KAPOSVÁRI EGYETEM. ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék. A doktori iskola vezetője HORN PÉTER. az MTA rendes tagja.
|
|
|
- Zsuzsanna Barnané
- 8 évvel ezelőtt
- Látták:
Átírás
1 KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője HORN PÉTER az MTA rendes tagja Témavezető NAGY ISTVÁN Tudományos munkatárs A BELTENYÉSZTETTSÉG ÉRTÉKELÉSE A MAGYAR FAJTATISZTA SERTÉSTENYÉSZTÉSI PROGRAMBAN Készítette VIGH ZSÓFIA KAPOSVÁR 2010

2 2
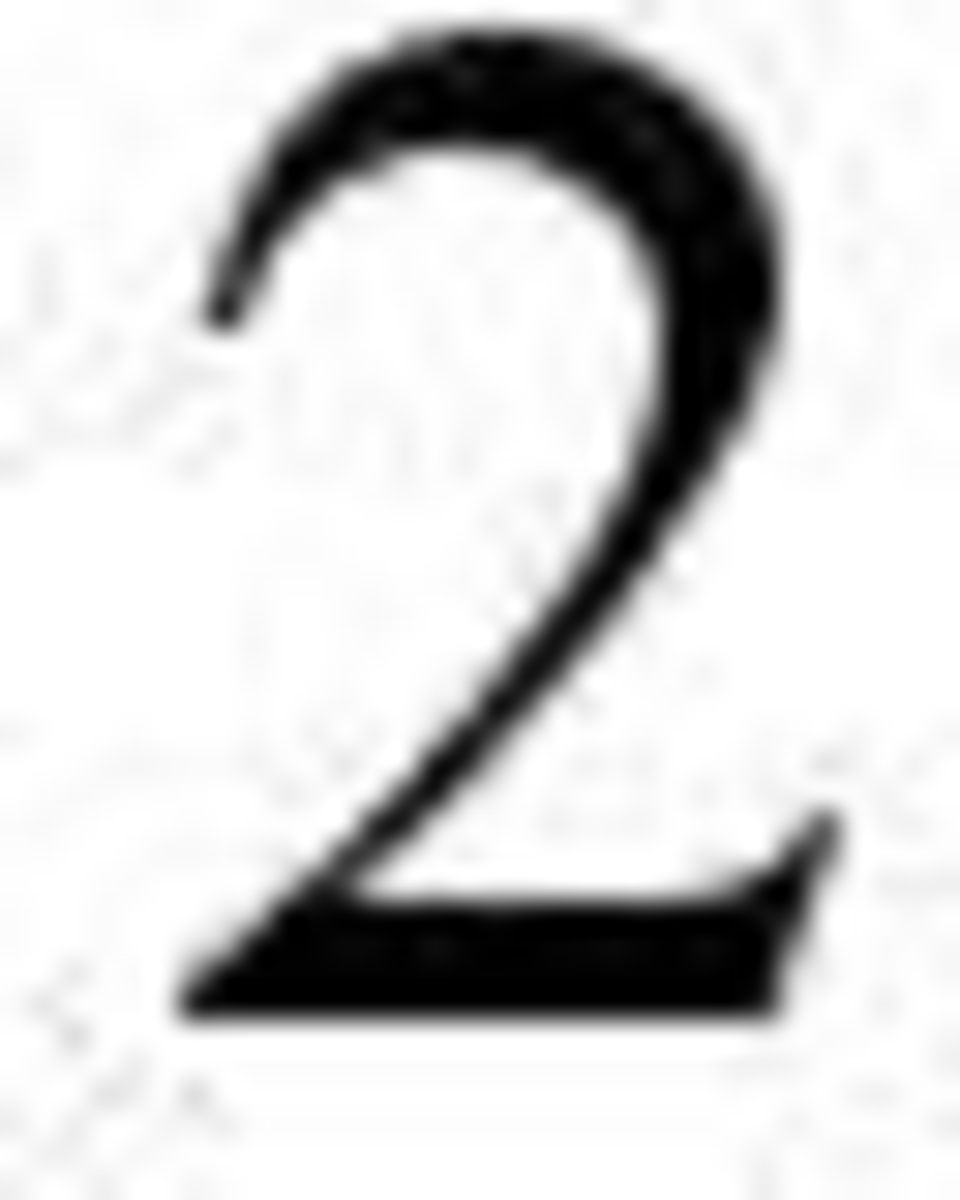
3 TARTALOMJEGYZÉK 1. BEVEZETÉS IRODALMI ÁTTEKINTÉS A beltenyésztés Beltenyésztés célja és kedvező hatásai A beltenyésztéshez közvetetten kapcsolódó mutatók A beltenyésztettség hatása multipara állatfajok reprodukciós és hízékonysági, valamint egyéb tulajdonságaira A beltenyésztettség hatása a hízékonysági és szaporasági tulajdonságokra sertésekben A hazai tenyészértékbecslési rendszer és annak beltenyésztési vonatkozásai A DISSZERTÁCIÓ CÉLKITŰZÉSEI ANYAG ÉS MÓDSZER Adatbázisok ÜSTV Adatok SZFTV adatok Pedigréanalízis, beltenyésztési együtthatók BLUP-modellek MNF fajta üzemi sajátteljesítmény-vizsgálati adatain futtatott modellváltozatok ML fajta üzemi sajátteljesítmény-vizsgálati adatain futtatott modellváltozatok A szaporasági és felnevelési teljesítmény-vizsgálati adatokon futtatott modellváltozatok EREDMÉNYEK ÉS ÉRTÉKELÉSÜK Pedigréanalízis Magyar nagyfehér hússertés Magyar lapálysertés Hízékonysági és vágási tulajdonságok Színhús % és egy életnapra jutó testsúly-gyarapodás Karajátmérő, szalonna 1 és szalonna 2 vastagság Szaporasági tulajdonságok MSE Beltenyésztési leromlás KÖVETKEZTETÉSEK, JAVASLATOK ÚJ TUDOMÁNYOS EREDMÉNYEK ÖSSZEFOGLALÁS

4 9. SUMMARY KÖSZÖNETNYILVÁNÍTÁS IRODALOMJEGYZÉK A DISSZERTÁCIÓ TÉMAKÖRÉBŐL MEGJELENT PUBLIKÁCIÓK Idegen nyelven megjelent tudományos közlemények Magyar nyelven megjelent tudományos közlemények Előadások idegen nyelvű konferenciákon Ismeretterjesztő közlemények A DISSZERTÁCIÓ TÉMAKÖRÉN KÍVÜLI PUBLIKÁCIÓK Idegen nyelven megjelent közlemények Magyar nyelven megjelent közlemények Proceedings-ben teljes terjedelemben megjelent közlemények Proceedings-ben megjelent magyar nyelvű közlemények Ismeretterjesztő közlemények SZAKMAI ÉLETRAJZ

5 1. BEVEZETÉS Az eredményes, jövedelmező állattenyésztés egyik alapja az új, korszerű módszerek alkalmazása mind a tenyészutánpótlás, mind pedig a végtermékelőállítás tekintetében. Hazánk sertéslétszáma a 90 -es évektől folyamatosan csökken, az egykori 10 millió egyed helyett 2008-ban már kevesebb, mint darabot tartottak számon (SZABÓ, 2009), pedig hagyományosan sok és sokféle sertéshúst fogyasztunk. Éppen ezért a tenyésztők alapvető érdeke az, hogy termelékenységüket a minőség folyamatos javításával fokozzák, melynek az egyik legfontosabb eszköze a nemesítés. A szelekció hatékonyságának és a genetikai előrehaladás mértékének növelése nem valósulhat meg pontos tenyészértékbecslő eljárás használata nélkül. Sertéstenyésztésünkben a tenyészérték-becslés alapvető feltételei, a teljesítményvizsgálatok és az adatgyűjtés, valamint a megfelelő matematikai-statisztikai módszerek már az 1990-as évektől megvalósultak (FARKAS, 2008). Rendelkezésre állnak a származásra vonatkozó, az üzemi sajátteljesítmény vizsgálati-, a központi vizsgálóállomási- és a reprodukciós adatok is több, mint 20 évre visszamenően, január 1-től pedig a tenyészérték-becslés és a szelekciós döntések az összetett BLUP-modellen alapuló indexen, az. ún. teljes tenyészérték index TT alapján történnek (MGSZH, 2009) Több tenyésztési integrációban (pl. KA-HYB) szerepet játszott a beltenyésztett vonalak kialakítása és a beltenyésztett egyedek felhasználása a tenyésztésben, bizonyos értékes gének rögzítése érdekében. A teljesítmény ily módon való fokozása azonban veszélyeket is rejthet magában, hiszen egy bizonyos szint felett a beltenyésztés már nem javítja, hanem rontja a szaporasági és hízékonysági eredményeket, azonban ez a szint fajonként, fajtánként, populációnként és akár egyedenként is eltérő lehet. Bár a 5

6 közelmúltban született átfogó tanulmány (FARKAS, 2008) komplex módon feldolgozta a hazai sertésállomány tenyészérték-becslési rendszerét, azonban a beltenyésztés kérdéskörében hazánkban eddig nem végeztek részletes kutatást. 6

7 2. IRODALMI ÁTTEKINTÉS 2.1. A beltenyésztés A beltenyésztés olyan egyedek párosítását jelenti, melyeknek egy vagy több őse közös (rokon egyedek). A rokonok részben azonos alléleket hordoznak, ezért utódaik nagyobb valószínűséggel lesznek homozigóták, mint véletlenszerű párosodás mellett. A nagymértékben homozigóta egyedek szervezetének élettani pufferkapacitása csökkent, ezért környezeti labilitásuk nagyobb, alkalmazkodóképességük kisebb. Mindez - főként, ha káros hatású recesszív gének homozigóta állapotban való manifesztációjával párosul - a beltenyésztési leromlásban jut kifejezésre (DOHY, 1989). Két egyed rokoni kapcsolata az állattenyésztés gyakorlatában azt jelenti, hogy egy vagy több közös ősük volt, amely közös ősüktől mindketten azonos géneket örökölhettek, ebből következően génkészletük egy része származásilag azonos lehet. A rokonság fokát a származásilag potenciálisan azonos génhányad nagysága határozza meg. Ez pedig a rokonok és a közös ős vagy ősök közötti nemzedékek számától függ (SZABÓ, 2004; DOHY, 1999). A magyar szakirodalomban találhatóak olyan megközelítések, mely szerint a rokontenyésztés a beltenyésztésnek egy magasabb fokát jelenti. Mindkét eljárás (a bel és rokontenyésztés) azonos irányba: a homozigozitás növekedésének irányába hat. A különbség köztük csupán a származásilag azonos gének homozigozitásának mértékében jelentkezik (SZABÓ, 2004; DOHY, 1999). Ugyanakkor a nemzetközi szakirodalomra ez a fajta megkülönböztetés nem jellemző. 7
.")
8 A beltenyésztés eredményeként bekövetkező homozigócia számszerű kifejezésére a legelterjedtebb mérőszám a Wright-féle beltenyésztési együttható (F). (SZABÓ, 2004; DOHY, 1999). Az F annak a valószínűsége, hogy az egyed valamely lókuszában lévő két gén származásilag azonos. A beltenyésztéssel nem csak a kívánatos, hanem a nemkívánatos géneket is felhalmozhatjuk és homozigóta állapotban fixálhatjuk a populációban (SZABÓ, 2004). A beltenyésztés káros hatásai már századok óta ismertek. DARWIN (1868; 1876) 57 növényfajt vizsgált 52 nemzetségből és 30 családból. A beltenyésztett növények átlagosan alacsonyabbak voltak, kisebb volt a súlyuk, később virágoztak és kevesebb magot hoztak. Az önbeporzó növények átlagosan 41 % csökkenést mutattak a maghozamban és 13 %-os elmaradást a magasságban. A beltenyésztés kérdéskörének elméleti alapjait WRIGHT (1922) dolgozta ki. Sok éves kutatás, illetve az evolúcióban, a mezőgazdaságban és a génmegőrzésben levő jelentősége ellenére a beltenyésztési leromlás genetikai alapja máig nem tisztázott kérdéskör (LYNCH és WALSH, 1998). Jelenleg két alaphipotézis létezik az evolúció szempontjából teljesen eltérő következményekkel. Az első hipotézis az ún. dominancia elmélet (CROW, 1952), mely azt feltételezi, hogy az egyes populációkban a beltenyésztési leromlást a recesszív gének okozzák, melyek arányát a szelekció-mutáció egyensúlya határozza meg. A második feltevés (CROW, 1952) szerint a beltenyésztési leromlást a heterozigozitás csökkenése okozza. Szintén megoldatlan kérdéskör a beltenyésztési leromlás heterogenitása (a nagy heterogenitás azt a feltevést támasztja alá, hogy a beltenyésztési leromlást kisszámú overdomináns lókusz okozza), melynek kifejezése a beltenyésztés káros hatásainak megértéséhez, illetve azok leküzdéséhez kiindulópontként szolgálhat. A beltenyésztési leromlás heterogenitását bizonyították 8
 57 növényfajt vizsgált 52 nemzetségből és 30 családból.")
9 tejtermelő-képességgel kapcsolatos tulajdonságok esetében tejelő szarvasmarha (MIGLIOR és mtsai, 1992), illetve sertések, valamint Tribolium castaneum (lisztbogár) reprodukciós tulajdonságaiban (HOHENBOKEN és mtsai, 1991). A populáció középértékének csökkenése olyan mennyiségi tulajdonságok esetében, mint az alomnagyság, a termékenység, az életképesség vagy a fitnesz a beltenyésztés hátrányos következménye lehet, melyet beltenyésztési leromlásnak nevezünk. A jelenséget több növény- és állatfajban (WRIGHT, 1977; LYNCH és WALSH, 1998), illetve humán vonatkozásban is megfigyelték (SCHULL, 1962). Számos szerző tapasztalt beltenyésztési leromlást különböző domesztikált állatfajok (BERESKIN és mtsai, 1968; MIGLIOR és mtsai, 1995; WIENER és mtsai, 1992), vadon élő állatfajok (HEDRICK és KALINOWSKI, 2000), fogságban tartott (nem domesztikált) állatfajok (CASSINELLO, 2005; KALINOWSKI és HEDRICK, 2001), valamint kísérleti állatfajok (HOLT és mtsai, 2005/a; HOLT és mtsai, 2005/b; WHITE, 1972) populációiban. A vadon élő kis létszámú, veszélyeztetett populációkban a beltenyésztettség tovább növeli a kihalás veszélyét (FRANKHAM, 2003). A domesztikált állatfajok esetében a leromlás leginkább a fitnesz tulajdonságok esetében figyelhető meg (FALCONER és MACKAY, 1996). A fitnesz tulajdonságoknál a beltenyésztés káros hatását erősíti, hogy az gyakran több értékmérőben egyszerre okoz teljesítménycsökkenést (pl. fialáskori alomnagyság, szopóskori elhullás). Ezen kívül a szaporasági teljesítménycsökkenés az anyát (ovulációs ráta) és az utódot (magzatkori elhullás) külön-külön érinti. A beltenyésztéssel kapcsolatos leromlás megnyilvánulásában tapasztalt nagy változékonyságot részben az okozhatja, hogy a káros allélek gyakorisága tulajdonságonként és populációnként eltérő lehet, amit az eltérő szelekciós nyomás okozhat (LYNCH és WALSH, 9
, illetve humán vonatkozásban is megfigyelték (SCHULL, 1962).")
