúságú és bázissorrendű amplifikált CDR3 szekvenciák a lymphomák közös klonális eredetére utalnak, a különböző hosszúságú termékek kizárják a klonális kapcsolatot 108 (12.ábra)

26 M 100 bp 12. ábra PCR vizsgálat a minták klonalitásának meghatározására. A CDR3 régióra tervezett specifikus primerekkel végzett PCR reakció segítségével is meghatározható a klonalitás és a két minta közötti kapcsolat is kimutatható. Az 1-es, 2-es minták klonális génátrendeződést mutatnak, azonban az termékek hossza nem azonos, ami arra utal, hogy a két minta klonális eredete különbözik. A 3-as, 4-es mintákban reaktív eredetre utaló futási mintázat látható. Az 5-ös, 6-os mintában is klonális génátrendeződés figyelhető meg, a termékek hossza megegyezik, ami a minták közös klonális eredetére utal. Kevés irodalmi adat áll rendelkezésre arra vonatkozóan, hogy a RS esetek hány százaléka mutat valódi klonális evolúciót a megelőző B-CLL-ből, és hány százalékban alakul ki de novo a B-CLL mellett. Az esetek nagy részében Southern blot 60, 109, 110 hibridizációval következtetnek a klonális eredetre. Eddig kevés olyan tanulmány jelent meg, ahol a klonalitást az IgV H gének szekvencia elemzésével mutatják ki. Ilyen az (a) egyik korábbi tanulmányunk, ahol a CLL és DLBCL minták mindössze négy Richter szindrómás betegben lettek összehasonlítva. Az eredmények azt mutatták, hogy a klonális progresszióban stabil, nem mutált IgV H szekvenciák vettek részt. 85 (b) Cherepakhin és munkatársai azonban ongoing jellegű szomatikus hipermutációt mutattak ki, igaz csak egy esetben
27 (c) Nakamura munkacsoportja két esetet közölt, ahol a DLBCL-es tumorklónok mutált IgV H szekvenciákkal rendelkeztek, de ezek a tumorklónok klonálisan függetlenek voltak az eredeti CLL-től. 63 Sajnos ezekből az adatokból az esetek alacsony száma miatt konklúziót levonni nagyon nehéz
28 IV. CÉLKITŰZÉS Tanulmányunkban a közelmúltban leírt eredményeket figyelembe véve arra kerestük a választ, hogy a Richter szindróma kialakulását az eddigi ismeretek mellett, milyen más genetikai hatások befolyásolják. Főbb kérdéseink a következők voltak: 1. A korábbiaknál nagyobb számú Richter szindrómás eset keresése, amelyhez az adott betegből mind a progressziót megelőző CLL, mind a DLBCL minta rendelkezésre áll. 2. Az ugyanazon betegekből származó CLL-es és DLBCL-es mintákban kimutatható-e klonális kapcsolat? 3. Az IgV H gének szomatikus hipermutációjának elemzése a CLL-es és a DLBCL-es mintákban. 4. Van-e összefüggés a lymphoma transzformáció és a CLL IgV H génjeinek mutációs profilja között?
29 V. ANYAG ÉS MÓDSZER V.1 Szövetminták Tanulmányunkhoz nyolc beteg szövettani mintáit használtuk, akiknél CLL-t, illetve DLBCL-be történő progressziót diagnosztizáltak. A betegek klinikai adatait az 1.táblázat tartalmazza. Vizsgálatainkhoz molekuláris biológiai vizsgálatok végzésére alkalmas fagyasztott biopsziás mintákat, illetve vérmintákat használtunk. A szövettani diagnózis hisztopatológiai, immunfenotípus és immungenotípus vizsgálatokon alapult a WHO lymphoma klasszifikációjának megfelelően. 8 Az összes DLBCL-es mintában a tumorsejtek aránya meghaladta a 90%-ot. Mind a nyolc esetben a CLL és az ennek megfelelő DLBCL azonos fenotípust mutatott (CD5 +, CD19 +, CD20 +, CD23 + ). V.2 DNS izolálás Genomikus DNS-t a szövetmintákból telített NaCl felhasználásával, kisózásos módszerrel nyertük. 112 A fagyasztott szövetmintákat 20 μm vastagságú szeletekre metszettük, majd 3ml maglízis pufferből (10 mm Tris-HCl, 400 mm NaCl, 2 mm EDTA), 20 μl 20%-os Na-dodecilszulfátból (SDS), és 50 μl Proteináz K-ból (1 mg Proteináz K, 2 mm EDTA) álló oldatban homogenizáltuk. A sejteket egy éjszakán át 37 C-on inkubáltuk, majd 1 ml telített NaCl hozzáadását követően az elegyet intenzíven ráztuk egy percig. 20 perces, 2500 fordulat/perc fordulatszámmal történő centrifugálást követően a felülúszóból a DNS-t kétszeres térfogatnyi 100%-os etanol segítségével csaptuk ki. A kivont DNS-t kétszeri 70%-os alkohollal történő mosás után 200 μl TE pufferben (10 mm Tris-HCl, 2 mm EDTA) oldottuk fel, koncentrációját 260 nm-es hullámhosszon fotométer (Gene Quant II, Cambridge, UK) segítségével határoztuk meg. A DNS minták tárolása 4 C-on történt. Vérmintákból a mononuclearis sejteket, Histopaque (Sigma, St. Louis, MO) segítségével, sűrűség-grádiens centrifugálással különítettük el. A szeparálást követően a DNS izolálása a sejtekből az előbb leírtaknak megfelelően történt
30 1. táblázat Nyolc beteg klinikai adatai, valamint a mintavétel helye és ideje. PB: perifériás vér; BM: csontvelő; LN: nyirokcsomó; CLL: chronicus lymphocytás leukaemia; DLBCL: diffúz nagy B-sejtes lymphoma; N: nő; F: férfi Eset Nem Életkor Minta Mintavétel Mintavétel Szövettan (év) dátuma helye 1 N 83 A BM CLL B LN DLBCL 2 N 92 A PB CLL B LN DLBCL 3 N 73 A PB CLL B LN DLBCL 4 F 63 A PB CLL B LN DLBCL 5 F 62 A PB CLL B LN DLBCL 6 F 51 A BM CLL B LN DLBCL 7 N 78 A PB CLL B LN DLBCL 8 N 77 A LN CLL B LN DLBCL
31 V.3 Az IgV H CDR3 régiójának PCR vizsgálata A mintapárok klonalitásának elemzéséhez az IgH-gén CDR3 régiójának szekvenciáját vizsgáltuk polimeráz láncreakcióval (PCR). A vizsgálathoz az IgH-gén 3. framework régiójában (FR3) előforduló veleszületett polimorfizmus kiküszöbölésére az FR3 régióra tervezett degenerált sense primer keveréket és az IgH kapcsolási régióra tervezett (J H ) univerzális antisense primert használtunk fel 85 (13. ábra). Primer Szekvencia FR3 5 - ACA CGG C[C/T][G/C] TGT ATT ACT GT -3 JH 5 - ACC TGA GGA GAC GGT GAC C ábra Az IgH CDR3 PCR reakciókban használt primerek szekvenciája és kötődési helyének sematikus ábrázolása. A PCR reakciók 25 μl végtérfogatban, 100 ng DNS templát, 10 μm sense és antisense primer, 1.5 mm dntp, 50 mm KCl, 10 mm Tris-HCl ph8, 1 mm MgCl 2, 1 unit (U) Taq polimeráz jelenlétében zajlottak. A Perkin-Elmer 2400 GeneAmp PCR készülékben végzett reakciók optimális körülményei a következők voltak: denaturálás 94 C-on 1 percig, annealing 57 C-on 1 percig, extenzió 72 C-on 1 percig, 30 cikluson keresztül. Negatív kontrollként minden reakciósorozathoz reaktív nyirokszövetből izolált DNS-t használtunk. Az amplifikálást követően a polimeráz láncreakció termékeinek detektálása ethidium-bromidot tartalmazó 3%-os agaróz gélelektroforézissel történt. Abban az esetben, ha a vizsgált DNS monoklonális sejtpopulációból származott, diszkrét DNS sávok megjelenése volt várható bázispár magasságában. Ha egy adott beteg mindkét biopsziás mintájából amplifikált PCR termékek azonos hosszúságot
.")
 Reaktív minta esetén a minta migrációs képe egy smear, amit a számos B-sejtből amplifikált különböző méretű termék ad. (5) V.")