10 1998). A változékonyságot azonban részben a beltenyésztési leromlásra vonatkozó becslési nehézségek is magyarázhatják (CURIK és mtsai, 2001). Ezek a statisztikai próba kis ereje, a kis mintanagyság és a különböző beltenyésztettségi szintekhez kapcsolódó heterogén varianciák lehetnek. A becsült beltenyésztési együtthatókat ezen kívül nagymértékben befolyásolja a pedigré hossza, illetve annak teljessége (KOENIG és SIMIANER, 2006). A beltenyésztési leromláson kívül a beltenyésztés, illetve a véletlen drift a genetikai és környezeti varianciák megváltozásához vezethet. Additív modell esetén feltételezve, hogy az allélek neutrálisak, a véletlen drift az additív genetikai variancia vonalakon belüli csökkenését okozza. A csökkenés mértéke a vonalakon belüli átlagos beltenyésztési együtthatóval arányos (FALCONER és MACKAY, 1996). A fenti jelenség főleg a fitnessszel szorosan nem kapcsolódó tulajdonságok esetében jellemző. A fitnesz tulajdonságokat azonban a nem additív génhatások is jelentősen befolyásolhatják. Ebben az esetben a genetikai variancia a beltenyésztés hatására átmenetileg akár növekedhet is (KRISTENSEN és mtsai, 2005). Ezzel párhuzamosan azonban jelentős beltenyésztési leromlás léphet fel, amit vélhetően az okozhat, hogy a genetikai variancia növekedését a káros recesszív allélek (mutáció) okozzák, melyek egyben a leromlásért is felelősek (KRISTENSEN és SORENSEN, 2005). Elméleti és empirikus bizonyítékok alapján a beltenyésztésnek számos káros hatása van, melyeket FRANKHAM (2003) foglalt össze. A beltenyésztés szaporasági teljesítményre, valamint mortalitásra gyakorolt hatása régóta ismert. Az édestestvér párosítások átlagosan 33 %-kal csökkentik a magzatkori életben maradást. Azok a populációk, melyeknek (folyamatos édestestvér párosítások mellett) az átlagos beltenyésztési együtthatója eléri a 80 %-ot, %-os arányban kiesnek a termelésből. A szisztematikus édestestvér párosítás a beltenyésztési együtthatók rendkívül 10

11 gyors populáció szintű növekedését okozza, ezért azt a domesztikált állatfajokban (eltekintve esetleges kísérleti céloktól) nem alkalmazzák. Amennyiben a beltenyésztési együttható növekedése folyamatos, de lassú ütemű, a természetes szelekció nagyobb eséllyel távolítja el a káros allélokat a populációból. Azonban még a lassú ütemű növekedés mellett is a populációk nagy arányban (mintegy 50 %-ban) kiesnek a termelésből, ha az átlagos beltenyésztési együttható megközelíti a korábban említett 80 %-ot. A környezeti változások (eddig ismeretlen betegségek, paraziták, környezetszennyezés, éghajlati változások stb.), folyamatosan befolyásolják a populációkat. Az új környezeti hatásokra adott "válasz" alapja a populációban rejlő genetikai változékonyság. A genetikailag különböző egyedek ezért eltérő módon reagálnak az új környezeti hatásokra. Az úgynevezett random drift a populáció átlagos beltenyésztési együtthatójával arányosan csökkenti az additív genetikai varianciát, mely megnehezíti a környezeti változásokhoz történő alkalmazkodást. A genetikai variancia csökkenése, valamint a betegségekre való hajlam, továbbá a parazitákkal szembeni csökkent ellenálló képesség kapcsolatát már számos fajra vonatkozóan megállapították Beltenyésztés célja és kedvező hatásai A beltenyésztés célja a kívánatos gének homozigóta formában való rögzítése (fixálása) a populációban. Így a beltenyésztésnek két alapvető genetikai hatása van, egyrészt a homozigozitás növekedése és a heterozigozitás csökkenése, másrészt az eredeti állományokon belül egymástól genetikailag jelentősen különböző részállományok, ún. beltenyésztett vonalak (családok) kialakítása. A beltenyésztés előidézi a genetikai variancia megváltozását, ezáltal a vonalakon belül mérséklődik, a 11
, folyamatosan befolyásolják a populációkat.")
12 vonalak között növekszik a genetikai variancia. A célszerűen beltenyésztett vonalak keresztezésével a tenyésztő előnyt tud szerezni a heterozigóta F 1 ivadékok genotípusos és fenotípusos egyöntetűségéből, ha az állományt a piacon azonos időben és közel azonos súlyban szükséges értékesíteni. A tenyésztő másik előnye, hogy egy beltenyésztett vonal keresztezéséből domináns génhatás esetében a nem rokontenyésztett szülők teljesítményét felülmúló heterozigóta ivadékok származnak. A beltenyésztés haszna az is, hogy valószínűsíti a káros recesszív gének egy lókuszra kerülését, így lehetővé válik az ilyen géneket hordozó egyedek selejtezése (SZABÓ, 2004). Több hibrid sertésfajta kialakításában és fenntartásában játszottak szerepet beltenyésztett vonalak, erre egy példa a Kaposvári Mezőgazdasági Főiskolán kitenyésztett és 1972-től államilag elismert KA-HYB fajta. Az 1970-es években mintegy 120 ezer termelő tartotta hazánkban és az évi hízósertés-kapacitás elérte a háromnegyed milliót (ANKER, 1973/a), ban pedig mintegy 1,6 millió KA-HYB sertés került vágásra (BALTAY, 1983). A fajta tenyésztése évekkel ezelőtt megszűnt. A KA-HYB vonalak részben rokontenyésztettek, ahol a rokontenyésztés lehetőleg a fajta, illetve állomány legjobb kanjára épül. Ezekben a vonalakban általában a kocák csökkent szaporaságával kell számolni, bár azonos rokontenyésztettségi koefficiensű kocák között nagy különbségek mutatkoznak az anyai teljesítmény tekintetében (HORN és mtsai, 1984). A KA-HYB kontinuens hibridsertést négy alapvonal (fajta) felhasználásával alakították ki: - az anyai populációt svéd nagy fehér hússertés szelektált, zárt vonalának és az angol lapálysertés mérsékelten rokontenyésztett vonalának keresztezésével; 12

13 - az apai populációt rokontenyésztett holland lapálysertés és angol nagy fehér vonalak keresztezésével állították elő (BALTAY, 1983). ANKER és RADNAI (1977) szerint a rokontenyésztés biztosítja a kanok megbízható örökítőképességét és lehetőséget ad, hogy a szelekcióval fokozható tulajdonságok terén az apaállatokban minél nagyobb értékben koncentrálódjék az értékes örökléses anyag. Rokontenyésztés kizárólag a KA-HYB kanvonalain belül történt, a termelésben soha. A vonalalapítók elsősorban additív tulajdonságokban kiváló teljesítményeit rokontenyésztéssel rögzítik azokban a vonalakban, ahol ez célszerűnek látszik (ANKER, 1973/b), majd az erősen rokontenyésztett és a kevésbé rokontenyésztett apaállatokat felexibilis módon váltogatták az újabb nemzedékek előállításakor. A már késznek tekinthető vonalakat tesztelték a kombinációkészség szempontjából, így az ilyen módon jelentkező legjobbak megbízhatóan reprodukálhatóak, mert minden vonalban megfelelő számú, ugyanazzal a genotípussal rendelkező egyed van (ANKER, 1970). A KA-HYB hibridsertés előállítás érdekében végzett kívánatos rokontenyésztettségi szintet ANKER (1970) az anyai alapvonalak esetében 0,10-0,15 körülinek, az apai alapvonalaknál pedig 0,25-0,30 körülinek tartotta. A hibridizálásban hagyományossá vált édestestvér-párosítási módszer helyett tapasztalata alapján a szülő-utód párosítás módszerét tartja célravezetőbbnek, mert így kevesebb rokontenyésztett nemzedék váltására van szükség egy kívánt mértékű homozigozitás eléréséhez, mint a testvérpárosításosnál, így az időtényező szempontjából is előnyösebb (ANKER, 1974). 13
, majd az erősen")
14 2.3. A beltenyésztéshez közvetetten kapcsolódó mutatók Beltenyésztési együttható (F, WRIGHT, 1922) Ugyanazon lókuszban származásilag azonos allélpárt hozdozó egyedek relatív gyakoriságát a populációban a beltenyésztési koefficiens fejezi ki. A beltenyésztési koefficiens egy adott lókuszra vonatkozólag annak a valószínűsége, hogy az egész populáció összes vonalaiból véletlenül kivett egyed a kérdéses lókuszban identikus homozigóta, azaz beltenyésztett. Egyedre kivetítve a beltenyésztési koefficiens annak valószínűségét fejezi ki, hogy a vizsgált egyed (mint valamely populáció véletlenszerűen kiválasztott tagja) a kérdéses lókuszban beltenyésztett. A beltenyésztési együttható függ a pedigré hosszától és teljességétől (BOICHARD és mtsai, 1997), tehát minél teljesebb és több nemzedékre visszavezethető pedigré áll rendelkezésre, annál megbízhatóbb a koefficiens. A program(csomag)ok ezt a paramétert VANRADEN (1992) és/vagy MEUWISSEN és LUO (1992) által készített algoritmus alapján számolják, ami egy rokonsági mátrix számítása. Effektív populációméret (Ne, WRIGHT, 1931; PIRCHNER, 1968; FALCONER és MACKAY, 1996) Az ún. effektív szám legtöbbször kisebb a tényleges számnál, mely abból adódik, hogy a háziállatok tenyésztésében a tenyészállatok eltérő mértékben járulnak hozzá az utódpopuláció génállományához. A valós populáció beltenyésztési rátája megegyezik egy attól kisebb, ám ideális szerkezetű populációéval. Ez utóbbi populációban lévő egyedek száma jelenti az effektív populációméretet. A beltenyésztési ráta ( F y ) alapján bármely populációban az effektív populációméret (Ne): 14

15 N e 1 2 F y L ahol F y = (F t F t-1 ) / (1 F t-1 ), a populáció beltenyésztettségének évenkénti növekedése, L-pedig a generációs intervallum. Az effektív populációméret rövid pedigréből nem becsülhető pontosan (TE BRAAKE és mtsai, 1994). Rövidnek az 5 teljes generáció ekvivalenst el nem érő pedigrét tekintjük (GUTIÉRREZ és mtsai, 2005). FRANKHAM és mtsai (2002) szerint kritikus effektív populációméretnek tekinthető az 50-es effektív létszám. Ez a szám az állattenyésztők gyakorlati tapasztalatán alapul, akik azt figyelték meg, hogy a szaporaságot érintő szelekció ellensúlyozhatja a beltenyésztési leromlást, amennyiben a beltenyésztési ráta generációnként kisebb, mint 1 % ( F=1/2N e ). Az effektív populációméretet a tenyészállatok ivararánya is befolyásolja. Alapító ősök effektív létszáma (fe, JAMES, 1972, LACY, 1989) Alapító ősök azok az egyedek, melyeknek mindkét szülője már ismeretlen a pedigrében. A populáció teljes génkészlete visszavezethető az alapító ősökig, melyek különböző mértékben járulnak hozzá az állomány genetikai diverzitásához. Ez a paraméter kevésbé érzékeny a pedigré teljességére, mint a beltenyésztési együttható (BOICHARD és mtsai, 1997). Nem alapító ősök effektív létszáma (fa, BOICHARD és mtsai, 1997) Azon ősök minimális létszáma (nem feltétlenül alapító ős), amellyel magyarázható a populáció teljes genetikai diverzitása. A nem alapító ősöket a populációhoz történő genetikai hozzájárulásuk alapján választjuk ki, azonban mivel egyes egyedek nem feltétlenül alapító ősök, ezért a rokonsági 15

16 kapcsolatok miatt a genetikai hozzájárulások között átfedések lehetnek (ezek összessége több lehet, mint 100 %). Ezért a nem alapító ősök esetében a marginális hozzájárulásukat vesszük figyelembe (a genetikai hozzájárulásoknak az átfedés-mentes részét használjuk). A nem alapító ősök létszáma alacsonyabb (vagy egyenlő) az alapító ősök effektív létszámánál. Alapító ősök genom ekvivalens értéke: (Ng, Chevalet és Rochambeau, 1986; MacCluer és mtsai, 1986; Lacy, 1989) Azt mutatja meg, hogy az alapító ősök által meghatározott populációban egy adott allélnek mennyi a fennmaradási valószínűsége (a referencia populációban). A paraméter kiszámításához sztochasztikus módszerre ( gene dropping ) van szükség. Minden alapító ős két egyedi allélt kap ( n ős esetében ez összesen 2n egyedi allél), majd a Mendeli szegregációt a pedigré alapján szimuláljuk egy előre meghatározott ismétlésszámmal (pl szer). Ezt követően a szimulációk összesítésével az allélek várható aránya ismertté válik. Ez a mérőszám minden olyan hatást figyelembe vesz, mely a génkészlet csökkenésében szerepet játszott, ezért az Ng érték általában kisebb, mint az fe és az fa. (Sölkner és mtsai., 1998). Az alapító ősök genom ekvivalens értéke közvetlenül összehasonlítható az fe és fa-val. Nem alapító ősök effektív létszámának és az alapító ősök effektív létszámának aránya (fa/fe arány) A két szám aránya jelzi, hogy a palacknyak effektus mennyire jellemző a vizsgált populációra. Palacknyak effektusnak hívjuk a populáció 16
 Azt mutatja meg, hogy az alapító ősök által meghatározott populációban egy adott allélnek")
17 létszámának és ezáltal a génkészletének beszűkülését. (BOICHARD és mtsai, 1997) Ha fe nagyobb, mint fa, akkor a palacknyak effektus szerepet játszott a populáció fennállása folyamán. (MAIGNEL és mtsai, 1996) Alapító ősök genom ekvivalens értékének és az alapító ősök effektív létszámának aránya (ng/fe arány) A populációkban fennálló drift (génsodródás) kifejezésére használatos arány. A génsodródás a szaporodás során véletlenszerűen bekövetkező allélgyakoriság változás a populációban. (BOICHARD és mtsai, 1997) A különböző szerzők által egyes állományokban becsült beltenyésztési együtthatót és a beltenyésztéshez közvetetten kapcsolódó mutatókat az 1-3. táblázatban tüntetem fel. 17

18 1. táblázat. Ló és szamár fajták esetében közölt beltenyésztési együtthatók (F), pedigré teljességek (Pt) és generációs intervallumok (GI) Szerző és fajta F (%) Pt GI (év) CERVANTES és mtsai (2008), spanyol arab ló 7,0 5,7 - GLAZEWSKA és JEZIERSKI (2004), : 10,24-12,6 lengyel arab ló 3,06-5,31 - : 8,64-13,92 GUTIÉRREZ és mtsai (2005), katalán szamár 3,36 1,96 - MOUREAUX és mtsai (1996) angol telivér francia ügető arab ló anglo-arab selle francais 1,02 1,86 3,08 1,17 0, ,6 11,8 9,7 11,5 11,7 PONCET és mtsai (2006), Franches-Montagnes ló 6,0 12,3-12,4 8,7 POSTA és mtsai (2006), magyar sportló 7,9-10,24 ROYO és mtsai (2007), asturcon ló 4,7 2,97 - VALERA és mtsai (2005), andalúz ló 8, ZECHNER és mtsai (2002), lipicai ló 10,81 15,2-18
, Franches-Montagnes ló 6,0 12,3-12,4 8,7 POSTA")
19 2. táblázat. Szarvasmarha és juh fajták esetében közölt beltenyésztési együtthatók (F), pedigré teljességek (Pt), generációs intervallumok(gi) és effektív populációméretek (Ne) Szerző és fajta F (%) Pt GI (év) Ne BAUMUNG és SÖLKNER (2002), 3 osztrák fajta Tux-Zillertal Carinthian Blond Original Pinzgau ,52 1,73 5, CANON és mtsai (1994) 2 helyi jelentőségű spanyol húsmarha fajta Casina Carrenana 1,2 0, ,3 5,4 - - GUTIÉRREZ és mtsai (2003) 8 helyi jelentőségű spanyol húsmarha fajta Alistana Asturiana de la Montana Asturiana de los Valles Avilena-Negra Iberica Bruna dels Pirineus Morucha Pirenaica Sayaguesa 1,09 1,55 0,48 2,50 0,25 2,20 1,60 3,13 1,53 1,56 1,08 2,23 0,81 1,22 2,97 1,73 4,08 4,55 4,30 5,70 5,52 4,93 6,08 3, KOENIG és SIMIANER (2006) német holstein fríz populáció 1,7 6,
 8 helyi jelentőségű spanyol húsmarha fajta Alistana Asturiana de la Montana Asturiana de los Valles Avilena-Negra Iberica Bruna dels")
20 Szerző és fajta F (%) Pt GI (év) Ne MAIGNEL és mtsai (1996) Abondance Pie Rouge des Plaines (PRP) Brown Swiss Tarentaise Simmental Montbéliarde Normande Holstein ,83 2,82 4,04 3,78 3,47 4,19 5,02 4, Mc PARLAND és mtsai (2007) Charolais Limousin Hereford Angus Simmental Holstein-Fries 0,54 0,57 2,19 1,31 1,35 1, ,17 6,71 6,03 6,09 6,54 6, PÉREZ TORRECILLAS és mtsai (2002) Chianina Maremmana Mucca Pisana 3,61 6,76 7,06 2,88 3,13 2, ,97 110,92 19,91 SORENSEN és mtsai (2005) dán holstein dán jersey dán vörös 3,9 3,4 1,4 7,20 7,36 6,77 4,6; 5,0 4,7; 5,2 4,8; 5, GOYACHE és mtsai (2003) Xalda juh 1,54 1,09 2,97 24,8 HUBY és mtsai (2003) 6 francia húsjuh fajta 1-3 6,7; 2,3; 6,7, 4,7, 4,2, 4,1 4,1; 4,3; 3,4, 4,1; 3,9; 3,
 Xalda juh 1,54 1,09 2,97 24,8 HUBY és mtsai (2003) 6 francia húsjuh fajta 1-3 6,7; 2,3;")
21 3. táblázat. Sertés, nyúl és kutya fajták esetében közölt beltenyésztési együtthatók (F), pedigré teljességek (Pt), generációs intervallumok (GI) és effektív populációméretek (Ne) Szerző és fajta F (%) Pt GI (év) Ne BAUMUNG és mtsai (2002) osztrák nagyfehér osztrák lapály pietrain 1,43 1,13 1,34 7,85 6,20 5, JANSSENS és mtsai (2005) belga lapály pietrain 3,15 2,36 9 8,8 2,5 3, GLAZEWSKA (2008), lengyel kopó 7, ,32 - LEROY és mtsai 12,4; 3,9, 3,5; 6,3; 8,1; 4,7; 3,8; 4,4; 4,9; 20, 76, 53, 33, (2006), 9 francia 5,4; 7,2; 6,1; 6,0; 5,9; 3,3; 5,6; 3,2; 4,6; 147, 40, 88, kutyafajta 3,3; 6,0; 6,2; 8,2; 5,2 4,9 70, 82 4,1; 4,5; 4,0 GYOVAI (2006), Pannon fehér nyúl 5,56 10,91 1,32, 1,15, 1, A beltenyésztettség hatása multipara állatfajok reprodukciós és hízékonysági, valamint egyéb tulajdonságaira WHITE (1972) két kísérletben szisztematikusan növelte a kísérleti egerek beltenyésztettségét annak megállapítása céljából, hogy ez milyen hatással van a növekedési és az anyai tulajdonságokra. Első kísérletében a beltenyésztettséggel lineárisan növekvő leromlást tapasztalt az anyák teljesítményében. Megállapította, hogy azok a fiókák, melyeket beltenyésztett anyák szoptattak, kisebb súlyt értek el 12 és 21 napos 21
22 korukra, mint azok, melyeket nem beltenyésztett anyák neveltek. A második kísérletben a beltenyésztettségi szinttel szignifikánsan növekvő leromlást tapasztalt a születési, a 12, a 21, a 42 és az 56 napos korban mért súlyban. Vizsgálatában a 0-tól 79 %-os beltenyésztettségi szinteken (0; 12,5; 25; 37,5; 50; 73 %-os beltenyésztési együtthatójú anyák és 0; 22; 38; 50; 59; 79 %-os beltenyésztési együtthatójú ivadékok) tapasztalt összes leromlás a két ivar esetében hasonló volt, a nőivarnál 13,3 %, a hímivarnál pedig 11,8 % a 21 napos korban mért súlyban. HOLT és mtsai (2005/a) 122 generáción keresztül alomlétszámra szelektált egérállományt vizsgáltak. 3 vonalat alakítottak ki, egyet a nagy alomlétszámra, egyet a kis alomlétszámra szelektálva, a harmadik pedig kontroll vonal volt, majd ezeket keresztezve összesen 8 vonal teljesítményét nézték 3 kísérleti periódusban. A kiindulási vonalak beltenyésztési koefficiense 0-ról indult, majd generációról generációra növekedett 0,33-1,19 %-kal, a legmagasabb szinten beltenyésztett vonal átlagos beltenyésztési koefficiense 0,64-es szintet ért el a 122. generációra. Eredményeik szerint a vonalak összes periódusban létrejött átlagos beltenyésztési leromlása 0,39 fiókában nyilvánult meg. Megállapították, hogy a beltenyésztési együttható 10 %-os növekedése az átlagos alomlétszámot 0,72-vel csökkenti. HOLT és mtsai (2005/b) egy másik kísérletben 124 generáción keresztül egy alomlétszámra szelektált egérállományt, egy kontroll vonalat, valamint az ezek keresztezésével előállított vonal teljesítményét vizsgálta a 124. és 125. generációban. A populáció mérete minden vonalban 40 és 70 nőstény egérből állt, az egyedek beltenyésztettsége 0,3-1,2 %-kal nőtt generációnként. Az állatok átlagos beltenyésztettségi szintje a vizsgált generációkban a szelektált állományban 39 %-os, a kontroll állományban 65 %-os volt. Eredményeik szerint a beltenyésztési együttható 10 %-os 22
23 növekedése 0,42-vel csökkenti az átlagos alomlétszámot. BOWMAN és FALCONER (1960) kísérletében ez a szám 0,59, MCCARTHY (1967) kutatása alapján pedig 0,46. MOURA és mtsai (2000) Botucatu nyúlfajtában végzett vizsgálatuk alapján megállapították, hogy az anyák beltenyésztési együtthatójának 10 %-os növekedése az élve született fiókák számát 0,805-el csökkentette. A fiókák beltenyésztési együtthatójának 10%-os növekedése átlagosan további 0,589 fiókával csökkentette az alomszámot. Kaliforniai és új-zélandi fehér nyúlfajtákban FERRAZ és mtsai (1991) ezzel szemben azt tapasztalták, hogy az anyák beltenyésztési együtthatójának növekedésével a 21 napos alomszám és alomsúly ugyan csökkent, a kapott teljesítménycsökkenés azonban egyik értékmérő esetében sem volt szignifikáns. BOKOR és mtsai (2008) magyarországi angol telivér állományban vizsgálták a beltenyésztés okozta leromlást a lovak versenyteljesítményében ló 1856 versenyen teljesített futásának adatain vizsgálták a végső, éves és általános handicap súlyt, mint teljesítmény-tulajdonságot figyelembe véve. Az elemzését követően azt állapították meg, hogy a beltenyésztés mértékének nem volt statisztikailag igazolható hatása az említett tulajdonságra A beltenyésztettség hatása a hízékonysági és szaporasági tulajdonságokra sertésekben BERESKIN és mtsai (1968) többváltozós regresszió analízissel keresték a választ arra, hogy a beltenyésztettségi szint milyen hatással van a kocák és az utódaik teljesítményére alom vizsgálata alapján megállapították, hogy a kocák beltenyésztettségi szintje szignifikáns leromlást okoz az 23
24 alomnagyságban (-0,25 malac/10 % F növekedésnél), az alomsúlyban (- 0,34kg/10 % F növekedésnél) és az átlagos választási súlyban (-1,4kg/10 % F növekedésnél). ANKER (1970) tapasztalata szerint a sertésnél a rokontenyésztés következtében létrejövő leromlás elég hamar, 0,10-0,15 körüli beltenyésztési koefficiens elérésekor jelentkezik és elsősorban a felnevelési veszteségek alakulásában észrevehető. RADNAI (1980) 979 rokontenyésztett és 1109 nem rokontenyésztett KA-HYB kan adatai alapján megállapította, hogy a rokontenyésztett - 12,5-37 %-os beltenyésztési együtthatójú - és a nem rokontenyésztett kanok tenyésztésre való alkalmasság tekintetében egymástól nem tértek el szignifikáns mértékben a gyakorlatban. DICKERSON és mtsai (1954) eredményei szerint a beltenyésztettségi szint 10 %-os növekedése esetén 1,16 kg-mal romlik a 154 napos súly, TRIEBLER és mtsai (1980) a beltenyésztési együttható 10 %-os növekedése esetén a választás utáni súlygyarapodás 1,32 kg-os csökkenéséről számoltak be. A fialásonként élve született ivadékok számában ROTHSCHILD és BIDANEL (1998) több szerző eredményei alapján arról számoltak be, hogy a kocák beltenyésztési együtthatójának 10 %-os növekedése esetén azok átlagosan 0,40 malaccal kevesebbet fialtak. A malacok beltenyésztési együtthatójának 10 %-os növekedése átlagosan további 0,29 malaccal csökkentette a fialásonként élve született ivadékok számát. A kocák 10 %-os beltenyésztettségi szintjénél a malacszám 0,20-0,23 - os csökkenéséről és 3,35 kg-os 21 napos alomsúly csökkenésről számoltak be JOHNSON (1990), valamint CULBERTSON és mtsai (1998). BELIC és mtsai (2002) svéd lapály populációban vizsgálták a gyors beltenyésztés hatását a kocák szaporasági tulajdonságaira. A 72 koca 24
25 beltenyésztési együtthatója 3,12 % és 31,25 % közötti volt. A szerzők szignifikáns beltenyésztési leromlást állapítottak meg a született malacok számában (-0,103 malac/1 % F növekedésnél), az élve született malacok számában (-0,091 malac/1 % F növekedésnél), a választási alomlétszámban (-0,078 malac/1 % F növekedésnél) és a halva született malacok számában (-0,088 malac/1 % F növekedésnél). BRANDT és MÖLLERS (1999) göttingeni minisertés populációban becsültek beltenyésztési leromlást szaporasági tulajdonságokra és súlygyarapodásra. A vizsgált 282 koca 6777 malacának átlagos beltenyésztettségi szintje 10 % körüli volt és az adatok közel normál eloszlást mutattak. Az almonként élve-halva született malacok arányában nem találtak beltenyésztési leromlást, sem a kocák, sem pedig a malacok esetében. Az átlagos és az egyedi születési súly szignifikáns összefüggést mutatott a koca beltenyésztettségével, míg az alom beltenyésztettségével nem találtak szignifikáns összefüggést. Eredményeik szerint a kocák beltenyésztési együtthatójának 10 %-os növekedése 70 grammal csökkentette az egyedi születési súlyt. A 6 hetes korban mért súlyra mind a koca, mind az alom beltenyésztettsége szignifikáns hatást gyakorol. Ennél a populációnál a kocák és az alom beltenyésztési együtthatójának 10 %-os növekedése egyaránt 250 g-os leromlást okoz a 6 hónap alatt elért súlygyarapodás tekintetében. RODRIGÁNEZ és mtsai (1998), valamint FARKAS és mtsai (2007) hasonló tulajdonságokat vizsgáltak lapály és nagy fehér hússertéseken. A vizsgált populációkban nem történt szándékos rokontenyésztés, ezért a beltenyésztettség növekedésének ütemében nagy eltérés nem volt. RODRIGÁNEZ és mtsai (1998) nagyfehér hússertés populációban 21,9 %- os beltenyésztési együttható mellett 0,89 malac leromlást becsült almonként az alomlétszámra, míg 1,7 malac leromlást almonként az élve született 25
26 malacok számára. A FARKAS és mtsai (2007) által vizsgált magyar nagyfehér hússertés populációban 1,79 beltenyésztési együttható mellett 0,113 malac/alom leromlást becsült az élve született malacok számára. 2,689%-os beltenyésztettség mellett magyar lapály populációban pedig az általa tapasztalt leromlás 0,113 malac/alom volt ugyanerre a tulajdonságra nézve. KÖCK és mtsai (2009) osztrák lapály és osztrák nagy fehér hússertés populációban folytattak vizsgálatokat, melyek során a teljes beltenyésztési együtthatót két részre osztották, új és régi beltenyésztésre. Az új beltenyésztési együttható az első öt generáció beltenyésztési együtthatója, a régi pedig a teljes beltenyésztési együttható, mely a rendelkezésre álló 41 évre visszamenő pedigré és az új beltenyésztési együttható különbsége. A régi és az új beltenyésztési együtthatót figyelembe véve is csökkent a malacok életképessége mindkét fajtában. Az alomra vonatkozó, új beltenyésztési együttható 10 %-os növekedése esetén a becsült beltenyésztés okozta leromlás 0,18 és 0,23 választott malac volt a nagyfehér és a lapálysertés fajtában, sorrendben. A régi, alomra vonatkozó beltenyésztési együttható 10 %-os növekedése esetén 0,31 és 1,91 választott malac belteltenyésztési leromlást tapasztaltak a nagyfehér hússertés és lapálysertés fajtában, sorrendben. A lapálysertés populációban a régi, alomra vonatkozó beltenyésztés nagyobb leromlást eredményezett, mint az új beltenyésztés A hazai tenyészértékbecslési rendszer és annak beltenyésztési vonatkozásai Hazánkban a tenyészértékbecslésben alkalmazott módszer 2008 óta a HENDERSON (1975) által kifejlesztett Best Linear Unbiased Prediction, azaz Legjobb Lineáris Torzítatlan Előrejelzés. Ezt megelőzően 26