32 mutattak (azonos magasságban helyezkedtek el a gélben), az az IgH génátrendeződés egyezésére, az adott mintapár klonális kapcsolatára utalt (14. ábra). 14. ábra Az IgH PCR termékek migrációs mintázatának sematikus ábrája. Abban az esetben, ha a mintapárok klonálisak, a migrációs mintázat mindkét mintánál azonos magasságban egy sávot mutat (1-2 minta). Ha mindkét minta monoklonális, de közöttük nincs klonális kapcsolat, akkor a sávok különböző magasságban helyezkednek el. (3-4) Reaktív minta esetén a minta migrációs képe egy smear, amit a számos B-sejtből amplifikált különböző méretű termék ad. (5) V.4 Az IgV H gének PCR amplifikációja, klónozása és szekvenálása A mintapárok DNS-ét a különböző V H (1-6) géncsaládokra tervezett specifikus sense és a J H régió konszenzus szakaszára tervezett antisense primerekkel amplifikáltuk külön-külön reakcióban (15. ábra). A PCR reakcióelegyek az V.3 pontban leírtakkal azonosak, azonban a ciklusparaméterek a következők voltak: denaturálás 30 másodperc, 95 C; annealing 30 másodperc, 60 C; elongáció 45 másodperc, 72 C, 35 cikluson át. A PCR termékeket pcr-2.1 vektorba klónoztuk a TOPO TA klónozó kit (Invitrogen, San Diego, CA) segítségével. A kompetens sejtek transzformációját követően, legalább 10 olyan kolóniának a plazmid DNS-ét izoláltuk, amelyek a megfelelő méretű inzertet tartalmazták. Az inzert jelenlétének meghatározására a plazmidban az inzerttől 5 és 3 irányban is megtalálható M13 szekvenciára specifikus PCR reakciót végeztünk el
33 Primer Szekvencia VH1-FR1 5 -GGC CTC AGT GAA GGT CTC CTG CAA G-3 VH2-FR1 5 -GTC TGG TCC TAC GCT GGT GAA ACC C-3 VH3-FR1 5 -CTG GGG GGT CCC TGA GAC TCT CCT G-3 VH4-FR1 5 -CTT CGG AGA CCC TGT CCC TCA CCT G-3 VH5-FR1 5 -CGG GGA GTC TCT GAA GAT CTC CTG T-3 VH6-FR1 5 -TCG CAG ACC CTC TCA CTC ACC TGT G-3 JH 5 -CTT ACC TGA GGA GAC GGT GAC C ábra Az IgV H gének PCR reakcióiban használt primerek szekvenciája. Ezt követően az inzertek szekvenálását BigDye 3.1 cycle sequencing kittel ABI 310 automata kapilláris elektroforézis készülékkel (Applied Biosystems, Foster City, CA) végeztük el (16. ábra). A kapott szekvenciákat összehasonlítottuk az Immunogenetics Information System (IMGT) adatbázisban lévő IgV H szekvenciákkal és meghatároztuk a legközelebbi csíravonal (germline) megfelelőjét. Ha a csíravonal szekvenciától való eltérés 2%-nál nagyobbnak mutatkozott, akkor a Casali-Chang formulát 113 alkalmaztuk (2. táblázat). A formula felhasználásával a CDR és az FR régió R mutációt elemezve az IgV H gének antigénszelekció általi érintettségére következtettünk
34 2. táblázat A Casali-Chang formula. (A) Az IgV H gén CDR és FR régióiban várható R mutációk száma. (B) Annak a valószínűsége, hogy az R mutáció a CDR ill. FR régióban van (C) Binomiális modell annak a valószínűségnek a meghatározására, hogy a megfigyelt R mutációk a véletlen folytán alakulnak-e ki. A. Rmut = N RF CDR v. FR L CDRrel v. Frrel Rmut a várt R mutációk száma N a megfigyelt mutációk száma RF a CDR és FR régiók szekvenciából adódó mutációs gyakoriság L a CDR és FR régió mérete B. q = RF CDR v. FR L CDRrel v. Frrel C. n! k p = q (1 q) k! n k! ( ) n k n megfigyelt mutációk száma k megfigyelt R mutációk száma a CDR illetve az FR régióban q annak a valószínűsége, hogy az R mutáció a CDR ill. FR régióban van p = annak a valószínűsége, hogy az R mutációk a véletlen folytán alakulnak ki



35 klónozás vektor PCR termék baktérium transzformációja Inzert ellenőrzése A helyes inzert mérete klónok az agar táptalajon kék nincs inzert fehér van inzert SZEKVENÁLÁS 16. ábra A klónozás és a szekvenálás sematikus folyamatábrája
 génátrendeződést mutatott. Öt esetben (1.-5.")
36 VI. EREDMÉNYEK VI.1 Klonális kapcsolat a CLL és a kialakuló DLBCL minták között A CLL és a DLBCL minták klonális kapcsolatát az IgV H gén CDR3 régiójára specifikus PCR-rel határoztuk meg. Minden, a vizsgálatban szereplő minta monoklonális Ig nehézlánc (IgH) génátrendeződést mutatott. Öt esetben (1.-5. esetek), az adott mintapárok DNS-éből származó PCR termékének hossza és az IgV H -D-J H szekvenciája azonosnak bizonyult, ami a tumorminták közös klonális eredetét támasztja alá. Három esetben (6.-8. esetek) az első és a második biopsziás minta PCR termékének hossza és az IgV H -D-J H szekvenciák sem egyeztek meg, ami az egy betegben kialakult CLL és DLBCL különböző klonális eredetére utal (17. ábra, 3. táblázat). M 1A. 1B. 2A. 2B. 3A. 3B. 6A. 6B. T1 T2 125 bp 100 bp 17. ábra A CDR3 régió PCR amplifikációját követő agaróz gélelektroforézis képe néhány reprezentatív mintával. Az 1.,2., 3. és 6. minta az egy beteghez tartozó CLL-ből (A) illetve az abból kialakult DLBCL-ből (B) származik. A migrációs mintázat alapján az 1.,2.,3. esetek szekvenciális mintái között klonális kapcsolat mutatható ki. A 6A és 6B minta esetén eltérő a migrációs mintázat, ami kizárja a közös klonális eredetet. M - DNS marker; T1, T2 negatív kontroll
.")

.")
37 VI.2 A mintákban előforduló V H géncsaládok A minták IgV H génjeit a 6 V H géncsaládra specifikus sense és a J H -ra specifikus antisense PCR segítségével amplifikáltuk. A minták azzal a V H specifikus primerrel adtak terméket, amely V H család a tumorklón IgH génjét alkotja (18. ábra). A teljes IgV H gén (FR1-CDR1-FR2-CDR2-FR3-CDR3) amplifikációja is alátámasztotta a monoklonális minták közötti klonális kapcsolatot vagy annak hiányát. 300 bp 300 bp 18. ábra 6 V H régió PCR amplifikációját követő agaróz gélelektroforézis két reprezentatív eset mintpárjain. Látható, hogy 2A. és a 2B. minta a V H 3 régióra specifikus primerekkel amplifikált és adott azonos méretű terméket. A 7A. minta a V H 1, a 7B. minta a V H 5-ben adott megfelelő méretű terméket. A klónozást és a kompetens baktérium transzformációját követően a klónozott PCR terméket egy újabb PCR reakció segítségével ellenőriztük (19. ábra). Megfelelő méretű terméket adó reakció esetén a mintákat szekvenáltuk (20A, B, C. ábra). 500 bp * * * * 19. ábra A pcr2.1 plazmid M13 szekvenciájára specifikus PCR amplifikáció egy reprezentatív minta esetén. A *-gal jelölt termékek tartalmazzák a megfelelő inzertet
38 Minden mintában a vizsgált IgV H -D-J H szekvenciák funkcionális génátrendeződést mutatnak, mivel nem tartalmaznak stop kodont vagy crippling mutációt. A minták V H génjeit az adatbázisban levő csíravonal szekvenciákkal hasonlítottuk össze. Négy CLL-es eset (1., 3., 4., 7.) a V H 1, három eset (2., 5., 6.) a V H 3 és egy eset (8. eset) a V H 4 családba tartozó génszegmenst hordozott. Öt esetben (1-5. eset) a DLBCL és a hozzá tartozó CLL-es minta azonos V H génszegmenst tartalmazott. Három esetben (6-8) a DLBCL-es minta és a CLL-es minta V H génszegmense különböző volt. A három DLBCL-es mintából kettőben (6., 8.) a V H 3, egyben (7.) pedig a V H 5 volt megtalálható (3. táblázat). VI.3 Az IgV H gének szomatikus mutációi A vizsgált IgV H gének csíravonal szekvenciától való báziseltéréseinek száma alapján a nyolc CLL-es esetet két csoportra tudtuk osztani. Azokat az eseteket, amelyeknél a báziseltérések száma kevesebb volt 2%-nál, a nem mutált csoportba soroltuk (1.-6. eset). Azokat az eseteket, amelyeknél a báziseltérések száma meghaladta a 2%-ot mutáltnak tekintettük (7.-8.). Abban az öt Richter szindrómás esetben, ahol a CLL és a DLBCL minták IgV H génjei azonosnak bizonyultak (1.-5. eset), az IgV H gének a nem mutált csoportba tartoztak. Három esetben (6.-8. eset), ahol az egy betegből diagnosztizált CLL-es és DLBCL-es minták IgV H génjei különbözőek voltak, a DLBCL-es minták IgV H génjei 91,8-96,9%-os homológiát mutattak a hozzájuk legközelebb eső csíravonal szekvenciákkal. Egy tumormintában sem észleltük az IgV H gének intraklonális diverzitását. VI.4 A szomatikus mutációk elemzése Egy génre gyakorolt negatív vagy pozitív szelekciós hatás hiányában a báziscserék amelyek aminosav cserében nyilvánulnak meg (R, replacement mutációk), illetve amelyek esetében a mutáció nem jár aminosav cserével (S, silent mutációk) az egész kódoló szekvenciában random módon helyezkednek el. Ha egy DNS szakaszon az R mutációk száma nagyobb, mint a véletlenből fakadóan várt
39 mutációk száma, akkor valószínűsíthető, hogy a génen pozitív szelekciós hatás érvényesült. 3. táblázat Nyolc Richter szindrómás betegből vett CLL és DLBCL mintákban előforduló V H géncsaládok és azok mutációinak és klonális kapcsolatának összehasonlítása. *, V H szekvenciák, ahol a CDR régióban több, az FR régióban kevesebb R mutációt találtunk, mint ami a véletlen folytán is létrejött volna (p<0.05); CLL, chronicus lymphocytás leukaemia; DLBCL, diffúz nagy B-sejtes lymphoma Eset Minta V H géncsalád Mutációk Száma Szekvencia homológia (%) Klonális kapcsolat 1 CLL V H Van DLBCL V H CLL V H Van DLBCL V H CLL V H Van DLBCL V H CLL V H Van DLBCL V H CLL V H Van DLBCL V H CLL V H Nincs DLBCL* V H CLL V H Nincs DLBCL V H CLL V H Nincs DLBCL* V H