27 Magyarországon a tenyészállatok értékelése, rangsorolása és kiválasztása a Hazel-féle szelekciós index módszer szerint történt (HAZEL, 1943). Ennek alkalmazása akkor célszerű, ha az egyedek illetve egyedcsoportok véletlen mintavételei ugyanazon populációból származnak, a környezeti hatások egy részétől eltekinthetünk (FARKAS, 2008). A Hazel-index meghatározásához a mérési eredményeket a súlyra és az ivarra korrigálták a megadott képletek alapján (OMMI, 2004), majd a vizsgálati egyedeknek több tulajdonságban mutatott teljesítményét egyetlen számmá vonták össze. Ez a módszer azonban nem vesz figyelembe olyan környezeti hatásokat évszak, tenyészet, év-hó, állomás melyekre szükséges lenne korrigálni (SZŐKE és KOMLÓSI, 2000). Ezeket a környezeti hatásokat képes korrigálni a BLUP eljárás, melynek használata akkor célszerű, ha az egyedek eloszlása tenyészértékük alapján a tenyészetek között véletlenszerű, tehát a legjobb és legrosszabb tenyészértékű egyedek származása az ún. jó és rossz körülményeket biztosító tenyészetekben egyaránt megtalálható. A BLUP módszert a gyakorlatban először az amerikai szarvasmarhatenyésztésben alkalmazták (VAN VLECK, 1977), Magyarországon szintén ebben az ágazatban került először bevezetésre, melynek körülményeiről ZSILINSZKY (1984, 1987) számolt be. E közleményeiben megállapította, hogy a korszerű matematikai és statisztikai ismereteken nyugvó BLUPalgoritmus segítségével a tenyészbikák közötti örökítőérték-különbségek nagy pontossággal állapíthatók meg, ezért az egykorú istállótársak összehasonlítására épülő korábbi tenyészérték-becslési eljárás átalakítása egyértelműen sikeres volt. Hazánkban a bikák tenyészérték-becslésének hivatalos módszere 1985 óta a BLUP, 1999-től pedig egyedmodellt alkalmaznak. 27
28 A módszer sertéstenyésztésben való alkalmazhatóságának nehézségeire BRUNS (1983) hívta fel a figyelmet, miszerint a tenyészkanok közötti torzítatlan genetikai összehasonlítás érdekében a környezeti tényezők (pl. állomás-, év-, évszakhatás, stb.) minőségében fennálló különbségeket, az ivadékok ivari hovatartozásából és genotípusából adódó egyéb hatásokat minden egyes tenyészkannak az összes többivel való szimultán összevetése révén vegyék figyelembe, valamint figyelmeztetett a buktatókra is, pl. kanok lokális használata, mesterséges termékenyítés korlátozott volta, stb. Hasonló álláspontot képviselt PRIBYL és VÁCHAL (1985), valamint THOMPSON és MEYER (1986) is. Azonban az 1980-as évek közepétől egyre több olyan közlemény látott napvilágot, melyek modellszámításokkal támasztották alá a BLUP módszer gyakorlati tenyésztőmunkában várható előnyeit (LUNDHEIM és ERIKSSON, 1984; HUDSON és KENNEDY, 1985; GROMAN és FLANAGAN, 1987; MABRY és mtsai, 1987; BELONSKY és KENNEDY, 1987), majd a 90-es évek elejétől az eljárást a nyugat-európai és a környező országokban is bevezették és ezzel jelentős szelekciós haladást értek el (KOMLÓSI, 1999). A témáról részletes áttekintést adnak THOLEN és GROENEVELD (1989), GROENEVELD és mtsai (1990), GROENEVELD és mtsai (1991), KOVAC és GROENEVELD (1990), GROENEVELD és KOVAC (1992) WOLF és mtsai (1998), GROENEVELD és PESKOVICOVA (1999), PESKOVICOVA és mtsai (2002) munkái. A módszer hivatalossá tétele Magyarországon csak január 1.-ével történt meg. Bevezetését sok éves kutatómunka előzte meg, a modellek kidolgozása már 1992-ben megkezdődött egy magyar-német együttműködés keretében Groeneveld professzor vezetésével, melynek eredményeiről számos publikáció jelent meg (GROENEVELD és mtsai, 1992; 1996; CSATÓ és RADNÓCZI, 1994; CSATÓ és mtsai, 2002; NAGY és mtsai, 2004). 28
29 Bevezetésének a feltételei már régóta biztosítottak voltak, úgy mint az üzemi sajátteljesítmény vizsgálati-, a központi vizsgálóállomási- és a reprodukciós adatok központi adatgyűjtése, valamint a megfelelő számítógépes kapacitás. Az OMMI is lépéseket tett a BLUP módszer alkalmazására, a saját fejlesztésű programcsomag tenyészetenkénti értékelést tett lehetővé apa modellel az üzemi saját- és reprodukciós teljesítmény területén. Az ígéretes eredmények után az összetett BLUPmodellen alapuló index több éven keresztül a szelekciós index mellett informatív, kísérleti jelleggel már feltüntetésre került (FARKAS, 2008). Jelenleg a szaporaság becslésére a született élőmalacok számának figyelembevételével egyváltozós modellt, míg az üzemi sajátteljesítmény vizsgálatra (ÜSTV), a központi hízékonyság és vágóérték vizsgálatra (HVT) és az üzemi ivadékvizsgálatra (ÜITV) a kijelölt paraméterek tekintetében többváltozós modellt használ a havonta esedékes tenyészérték-becslésre az MGSZH. A tenyészértékekből egy ún. TT indexet (teljes tenyészérték index) alakítanak ki a következő értékmérők tenyészértékének szerepeltetésével: hízlalási nap (HVT), értékes húsrészek aránya (HVT), életnap (ÜSTV, ÜITV) és színhús százalék (ÜSTV, ÜITV). BELONSKY és KENNEDY (1988) rámutat arra, hogy ha egy populációt a BLUP-tenyészértékek alapján szelektáltak, akkor az a beltenyésztési ráta növekedését okozza. MEHRABANI-YEGANEH és mtsai (2000) viszont azt közölték, hogy a beltenyésztési együttható szerepeltetése a BLUP modellben nem változtatja meg az egyedek tenyészértékének rangsorát. 29
30 3. A DISSZERTÁCIÓ CÉLKITŰZÉSEI Munkám során a következő célokat tűztem ki: A rendelkezésre álló származási adatokból a magyar nagyfehér (MNF) és a magyar lapálysertés (ML) fajtában a pedigréanalízis elvégzése, a pedigré minőségének (teljességének) meghatározása, ezáltal az állomány beltenyésztettségének értékelése. Továbbá a beltenyésztési leromlás vizsgálata a MNF és ML fajtában az üzemi sajátteljesítmény vizsgálati adatok (egy életnapra jutó napi testsúly-gyarapodás, színhús százalék, karajátmérő, szalonna 1 vastagság, szalonna 2 vastagság), valamint szaporasági- és felnevelési teljesítményvizsgálat (élve született malacok száma és a holtan született malacok száma) alapján. Ezeken túlmenően a legpontosabb modellváltozatok megtalálása, beleértve annak vizsgálatát is, hogy hány generációt érdemes figyelembe venni, mellyel pontosabbá és reálisabbá tehető a magyar sertésállományokra vonatkozó tenyészérték-becslés. 30
31 4. ANYAG ÉS MÓDSZER 4.1. Adatbázisok ÜSTV Adatok Magyar nagyfehér hússertés A vizsgálat során az OMMI (az MGSZH jogelődje) által gyűjtött Üzemi sajátteljesítmény vizsgálati (ÜSTV) adatokat elemeztem, melynek keretében között vizsgált sertés eredményeit értékeltem. A vizsgált egyedek magyar nagyfehér hússertések voltak (MNF), melyeket 67 tenyészetben tartottak. Az adatokhoz tartozó pedigrében tól ig született egyedek szerepeltek. A teljesítményvizsgálatot megelőzően az egyedeket ivar szerint elkülönítve és falkásítva kell elhelyezni úgy, hogy egy egyedre 60 kg-os egyedi súlyig legalább 0,6m 2, 60 kg felett legalább 1,0 m² terület jusson. A vizsgálati időszakban a takarmányozást egységesen, ad libitum etetéssel kell végezni hosszabb távon azonos minőségű, amely lehetővé teszi a tenyészállatok genetikai értékének realizálását, a minőségi tenyészállatnevelést. (MGSZH, 2009). A kan és kocasüldők értékmérő tulajdonságait Üzemi sajátteljesítmény vizsgálat (ÜSTV) keretében mérték élő egyedeken kg közötti testsúlyban. A vizsgált ÜSTV - értékmérőket és azok statisztikai jellemzőit a 4. táblázatban mutatom be. 31
32 4. táblázat. A vizsgált ÜSTV-értékmérők és azok statisztikai jellemzői MNF fajtában Elemszám Vizsgálati Átlag Szórás Min. Max. idő Szinhus (%) ,02 1,98 45,54 69,87 NapiGyar ,91 61,67 270,0 924,53 (g/nap) Karaj (mm) ,26 6,29 22,0 80,0 SZ1 (mm) ,3 3,44 2,0 36,0 SZ2 (mm) ,45 2,76 2,00 31,00 A színhús százalékot az MGSZH által engedélyezett, hitelesített ultrahangos készülékkel mérik. Az egy életnapra jutó testsúly-gyarapodást a vizsgálat végén mért testsúly, illetve életkor hányadosa alapján számítják. A karajátmérőt hátulról a 3. és a 4. borda között a gerincvonaltól 6 cm- re oldalirányban, a szalonna 1 vastagságot a sertés bal oldalán a 3. és a 4. ágyékcsigolya között a gerincvonaltól 8 cm-re oldalirányban, a szalonna 2 vastagságot hátulról a 3. és a 4. borda között a gerincvonaltól 6 cm- re oldalirányban vizsgálják Magyar lapálysertés A vizsgálatot magyar lapálysertés között - az MGSZH által - gyűjtött ÜSTV adatain végeztem, melyeket 97 tenyészetben tartottak. A származást rögzítő pedigrében az től ig született egyedek szerepeltek. A vizsgálatra vonatkozó előírások megegyeznek a magyar nagyfehér fajtánál ismertetekkel, azokat a Sertés Teljesítményvizsgálati Kódexben (MGSZH, 2009) rögzítetteknek 32
33 megfelelően kell végezni. A vizsgált ÜSTV - értékmérőket és azok statisztikai jellemzőit az 5. táblázatban mutatom be. 5. táblázat. A vizsgált ÜSTV-értékmérők és azok statisztikai jellemzői ML fajtában Elemszám Vizsgálati Átlag Szórás Min. Max. idő Szinhus (%) ,61 2,14 44,59 69,99 NapiGyar ,38 62,17 295,0 902,0 (g/nap) Karaj (mm) ,74 6,05 20,0 80,0 SZ1 (mm) ,90 3,23 2,0 32,0 SZ2 (mm) ,22 2,78 2,0 31, SZFTV adatok Magyar nagyfehér hússertés A dolgozathoz felhasznált Szaporasági és felnevelési vizsgálati adatokat szintén az MGSZH bocsájtotta rendelkezésemre, 1979 és 2009 között gyűjtött magyar nagyfehér koca fialását elemeztem, melyek 193 tenyészetből származtak. A pedigrében az egyedek től ig szerepeltek. A kocák elhelyezésére, takarmányozására és az adatgyűjtés körülményeire szintén a Sertés Teljesítményvizsgálati Kódex (MGSZH, 2009) alapján került sor. Ennek értelmében a vizsgálatba vont telepen az állatok tartása és takarmányozása a fajta igényeinek megfelelően kell, hogy történjen a genetikai érték realizálása érdekében. A vizsgálatba vont értékmérők az élve született malacok száma (Észmsz) és a holtan született malacok száma (Hszmsz) volt. 33
34 tüntetem fel. A tulajdonságok alapstatisztikáit a 6. táblázatban fialásonként 6. táblázat. A vizsgált SZFTV-értékmérők és azok statisztikai jellemzői, fialásonként (MNF) Tulajdonság Elemszám Átlag Szórás Élve született malacok száma (Észmsz), összes ,95 2,70 Észmsz - 1. fialás ,23 2,59 Észmsz - 2. fialás ,97 2,66 Észmsz - 3. fialás ,45 2,67 Észmsz - 4. fialás ,55 2,69 Észmsz - 5. fialás ,47 2,69 Észmsz - 6. fialás ,33 2,69 Észmsz - 7. fialás ,11 2,70 Észmsz - 8. fialás ,94 2,68 Észmsz - 9. fialás ,70 2,70 Észmsz fialás ,51 2,69 Észmsz fialás ,27 2,61 Észmsz fialás ,05 2,59 Észmsz fialás 998 8,83 2,55 Észmsz fialás 560 8,73 2,44 Észmsz fialás 275 8,26 2,66 Holtan született malacok száma (Hszmsz), összes ,46 1,10 Hszmsz - 1. fialás ,44 1,07 Hszmsz - 2. fialás ,38 0,97 Hszmsz - 3. fialás ,42 1,03 Hszmsz - 4. fialás ,46 1,10 Hszmsz - 5. fialás ,50 1,16 Hszmsz - 6. fialás ,54 1,20 Hszmsz - 7. fialás ,58 1,27 Hszmsz - 8. fialás ,6 1,29 Hszmsz - 9. fialás ,63 1,30 Hszmsz fialás ,63 1,30 Hszmsz fialás ,64 1,32 Hszmsz fialás ,60 1,29 34
35 Hszmsz fialás 998 0,49 1,10 Hszmsz fialás 560 0,53 1,18 Hszmsz fialás 275 0,45 1, Magyar lapálysertés A dolgozathoz felhasznált Szaporasági és felnevelési vizsgálati adatokat szintén az MGSZH bocsájtotta rendelkezésemre, 1985 és 2005 között gyűjtött magyar lapály koca fialását elemeztem, melyek 57 tenyészetből származtak. A pedigrében az től ig született egyedek szerepeltek. A vizsgálat során csak a legalább 4 fialással rendelkező kocák adatait vettem figyelembe. A kocák elhelyezésére, takarmányozására és az adatgyűjtés körülményeire szintén a Sertés Teljesítményvizsgálati Kódex (MGSZH, 2009) alapján került sor. Ennek értelmében a vizsgálatba vont telepen az állatok tartása és takarmányozása a fajta igényeinek megfelelően kell, hogy történjen a genetikai érték realizálása érdekében. A vizsgálatba vont értékmérők az élve született malacok száma (Észmsz) és a holtan született malacok száma (Hszmsz) volt. A tulajdonságok alapstatisztikáit a 7. táblázatban fialásonként tüntetem fel. 35
36 7. táblázat. A vizsgált SZFTV-értékmérők és azok statisztikai jellemzői, fialásonként (ML) Tulajdonság Elemszám Átlag Szórás Élve született malacok száma (Észmsz), összes ,98 2,26 Észmsz - 1. fialás ,65 1,79 Észmsz - 2. fialás ,94 2,16 Észmsz - 3. fialás ,31 2,21 Észmsz - 4. fialás ,28 2,36 Észmsz - 5. fialás ,21 2,36 Észmsz - 6. fialás ,06 2,39 Észmsz - 7. fialás ,82 2,39 Észmsz - 8. fialás ,55 2,42 Észmsz - 9. fialás ,35 2,39 Észmsz fialás ,21 2,34 Észmsz fialás 848 8,90 2,37 Észmsz fialás 436 8,78 2,55 Észmsz fialás 209 8,36 2,26 Észmsz fialás 87 8,06 2,48 Észmsz fialás 32 8,46 2,73 Holtan született malacok száma (Hszmsz), összes ,46 1,04 Hszmsz - 1. fialás ,34 0,81 Hszmsz - 2. fialás ,34 0,83 Hszmsz - 3. fialás ,38 0,89 Hszmsz - 4. fialás ,45 1,02 Hszmsz - 5. fialás ,52 1,11 Hszmsz - 6. fialás ,57 1,20 Hszmsz - 7. fialás ,62 1,30 Hszmsz - 8. fialás ,71 1,43 Hszmsz - 9. fialás ,71 1,38 Hszmsz fialás ,79 1,43 Hszmsz fialás 848 0,82 1,51 Hszmsz fialás 436 0,77 1,26 Hszmsz fialás 209 0,73 1,29 Hszmsz fialás 87 0,91 1,55 Hszmsz fialás 32 0,71 1,65 36
37 4.2 Pedigréanalízis, beltenyésztési együtthatók Az adatok elemzését mind az ÜSTV, mind pedig az SZFTVtulajdonságok esetében pedigréanalízissel kezdtem, melyhez a PEDUTIL, az NGEN és a VANRAD programokat használtam. Első lépésben a PEDUTIL program futtatásával kiválasztottam a pedigréből a mérési eredménnyel rendelkező egyedek származási adatait. Ezt követően BOICHARD és mtsai (1997) által kidolgozott képlet alapján megállapítottam az egyedekre vonatkozó pedigré-teljességet az NGEN programmal, azaz hogy bármely egyed származása hány teljes generációra nézve ismert (teljes generáció ekvivalens), az alábbi képlet alapján: 1/N ( j=1 N i=1 Mj 1/2 gij ) ahol N a vizsgált populációban található egyedek száma, M j a j-ik egyednek a pedigrében szereplő valamennyi ősének száma, g ij a generációk száma a j- ik egyed és annak i-ik őse között. A pedigré teljességet az ÜSTV-értékmérőknél három, négy, öt, hat, illetve hét (pt3, pt4, pt5, pt 6, pt7), míg az SZFTV vizsgálatok adatain három, négy, nyolc, kilenc és tíz (pt3, pt4, pt8, pt9, pt10) generációra visszamenően vizsgáltam. Ennek oka, hogy az SZFTV adatbázisokhoz hosszabb és teljesebb pedigré állt rendelkezésre, mint az ÜSTV során vizsgált egyedekhez, így a fialási adatokkal rendelkező tenyészkocák esetében indokolt volt a származást több generációra visszemenően nézni. 37
38 Ezután minden egyedre kiszámítottam a beltenyésztési együtthatót (F) (WRIGHT, 1922) a VANRAD program segítségével. Adott (X) egyed beltenyésztési koefficiense: F X = (1/2) n+n +1 (1+F A ) ahol A az X egyed apai és anyai származási láncában levő közös ős, n és n az X egyed és A ős közötti generációk száma az apai (n), illetve az anyai (n ) ágon, F A a közös ős beltenyésztési koefficiense. A jel az X egyed apai és anyai származási láncában levő összes közös ősre vonatkozó összegzést jelent. A beltenyésztési együtthatót szintén három, négy, öt, hat és hét generációra visszamenően néztem (f3, f4, f5, f6, f7) az ÜSTV adatbázisban szereplő egyedek, míg három, négy, nyolc, kilenc és tíz generációra visszamenően (f3, f4 f8, f9, f10) az SZFTV adatbázisban szereplő egyedek származása esetében. Mindhárom program a PEDIG szoftvercsomag része (BOICHARD, 2002). Ezen számítások elvégzését követően a pedigréanalízisből származó eredményeket a mérési adatokat tartalmazó adatbázishoz illesztettem A leíró statisztikai számításokhoz és az adatok ábrázolásához a SAS szoftvert használtam (SAS Institute Inc., 2004) BLUP-modellek Az adatbázisok előkészítését követően REML és BLUP módszerekkel genetikai paraméterbecslést és tenyészértékbecslést végeztem a beltenyésztési együtthatókat és a pedigrételjességet is tartalmazó 38
-Vese körüli zsír tömege (g) (12) ,68 2,18
 (12) ,68 2,18") KUTATÁSI JELENTÉS a Kvantitatív genetikai vizsgálatok a Pannon fehér nyúlpopulációban c. ifjúsági OTKA téma (2005-2007) eredményeiről. A kutasási munka célkitűzése, hogy a Kaposvári Egyetem Pannon fehér
KUTATÁSI JELENTÉS a Kvantitatív genetikai vizsgálatok a Pannon fehér nyúlpopulációban c. ifjúsági OTKA téma (2005-2007) eredményeiről. A kutasási munka célkitűzése, hogy a Kaposvári Egyetem Pannon fehér
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI
 ÉRTEKEZÉS TÉZISEI") DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője DR. HORN PÉTER az MTA rendes tagja Témavezető DR. NAGY ISTVÁN Tudományos
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője DR. HORN PÉTER az MTA rendes tagja Témavezető DR. NAGY ISTVÁN Tudományos
ÁLLATTENYÉSZTÉSI GENETIKA
 TÁMOP-4.1.2-08/1/A-2009-0010 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by
TÁMOP-4.1.2-08/1/A-2009-0010 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by
Általános állattenyésztés
 Általános állattenyésztés 10. Előadás Tenyésztési (párosítási) eljárások 1. Előadás-vázlat Fajtatiszta tenyésztés Kombinációs párosítás Vérfrissítés Rokontenyésztés, beltenyésztés Vérvonaltenyésztés Szintetikus
Általános állattenyésztés 10. Előadás Tenyésztési (párosítási) eljárások 1. Előadás-vázlat Fajtatiszta tenyésztés Kombinációs párosítás Vérfrissítés Rokontenyésztés, beltenyésztés Vérvonaltenyésztés Szintetikus
DOKTORI (PhD) ÉRTEKEZÉS NAGYNÉ KISZLINGER HENRIETTA KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR
 ÉRTEKEZÉS NAGYNÉ KISZLINGER HENRIETTA KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR") DOKTORI (PhD) ÉRTEKEZÉS NAGYNÉ KISZLINGER HENRIETTA KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR 2014 KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR Állatgenetikai és Biotechnológiai Tanszék
DOKTORI (PhD) ÉRTEKEZÉS NAGYNÉ KISZLINGER HENRIETTA KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR 2014 KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR Állatgenetikai és Biotechnológiai Tanszék
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM
 ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM") DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR Állatgenetikai és Biotechnológiai Tanszék A doktori iskola vezetője KOVÁCS MELINDA az MTA levelező tagja Témavezető NAGY
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM AGRÁR- ÉS KÖRNYEZETTUDOMÁNYI KAR Állatgenetikai és Biotechnológiai Tanszék A doktori iskola vezetője KOVÁCS MELINDA az MTA levelező tagja Témavezető NAGY
DOKTORI (PhD) ÉRTEKEZÉS GYOVAI PETRA KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR
 ÉRTEKEZÉS GYOVAI PETRA KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR") DOKTORI (PhD) ÉRTEKEZÉS GYOVAI PETRA KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR 2011 KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Mezıgazdasági Termékfeldolgozás és Minısítés Tanszék A doktori iskola vezetıje DR. HORN
DOKTORI (PhD) ÉRTEKEZÉS GYOVAI PETRA KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR 2011 KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Mezıgazdasági Termékfeldolgozás és Minısítés Tanszék A doktori iskola vezetıje DR. HORN
A VÁGÁSI KOR, A VÁGÁSI SÚLY ÉS A ROSTÉLYOS KERESZTMETSZET ALAKULÁSA FEHÉR KÉK BELGA ÉS CHAROLAIS KERESZTEZETT HÍZÓBIKÁK ESETÉBEN
 A vágási kor, a vágási súly és a rostélyos keresztmetszet alakulása fehér kék belga és charolais keresztezett hízóbikák esetében 1 () A VÁGÁSI KOR, A VÁGÁSI SÚLY ÉS A ROSTÉLYOS KERESZTMETSZET ALAKULÁSA
A vágási kor, a vágási súly és a rostélyos keresztmetszet alakulása fehér kék belga és charolais keresztezett hízóbikák esetében 1 () A VÁGÁSI KOR, A VÁGÁSI SÚLY ÉS A ROSTÉLYOS KERESZTMETSZET ALAKULÁSA
KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN
 MTA DOKTORI ÉRTEKEZÉS TÉZISEI KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN DR. NAGY ISTVÁN Ph.D., mezőgazdaságtudomány KAPOSVÁR 2016 1 1. BEVEZETÉS A kvantitatív genetika módszereit az állattenyésztők
MTA DOKTORI ÉRTEKEZÉS TÉZISEI KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN DR. NAGY ISTVÁN Ph.D., mezőgazdaságtudomány KAPOSVÁR 2016 1 1. BEVEZETÉS A kvantitatív genetika módszereit az állattenyésztők
A SZŐKE MANGALICA FAJTÁK GENETIKAI SZERKEZETÉNEK ÉRTÉKELÉSE
 A SZŐKE MANGALICA FAJTÁK GENETIKAI SZERKEZETÉNEK ÉRTÉKELÉSE Készítette: Dr. Posta János Debrecen Tartalomjegyzék. Bevezetés.... Szakirodalmi áttekintés.... A mangalica sertés.... Pedigréanalízis a sertéstenyésztésben....
A SZŐKE MANGALICA FAJTÁK GENETIKAI SZERKEZETÉNEK ÉRTÉKELÉSE Készítette: Dr. Posta János Debrecen Tartalomjegyzék. Bevezetés.... Szakirodalmi áttekintés.... A mangalica sertés.... Pedigréanalízis a sertéstenyésztésben....
Sodródás Evolúció neutrális elmélete
 Sodródás Evolúció neutrális elmélete Egy kísérlet Drosophila Drosophila pseudoobscura 8 hím + 8 nőstény/tenyészet 107 darab tenyészet Minden tenyészet csak heterozigóta egyedekkel indul a neutrális szemszín
Sodródás Evolúció neutrális elmélete Egy kísérlet Drosophila Drosophila pseudoobscura 8 hím + 8 nőstény/tenyészet 107 darab tenyészet Minden tenyészet csak heterozigóta egyedekkel indul a neutrális szemszín
A Hardy-Weinberg egyensúly. 2. gyakorlat
 A Hardy-Weinberg egyensúly 2. gyakorlat A Hardy-Weinberg egyensúly feltételei: nincs szelekció nincs migráció nagy populációméret (nincs sodródás) nincs mutáció pánmixis van allélgyakoriság azonos hímekben
A Hardy-Weinberg egyensúly 2. gyakorlat A Hardy-Weinberg egyensúly feltételei: nincs szelekció nincs migráció nagy populációméret (nincs sodródás) nincs mutáció pánmixis van allélgyakoriság azonos hímekben
MAGYAR JUHTENYÉSZTŐK ÉS KECSKETENYÉSZTŐK SZÖVETSÉGE
 MAGYAR JUHTENYÉSZTŐK ÉS KECSKETENYÉSZTŐK SZÖVETSÉGE Cím: 34 Budapest, Lőportár u. 6., Levélcím: 242 Bp. Pf. 365 Tel.: (06) 42-5030, Fax: (06) 42-503, E-mail: iroda@mjksz.hu www.mjksz.hu Beltenyésztettség
MAGYAR JUHTENYÉSZTŐK ÉS KECSKETENYÉSZTŐK SZÖVETSÉGE Cím: 34 Budapest, Lőportár u. 6., Levélcím: 242 Bp. Pf. 365 Tel.: (06) 42-5030, Fax: (06) 42-503, E-mail: iroda@mjksz.hu www.mjksz.hu Beltenyésztettség
DOKTORI (PhD) ÉRTEKEZÉS FARKAS JÁNOS KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR
 ÉRTEKEZÉS FARKAS JÁNOS KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR") DOKTORI (PhD) ÉRTEKEZÉS FARKAS JÁNOS KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR 2008 KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője: HORN PÉTER MTA rendes
DOKTORI (PhD) ÉRTEKEZÉS FARKAS JÁNOS KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR 2008 KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője: HORN PÉTER MTA rendes
Érdemes vakcinázni a sertések 1-es típusú parvovírusa ellen
 1. oldal (összes: 6) 2. oldal (összes: 6) A PPV1 által okozott szaporodásbiológiai zavarok megelőzésére a tenyészkocák vakcinázását széles körben alkalmazzák. Fogékony sertésekben a PPV1 fertőződést követően
1. oldal (összes: 6) 2. oldal (összes: 6) A PPV1 által okozott szaporodásbiológiai zavarok megelőzésére a tenyészkocák vakcinázását széles körben alkalmazzák. Fogékony sertésekben a PPV1 fertőződést követően
A versenyteljesítmény alapú szelekció lehetőségei a telivér- és ügetőtenyésztésben
 II. Anker Alfonz emléknap és nemzetközi galambásztalálkozó Kaposvári Egyetem 2012. november 17. A versenyteljesítmény alapú szelekció lehetőségei a telivér- és ügetőtenyésztésben Dr. Bokor Árpád Kaposvári
II. Anker Alfonz emléknap és nemzetközi galambásztalálkozó Kaposvári Egyetem 2012. november 17. A versenyteljesítmény alapú szelekció lehetőségei a telivér- és ügetőtenyésztésben Dr. Bokor Árpád Kaposvári
A nyulak is szenvednek a melegtől - és romlanak a szaporasági mutatók
 A nyulak is szenvednek a melegtől - és romlanak a szaporasági mutatók Ugyanakkor nem lehet tehetetlenül figyelni az emberi tevékenység által bekövetkező kedvezőtlen változásokat. A hőstressz negatív hatásai
A nyulak is szenvednek a melegtől - és romlanak a szaporasági mutatók Ugyanakkor nem lehet tehetetlenül figyelni az emberi tevékenység által bekövetkező kedvezőtlen változásokat. A hőstressz negatív hatásai
Magyar nagyfehér hússertés kanok. Tisztelt Sertéstenyésztő, Sertéstartó! Használja ki tapasztalatunkat és a garantált minőséget!
 TENYÉSZKANOK 2009 Tisztelt Sertéstenyésztő, Sertéstartó! Ön az Országos Mesterséges Termékenyítő Zrt. kankatalógusát tartja kezében. Társaságunk nevéhez fűződik a szarvasmarha és a sertés mesterséges termékenyítés
TENYÉSZKANOK 2009 Tisztelt Sertéstenyésztő, Sertéstartó! Ön az Országos Mesterséges Termékenyítő Zrt. kankatalógusát tartja kezében. Társaságunk nevéhez fűződik a szarvasmarha és a sertés mesterséges termékenyítés
LIMOUSIN TENYÉSZÜSZŐK VÁLASZTÁSI MUTATÓINAK VIZSGÁLATA
 Limousin tenyészüszők választási mutatóinak vizsgálata 1 () LIMOUSIN TENYÉSZÜSZŐK VÁLASZTÁSI MUTATÓINAK VIZSGÁLATA KOVÁCS A. 1 -BÁDER E. 1 - MIHÁLYFI I. 2 -GYÖRKÖS I. 3 1 Nyugat-Magyarországi Egyetem Mezőgazdaság-
Limousin tenyészüszők választási mutatóinak vizsgálata 1 () LIMOUSIN TENYÉSZÜSZŐK VÁLASZTÁSI MUTATÓINAK VIZSGÁLATA KOVÁCS A. 1 -BÁDER E. 1 - MIHÁLYFI I. 2 -GYÖRKÖS I. 3 1 Nyugat-Magyarországi Egyetem Mezőgazdaság-
A Kaposvári Egyetem nyúltenyésztési programja PANNON NYÚLTENYÉSZTÉSI PROGRAM. Anyai vonal. Pannon fehér. Nagytestű vonal
 A HÁROM GENOTÍPUS FŐBB JELLEMZŐI: A Kaposvári Egyetem nyúltenyésztési programja PANNON NYÚLTENYÉSZTÉSI PROGRAM A Kaposvári Egyetem három állomány szelekcióját végzi Anyai vonal Pannon fehér Nagytestű vonal
A HÁROM GENOTÍPUS FŐBB JELLEMZŐI: A Kaposvári Egyetem nyúltenyésztési programja PANNON NYÚLTENYÉSZTÉSI PROGRAM A Kaposvári Egyetem három állomány szelekcióját végzi Anyai vonal Pannon fehér Nagytestű vonal
Populációgenetikai. alapok
 Populációgenetikai alapok Populáció = egyedek egy adott csoportja Az egyedek eltérnek egymástól morfológiailag, de viselkedésüket tekintve is = genetikai különbségek Fenotípus = külső jellegek morfológia,
Populációgenetikai alapok Populáció = egyedek egy adott csoportja Az egyedek eltérnek egymástól morfológiailag, de viselkedésüket tekintve is = genetikai különbségek Fenotípus = külső jellegek morfológia,
HOLSTEIN-FRÍZ KERESZTEZETT TEHÉNÁLLOMÁNYOK KÜLLEMI TULAJDONSÁGAINAK ALAKULÁSA
 Holstein-fríz keresztezett tehénállományok küllemi tulajdonságainak alakulása 1(6) HOLSTEIN-FRÍZ KERESZTEZETT TEHÉNÁLLOMÁNYOK KÜLLEMI TULAJDONSÁGAINAK ALAKULÁSA BÁDER P. 1 - BÁDER E. 1 BARTYIK J 2.- PORVAY
Holstein-fríz keresztezett tehénállományok küllemi tulajdonságainak alakulása 1(6) HOLSTEIN-FRÍZ KERESZTEZETT TEHÉNÁLLOMÁNYOK KÜLLEMI TULAJDONSÁGAINAK ALAKULÁSA BÁDER P. 1 - BÁDER E. 1 BARTYIK J 2.- PORVAY
A SERTÉSTENYÉSZTÉS ÉVI EREDMÉNYEI
 A SERTÉSTENYÉSZTÉS 2013. ÉVI EREDMÉNYEI 1 Készítette: Novozánszky Gábor Lektorálta: Németh Csaba állattenyésztési igazgató Kiadja: Nemzeti Élelmiszerlánc-biztonsági Hivatal 2 Bevezetés A Nemzeti Élelmiszerlánc-biztonsági
A SERTÉSTENYÉSZTÉS 2013. ÉVI EREDMÉNYEI 1 Készítette: Novozánszky Gábor Lektorálta: Németh Csaba állattenyésztési igazgató Kiadja: Nemzeti Élelmiszerlánc-biztonsági Hivatal 2 Bevezetés A Nemzeti Élelmiszerlánc-biztonsági
Az evolúció folyamatos változások olyan sorozata, melynek során bizonyos populációk öröklődő jellegei nemzedékről nemzedékre változnak.