40 Ezzel ellentétben, ha az észlelt R mutációk száma kevesebb, mint a várt mutációk száma, az a negatív szelekciós nyomással magyarázható, amely így tartósítja azt a struktúrát, amelyet a gén kódol. A mutált DLBCL-es esetek szomatikus mutációinak elemzéséhez a Casali-Chang formulát alkalmaztuk 113. A modell binomiális eloszlást használ, ami alapján kiszámolható, hogy a CDR régiók többlet R mutációi illetőleg az FR régió R mutációinak hiánya a véletlen folytán alakul-e ki. A számolás eredményeit a 4. táblázat tartalmazza, amiből kiderül, hogy a három esetből kettőben a CDR régiók a vártnál több, az FR régiók a vártnál kevesebb R mutációt hordoztak, tehát antigén szelekciós hatásra alakultak ki. 4. táblázat Az IgV H gének analízise azokban a Richter szindrómás betegekben, ahol a DLBCL-es minták mutáltnak bizonyultak. DLBCL, diffúz nagy B-sejtes lymphoma; R, az észlelt és zárójelben feltüntetve a várt R mutációk száma; S, az észlelt S mutációk száma; p, valószínűség; *, szignifikáns különbség (p<0.05) CDR1 és CDR2 FR1, FR2 és FR3 Eset Minta V H géncsalád R S P R S P 6 DLBCL VH (1.63) * 2 (6.90) * 7 DLBCL VH (1.80) (5.10) DLBCL VH (5.00) * 9 (15.9) *
41 IGVH1-8 CAG GTG CAG CTG GTG CAG TCT GGG GCT GAG GTG AAG AAG CCT GGG GCC TCA GTG AAG GTC TCC TGC 1. eset (CLL) eset (Richter) CDR1 IGVH1-8 AAG GCT TCT GGA TAC ACC TTC ACC AGT TAT GAT ATC AAC TGG GTG CGA CAG GCC ACT GGA CAA GGG 1. eset (CLL) C eset (Richter) C CDR2 IGVH1-8 CTT GAG TGG ATG GGA TGG ATG AAC CCT AAC AGT GGT AAC ACA GGC TAT GCA CAG AAG TTC CAG GGC 1. eset (CLL) eset (Richter) IGVH1-8 AGA GTC ACC ATG ACC AGG AAC ACC TCC ATA AGC ACA GCC TAC ATG GAG CTG AGC AGC CTG AGA TCT 1. eset (CLL) T eset (Richter) T CDR3 IGVH1-8 GAG GAC ACG GCC GTG TAT TAC TGT eset (CLL) GCG AGA TCC ATT TAT TAC TAT GAT AGT AGT GGT TAC 1. eset (Richter) A. ábra Az 1. eset IgVH szekvenciája. Ebben az esetben a CLL és a DLBCL szekvenciája (IgVH1-8) megegyezik és mindkettő ugyanazt a két mutációt hordozza. Piros betű: replacement mutáció, Kék betű: silent mutáció
42 IGVH3-11 CAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTC AAG CCT GGA GGG TCC CTG AGA CTC TCC TGT 2. eset (CLL) eset (Richter) CDR1 IGVH3-11 GCA GCC TCT GGA TTC ACC TTC AGT GAC TAC TAC ATG AGC TGG ATC CGC CAG GCT CCA GGG AAG GGG 2. eset (CLL) eset (Richter) CDR2 IGVH3-11 CTG GAG TGG GTT TCA TAC ATT AGT AGT AGT GGT AGT ACC ATA TAC TAC GCA GAC TCT GTG AAG GGC 2. eset (CLL) eset (Richter) IGVH3-11 CGA TTC ACC ATC TCC AGG GAC AAC GCC AAG AAC TCA CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC 2. eset (CLL) eset (Richter) CDR3 IGVH3-11 GAG GAC ACG GCC GTG TAT TAC TGT eset (CLL) GCG AGA GAT TGG GCC GCC TGG TGG TTC GGG GAG TCC CAC TTT 2. eset (Richter) GCG AGA GAT TGG GCC GCC TGG TGG TTC GGG GAG TCC CAC TTT IGVH eset (CLL) GAC TAC 2. eset (Richter) GAC TAC 20B. ábra Az 2. eset IgVH szekvenciája. Ebben az esetben a CLL és a DLBCL szekvenciája (IgVH3-11) megegyezik és egyik eset sem hordoz mutációt
43 IGVH1-18 CAG GTT CAG CTG GTG CAG TCT GGA GCT GAG GTG AAG AAG CCT GGG GCC TCA GTG AAG GTC TCC TGC 7. eset (CLL) CDR1 IGVH1-18 AAG GCT TCT GGT TAC ACC TTT ACC AGC TAT GGT ATC AGC TGG GTG CGA CAG GCC CCT GGA CAA GGG 7. eset (CLL) GT -A T C CDR2 IGVH1-18 CTT GAG TGG ATG GGA TGG ATC AGC GCT TAC AAT GGT AAC ACA AAC TAT GCA CAG AAG CTC CAG GGC 7. eset (CLL) G C T- --G IGVH1-18 AGA GTC ACC ATG ACC ACA GAC ACA TCC ACG AGC ACA GCC TAC ATG GAG CTG AGG AGC CTG AGA TCT 7. eset (CLL) A CDR3 IGVH1-18 GAC GAC ACG GCC GTG TAT TAC TGT eset (CLL) GCG AGA GAT CAT CTT CCC ACT ACG ATT TTT IGVH5-51 GAG GTG CAG CTG GTG CAG TCT GGA GCA GAG GTG AAA AAG CCC GGG GAG TCT CTG AAG ATC TCC TGT 7. eset (Richter) CDR1 IGVH5-51 AAG GGT TCT GGA TAC AGC TTT ACC AGC TAC TGG ACC GGC TGG GTG CGC CAG ATG CCC GGG AAA GGC 7. eset (Richter) --- -A C A- -A CDR2 IGVH5-51 TTG GAG TGG ATG GGG ATC ATC TAT CCT GGT GAC TCT GAT ACC AGA TAC AGC CCG TCC TTC CAA GGC 7. eset (Richter) A IGVH5-51 CAG GTC ACC ATC TCA GCC GAC AAG TCC ATC AGC ACC GCC TAC CTG CAG TGG AGC AGC CTG AAG GCC 7. eset (Richter) C T T --A CDR3 IGVH5-51 TCG GAC ACC GCC ATG TAT TAC TGT eset (Richter) GCG AGA CAG ACT ACA CTG ACT ATG TCC ATT GAC GAC 20C. ábra A 7. eset IgVH szekvenciája. Ebben az esetben a CLL (IgVH1-18) és a DLBCL (IgVH5-51) szekvenciája nem egyezett meg és mindkettő tartalmaz mutációkat. Piros betű: replacement mutáció, Kék betű: silent mutáció
44 VII. MEGBESZÉLÉS A hisztológiai transzformáció jelensége follicularis lymphoma, köpenysejtes lymphoma és CLL esetén jöhet létre, amikoris a progressziót követően sokkal agresszívebb lefolyású betegség alakul ki. Richter szindrómának nevezzük az alacsony malignitású CLL talaján másodlagosan kialakuló magas malignitású DLBCL-t. A folyamat során a betegség terápia rezisztenssé és klinikailag rossz prognózisúvá válik. Munkánk során arra kerestük a választ, hogy a Richter szindróma mutált vagy nem mutált IgV H génekkel rendelkező CLL-ből alakul-e ki. Ehhez az IgV H gének mutációs státuszát elemeztük olyan betegek mintapárjaiban, ahol az egyik minta a betegség korai CLL-es stádiumából származik, a másik pedig a Richter szindróma fennállását megállapító DLBCL-t tartalmazó nyirokcsomó volt. A vizsgálat időpontjában nyolc beteg mintapárja állt rendelkezésünkre, ami kevésnek tűnhet, azonban a RS előfordulását és a szekvenciális minták ritkaságát figyelembe véve mindenképpen jelentős. Az 1-3. esetben a CLL és a DLBCL diagnózisa gyakorlatilag egyidőben került felállításra. Ezekben az esetekben a DLBCL okozta nyirokcsomómegnagyobbodás miatt végzett teljes kivizsgálás során derült fény a feltehetően korábban meglévő, panaszt nem okozó CLL-re. Eredményeink arra utalnak, hogy a DLBCL mind mutált, mind nem mutált IgV H génnel rendelkező CLL-es betegekben is kialakulhat, de csak a nem mutált IgV H gént hordozó DLBCL fejlődik ki a CLL-es tumorklónból, amely szintén nem mutált. A mutált IgV H génnel rendelkező DLBCL-ek klonálisan független, másodlagos tumorként jönnek létre. Munkacsoportunknak e dolgozat alapját képző cikk megjelenése óta 5 újabb Richter szindrómás esetet sikerült vizsgálni. Az újabb esetek IgV H génjeinek szekvenálása és elemzése megerősíti a fent leírtakat. 114 A RS során a DLBCL kialakulása biológiai szempontból két különböző utat foglal magába. A DLBCL kifejlődhet a már meglévő CLL-es tumorklónból, vagy kialakulhat mint egy másodlagos, független tumor 47, 85. Ezt figyelembe véve javasolták a következő terminológiák használatát. A lymphoma transzformáció olyan eseteket jelöl, ahol a DLBCL a CLL-es klónból alakul ki. Ezzel ellentétben a kompozit lymphoma fogalmát azokra az esetekre tartják fenn, ahol a DLBCL és a CLL klonálisan függetlenek egymástól 115 (21. ábra)






45 21. ábra A V H gének mutációs státusza és a DLBCL kialakulásának útvonalai közötti kapcsolat Richter szindrómában. (A, lymphoma transzformáció) A CLL klonális transzformációja DLBCL-be a nem mutált V H génnel rendelkező CLL-es esetekben történik. (B, kompozit lymphoma) A CLL-től klonálisan független DLBCL, amely mutált V H gént hordoz, kialakulhat mutált illetve nem mutált V H géneket hordozó CLL mellett is. A munkánk során végzett IgV H gén mutációs státuszának analízise arra enged következtetni, hogy a lymphoma transzformáció -val kialakult DLBCL csak nem mutált IgV H génnel rendelkező CLL-es betegekben fordul elő. A nem mutált CLL gyakran hordoz egyéb genetikai eltéréseket is, amelyek között meg kell említeni a 12-es triszómiát, a 17p és a 11q deléciót. 18, 116, 117 Ezeket az eltéréseket leírták a transzformált DLBCL klónokban is, de leírtak más a Richter transzformációval társuló molekuláris és kromoszómális szintű anomáliát is 92, 118 (5. táblázat). Ezen tanulmányok alapján arra
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása. Uher Ferenc, PhD, DSc
 A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Az immunglobulinok szerkezete Fab V L V H C L C H 1 C H 1 Az egér immunglobulin géncsaládok szerveződése Hlánc
Immunológia alapjai előadás. A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és
 Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia alapjai 15-16. előadás A humorális immunválasz formái és lefolyása: extrafollikuláris reakció és csíracentrum reakció, affinitás-érés és izotípusváltás. A B-sejt fejlődés szakaszai HSC Primer
Immunológia 4. A BCR diverzitás kialakulása
 Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Immunológia 4. A BCR diverzitás kialakulása 2017. október 4. Bajtay Zsuzsa A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Immunológiai módszerek a klinikai kutatásban 3. előadás Az immunrendszer molekuláris elemei: antigén, ellenanyag, Ig osztályok Az antigén meghatározása Detre László: antitest generátor - Régi meghatározás:
Klonális szelekció a follikuláris lymphomák csontvelői terjedése során
 Klonális szelekció a follikuláris lymphomák csontvelői terjedése során Doktori értekezés Dr. Bognár Ágnes Semmelweis Egyetem Patológiai Doktori Iskola Témavezető: Dr. Szepesi Ágota egyetemi adjunktus,
Klonális szelekció a follikuláris lymphomák csontvelői terjedése során Doktori értekezés Dr. Bognár Ágnes Semmelweis Egyetem Patológiai Doktori Iskola Témavezető: Dr. Szepesi Ágota egyetemi adjunktus,
4. A humorális immunválasz október 12.
 4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
4. A humorális immunválasz 2016. október 12. A klónszelekciós elmélet sarokpontjai: Monospecifictás: 1 sejt 1-féle specificitású receptor Az antigén receptorhoz kötődése aktiválja a limfocitát A keletkező
GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN
 GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN PhD értekezés tézisei Dr. Fülöp Zsolt Témavezeto : Dr. Matolcsy András Programvezeto : Dr. Jeney
GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN PhD értekezés tézisei Dr. Fülöp Zsolt Témavezeto : Dr. Matolcsy András Programvezeto : Dr. Jeney
Doktori tézisek. Dr. Reiniger Lilla. Semmelweis Egyetem Patológia Tudományok Doktori Iskolája. Témavezető: Dr. Szepesi Ágota egyetemi adjunktus, PhD
 Az aktiváció-indukált citidin deamináz és az aberráns szomatikus hipermutáció szerepe a krónikus lymphocytás leukaemia Richter és prolymphocytás transzformációjában Doktori tézisek Dr. Reiniger Lilla Semmelweis
Az aktiváció-indukált citidin deamináz és az aberráns szomatikus hipermutáció szerepe a krónikus lymphocytás leukaemia Richter és prolymphocytás transzformációjában Doktori tézisek Dr. Reiniger Lilla Semmelweis
Lehetőségek és korlátok a core-biopszián alapuló limfóma diagnosztikában
 Lehetőségek és korlátok a core-biopszián alapuló limfóma diagnosztikában Strausz Tamás Tóth Erika Országos Onkológiai Intézet Sebészeti és Molekuláris Patológiai Osztály FiPaT 2012 Elvi alapok WHO klasszifikáció
Lehetőségek és korlátok a core-biopszián alapuló limfóma diagnosztikában Strausz Tamás Tóth Erika Országos Onkológiai Intézet Sebészeti és Molekuláris Patológiai Osztály FiPaT 2012 Elvi alapok WHO klasszifikáció
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata
 sejtek mikro RNS (mir) profiljának vizsgálata") Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Lymphoma sejtvonalak és gyerekkori leukémia (ALL) sejtek mikro RNS (mir) profiljának vizsgálata Dr. Nemes Karolina, Márk Ágnes, Dr. Hajdu Melinda, Csorba Gézáné, Dr. Kopper László, Dr. Csóka Monika, Dr.
Az ellenanyagok szerkezete és funkciója. Erdei Anna Immunológiai Tanszék ELTE
 Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Az ellenanyagok szerkezete és funkciója Erdei Anna Immunológiai Tanszék ELTE Bev. 1. ábra Immunhomeosztázis A veleszületett és az adaptív immunrendszer szorosan együttműködik az immunhomeosztázis fenntartásáért
Kappelmayer János. Malignus hematológiai megbetegedések molekuláris háttere. MOLSZE IX. Nagygyűlése. Bük, 2005 szeptember
 Kappelmayer János Malignus hematológiai megbetegedések molekuláris háttere MOLSZE IX. Nagygyűlése Bük, 2005 szeptember 29-30. Laboratóriumi vizsgálatok hematológiai malignómákban Általános laboratóriumi
Kappelmayer János Malignus hematológiai megbetegedések molekuláris háttere MOLSZE IX. Nagygyűlése Bük, 2005 szeptember 29-30. Laboratóriumi vizsgálatok hematológiai malignómákban Általános laboratóriumi
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Lymphomák molekuláris patológiai diagnosztikája
 OTKA nyilvántartási szám:t/f 043323 Lymphomák molekuláris patológiai diagnosztikája ZÁRÓJELENTÉS A lymphomák diagnosztikája és klasszifikációja az elmúlt 10 évben jelentősen átalakult. A jelenlegi osztályozás
OTKA nyilvántartási szám:t/f 043323 Lymphomák molekuláris patológiai diagnosztikája ZÁRÓJELENTÉS A lymphomák diagnosztikája és klasszifikációja az elmúlt 10 évben jelentősen átalakult. A jelenlegi osztályozás
Immunológia I. 4. előadás. Kacskovics Imre
 Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Immunológia I. 4. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán 3.2. ábra A hemopoetikus őssejt aszimmetrikus osztódása 3.3. ábra
Az Ig génátrendeződés
 Az Ig génátrendeződés Háromféle változás játszódik le a molekula szerkezetét tekintve: B sejtek fejlődése alatt: VDJ átrendeződés (rekombináció) IgH izotípusváltás rekombináció (CSR) Szomatikus hipermutáció
Az Ig génátrendeződés Háromféle változás játszódik le a molekula szerkezetét tekintve: B sejtek fejlődése alatt: VDJ átrendeződés (rekombináció) IgH izotípusváltás rekombináció (CSR) Szomatikus hipermutáció
Molekuláris vizsgálatok B- és T-sejtes extranodalis Non-Hodgkin. limfómákban. Dr. Gurbity Pálfi Tímea. Összefoglalás
 1 W L 1 Molekuláris vizsgálatok B- és T-sejtes extranodalis Non-Hodgkin limfómákban Dr. Gurbity Pálfi Tímea Összefoglalás A malignus limfómák korai felismerése, a pontos patológiai diagnózis, valamint
1 W L 1 Molekuláris vizsgálatok B- és T-sejtes extranodalis Non-Hodgkin limfómákban Dr. Gurbity Pálfi Tímea Összefoglalás A malignus limfómák korai felismerése, a pontos patológiai diagnózis, valamint
Immunológia I. 2. előadás. Kacskovics Imre (imre.kacskovics@ttk.elte.hu)
") Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia I. 2. előadás Kacskovics Imre (imre.kacskovics@ttk.elte.hu) Az immunválasz kialakulása A veleszületett és az adaptív immunválasz összefonódása A veleszületett immunválasz mechanizmusai A veleszületett
Immunológia Alapjai. 13. előadás. Elsődleges T sejt érés és differenciálódás
 Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
Immunológia Alapjai 13. előadás Elsődleges T sejt érés és differenciálódás A T és B sejt receptor eltérő szerkezetű A T sejt receptor komplex felépítése + DOMÉNES SZERKEZET αβ ΤcR SP(CD4+ vagy CD8+) γδ
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
DNS-szekvencia meghatározás
 DNS-szekvencia meghatározás Gilbert 1980 (1958) Sanger 3-1 A DNS-polimerázok jellemzői 5'-3' polimeráz aktivitás 5'-3' exonukleáz 3'-5' exonukleáz aktivitás Az új szál szintéziséhez kell: templát DNS primer
DNS-szekvencia meghatározás Gilbert 1980 (1958) Sanger 3-1 A DNS-polimerázok jellemzői 5'-3' polimeráz aktivitás 5'-3' exonukleáz 3'-5' exonukleáz aktivitás Az új szál szintéziséhez kell: templát DNS primer
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van.
 In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van. Kneif Józsefné PTE KK Pathologiai Intézet Budapest 2017. 05. 26 Kromoszóma rendellenesség kimutatás PCR technika: izolált nukleinsavak
In Situ Hibridizáció a pathologiai diagnosztikában és ami mögötte van. Kneif Józsefné PTE KK Pathologiai Intézet Budapest 2017. 05. 26 Kromoszóma rendellenesség kimutatás PCR technika: izolált nukleinsavak
A genetikai lelet értelmezése monogénes betegségekben
 A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
A genetikai lelet értelmezése monogénes betegségekben Tory Kálmán Semmelweis Egyetem, I. sz. Gyermekklinika A ~20 ezer fehérje-kódoló gén a 23 pár kromoszómán A kromoszómán található bázisok száma: 250M
AZ ABERRÁNS SZOMATIKUS HIPERMUTÁCIÓ ÉS AZ AKTIVÁCIÓ-INDUKÁLT CITIDIN DEAMINÁZ SZEREPE A MEDIASTINALIS NAGY B-SEJTES LYMPHOMA PATOGENEZISÉBEN
 AZ ABERRÁNS SZOMATIKUS HIPERMUTÁCIÓ ÉS AZ AKTIVÁCIÓ-INDUKÁLT CITIDIN DEAMINÁZ SZEREPE A MEDIASTINALIS NAGY B-SEJTES LYMPHOMA PATOGENEZISÉBEN Doktori értekezés Semmelweis Egyetem Patológiai Tudományok Doktori
AZ ABERRÁNS SZOMATIKUS HIPERMUTÁCIÓ ÉS AZ AKTIVÁCIÓ-INDUKÁLT CITIDIN DEAMINÁZ SZEREPE A MEDIASTINALIS NAGY B-SEJTES LYMPHOMA PATOGENEZISÉBEN Doktori értekezés Semmelweis Egyetem Patológiai Tudományok Doktori
Tóth Erika Sebészeti és Molekuláris Patológia Osztály Országos Onkológiai Intézet. Frank Diagnosztika Szimpózium, DAKO workshop 2012.