 Evolúció Az evolúció folyamatos változások olyan sorozata, melynek során bizonyos populációk öröklődő jellegei nemzedékről nemzedékre változnak. Latin eredetű szó, jelentése: kibontakozás Időben egymást
Evolúció Az evolúció folyamatos változások olyan sorozata, melynek során bizonyos populációk öröklődő jellegei nemzedékről nemzedékre változnak. Latin eredetű szó, jelentése: kibontakozás Időben egymást
Tenyésztési eljárások a szarvasmarha-tenyésztésben
 Tenyésztési eljárások a szarvasmarha-tenyésztésben Tenyésztési cél megválasztása Tenyésztési cél fogalma Tulajdonságok kiválasztása Tenyésztési eljárások Additív génhatásokat kihasználó Nem additív génhatásokat
Tenyésztési eljárások a szarvasmarha-tenyésztésben Tenyésztési cél megválasztása Tenyésztési cél fogalma Tulajdonságok kiválasztása Tenyésztési eljárások Additív génhatásokat kihasználó Nem additív génhatásokat
ÁLLATTENYÉSZTÉSI GENETIKA
 TÁMOP-4.1.2-08/1/A-2009-0010 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by
TÁMOP-4.1.2-08/1/A-2009-0010 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by
ELTÉRŐ TARTÁSTECHNOLÓGIÁK HATÁSA A TEJELŐ TEHÉNÁLLOMÁNYOK ÉLETTARTAMÁRA. Báder Ernő - Kertész Tamás Kertészné, Győrffy Eszter- Kovács Anita
 Eltérő tartástechnológiák hatása a tejelő tehénállományok élettartamára 1 (6) ELTÉRŐ TARTÁSTECHNOLÓGIÁK HATÁSA A TEJELŐ TEHÉNÁLLOMÁNYOK ÉLETTARTAMÁRA Báder Ernő - Kertész Tamás Kertészné, Győrffy Eszter-
Eltérő tartástechnológiák hatása a tejelő tehénállományok élettartamára 1 (6) ELTÉRŐ TARTÁSTECHNOLÓGIÁK HATÁSA A TEJELŐ TEHÉNÁLLOMÁNYOK ÉLETTARTAMÁRA Báder Ernő - Kertész Tamás Kertészné, Győrffy Eszter-
PEST MEGYEI KORMÁNYHIVATAL
 PEST MEGYEI KORMÁNYHIVATAL Élelmiszerlánc-biztonsági, Földhivatali, Növény- és Talajvédelmi, Erdészeti Főosztály Állategészségügyi Osztály 1135 Budapest, Lehel u. 43.47. SERTÉS HÍZÉKONYSÁGI ÉS VÁGÁSI VÉGTERMÉKTESZT
PEST MEGYEI KORMÁNYHIVATAL Élelmiszerlánc-biztonsági, Földhivatali, Növény- és Talajvédelmi, Erdészeti Főosztály Állategészségügyi Osztály 1135 Budapest, Lehel u. 43.47. SERTÉS HÍZÉKONYSÁGI ÉS VÁGÁSI VÉGTERMÉKTESZT
Juh és szarvasmarha tenyésztési programok fejlesztését megalapozó kutatások
 Komlósi István Juh és szarvasmarha tenyésztési programok fejlesztését megalapozó kutatások című MTA doktori értekezésének a bírálata A Magyar Tudományos Akadémia Doktori Tanácsának a megtisztelő felkérése
Komlósi István Juh és szarvasmarha tenyésztési programok fejlesztését megalapozó kutatások című MTA doktori értekezésének a bírálata A Magyar Tudományos Akadémia Doktori Tanácsának a megtisztelő felkérése
A sertéshústermelés takarmányozásának a hatékonysága
 Feed Price/100 kg A sertéshústermelés takarmányozásának a hatékonysága A sertéshústermelés hatékonyságát alapvetően befolyásolja a takarmányárak és a sertés felvásárlási árának szintje és aránya. A takarmány
Feed Price/100 kg A sertéshústermelés takarmányozásának a hatékonysága A sertéshústermelés hatékonyságát alapvetően befolyásolja a takarmányárak és a sertés felvásárlási árának szintje és aránya. A takarmány
Nincs öntermékenyítés, de a véges méret miatt a párosodó egyedek bizonyos valószínűséggel rokonok, ezért kerül egy
 Véges populációméret okozta beltenyésztettség incs öntermékenyítés, de a véges méret miatt a párosodó egyedek bizonyos valószínűséggel rokonok, ezért kerül egy utódba 2 IBD allél Előadásról: -F t (-/2)
Véges populációméret okozta beltenyésztettség incs öntermékenyítés, de a véges méret miatt a párosodó egyedek bizonyos valószínűséggel rokonok, ezért kerül egy utódba 2 IBD allél Előadásról: -F t (-/2)
KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN
 MTA DOKTORI ÉRTEKEZÉS KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN DR. NAGY ISTVÁN Ph.D., mezőgazdaságtudomány KAPOSVÁR 2016 1 Tartalom 1. BEVEZETÉS... 4 1.1 Előzmények... 4 1.2. Célkitűzés...
MTA DOKTORI ÉRTEKEZÉS KVANTITATÍV GENETIKAI VIZSGÁLATOK MULTIPARA ÁLLATFAJOKBAN DR. NAGY ISTVÁN Ph.D., mezőgazdaságtudomány KAPOSVÁR 2016 1 Tartalom 1. BEVEZETÉS... 4 1.1 Előzmények... 4 1.2. Célkitűzés...
Altruizmus. Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között?
ét növeli. Lehet-e önző egyedek között?") Altruizmus Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között? Altruizmus rokonok között A legtöbb másolat az adott génről vagy az egyed
Altruizmus Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között? Altruizmus rokonok között A legtöbb másolat az adott génről vagy az egyed
y ij = µ + α i + e ij
 Elmélet STATISZTIKA 3. Előadás Variancia-analízis Lineáris modellek A magyarázat a függő változó teljes heterogenitásának két részre bontását jelenti. A teljes heterogenitás egyik része az, amelynek okai
Elmélet STATISZTIKA 3. Előadás Variancia-analízis Lineáris modellek A magyarázat a függő változó teljes heterogenitásának két részre bontását jelenti. A teljes heterogenitás egyik része az, amelynek okai
Beszámoló feltöltése (zárójelentés)
") Beszámoló feltöltése (zárójelentés) Az OTKA kutatási feladat keretében született eredményeket folyamatosan publikáltuk, ezért e helyen a beszámolási útmutató szerint az elvégzett munkát csupán röviden
Beszámoló feltöltése (zárójelentés) Az OTKA kutatási feladat keretében született eredményeket folyamatosan publikáltuk, ezért e helyen a beszámolási útmutató szerint az elvégzett munkát csupán röviden
Genetikai fejlesztések az állatjólét szolgálatában. Bikal november 22.
 Genetikai fejlesztések az állatjólét szolgálatában Bikal 2018. november 22. Alacsonyabb malac elhullás Alacsonyabb hizlaldai kiesés Javuló koca élettartam Magasabb hatékonyság Herélés elkerülése Állatjólét
Genetikai fejlesztések az állatjólét szolgálatában Bikal 2018. november 22. Alacsonyabb malac elhullás Alacsonyabb hizlaldai kiesés Javuló koca élettartam Magasabb hatékonyság Herélés elkerülése Állatjólét
JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK
 MTA DOKTORI ÉRTEKEZÉS JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK Komlósi István 2012 TARTALOMJEGYZÉK 1. BEVEZETÉS A TENYÉSZTÉSI PROGRAM. 4. 2. CÉLKITŰZÉSEK.. 11. 3. MÓDSZERTANI
MTA DOKTORI ÉRTEKEZÉS JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK Komlósi István 2012 TARTALOMJEGYZÉK 1. BEVEZETÉS A TENYÉSZTÉSI PROGRAM. 4. 2. CÉLKITŰZÉSEK.. 11. 3. MÓDSZERTANI
Altruizmus. Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között?
ét növeli. Lehet-e önző egyedek között?") Altruizmus Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között? Altruizmus rokonok között A legtöbb másolat az adott génről vagy az egyed
Altruizmus Altruizmus: a viselkedés az adott egyed fitneszét csökkenti, de másik egyed(ek)ét növeli. Lehet-e önző egyedek között? Altruizmus rokonok között A legtöbb másolat az adott génről vagy az egyed
A genetikai sodródás
 A genetikai sodródás irányított, nem véletlenszerű Mindig a jobb nyer! természetes szelekció POPULÁCIÓ evolúció POPULÁCIÓ A kulcsszó: változékonyság a populáción belül POPULÁCIÓ nem irányított, véletlenszerű
A genetikai sodródás irányított, nem véletlenszerű Mindig a jobb nyer! természetes szelekció POPULÁCIÓ evolúció POPULÁCIÓ A kulcsszó: változékonyság a populáción belül POPULÁCIÓ nem irányított, véletlenszerű
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI
 ÉRTEKEZÉS TÉZISEI") DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője: HORN PÉTER MTA rendes tagja Témavezető: DR. CSATÓ LÁSZLÓ a mezőgazdaság
DOKTORI (PhD) ÉRTEKEZÉS TÉZISEI KAPOSVÁRI EGYETEM ÁLLATTUDOMÁNYI KAR Sertés- és Kisállattenyésztési Tanszék A doktori iskola vezetője: HORN PÉTER MTA rendes tagja Témavezető: DR. CSATÓ LÁSZLÓ a mezőgazdaság
STATISZTIKA. A maradék független a kezelés és blokk hatástól. Maradékok leíró statisztikája. 4. A modell érvényességének ellenőrzése
 4. A modell érvényességének ellenőrzése STATISZTIKA 4. Előadás Variancia-analízis Lineáris modellek 1. Függetlenség 2. Normális eloszlás 3. Azonos varianciák A maradék független a kezelés és blokk hatástól
4. A modell érvényességének ellenőrzése STATISZTIKA 4. Előadás Variancia-analízis Lineáris modellek 1. Függetlenség 2. Normális eloszlás 3. Azonos varianciák A maradék független a kezelés és blokk hatástól
Szelekció. Szelekció. A szelekció típusai. Az allélgyakoriságok változása 3/4/2013
 Szelekció Ok: több egyed születik, mint amennyi túlél és szaporodni képes a sikeresség mérése: fitnesz Szelekció Ok: több egyed születik, mint amennyi túlél és szaporodni képes a sikeresség mérése: fitnesz
Szelekció Ok: több egyed születik, mint amennyi túlél és szaporodni képes a sikeresség mérése: fitnesz Szelekció Ok: több egyed születik, mint amennyi túlél és szaporodni képes a sikeresség mérése: fitnesz
Módszertani Intézeti Tanszéki Osztály. A megoldás részletes mellékszámítások hiányában nem értékelhető!
 BGF KKK Módszertani Intézeti Tanszéki Osztály Budapest, 2012.. Név:... Neptun kód:... Érdemjegy:..... STATISZTIKA II. VIZSGADOLGOZAT Feladatok 1. 2. 3. 4. 5. 6. Összesen Szerezhető pontszám 21 20 7 22
BGF KKK Módszertani Intézeti Tanszéki Osztály Budapest, 2012.. Név:... Neptun kód:... Érdemjegy:..... STATISZTIKA II. VIZSGADOLGOZAT Feladatok 1. 2. 3. 4. 5. 6. Összesen Szerezhető pontszám 21 20 7 22
A BREEDPLAN-t a nagy húsmarhatenyésztı országok széles körben használják Magyarország
 BSI.1 1 A BREEDPLAN-t a nagy húsmarhatenyésztı országok széles körben használják Magyarország BSI.2 2 MI A TENYÉSZÉRTÉK? Egy állat GENETIKAI ÉRTÉKÉNEK becslése egy adott tulajdonságban pl. 400-napos súlyban,
BSI.1 1 A BREEDPLAN-t a nagy húsmarhatenyésztı országok széles körben használják Magyarország BSI.2 2 MI A TENYÉSZÉRTÉK? Egy állat GENETIKAI ÉRTÉKÉNEK becslése egy adott tulajdonságban pl. 400-napos súlyban,
Kvantitatív genetika Nagy, István
 Kvantitatív genetika Nagy, István Kvantitatív genetika írta Nagy, István Publication date 2011 Tartalom... iv... v... vi... vii 1. Bevezetés a kvantitatív genetikába... 1 2. Statisztikai fogalmak... 3
Kvantitatív genetika Nagy, István Kvantitatív genetika írta Nagy, István Publication date 2011 Tartalom... iv... v... vi... vii 1. Bevezetés a kvantitatív genetikába... 1 2. Statisztikai fogalmak... 3
TENYÉSZTÉSSZERVEZÉS. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 TENYÉSZTÉSSZERVEZÉS Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A tenyésztési hatóság felülvizsgálati tevékenysége Előadás áttekintése Az állattenyésztési törvény és a különböző
TENYÉSZTÉSSZERVEZÉS Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A tenyésztési hatóság felülvizsgálati tevékenysége Előadás áttekintése Az állattenyésztési törvény és a különböző
FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI
 FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI statisztika 8 VIII. REGREssZIÓ 1. A REGREssZIÓs EGYENEs Két valószínűségi változó kapcsolatának leírására az eddigiek alapján vagy egy numerikus
FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI statisztika 8 VIII. REGREssZIÓ 1. A REGREssZIÓs EGYENEs Két valószínűségi változó kapcsolatának leírására az eddigiek alapján vagy egy numerikus
A mangalica sertés takarmányozásának sajátosságai. Sárközi Tamás UBM Feed kft
 A mangalica sertés takarmányozásának sajátosságai Sárközi Tamás UBM Feed kft Az előadás tárgya 1, Malac takarmányozás 2, Koca takarmányozás 3, Hízó takarmányozás 4, Malacszám növelésének lehetősége A SERTÉSTAKARMÁNYOZÁSI
A mangalica sertés takarmányozásának sajátosságai Sárközi Tamás UBM Feed kft Az előadás tárgya 1, Malac takarmányozás 2, Koca takarmányozás 3, Hízó takarmányozás 4, Malacszám növelésének lehetősége A SERTÉSTAKARMÁNYOZÁSI
Statisztika I. 8. előadás. Előadó: Dr. Ertsey Imre
 Statisztika I. 8. előadás Előadó: Dr. Ertsey Imre Minták alapján történő értékelések A statisztika foglalkozik. a tömegjelenségek vizsgálatával Bizonyos esetekben lehetetlen illetve célszerűtlen a teljes
Statisztika I. 8. előadás Előadó: Dr. Ertsey Imre Minták alapján történő értékelések A statisztika foglalkozik. a tömegjelenségek vizsgálatával Bizonyos esetekben lehetetlen illetve célszerűtlen a teljes
Biomatematika 13. Varianciaanaĺızis (ANOVA)
") Szent István Egyetem Állatorvos-tudományi Kar Biomatematikai és Számítástechnikai Tanszék Biomatematika 13. Varianciaanaĺızis (ANOVA) Fodor János Copyright c Fodor.Janos@aotk.szie.hu Last Revision Date:
Szent István Egyetem Állatorvos-tudományi Kar Biomatematikai és Számítástechnikai Tanszék Biomatematika 13. Varianciaanaĺızis (ANOVA) Fodor János Copyright c Fodor.Janos@aotk.szie.hu Last Revision Date:
ÁLLATTENYÉSZTÉSI GENETIKA
 TÁMOP-4..2-08//A-2009-000 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by European
TÁMOP-4..2-08//A-2009-000 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by European
SZARVASMARHA TELJESÍTMÉNYVIZSGÁLATI KÓDEX 2002.