 Tóth Erika Sebészeti és Molekuláris Patológia Osztály Országos Onkológiai Intézet Frank Diagnosztika Szimpózium, DAKO workshop 2012.december 7 Follicularis hám eredetű daganatok Jól differenciált tumorok
Tóth Erika Sebészeti és Molekuláris Patológia Osztály Országos Onkológiai Intézet Frank Diagnosztika Szimpózium, DAKO workshop 2012.december 7 Follicularis hám eredetű daganatok Jól differenciált tumorok
AZ ABERRÁNS HIPERMUTÁCIÓ OKOZTA GENETIKAI INSTABILITÁS SZEREPE A B-SEJTES NON-HODGKIN LYMPHOMÁK KIALAKULÁSÁBAN ÉS TRANSZFORMÁCIÓJÁBAN
 (OTKA T049306 ) AZ ABERRÁNS HIPERMUTÁCIÓ OKOZTA GENETIKAI INSTABILITÁS SZEREPE A B-SEJTES NON-HODGKIN LYMPHOMÁK KIALAKULÁSÁBAN ÉS TRANSZFORMÁCIÓJÁBAN TEMATIKUS OTKA PÁLYÁZAT ÉLETTUDOMÁNYOK Matolcsy András
(OTKA T049306 ) AZ ABERRÁNS HIPERMUTÁCIÓ OKOZTA GENETIKAI INSTABILITÁS SZEREPE A B-SEJTES NON-HODGKIN LYMPHOMÁK KIALAKULÁSÁBAN ÉS TRANSZFORMÁCIÓJÁBAN TEMATIKUS OTKA PÁLYÁZAT ÉLETTUDOMÁNYOK Matolcsy András
GENETIKAI INSTABILITÁS FOLLICULARIS LYMPHOMA HISZTOLÓGIAI TRANSZFORMÁCIÓJA SORÁN. PhD értekezés tézisei. Dr. Nagy Mónika
 GENETIKAI INSTABILITÁS FOLLICULARIS LYMPHOMA HISZTOLÓGIAI TRANSZFORMÁCIÓJA SORÁN PhD értekezés tézisei Dr. Nagy Mónika Témavezető: Programvezetők: Dr. Matolcsy András Dr. Kelényi Gábor Dr.Kellermayer Miklós
GENETIKAI INSTABILITÁS FOLLICULARIS LYMPHOMA HISZTOLÓGIAI TRANSZFORMÁCIÓJA SORÁN PhD értekezés tézisei Dr. Nagy Mónika Témavezető: Programvezetők: Dr. Matolcsy András Dr. Kelényi Gábor Dr.Kellermayer Miklós
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői.
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
CD8 pozitív primér bőr T-sejtes limfómák 14 eset kapcsán
 CD8 pozitív primér bőr T-sejtes limfómák 14 eset kapcsán Szepesi Ágota 1, Csomor Judit 1, Marschalkó Márta 2 Erős Nóra 2, Kárpáti Sarolta 2, Matolcsy András 1 Semmelweis Egyetem I. Patológiai és Kísérlet
CD8 pozitív primér bőr T-sejtes limfómák 14 eset kapcsán Szepesi Ágota 1, Csomor Judit 1, Marschalkó Márta 2 Erős Nóra 2, Kárpáti Sarolta 2, Matolcsy András 1 Semmelweis Egyetem I. Patológiai és Kísérlet
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Immunológiai módszerek a klinikai kutatásban 6. előadás Humorális és celluláris immunválasz A humorális (B sejtes) immunválasz lépései Antigén felismerés B sejt aktiváció: proliferáció, differenciálódás
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a PD-L1 és PD-1 fehérjék túlélésre gyakorolt hatása
 Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Tüdő adenocarcinomásbetegek agyi áttéteiben jelenlévő immunsejtek, valamint a és PD-1 fehérjék túlélésre gyakorolt hatása Téglási Vanda, MoldvayJudit, Fábián Katalin, Csala Irén, PipekOrsolya, Bagó Attila,
Immunológia alapjai előadás. Az immunológiai felismerés molekuláris összetevői
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői Az antigén fogalma. Antitestek, T- és B- sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Engedélyszám: 18211-2/2011-EAHUF Verziószám: 1. 2460-06 Humángenetikai vizsgálatok követelménymodul szóbeli vizsgafeladatai
 1. feladat Ismertesse a gyakorlaton lévő szakasszisztens hallgatóknak a PCR termékek elválasztása céljából végzett analitikai agaróz gélelektroforézis során használt puffert! Az ismertetés során az alábbi
1. feladat Ismertesse a gyakorlaton lévő szakasszisztens hallgatóknak a PCR termékek elválasztása céljából végzett analitikai agaróz gélelektroforézis során használt puffert! Az ismertetés során az alábbi
MIKROSZATELIT DNS- VIZSGÁLATOK A MOCSÁRI TEKNŐS NÉGY DUNÁNTÚLI ÁLLOMÁNYÁN
 MIKROSZATELIT DNS- VIZSGÁLATOK A MOCSÁRI TEKNŐS NÉGY DUNÁNTÚLI ÁLLOMÁNYÁN Molnár Tamás 1, Lanszki József 1, Magyary István 1, Jeney Zsigmond 2, Lehoczky István 2 1 Kaposvári Egyetem Állattudományi Kar,
MIKROSZATELIT DNS- VIZSGÁLATOK A MOCSÁRI TEKNŐS NÉGY DUNÁNTÚLI ÁLLOMÁNYÁN Molnár Tamás 1, Lanszki József 1, Magyary István 1, Jeney Zsigmond 2, Lehoczky István 2 1 Kaposvári Egyetem Állattudományi Kar,
A Hardy-Weinberg egyensúly. 2. gyakorlat
 A Hardy-Weinberg egyensúly 2. gyakorlat A Hardy-Weinberg egyensúly feltételei: nincs szelekció nincs migráció nagy populációméret (nincs sodródás) nincs mutáció pánmixis van allélgyakoriság azonos hímekben
A Hardy-Weinberg egyensúly 2. gyakorlat A Hardy-Weinberg egyensúly feltételei: nincs szelekció nincs migráció nagy populációméret (nincs sodródás) nincs mutáció pánmixis van allélgyakoriság azonos hímekben
A termesztett búza diploid őseinek molekuláris citogenetikai elemzése: pachytén- és fiber-fish.
 OTKA K67808 zárójelentés 2012. A termesztett búza diploid őseinek molekuláris citogenetikai elemzése: pachytén- és fiber-fish. A fluoreszcens in situ hibridizáció (FISH) olyan technikai fejlettséget ért
OTKA K67808 zárójelentés 2012. A termesztett búza diploid őseinek molekuláris citogenetikai elemzése: pachytén- és fiber-fish. A fluoreszcens in situ hibridizáció (FISH) olyan technikai fejlettséget ért
A krónikus myeloid leukémia kezelésének finanszírozási protokollja (eljárásrend)
") A krónikus myeloid leukémia kezelésének finanszírozási protokollja (eljárásrend) Országos Egészségbiztosítási Pénztár Elemzési, Orvosszakértői és Szakmai Ellenőrzési Főosztály Budapest, 2013. június 26.
A krónikus myeloid leukémia kezelésének finanszírozási protokollja (eljárásrend) Országos Egészségbiztosítási Pénztár Elemzési, Orvosszakértői és Szakmai Ellenőrzési Főosztály Budapest, 2013. június 26.
Immunológia alapjai előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok:
 Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Immunológia alapjai 3 4. előadás Az immunológiai felismerés molekuláris összetevői. Az antigén fogalma. Antitestek, T- és B-sejt receptorok: molekuláris szerkezet, funkciók, alcsoportok Az antigén meghatározása
Molekuláris genetikai vizsgáló. módszerek az immundefektusok. diagnosztikájában
 Molekuláris genetikai vizsgáló módszerek az immundefektusok diagnosztikájában Primer immundefektusok A primer immundeficiencia ritka, veleszületett, monogénes öröklődésű immunhiányos állapot. Családi halmozódást
Molekuláris genetikai vizsgáló módszerek az immundefektusok diagnosztikájában Primer immundefektusok A primer immundeficiencia ritka, veleszületett, monogénes öröklődésű immunhiányos állapot. Családi halmozódást
Kórélettan hematológiai esetek
 8. eset Középkorú nőbeteg. Két hete gyengébb, fáradékonyabb. Háziorvosa laborvizsgálatot kért, a lelet ismeretében kórházba utalta. Fehérvérsejt 62,7 G/l H 4 10 G/l Neutrophil % 40 70 % Neutrophil (abs)
8. eset Középkorú nőbeteg. Két hete gyengébb, fáradékonyabb. Háziorvosa laborvizsgálatot kért, a lelet ismeretében kórházba utalta. Fehérvérsejt 62,7 G/l H 4 10 G/l Neutrophil % 40 70 % Neutrophil (abs)
Antigén-felismerő receptorok (BCR, TCR) GYTK Immunológia
 GYTK Immunológia") Antigén-felismerő receptorok (BCR, TCR) GYTK Immunológia Dr Pállinger Éva Genetikai Sejt- és Immunbiológiai Intézet Nem antigén- specifikus antigénreceptorok Opszonizáló receptorok Mintázat felismerő receptorok
Antigén-felismerő receptorok (BCR, TCR) GYTK Immunológia Dr Pállinger Éva Genetikai Sejt- és Immunbiológiai Intézet Nem antigén- specifikus antigénreceptorok Opszonizáló receptorok Mintázat felismerő receptorok
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Prenatalis diagnosztika lehetőségei mikor, hogyan, miért? Dr. Almássy Zsuzsanna Heim Pál Kórház, Budapest Toxikológia és Anyagcsere Osztály
 Prenatalis diagnosztika lehetőségei mikor, hogyan, miért? Dr. Almássy Zsuzsanna Heim Pál Kórház, Budapest Toxikológia és Anyagcsere Osztály Definíció A prenatális diagnosztika a klinikai genetika azon
Prenatalis diagnosztika lehetőségei mikor, hogyan, miért? Dr. Almássy Zsuzsanna Heim Pál Kórház, Budapest Toxikológia és Anyagcsere Osztály Definíció A prenatális diagnosztika a klinikai genetika azon
Új könnyűlánc diagnosztika. Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006
 Új könnyűlánc diagnosztika Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006 1845 Bence Jones Protein vizelet fehérje 1922 BJP I-II típus 1956 BJP
Új könnyűlánc diagnosztika Dr. Németh Julianna Országos Gyógyintézeti Központ Immundiagnosztikai Osztály MLDT-MIT Továbbképzés 2006 1845 Bence Jones Protein vizelet fehérje 1922 BJP I-II típus 1956 BJP
PLAZMASEJT OKOZTA BETEGSÉGEK, MYELOMA MULTIPLEX, LYMPHOMÁK
 PLAZMASEJT OKOZTA BETEGSÉGEK, MYELOMA MULTIPLEX, LYMPHOMÁK Belgyógyászat alapjai fogorvosoknak Reismann Péter SE ÁOK II.sz. Belgyógyászati Klinika 2015.09.15 Fehérvérsejtképzés, myelopoesis Haemopoetikus
PLAZMASEJT OKOZTA BETEGSÉGEK, MYELOMA MULTIPLEX, LYMPHOMÁK Belgyógyászat alapjai fogorvosoknak Reismann Péter SE ÁOK II.sz. Belgyógyászati Klinika 2015.09.15 Fehérvérsejtképzés, myelopoesis Haemopoetikus
MAGYOT évi Tudományos Szimpóziuma Május 5-6, Budapest
 MAGYOT 2017. évi Tudományos Szimpóziuma Május 5-6, Budapest A petefészekrákok kezelésében nem régen került bevezetésre egy újabb fenntartó kezelés BRCA mutációt hordozó (szomatikus vagy germinális) magas
MAGYOT 2017. évi Tudományos Szimpóziuma Május 5-6, Budapest A petefészekrákok kezelésében nem régen került bevezetésre egy újabb fenntartó kezelés BRCA mutációt hordozó (szomatikus vagy germinális) magas
Immunológia alapjai. Az immunválasz szupressziója Előadás. A szupresszióban részt vevő sejtes és molekuláris elemek
 Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Immunológia alapjai 19 20. Előadás Az immunválasz szupressziója A szupresszióban részt vevő sejtes és molekuláris elemek Mi a szupresszió? Általános biológiai szabályzó funkció. Az immunszupresszió az
Többgénes jellegek. 1. Klasszikus (poligénes) mennyiségi jellegek. 2.Szinte minden jelleg több gén irányítása alatt áll
 mennyiségi jellegek. 2.Szinte minden jelleg több gén irányítása alatt áll") Többgénes jellegek Többgénes jellegek 1. 1. Klasszikus (poligénes) mennyiségi jellegek Multifaktoriális jellegek: több gén és a környezet által meghatározott jellegek 2.Szinte minden jelleg több gén irányítása
Többgénes jellegek Többgénes jellegek 1. 1. Klasszikus (poligénes) mennyiségi jellegek Multifaktoriális jellegek: több gén és a környezet által meghatározott jellegek 2.Szinte minden jelleg több gén irányítása
Immunológiai módszerek a klinikai kutatásban
 Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Immunológiai módszerek a klinikai kutatásban 2. előadás A veleszületett és specifikus immunrendszer sejtjei Vérképzés = Haematopeiesis, differenciálódás Kék: ősssejt Sötétkék: éretlen sejtek Barna: érett
Az X kromoszóma inaktívációja. A kromatin szerkezet befolyásolja a génexpressziót
 Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Az X kromoszóma inaktívációja A kromatin szerkezet befolyásolja a génexpressziót Férfiak: XY Nők: XX X kromoszóma: nagy méretű több mint 1000 gén Y kromoszóma: kis méretű, kevesebb, mint 100 gén Kompenzációs
Immunológia alapjai előadás. Az antigén-receptor gének szerveződése és átrendeződése. Primer B-sejt fejlődés
 Immunológia alapjai 11-12. előadás Az antigén-receptor gének szerveződése és átrendeződése. Primer B-sejt fejlődés Az antigén-receptor gének kifejeződésének főbb kérdései Minden testi sejt tartalmaz TcR/BcR
Immunológia alapjai 11-12. előadás Az antigén-receptor gének szerveződése és átrendeződése. Primer B-sejt fejlődés Az antigén-receptor gének kifejeződésének főbb kérdései Minden testi sejt tartalmaz TcR/BcR
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása
 A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Délpesti Centrumkórház Országos Hematológiai és Infektológiai Intézet, Budapest Az immunglobulinok szerkezete
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Délpesti Centrumkórház Országos Hematológiai és Infektológiai Intézet, Budapest Az immunglobulinok szerkezete
Algaközösségek ökológiai, morfológiai és genetikai diverzitásának összehasonlítása szentély jellegű és emberi használatnak kitett élőhelykomplexekben
 Algaközösségek ökológiai, morfológiai és genetikai diverzitásának összehasonlítása szentély jellegű és emberi használatnak kitett élőhelykomplexekben Duleba Mónika Környezettudományi Doktori Iskola I.
Algaközösségek ökológiai, morfológiai és genetikai diverzitásának összehasonlítása szentély jellegű és emberi használatnak kitett élőhelykomplexekben Duleba Mónika Környezettudományi Doktori Iskola I.
Antigén, Antigén prezentáció
 Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Antigén, Antigén prezentáció Biológiai Intézet Immunológiai Tanszék Bajtay Zsuzsa ELTE, TTK Biológiai Intézet Immunológiai Tanszék ORFI Klinikai immunológia tanfolyam, 2019. február. 26 Bev. 2. ábra Az
Immunológia Világnapja
 a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
a Magyar Tudományos Akadémia Biológiai Osztály, Immunológiai Bizottsága és a Magyar Immunológiai Társaság Immunológia Világnapja - 2016 Tumorbiológia Dr. Tóvári József, Országos Onkológiai Intézet Mágikus
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Az adaptív immunválasz kialakulása. Erdei Anna Immunológiai Tanszék ELTE
 Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Az adaptív immunválasz kialakulása Erdei Anna Immunológiai Tanszék ELTE NK sejt T Bev. 1. ábra Immunhomeosztázis A veleszületett immunrendszer elemei nélkül nem alakulhat ki az adaptív immunválasz A veleszületett
Doktori értekezés. Dr. Reiniger Lilla. I.sz. Patológiai és Kísérleti Rákkutató Intézet
 Az aktiváció-indukált citidin deamináz és az aberráns szomatikus hipermutáció szerepe a krónikus lymphocytás leukaemia Richter és prolymphocytás transzformációjában Doktori értekezés Dr. Reiniger Lilla
Az aktiváció-indukált citidin deamináz és az aberráns szomatikus hipermutáció szerepe a krónikus lymphocytás leukaemia Richter és prolymphocytás transzformációjában Doktori értekezés Dr. Reiniger Lilla
Immunitás és evolúció
 Immunitás és evolúció (r)evolúció az immunrendszerben Az immunrendszer evolúciója Müller Viktor ELTE Növényrendszertani, Ökológiai és Elméleti Biológiai Tanszék http://ramet.elte.hu/~viktor Az immunitás
Immunitás és evolúció (r)evolúció az immunrendszerben Az immunrendszer evolúciója Müller Viktor ELTE Növényrendszertani, Ökológiai és Elméleti Biológiai Tanszék http://ramet.elte.hu/~viktor Az immunitás
67. Pathologus Kongresszus
 A kemoirradiáció okozta oncocytás átalakulás szövettani, immunhisztokémiai, ultrastruktúrális jellemzői és lehetséges prognosztikus jelentősége rectum adenocarcinomákban 67. Pathologus Kongresszus Bogner
A kemoirradiáció okozta oncocytás átalakulás szövettani, immunhisztokémiai, ultrastruktúrális jellemzői és lehetséges prognosztikus jelentősége rectum adenocarcinomákban 67. Pathologus Kongresszus Bogner
Esetbemutatás. Dr. Iván Mária Uzsoki Kórház 2013.11.07.
 Esetbemutatás Dr. Iván Mária Uzsoki Kórház 2013.11.07. Esetbemutatás I. 26 éves férfi 6 héttel korábban bal oldali herében elváltozást észlelt,majd 3 héttel később haemoptoe miatt kereste fel orvosát antibiotikumos
Esetbemutatás Dr. Iván Mária Uzsoki Kórház 2013.11.07. Esetbemutatás I. 26 éves férfi 6 héttel korábban bal oldali herében elváltozást észlelt,majd 3 héttel később haemoptoe miatt kereste fel orvosát antibiotikumos
Dr. Máthéné Dr. Szigeti Zsuzsanna és munkatársai
 Kar: TTK Tantárgy: CITOGENETIKA Kód: AOMBCGE3 ECTS Kredit: 3 A tantárgyat oktató intézet: TTK Mikrobiális Biotechnológiai és Sejtbiológiai Tanszék A tantárgy felvételére ajánlott félév: 3. Melyik félévben
Kar: TTK Tantárgy: CITOGENETIKA Kód: AOMBCGE3 ECTS Kredit: 3 A tantárgyat oktató intézet: TTK Mikrobiális Biotechnológiai és Sejtbiológiai Tanszék A tantárgy felvételére ajánlott félév: 3. Melyik félévben
Hátterükben egyetlen gén áll, melynek általában számottevő a viselkedésre gyakorolt hatása, öröklési mintázata jellegzetes.