 SZARVASMARHA TELJESÍTMÉNYVIZSGÁLATI KÓDEX 3. 2002. Készítette: a Szarvasmarha Teljesítményvizsgálati Kódex Szerkesztő Bizottság Dr. Sebestyén Sándor, a bizottság elnöke Országos Mezőgazdasági Minősítő
SZARVASMARHA TELJESÍTMÉNYVIZSGÁLATI KÓDEX 3. 2002. Készítette: a Szarvasmarha Teljesítményvizsgálati Kódex Szerkesztő Bizottság Dr. Sebestyén Sándor, a bizottság elnöke Országos Mezőgazdasági Minősítő
Intelligens Rendszerek Elmélete. Párhuzamos keresés genetikus algoritmusokkal
 Intelligens Rendszerek Elmélete Dr. Kutor László Párhuzamos keresés genetikus algoritmusokkal http://mobil.nik.bmf.hu/tantargyak/ire.html login: ire jelszó: IRE0 IRE / A természet általános kereső algoritmusa:
Intelligens Rendszerek Elmélete Dr. Kutor László Párhuzamos keresés genetikus algoritmusokkal http://mobil.nik.bmf.hu/tantargyak/ire.html login: ire jelszó: IRE0 IRE / A természet általános kereső algoritmusa:
Az egyetlen automatizált állományelemző program.
 Az egyetlen automatizált állományelemző program. Bemutatkozás Peter van Beek, MSc Key Account Manager, Európa és Közel- Kelet Holland tenyésztői családból származom A Wageningen Egyetemen szereztem meg
Az egyetlen automatizált állományelemző program. Bemutatkozás Peter van Beek, MSc Key Account Manager, Európa és Közel- Kelet Holland tenyésztői családból származom A Wageningen Egyetemen szereztem meg
Populációbecslés és monitoring. Eloszlások és alapstatisztikák
 Populációbecslés és monitoring Eloszlások és alapstatisztikák Eloszlások Az eloszlás megadja, hogy milyen valószínűséggel kapunk egy adott intervallumba tartozó értéket, ha egy olyan populációból veszünk
Populációbecslés és monitoring Eloszlások és alapstatisztikák Eloszlások Az eloszlás megadja, hogy milyen valószínűséggel kapunk egy adott intervallumba tartozó értéket, ha egy olyan populációból veszünk
FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI
 FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI statisztika 4 IV. MINTA, ALAPsTATIsZTIKÁK 1. MATEMATIKAI statisztika A matematikai statisztika alapfeladatát nagy általánosságban a következőképpen
FEGYVERNEKI SÁNDOR, Valószínűség-sZÁMÍTÁs És MATEMATIKAI statisztika 4 IV. MINTA, ALAPsTATIsZTIKÁK 1. MATEMATIKAI statisztika A matematikai statisztika alapfeladatát nagy általánosságban a következőképpen
Matematikai alapok és valószínőségszámítás. Statisztikai becslés Statisztikák eloszlása
 Matematikai alapok és valószínőségszámítás Statisztikai becslés Statisztikák eloszlása Mintavétel A statisztikában a cél, hogy az érdeklõdés tárgyát képezõ populáció bizonyos paramétereit a populációból
Matematikai alapok és valószínőségszámítás Statisztikai becslés Statisztikák eloszlása Mintavétel A statisztikában a cél, hogy az érdeklõdés tárgyát képezõ populáció bizonyos paramétereit a populációból
XXI. századi lehetőségek a húsmarhatenyésztésben. Dr. Komlósi István egyetemi tanár Debreceni Egyetem
 XXI. századi lehetőségek a húsmarhatenyésztésben Dr. Komlósi István egyetemi tanár Debreceni Egyetem Tenyészcélok a húsmarhatenyésztésben Termelési Reprodukciós Vágási tulajdonságok Termelési tulajdonságok
XXI. századi lehetőségek a húsmarhatenyésztésben Dr. Komlósi István egyetemi tanár Debreceni Egyetem Tenyészcélok a húsmarhatenyésztésben Termelési Reprodukciós Vágási tulajdonságok Termelési tulajdonságok
Természetes szelekció és adaptáció
 Természetes szelekció és adaptáció Amiről szó lesz öröklődő és variábilis fenotípus természetes szelekció adaptáció evolúció 2. Természetes szelekció Miért fontos a természetes szelekció (TSZ)? 1. C.R.
Természetes szelekció és adaptáció Amiről szó lesz öröklődő és variábilis fenotípus természetes szelekció adaptáció evolúció 2. Természetes szelekció Miért fontos a természetes szelekció (TSZ)? 1. C.R.
Statisztika I. 11. előadás. Előadó: Dr. Ertsey Imre
 Statisztika I. 11. előadás Előadó: Dr. Ertsey Imre Összefüggés vizsgálatok A társadalmi gazdasági élet jelenségei kölcsönhatásban állnak, összefüggnek egymással. Statisztika alapvető feladata: - tényszerűségek
Statisztika I. 11. előadás Előadó: Dr. Ertsey Imre Összefüggés vizsgálatok A társadalmi gazdasági élet jelenségei kölcsönhatásban állnak, összefüggnek egymással. Statisztika alapvető feladata: - tényszerűségek
Kettőnél több csoport vizsgálata. Makara B. Gábor MTA Kísérleti Orvostudományi Kutatóintézet
 Kettőnél több csoport vizsgálata Makara B. Gábor MTA Kísérleti Orvostudományi Kutatóintézet Gyógytápszerek (kilokalória/adag) Három gyógytápszer A B C 30 5 00 10 05 08 40 45 03 50 35 190 Kérdések: 1. Van-e
Kettőnél több csoport vizsgálata Makara B. Gábor MTA Kísérleti Orvostudományi Kutatóintézet Gyógytápszerek (kilokalória/adag) Három gyógytápszer A B C 30 5 00 10 05 08 40 45 03 50 35 190 Kérdések: 1. Van-e
Adatok statisztikai értékelésének főbb lehetőségei
 Adatok statisztikai értékelésének főbb lehetőségei 1. a. Egy- vagy kétváltozós eset b. Többváltozós eset 2. a. Becslési problémák, hipotézis vizsgálat b. Mintázatelemzés 3. Szint: a. Egyedi b. Populáció
Adatok statisztikai értékelésének főbb lehetőségei 1. a. Egy- vagy kétváltozós eset b. Többváltozós eset 2. a. Becslési problémák, hipotézis vizsgálat b. Mintázatelemzés 3. Szint: a. Egyedi b. Populáció
Tejtermelési adatok vizsgálata egy hazai magyartarka tenyészetben Analysis of milk production data in a Hungarian Simmental Cattle farm
 Egri Edit 1, 2 - Holló István 2 - Húth Balázs 2 - Holló Gabriella 2 Tejtermelési adatok vizsgálata egy hazai magyartarka tenyészetben Analysis of milk production data in a Hungarian Simmental Cattle farm
Egri Edit 1, 2 - Holló István 2 - Húth Balázs 2 - Holló Gabriella 2 Tejtermelési adatok vizsgálata egy hazai magyartarka tenyészetben Analysis of milk production data in a Hungarian Simmental Cattle farm
A HAZAI SERTÉSÁLLOMÁNYOK GENETIKAI POTENCIÁLJÁNAK ÉRTÉKELÉSE, TELJESÍTMÉNYVIZSGÁLATI EREDMÉNYEIK ALAPJÁN
 ÁLLATTENYÉSZTÉS ÉS TAKARMÁNYOZÁS, 2009. 58. 5. 397 410. 397 A HAZAI SERTÉSÁLLOMÁNYOK GENETIKAI POTENCIÁLJÁNAK ÉRTÉKELÉSE, TELJESÍTMÉNYVIZSGÁLATI EREDMÉNYEIK ALAPJÁN RADNÓCZI LÁSZLÓ KÖVÉR GYÖRGY FARKAS
ÁLLATTENYÉSZTÉS ÉS TAKARMÁNYOZÁS, 2009. 58. 5. 397 410. 397 A HAZAI SERTÉSÁLLOMÁNYOK GENETIKAI POTENCIÁLJÁNAK ÉRTÉKELÉSE, TELJESÍTMÉNYVIZSGÁLATI EREDMÉNYEIK ALAPJÁN RADNÓCZI LÁSZLÓ KÖVÉR GYÖRGY FARKAS
Számítógépes döntéstámogatás. Genetikus algoritmusok
 BLSZM-10 p. 1/18 Számítógépes döntéstámogatás Genetikus algoritmusok Werner Ágnes Villamosmérnöki és Információs Rendszerek Tanszék e-mail: werner.agnes@virt.uni-pannon.hu BLSZM-10 p. 2/18 Bevezetés 1950-60-as
BLSZM-10 p. 1/18 Számítógépes döntéstámogatás Genetikus algoritmusok Werner Ágnes Villamosmérnöki és Információs Rendszerek Tanszék e-mail: werner.agnes@virt.uni-pannon.hu BLSZM-10 p. 2/18 Bevezetés 1950-60-as
Sebes pisztráng ivadékok Myxobolus cerebralis (Myxozoa) okozta kergekórra való fogékonysága a tenyészállomány genetikai diverzitásának függvényében
 okozta kergekórra való fogékonysága a tenyészállomány genetikai diverzitásának függvényében") Sebes pisztráng ivadékok Myxobolus cerebralis (Myxozoa) okozta kergekórra való fogékonysága a tenyészállomány genetikai diverzitásának függvényében Eszterbauer Edit, Forró Barbara, Tolnai Zoltán, Guti
Sebes pisztráng ivadékok Myxobolus cerebralis (Myxozoa) okozta kergekórra való fogékonysága a tenyészállomány genetikai diverzitásának függvényében Eszterbauer Edit, Forró Barbara, Tolnai Zoltán, Guti
Evolúció. Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet
 Evolúció Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet Mi az evolúció? Egy folyamat: az élőlények tulajdonságainak változása a környezethez való alkalmazkodásra Egy
Evolúció Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet Mi az evolúció? Egy folyamat: az élőlények tulajdonságainak változása a környezethez való alkalmazkodásra Egy
Dr. habil. Maróti György
 infokommunikációs technológiák III.8. MÓDSZER KIDOLGOZÁSA ALGORITMUSOK ÁTÜLTETÉSÉRE KIS SZÁMÍTÁSI TELJESÍTMÉNYŰ ESZKÖZÖKBŐL ÁLLÓ NÉPES HETEROGÉN INFRASTRUKTÚRA Dr. habil. Maróti György maroti@dcs.uni-pannon.hu
infokommunikációs technológiák III.8. MÓDSZER KIDOLGOZÁSA ALGORITMUSOK ÁTÜLTETÉSÉRE KIS SZÁMÍTÁSI TELJESÍTMÉNYŰ ESZKÖZÖKBŐL ÁLLÓ NÉPES HETEROGÉN INFRASTRUKTÚRA Dr. habil. Maróti György maroti@dcs.uni-pannon.hu
Populációbecslések és monitoring 1. gyakorlat. Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb
 Populációbecslések és monitoring 1. gyakorlat Nem minden állat látható fogásos módszerek Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb 1. Egyszerű arányváltozás - zárt populáció,
Populációbecslések és monitoring 1. gyakorlat Nem minden állat látható fogásos módszerek Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb 1. Egyszerű arányváltozás - zárt populáció,
Populációbecslések és monitoring 1. gyakorlat. Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb
 Populációbecslések és monitoring 1. gyakorlat Nem minden állat látható fogásos módszerek Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb 1. Egyszerű arányváltozás - zárt populáció,
Populációbecslések és monitoring 1. gyakorlat Nem minden állat látható fogásos módszerek Elvonásos módszerek az adatokat pl. a vadászok is gyűjthetik, olcsóbb 1. Egyszerű arányváltozás - zárt populáció,
Evolúció. Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet
 Evolúció Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet Mi az evolúció? Egy folyamat: az élőlények tulajdonságainak változása a környezethez való alkalmazkodásra Egy
Evolúció Dr. Szemethy László egyetemi docens Szent István Egyetem VadVilág Megőrzési Intézet Mi az evolúció? Egy folyamat: az élőlények tulajdonságainak változása a környezethez való alkalmazkodásra Egy
A HÁZINYULAK NAGYÜZEMI TARTÁSÁNAK KÖVETELMÉNYEI
 TÁMOP-4.1.1.F-14/1/KONV-2015-0006 Egység, alprojekt/alprogram neve Rendezvény cím, dátum A HÁZINYULAK NAGYÜZEMI TARTÁSÁNAK KÖVETELMÉNYEI Előadó neve SZÁRMAZÁS, ELTERJEDÉS, DOMESZTIKÁCIÓ Nyúlszerű rágcsálók
TÁMOP-4.1.1.F-14/1/KONV-2015-0006 Egység, alprojekt/alprogram neve Rendezvény cím, dátum A HÁZINYULAK NAGYÜZEMI TARTÁSÁNAK KÖVETELMÉNYEI Előadó neve SZÁRMAZÁS, ELTERJEDÉS, DOMESZTIKÁCIÓ Nyúlszerű rágcsálók
Kvantitatív genetikai alapok április
 Kvantitatív genetikai alapok 2018. április A vizsgálható tulajdonságok köre: egyed - szám Egyedek morfológiai tulajdonságai: testméretek, arányok, testtömeg Egyedek fiziológiai tulajdonságai: vérnyomás,
Kvantitatív genetikai alapok 2018. április A vizsgálható tulajdonságok köre: egyed - szám Egyedek morfológiai tulajdonságai: testméretek, arányok, testtömeg Egyedek fiziológiai tulajdonságai: vérnyomás,
Kettőnél több csoport vizsgálata. Makara B. Gábor
 Kettőnél több csoport vizsgálata Makara B. Gábor Három gyógytápszer elemzéséből az alábbi energia tartalom adatok származtak (kilokalória/adag egységben) Három gyógytápszer elemzésébô A B C 30 5 00 10
Kettőnél több csoport vizsgálata Makara B. Gábor Három gyógytápszer elemzéséből az alábbi energia tartalom adatok származtak (kilokalória/adag egységben) Három gyógytápszer elemzésébô A B C 30 5 00 10
1. Magyar szürke szarvasmarha tenyésztési programhoz kapcsolódó meg nem felelések
 4B. számú melléklet: Az egyes fajták tenyésztési program előírásainak meg nem felelése a súlyosság és tartósság szempontjából az ellenőri megállapítások elvégzéséhez 1. Magyar szürke szarvasmarha tenyésztési
4B. számú melléklet: Az egyes fajták tenyésztési program előírásainak meg nem felelése a súlyosság és tartósság szempontjából az ellenőri megállapítások elvégzéséhez 1. Magyar szürke szarvasmarha tenyésztési
Biomatematika 2 Orvosi biometria
 Biomatematika 2 Orvosi biometria 2017.02.05. Orvosi biometria (orvosi biostatisztika) Statisztika: tömegjelenségeket számadatokkal leíró tudomány. A statisztika elkészítésének menete: tanulmányok (kísérletek)