 Múlt órán: Lehetséges tesztfeladatok: Kitől származik a variáció-szelekció paradigma, mely szerint az egyéni, javarészt öröklött különbségek között a társadalmi harc válogat? Fromm-Reichmann Mill Gallton
Múlt órán: Lehetséges tesztfeladatok: Kitől származik a variáció-szelekció paradigma, mely szerint az egyéni, javarészt öröklött különbségek között a társadalmi harc válogat? Fromm-Reichmann Mill Gallton
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei
 A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
A keringı tumor markerek klinikai alkalmazásának aktuális kérdései és irányelvei A TM vizsgálatok alapkérdései A vizsgálatok célja, információértéke? Az alkalmazás területei? Hogyan válasszuk ki az alkalmazott
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A citológia és a genetika társtudománya Citogenetika A kromoszómák eredetét, szerkezetét, genetikai funkcióját,
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A citológia és a genetika társtudománya Citogenetika A kromoszómák eredetét, szerkezetét, genetikai funkcióját,
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai
 Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak
Az áramlási citométer és sejtszorter felépítése és működése, diagnosztikai alkalmazásai Az áramlási citométer és sejtszorter felépítése és működése Kereskedelmi forgalomban kapható készülékek 1 Fogalmak
A növény inváziójában szerepet játszó bakteriális gének
 A növény inváziójában szerepet játszó bakteriális gének merisztéma korai szimbiotikus zóna késői szimbiotikus zóna öregedési zóna gyökér keresztmetszet NODULÁCIÓ növényi jel Rhizobium meliloti rhizobium
A növény inváziójában szerepet játszó bakteriális gének merisztéma korai szimbiotikus zóna késői szimbiotikus zóna öregedési zóna gyökér keresztmetszet NODULÁCIÓ növényi jel Rhizobium meliloti rhizobium
Az érett T-sejtes lymphomák funkcionális patogenezise Krenács László PhD Részletes beszámoló a zárójelentéshez KON-46663
 Az érett T-sejtes lymphomák funkcionális patogenezise Krenács László PhD Részletes beszámoló a zárójelentéshez KON-46663 A perifériás (vagy post-thymicus) T-sejtes lymphomák (PTL) az érett T-sejtek daganatai,
Az érett T-sejtes lymphomák funkcionális patogenezise Krenács László PhD Részletes beszámoló a zárójelentéshez KON-46663 A perifériás (vagy post-thymicus) T-sejtes lymphomák (PTL) az érett T-sejtek daganatai,
mtorc1 and C2 komplexhez köthető aktivitás különbségek és ennek jelentősége humán lymphomákbanés leukémiákban
 mtorc1 and C2 komplexhez köthető aktivitás különbségek és ennek jelentősége humán lymphomákbanés leukémiákban Sebestyén A, Márk Á, Nagy N, Hajdu M, Csóka M*, SticzT, Barna G, Nemes K*, Váradi Zs*, KopperL
mtorc1 and C2 komplexhez köthető aktivitás különbségek és ennek jelentősége humán lymphomákbanés leukémiákban Sebestyén A, Márk Á, Nagy N, Hajdu M, Csóka M*, SticzT, Barna G, Nemes K*, Váradi Zs*, KopperL
Dr. Nemes Nagy Zsuzsa Szakképzés Karl Landsteiner Karl Landsteiner:
 Az AB0 vércsoport rendszer Dr. Nemes Nagy Zsuzsa Szakképzés 2011 Az AB0 rendszer felfedezése 1901. Karl Landsteiner Landsteiner szabály 1901 Karl Landsteiner: Munkatársai vérmintáit vizsgálva fedezte fel
Az AB0 vércsoport rendszer Dr. Nemes Nagy Zsuzsa Szakképzés 2011 Az AB0 rendszer felfedezése 1901. Karl Landsteiner Landsteiner szabály 1901 Karl Landsteiner: Munkatársai vérmintáit vizsgálva fedezte fel
I. MELLÉKLET ALKALMAZÁSI ELŐÍRÁS
 I. MELLÉKLET ALKALMAZÁSI ELŐÍRÁS 1 1. A GYÓGYSZER MEGNEVEZÉSE VPRIV 200 egység por oldatos infúzióhoz. 2. MINŐSÉGI ÉS MENNYISÉGI ÖSSZETÉTEL Egy injekciós üveg 200 egység* velagluceráz-alfát tartalmaz.
I. MELLÉKLET ALKALMAZÁSI ELŐÍRÁS 1 1. A GYÓGYSZER MEGNEVEZÉSE VPRIV 200 egység por oldatos infúzióhoz. 2. MINŐSÉGI ÉS MENNYISÉGI ÖSSZETÉTEL Egy injekciós üveg 200 egység* velagluceráz-alfát tartalmaz.
GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN
 GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN PhD értekezés Dr. Fülöp Zsolt Ferenc Témavezeto: Dr. Matolcsy András Semmelweis Egyetem Patológiai
GENETIKAI INSTABILITÁS VIZSGÁLATA A B-SEJTES KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA RICHTER TRANSZFORMÁCIÓJA SORÁN PhD értekezés Dr. Fülöp Zsolt Ferenc Témavezeto: Dr. Matolcsy András Semmelweis Egyetem Patológiai
TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA. Dr. Kurgyis Zsuzsanna
 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA PhD tézis Dr. Kurgyis Zsuzsanna Bőrgyógyászati és Allergológiai Klinika Szegedi Tudományegyetem Szeged 2017 2 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA
TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA TUMOR-SZTRÓMA SEJTFÚZIÓ HATÁSÁRA PhD tézis Dr. Kurgyis Zsuzsanna Bőrgyógyászati és Allergológiai Klinika Szegedi Tudományegyetem Szeged 2017 2 TUMORSEJTEK FENOTÍPUS-VÁLTOZÁSA
Populációgenetikai. alapok
 Populációgenetikai alapok Populáció = egyedek egy adott csoportja Az egyedek eltérnek egymástól morfológiailag, de viselkedésüket tekintve is = genetikai különbségek Fenotípus = külső jellegek morfológia,
Populációgenetikai alapok Populáció = egyedek egy adott csoportja Az egyedek eltérnek egymástól morfológiailag, de viselkedésüket tekintve is = genetikai különbségek Fenotípus = külső jellegek morfológia,
A hólyagrák aktuális kérdései a pathologus szemszögéből. Iványi Béla SZTE Pathologia
 A hólyagrák aktuális kérdései a pathologus szemszögéből Iványi Béla SZTE Pathologia Hólyagrák 7. leggyakoribb carcinoma Férfi : nő arány 3.5 : 1 Csúcsa: 60-80 év között Oka: jórészt ismeretlen Rizikótényezők:
A hólyagrák aktuális kérdései a pathologus szemszögéből Iványi Béla SZTE Pathologia Hólyagrák 7. leggyakoribb carcinoma Férfi : nő arány 3.5 : 1 Csúcsa: 60-80 év között Oka: jórészt ismeretlen Rizikótényezők:
A köpenysejtes limfómákról
 A köpenysejtes limfómákról Dr. Schneider Tamás Országos Onkológiai Intézet Lymphoma Centrum Limfóma Világnap 2014. szeptember 15. Limfómám van! Limfómám van! Igen? De milyen típusú? Gyógyítható? Nem gyógyítható?
A köpenysejtes limfómákról Dr. Schneider Tamás Országos Onkológiai Intézet Lymphoma Centrum Limfóma Világnap 2014. szeptember 15. Limfómám van! Limfómám van! Igen? De milyen típusú? Gyógyítható? Nem gyógyítható?
ADATBÁNYÁSZAT I. ÉS OMICS
 Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ADATBÁNYÁSZAT
Az élettudományi-klinikai felsőoktatás gyakorlatorientált és hallgatóbarát korszerűsítése a vidéki képzőhelyek nemzetközi versenyképességének erősítésére TÁMOP-4.1.1.C-13/1/KONV-2014-0001 ADATBÁNYÁSZAT
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE
 Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
Az immunrendszer működésében résztvevő sejtek Erdei Anna Immunológiai Tanszék ELTE Tanárszakosok, 2017. Bev. 2. ábra Az immunválasz kialakulása 3.1. ábra A vérsejtek képződésének helyszínei az élet folyamán
HAPMAP -2010 Nemzetközi HapMap Projekt. SNP GWA Haplotípus: egy kromoszóma szegmensen lévő SNP mintázat
 HAPMAP -2010 Nemzetközi HapMap Projekt A Nemzetközi HapMap Project célja az emberi genom haplotípus* térképének(hapmap; haplotype map) megszerkesztése, melynek segítségével katalogizálni tudjuk az ember
HAPMAP -2010 Nemzetközi HapMap Projekt A Nemzetközi HapMap Project célja az emberi genom haplotípus* térképének(hapmap; haplotype map) megszerkesztése, melynek segítségével katalogizálni tudjuk az ember
Az FNA a leggyakrabban használt módszer a pajzsmirigy nodulusok tisztázásában Az FNA használata előtt a pajzsmirigy műtétek 14%-ban volt malignitás
 Kovács Ilona Az FNA a leggyakrabban használt módszer a pajzsmirigy nodulusok tisztázásában Az FNA használata előtt a pajzsmirigy műtétek 14%-ban volt malignitás (Hamberger et al. 1982) FNA használata mellett
Kovács Ilona Az FNA a leggyakrabban használt módszer a pajzsmirigy nodulusok tisztázásában Az FNA használata előtt a pajzsmirigy műtétek 14%-ban volt malignitás (Hamberger et al. 1982) FNA használata mellett
Follicularis lymphomák komplex diagnosztikája
 Follicularis lymphomák komplex diagnosztikája Eredeti közlemény Tóth Erika, 1 Schneider Tamás, 2 Melegh Zsombor, 1 Csernák Erzsébet, 1 Udvarhelyi Nóra, 3 Rosta András, 2 Szentirmay Zoltán 1 Országos Onkológiai
Follicularis lymphomák komplex diagnosztikája Eredeti közlemény Tóth Erika, 1 Schneider Tamás, 2 Melegh Zsombor, 1 Csernák Erzsébet, 1 Udvarhelyi Nóra, 3 Rosta András, 2 Szentirmay Zoltán 1 Országos Onkológiai
Áramlási citometria, sejtszeparációs technikák. Immunológiai és Biotechnológiai Intézet ÁOK, PTE
 Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Áramlási citometria, sejtszeparációs technikák Immunológiai és Biotechnológiai Intézet ÁOK, PTE Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára
Kryoglobulinaemia kezelése. Domján Gyula. Semmelweis Egyetem I. Belklinika. III. Terápiás Aferezis Konferencia, 2014. Debrecen
 Kryoglobulinaemia kezelése Domján Gyula Semmelweis Egyetem I. Belklinika Mi a kryoglobulin? Hidegben kicsapódó immunglobulin Melegben visszaoldódik (37 C-on) Klasszifikáció 3 csoport az Ig komponens
Kryoglobulinaemia kezelése Domján Gyula Semmelweis Egyetem I. Belklinika Mi a kryoglobulin? Hidegben kicsapódó immunglobulin Melegben visszaoldódik (37 C-on) Klasszifikáció 3 csoport az Ig komponens
A kromoszómák kialakulása előtt a DNS állomány megkettőződik. A két azonos információ tartalmú DNS egymás mellé rendeződik és egy kromoszómát alkot.
 Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
Kromoszómák, Gének A kromoszóma egy hosszú DNS szakasz, amely a sejt életének bizonyos szakaszában (a sejtosztódás előkészítéseként) tömörödik, így fénymikroszkóppal láthatóvá válik. A kromoszómák két
A tüdőcitológia jelentősége a tüdődaganatok neoadjuváns kezelésének tervezésében
 A tüdőcitológia jelentősége a tüdődaganatok neoadjuváns kezelésének tervezésében Pápay Judit SE, I.sz.Patológiai és Kísérleti Rákkutató Intézet 70. Patológus Kongresszus Siófok 2011. szept. 29 - okt.1.
A tüdőcitológia jelentősége a tüdődaganatok neoadjuváns kezelésének tervezésében Pápay Judit SE, I.sz.Patológiai és Kísérleti Rákkutató Intézet 70. Patológus Kongresszus Siófok 2011. szept. 29 - okt.1.
DNS molekulák elválasztása agaróz gélelektroforézissel és kapilláris elektroforézissel
 DNS molekulák elválasztása agaróz gélelektroforézissel és kapilláris elektroforézissel Gyakorlat helye: BIOMI Kft. Gödöllő, Szent-Györgyi A. u. 4. (Nemzeti Agrárkutatási és Innovációs Központ épülete volt
DNS molekulák elválasztása agaróz gélelektroforézissel és kapilláris elektroforézissel Gyakorlat helye: BIOMI Kft. Gödöllő, Szent-Györgyi A. u. 4. (Nemzeti Agrárkutatási és Innovációs Központ épülete volt
Adaptív/anticipatív immunrendszer. Egyedi, klonális receptorok. szignáljainak kontrollja alatt áll
 Adaptív/anticipatív immunrendszer Egyedi, klonális receptorok A természetes immunrendszer DANGER szignáljainak kontrollja alatt áll EHRLICH oldallánc-elmélete Alapfogalmak: antigén epitop haptén karrier
Adaptív/anticipatív immunrendszer Egyedi, klonális receptorok A természetes immunrendszer DANGER szignáljainak kontrollja alatt áll EHRLICH oldallánc-elmélete Alapfogalmak: antigén epitop haptén karrier
Egyénre szabott terápiás választás myeloma multiplexben
 Egyénre szabott terápiás választás myeloma multiplexben Dr. Mikala Gábor Főv. Önk. Egy. Szt István és Szent László Kórház Haematológiai és Őssejt-Transzplantációs Osztály Mit kell tudni a myelomáról? Csontvelőből
Egyénre szabott terápiás választás myeloma multiplexben Dr. Mikala Gábor Főv. Önk. Egy. Szt István és Szent László Kórház Haematológiai és Őssejt-Transzplantációs Osztály Mit kell tudni a myelomáról? Csontvelőből
11. Dr. House. Biokémiai és sejtbiológiai módszerek alkalmazása az orvoslásban
 11. Dr. House. Biokémiai és sejtbiológiai módszerek alkalmazása az orvoslásban HIV fertőzés kimutatása (fiktív) esettanulmány 35 éves nő, HIV fertőzöttség gyanúja. Két partner az elmúlt időszakban. Fertőzött-e
11. Dr. House. Biokémiai és sejtbiológiai módszerek alkalmazása az orvoslásban HIV fertőzés kimutatása (fiktív) esettanulmány 35 éves nő, HIV fertőzöttség gyanúja. Két partner az elmúlt időszakban. Fertőzött-e
PrenaTest Újgenerációs szekvenálást és z-score
 PrenaTest Újgenerációs szekvenálást és z-score számítást alkalmazó, nem-invazív prenatális molekuláris genetikai teszt a magzati 21-es triszómia észlelésére, anyai vérből végzett DNS izolálást követően
PrenaTest Újgenerációs szekvenálást és z-score számítást alkalmazó, nem-invazív prenatális molekuláris genetikai teszt a magzati 21-es triszómia észlelésére, anyai vérből végzett DNS izolálást követően
KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA SEJTEK ÉS A MIKORKÖRNYEZET KÖLCSÖNHATÁSÁNAK JELENTŐSÉGE A BETEGSÉG PROGRESSZIÓJÁBAN. Dr.
 KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA SEJTEK ÉS A MIKORKÖRNYEZET KÖLCSÖNHATÁSÁNAK JELENTŐSÉGE A BETEGSÉG PROGRESSZIÓJÁBAN Doktori Tézisek Dr. Plander Márk Semmelweis Egyetem Patológiai Tudományok Doktori Iskola
KRÓNIKUS LYMPHOCYTÁS LEUKÉMIA SEJTEK ÉS A MIKORKÖRNYEZET KÖLCSÖNHATÁSÁNAK JELENTŐSÉGE A BETEGSÉG PROGRESSZIÓJÁBAN Doktori Tézisek Dr. Plander Márk Semmelweis Egyetem Patológiai Tudományok Doktori Iskola
Áramlási citometria 2011. / 4. Immunológiai és Biotechnológiai Intézet PTE KK
 Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
Áramlási citometria 2011. / 4 Immunológiai és Biotechnológiai Intézet PTE KK Az áramlási citometria elve Az áramlási citometria ( Flow cytometria ) sejtek gyors, multiparaméteres vizsgálatára alkalmas
A sejtes mikrokörnyezet szerepe a follicularis lymphoma csontvelői terjedésében
 A sejtes mikrokörnyezet szerepe a follicularis lymphoma csontvelői terjedésében Doktori értekezés Dr. Semmelweis Egyetem Patológiai Tudományok Doktori Iskola Témavezető: Dr. Matolcsy András egyetemi tanár,
A sejtes mikrokörnyezet szerepe a follicularis lymphoma csontvelői terjedésében Doktori értekezés Dr. Semmelweis Egyetem Patológiai Tudományok Doktori Iskola Témavezető: Dr. Matolcsy András egyetemi tanár,
MUTÁCIÓK. A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik.
 MUTÁCIÓK A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik. Pontmutáció: A kromoszóma egy génjében pár nukleotidnál következik be változás.
MUTÁCIÓK A mutáció az örökítő anyag spontán, maradandó megváltozása, amelynek során új genetikai tulajdonság keletkezik. Pontmutáció: A kromoszóma egy génjében pár nukleotidnál következik be változás.
Hasi tumorok gyermekkorban
 Hasi tumorok gyermekkorban Dr. Bartyik Katalin SZTE ÁOK Gyermekgyógyászati Klinika és Gyermekegészségügyi Központ, Szeged A gyermekkori tumorok a halálozási oki tényezői közül a 2. helyen állnak a balesetek,
Hasi tumorok gyermekkorban Dr. Bartyik Katalin SZTE ÁOK Gyermekgyógyászati Klinika és Gyermekegészségügyi Központ, Szeged A gyermekkori tumorok a halálozási oki tényezői közül a 2. helyen állnak a balesetek,
Katasztrófális antifoszfolipid szindróma
 Katasztrófális antifoszfolipid szindróma Gadó Klára Semmelweis Egyetem, I.sz. Belgyógyászati Klinika Antifoszfolipid szindróma Artériás és vénás thrombosis Habituális vetélés apl antitest jelenléte Mi
Katasztrófális antifoszfolipid szindróma Gadó Klára Semmelweis Egyetem, I.sz. Belgyógyászati Klinika Antifoszfolipid szindróma Artériás és vénás thrombosis Habituális vetélés apl antitest jelenléte Mi
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása
 A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Délpesti Centrumkórház Országos Hematológiai és Infektológiai Intézet, Hematológiai és Őssejttranszplantációs
A B sejtek érése, aktivációja, az immunglobulin osztályok kialakulása Uher Ferenc, PhD, DSc Délpesti Centrumkórház Országos Hematológiai és Infektológiai Intézet, Hematológiai és Őssejttranszplantációs
A csontvelõ-átültetés után kialakuló B-sejt-készlet a korlátozott ellenanyag-sokféleség molekuláris alapjai
 A csontvelõ-átültetés után kialakuló B-sejt-készlet a korlátozott ellenanyag-sokféleség molekuláris alapjai Vas Virág, Pálóczi Katalin dr. és Uher Ferenc dr. Országos Haematológiai és Immunológiai Intézet,
A csontvelõ-átültetés után kialakuló B-sejt-készlet a korlátozott ellenanyag-sokféleség molekuláris alapjai Vas Virág, Pálóczi Katalin dr. és Uher Ferenc dr. Országos Haematológiai és Immunológiai Intézet,
Hivatalos Bírálat Dr. Gődény Mária
 Hivatalos Bírálat Dr. Gődény Mária:,,Multiparametrikus MR vizsgálat prognosztikai és prediktív faktorokat meghatározó szerepe fej-nyaki tumoroknál, valamint a kismedence főbb daganat csoportjaiban című
Hivatalos Bírálat Dr. Gődény Mária:,,Multiparametrikus MR vizsgálat prognosztikai és prediktív faktorokat meghatározó szerepe fej-nyaki tumoroknál, valamint a kismedence főbb daganat csoportjaiban című