Biomatematika 2 Orvosi biometria 2017.02.05. Orvosi biometria (orvosi biostatisztika) Statisztika: tömegjelenségeket számadatokkal leíró tudomány. A statisztika elkészítésének menete: tanulmányok (kísérletek)
[Biomatematika 2] Orvosi biometria
![[Biomatematika 2] Orvosi biometria](/thumbs/54/33134285.jpg "[Biomatematika 2] Orvosi biometria") [Biomatematika 2] Orvosi biometria 2016.02.08. Orvosi biometria (orvosi biostatisztika) Statisztika: tömegjelenségeket számadatokkal leíró tudomány. A statisztika elkészítésének menete: tanulmányok (kísérletek)
[Biomatematika 2] Orvosi biometria 2016.02.08. Orvosi biometria (orvosi biostatisztika) Statisztika: tömegjelenségeket számadatokkal leíró tudomány. A statisztika elkészítésének menete: tanulmányok (kísérletek)
Populációbecslések és monitoring
 Populációbecslések és monitoring A becslés szerepe az ökológiában és a vadgazdálkodásban. A becslési módszerek csoportosítása. Teljes számlálás. Statisztikai alapfogalmak. Fontos lehet tudnunk, hogy hány
Populációbecslések és monitoring A becslés szerepe az ökológiában és a vadgazdálkodásban. A becslési módszerek csoportosítása. Teljes számlálás. Statisztikai alapfogalmak. Fontos lehet tudnunk, hogy hány
Perspektívák a sertések precíziós takarmányozásában. Halas Veronika, PhD Kaposvári Egyetem Takarmányozástani tanszék
 Perspektívák a sertések precíziós takarmányozásában Halas Veronika, PhD Kaposvári Egyetem Takarmányozástani tanszék Az EU országok vágott sertés kibocsájtása (millió hízó) Eurostat (2014) 1000 koca Az
Perspektívák a sertések precíziós takarmányozásában Halas Veronika, PhD Kaposvári Egyetem Takarmányozástani tanszék Az EU országok vágott sertés kibocsájtása (millió hízó) Eurostat (2014) 1000 koca Az
ÁLLATTENYÉSZTÉSI GENETIKA
 TÁMOP-4..-08//A-009-000 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by European
TÁMOP-4..-08//A-009-000 project ÁLLATTENYÉSZTÉSI GENETIKA University of Debrecen University of West Hungary University of Pannonia The project is supported by the European Union and co-financed by European
A Markowitz modell: kvadratikus programozás
 A Markowitz modell: kvadratikus programozás Harry Markowitz 1990-ben kapott Közgazdasági Nobel díjat a portfolió optimalizálási modelljéért. Ld. http://en.wikipedia.org/wiki/harry_markowitz Ennek a legegyszer
A Markowitz modell: kvadratikus programozás Harry Markowitz 1990-ben kapott Közgazdasági Nobel díjat a portfolió optimalizálási modelljéért. Ld. http://en.wikipedia.org/wiki/harry_markowitz Ennek a legegyszer
93/2008. (VII. 24.) FVM rendelet
 FVM rendelet") 93/2008. (VII. 24.) FVM rendelet a védett őshonos állatfajták genetikai fenntartásának rendjéről Az állattenyésztésről szóló 1993. évi CXIV. törvény 49. (1) bekezdés a) pont 6., 7. és 8. alpontjaiban kapott
93/2008. (VII. 24.) FVM rendelet a védett őshonos állatfajták genetikai fenntartásának rendjéről Az állattenyésztésről szóló 1993. évi CXIV. törvény 49. (1) bekezdés a) pont 6., 7. és 8. alpontjaiban kapott
Transzformátor, Mérőtranszformátor Állapot Tényező szakértői rendszer Vörös Csaba Tarcsa Dániel Németh Bálint Csépes Gusztáv
 Transzformátor, Mérőtranszformátor Állapot Tényező szakértői rendszer Vörös Csaba Tarcsa Dániel Németh Bálint Csépes Gusztáv Áttekintés A Rendszer jelentősége Állapotjellemzők MérőTranszformátor Állapot
Transzformátor, Mérőtranszformátor Állapot Tényező szakértői rendszer Vörös Csaba Tarcsa Dániel Németh Bálint Csépes Gusztáv Áttekintés A Rendszer jelentősége Állapotjellemzők MérőTranszformátor Állapot
Populációbecslések és monitoring
 Populációbecslések és monitoring A becslés szerepe az ökológiában és a vadgazdálkodásban. A becslési módszerek csoportosítása. Teljes számlálás. Statisztikai alapfogalmak. Fontos lehet tudnunk, hogy hány
Populációbecslések és monitoring A becslés szerepe az ökológiában és a vadgazdálkodásban. A becslési módszerek csoportosítása. Teljes számlálás. Statisztikai alapfogalmak. Fontos lehet tudnunk, hogy hány
RÉGI FAJTA ÚJ ERÉNYEKKEL A MAGYARTARKA TENYÉSZTÉS EREDMÉNYEI
 RÉGI FAJTA ÚJ ERÉNYEKKEL A MAGYARTARKA TENYÉSZTÉS EREDMÉNYEI Dr. HÚTH Balázs tenyésztés- és marketingvezető Tarka Marhafesztivál Bonyhád 2012. augusztus 10-11. A fajta szerepe a hazai mezőgazdaságban A
RÉGI FAJTA ÚJ ERÉNYEKKEL A MAGYARTARKA TENYÉSZTÉS EREDMÉNYEI Dr. HÚTH Balázs tenyésztés- és marketingvezető Tarka Marhafesztivál Bonyhád 2012. augusztus 10-11. A fajta szerepe a hazai mezőgazdaságban A
Állatállomány, 2013. június 1., (előzetes adatok)
") 213/85 Összeállította: Központi Statisztikai Hivatal www.ksh.hu VII. évfolyam 85. szám 213. október 25. Állatállomány, 213. június 1., (előzetes adatok) A tartalomból 1 Bevezető 1 Szarvasmarha-állomány
213/85 Összeállította: Központi Statisztikai Hivatal www.ksh.hu VII. évfolyam 85. szám 213. október 25. Állatállomány, 213. június 1., (előzetes adatok) A tartalomból 1 Bevezető 1 Szarvasmarha-állomány
KUTATÁSMÓDSZERTAN 4. ELŐADÁS. A minta és mintavétel
 KUTATÁSMÓDSZERTAN 4. ELŐADÁS A minta és mintavétel 1 1. A MINTA ÉS A POPULÁCIÓ VISZONYA Populáció: tágabb halmaz, alapsokaság a vizsgálandó csoport egésze Minta: részhalmaz, az alapsokaság azon része,
KUTATÁSMÓDSZERTAN 4. ELŐADÁS A minta és mintavétel 1 1. A MINTA ÉS A POPULÁCIÓ VISZONYA Populáció: tágabb halmaz, alapsokaság a vizsgálandó csoport egésze Minta: részhalmaz, az alapsokaság azon része,
TENYÉSZTÉSSZERVEZÉS. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010
 TENYÉSZTÉSSZERVEZÉS Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A gazdasági állatfajok elismert tenyésztőszervezeteinek ismertetése Tenyésztőszervezeti munka a lótenyésztésben
TENYÉSZTÉSSZERVEZÉS Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A gazdasági állatfajok elismert tenyésztőszervezeteinek ismertetése Tenyésztőszervezeti munka a lótenyésztésben
129/2004. (VIII. 25.) FVM rendelet
 FVM rendelet") 129/2004. (VIII. 25.) FVM rendelet a tenyészállat, illetve szaporítóanyag behozatalának és kivitelének szakmai előírásairól Az állattenyésztésről szóló 1993. évi CXIV. törvény (a továbbiakban: Tv.) 49.
129/2004. (VIII. 25.) FVM rendelet a tenyészállat, illetve szaporítóanyag behozatalának és kivitelének szakmai előírásairól Az állattenyésztésről szóló 1993. évi CXIV. törvény (a továbbiakban: Tv.) 49.
Állatállomány, 2012. június 1.
 212/56 Összeállította: Központi Statisztikai Hivatal www.ksh.hu VI. évfolyam 56. szám 212. augusztus 3. Állatállomány, 212. június 1. A tartalomból 1 Bevezető 1 Szarvasmarha-állomány 2 állomány 3 Baromfiállomány
212/56 Összeállította: Központi Statisztikai Hivatal www.ksh.hu VI. évfolyam 56. szám 212. augusztus 3. Állatállomány, 212. június 1. A tartalomból 1 Bevezető 1 Szarvasmarha-állomány 2 állomány 3 Baromfiállomány
Mangalica tanácskozás Debrecen 2014. Augusztus 18. Dr. Radnóczi László
 A minisztérium feladatai a védett őshonos állatfajták megőrzésével és genetikai fenntartásával kapcsolatban Mangalica tanácskozás Debrecen 2014. Augusztus 18. Dr. Radnóczi László Jogi szabályozás -az
A minisztérium feladatai a védett őshonos állatfajták megőrzésével és genetikai fenntartásával kapcsolatban Mangalica tanácskozás Debrecen 2014. Augusztus 18. Dr. Radnóczi László Jogi szabályozás -az
6. Előadás. Vereb György, DE OEC BSI, október 12.
 6. Előadás Visszatekintés: a normális eloszlás Becslés, mintavételezés Reprezentatív minta A statisztika, mint változó Paraméter és Statisztika Torzítatlan becslés A mintaközép eloszlása - centrális határeloszlás
6. Előadás Visszatekintés: a normális eloszlás Becslés, mintavételezés Reprezentatív minta A statisztika, mint változó Paraméter és Statisztika Torzítatlan becslés A mintaközép eloszlása - centrális határeloszlás
y ij = µ + α i + e ij STATISZTIKA Sir Ronald Aylmer Fisher Példa Elmélet A variancia-analízis alkalmazásának feltételei Lineáris modell
 Példa STATISZTIKA Egy gazdálkodó k kukorica hibrid termesztése között választhat. Jelöljük a fajtákat A, B, C, D-vel. Döntsük el, hogy a hibridek termesztése esetén azonos terméseredményre számíthatunk-e.
Példa STATISZTIKA Egy gazdálkodó k kukorica hibrid termesztése között választhat. Jelöljük a fajtákat A, B, C, D-vel. Döntsük el, hogy a hibridek termesztése esetén azonos terméseredményre számíthatunk-e.
BIOLÓGIA HÁZIVERSENY 1. FORDULÓ BIOKÉMIA, GENETIKA BIOKÉMIA, GENETIKA
 BIOKÉMIA, GENETIKA 1. Nukleinsavak keresztrejtvény (12+1 p) 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 1. A nukleinsavak a.-ok összekapcsolódásával kialakuló polimerek. 2. Purinvázas szerves bázis, amely az
BIOKÉMIA, GENETIKA 1. Nukleinsavak keresztrejtvény (12+1 p) 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 1. A nukleinsavak a.-ok összekapcsolódásával kialakuló polimerek. 2. Purinvázas szerves bázis, amely az
JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK. MTA Doktori értekezés tézisei. Komlósi István
 JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK MTA Doktori értekezés tézisei Komlósi István 2012 1. BEVEZETÉS A Tenyésztési Program A tenyésztési program elemei: 1. a tenyészcél
JUH ÉS SZARVASMARHA TENYÉSZTÉSI PROGRAMOK FEJLESZTÉSÉT MEGALAPOZÓ KUTATÁSOK MTA Doktori értekezés tézisei Komlósi István 2012 1. BEVEZETÉS A Tenyésztési Program A tenyésztési program elemei: 1. a tenyészcél
KÖVETKEZTETŐ STATISZTIKA
 ÁVF GM szak 2010 ősz KÖVETKEZTETŐ STATISZTIKA A MINTAVÉTEL BECSLÉS A sokasági átlag becslése 2010 ősz Utoljára módosítva: 2010-09-07 ÁVF Oktató: Lipécz György 1 A becslés alapfeladata Pl. Hányan láttak
ÁVF GM szak 2010 ősz KÖVETKEZTETŐ STATISZTIKA A MINTAVÉTEL BECSLÉS A sokasági átlag becslése 2010 ősz Utoljára módosítva: 2010-09-07 ÁVF Oktató: Lipécz György 1 A becslés alapfeladata Pl. Hányan láttak
Hátterükben egyetlen gén áll, melynek általában számottevő a viselkedésre gyakorolt hatása, öröklési mintázata jellegzetes.
 Múlt órán: Lehetséges tesztfeladatok: Kitől származik a variáció-szelekció paradigma, mely szerint az egyéni, javarészt öröklött különbségek között a társadalmi harc válogat? Fromm-Reichmann Mill Gallton
Múlt órán: Lehetséges tesztfeladatok: Kitől származik a variáció-szelekció paradigma, mely szerint az egyéni, javarészt öröklött különbségek között a társadalmi harc válogat? Fromm-Reichmann Mill Gallton
Matematikai statisztika c. tárgy oktatásának célja és tematikája
 Matematikai statisztika c. tárgy oktatásának célja és tematikája 2015 Tematika Matematikai statisztika 1. Időkeret: 12 héten keresztül heti 3x50 perc (előadás és szeminárium) 2. Szükséges előismeretek:
Matematikai statisztika c. tárgy oktatásának célja és tematikája 2015 Tematika Matematikai statisztika 1. Időkeret: 12 héten keresztül heti 3x50 perc (előadás és szeminárium) 2. Szükséges előismeretek:
25 év a magyartarka tenyésztésben
 25 év a magyartarka tenyésztésben Regionális közgyűlések 2014. február 25. - március 6. Dr. Füller Imre ügyvezető igazgató Az egyesület megalakulása és főbb állomásai 1989. június 16. Bonyhád, megalakult
25 év a magyartarka tenyésztésben Regionális közgyűlések 2014. február 25. - március 6. Dr. Füller Imre ügyvezető igazgató Az egyesület megalakulása és főbb állomásai 1989. június 16. Bonyhád, megalakult
Matematikai alapok és valószínőségszámítás. Középértékek és szóródási mutatók
 Matematikai alapok és valószínőségszámítás Középértékek és szóródási mutatók Középértékek A leíró statisztikák talán leggyakrabban használt csoportját a középértékek jelentik. Legkönnyebben mint az adathalmaz
Matematikai alapok és valószínőségszámítás Középértékek és szóródási mutatók Középértékek A leíró statisztikák talán leggyakrabban használt csoportját a középértékek jelentik. Legkönnyebben mint az adathalmaz