AZ INTERMEDIER ANYAGCSERE ENDOPLAZMÁS RETIKULUMHOZ KÖTÖTT REAKCIÓINAK INVENTÁRIUMA
|
|
|
- Máté Papp
- 8 évvel ezelőtt
- Látták:
Átírás
1 AZ ENDOPLAZMÁS RETIKULUM Az endoplazmás retikulum (ER) ciszternák és tubulusok hálózata, mely a magmembránt is magába foglalja. A legtöbb eukarióta sejt legnagyobb organelluma, melyet folyamatos membrán határol. A májsejtben, ahol az ER igen fejlett, a sejttérfogat 10, a sejtfehérje 20, illetve a membránok több mint 90 százalékát képviseli. Néhány sejttípusban viszont szinte kizárólag a magmembrán alkotja az ER hálózatát. AZ INTERMEDIER ANYAGCSERE ENDOPLAZMÁS RETIKULUMHOZ KÖTÖTT REAKCIÓINAK INVENTÁRIUMA Az ER-ban található enzimek többnyire nem véletlenül működnek ebben az organellumban. A ER lokalizáció legjellemzőbb okai: a) A reakció termékei exportra készülnek és a vezikuláris transzport révén távoznak a sejtből (szekréciós fehérjék, lipoproteinek). b) A reakcióútban szereplő enzimek láncolatot képeznek - szubsztrát channeling - pl. elektrontranszfer lánc (mikroszómális elektrontranszfer a biotranszformáció első fázisában). c) Az enzimkatalízis optimumához szükséges az ER lumenében található sajátos környezet (chaperonok, UDP-glukuronoziltranszferázok). d) A reakció szubsztrátjai és/vagy termékei lipidoldékony vegyületek lévén az ER membránjában oldva találhatók (citokróm P450 enzimek, koleszterol szintézis, deszaturáz, stb). Az ER-ben található legfontosabb reakciók: a) szénhidrát anyagcsere: glukóz-6-foszfatáz rendszer, hexuronsav ciklus befejező lépései, gulonolakton oxidáz; b) lipid anyagcsere: zsírsavlánc elongációja, deszaturáció, triglicerid szintézis, koleszterol és más biológiai izoprének szintézise, koleszterol észteresítése (ACAT), lipoprotein szintézis, foszfolipid bioszintézis legtöbb lépése; 1

2 c) biotranszformációs reakciók: citokróm P450 izoenzimek által katalizált reakciók (szteroidok egyes hidroxilációi, D-vitamin aktiválódása, xenobiotikumok oxigenálása), glukuronidáció (bilirubin, szteroidok és xenobiotikumok); d) szekréciós fehérjék szintézise és poszttranszlációs módosítása (prolil- és lizilhidroxiláció, diszulfid híd képződés, γ-karboxiláció, N- és O-glikozilálás). (A fenti reakciók részletesen megtalálhatók a tankönyvben, illetve korábbi konzultációs anyagokban. A továbbiakhoz célszerű a glukóz-6-foszfatáz rendszer, az UDP-glukuronoziltranszferázok, valamint az ER-hez kötött riboszómákon történő fehérjeszintézis és poszttranszlációs módosítások átismétlése.) AZ ENDOPLAZMÁS RETIKULUM TRANSZPORTFOLYAMATAI Az endoplazmás retikulum luminális kompartimentumának összetétele nem teljesen ismert, de számos molekula tekintetében jelentősen különbözik a citoszólétól. Jól ismert pl. a citoszólnál több mint három nagyságrenddel magasabb intraluminális kálcium koncentráció. A protonkoncentráció szintén magasabb a lumenben. A fehérjeszekrécióban résztvevő sejtek ER-ában a fehérjekoncentráció is igen magas. A kis molekulák közül ismert, hogy a glutation diszulfid és az aszkorbinsav koncentrációja magasabb, míg a (redukált) glutationé alacsonyabb a lumenben. Az intermedier anyagcsere legtöbb köztitermékének intraluminális koncentrációja a metodikai nehézségek miatt (l. később) nem ismert. Feltehető, hogy legtöbbjük nincs vagy csak igen alacsony koncentrációban van jelen az ER lumenben. A transzport célja: 1. A szekréciós és membránfehérjék poszttranszlációs módosításához szükséges szubsztátok szállítása befelé és a keletkezett végtermékek szállítása kifelé (ATP/ADP, cukor nukleotidok/nukleozid monofoszfátok, aszkorbát/dehidroaszkorbát, glutation) 2. A sejtből részben vagy teljesen exocitózissal távozó termékek prekurzorainak szállítása intraluminális aktív centrummal rendelkező enzimekhez (UDP-glukuronsav, glukóz-6-foszfát, dehidroaszkorbát) 3. Az általában regulációs okokból intraluminális enzimek szubsztrátjainak és termékeinek szállítása (UDP-glukuronsav, glukuronidok, UMP, glukóz-6-foszfát, glukóz, foszfát) A transzport mechanizmusa: 1. Aktív transzport: Ca 2+ -ATPáz (SERCA), H + -ATPáz 2. Csatornák: Ins(1,4,5)P 3 -receptor, ryanodine receptor 3. Facilitált diffúzió 2

3 3.1. Uniporterek: glukóz, glukóz-6-foszfát, Pi, PP, glutation, dehidroaszkorbát, kisebb glukuronidok (ms<300) 3.2. Antiporterek: ATP, nukleotid-cukrok, nagyobb glukuronidok (ms>300) A traszport vizsgálata A jelenlegi módszerek nem alkalmasak a transzporterek ligandjainak in vivo kimutatására az ER lumenében. Ezért ismereteink in vitro rekonstruált kísérleti rendszereken, mikroszómákon és proteoliposzómákon végzett kísérletekből származnak. A sejt homogenizálása során az ER rendszere feltöredezik és kisméretű vezikulákat (mikroszómák) alkot. A mikroszómák membránja megőrzi eredeti irányultságát (a citoszól felőli oldal van kívül), de elvész az ER esetleges (bár vitatott) heterogenitása. A tisztított transzporter liposzómába építhető. A transzport vizsgálatának módszerei mikroszómában és proteoliposzómában fényszórás gyors szűrés gyors precipitáció elve ozmotikus hatásra bekövetkező zsugorodás vagy tágulás befolyásolja a vezikulák fényszórását az inkubációs közeg és a vezikulák gyors elválasztása szűréssel az inkubációs közeg és a vezikulák gyors elválasztása polietilénglikol precipitációval és centrifugálással detektálás időigénye real time ~5 s 1-2 min szükséges ligandkoncentráció magas (5-100 mm), a legtöbb ligand esetében nem fiziológiás tetszőleges (de jelzett ligandot igényel) tetszőleges, a detektáláshoz alkalmazott módszertől függ szükséges fehérje µg µg 1-5 mg szükséges eszköz fluoriméter folyadék szcintillátor HPLC A transzporterek molekuláris azonosítása a) biokémiai megközelítés: a transzport funkcionális jellemzése membránfehérjék frakcionálása és tisztítása rekonstrukció proteoliposzómában b) molekuláris biológiai megközelítés: transzporter gének keresése bakteriális analógiák alapján molekuláris biológiai és biokémiai igazolás 3
 alkot.")
4 A transzporterek rekonstrukciója proteoliposzómában A rekonstrukció több fontos célt szolgálhat: a) a tisztítás ellenőrzése; b) a transzport mechanimusának tanulmányozása (pl. antiport mechanizmus) c) kinetikai mérések; d) a transzporter és a membrán kölcsönhatásának vizsgálata; e) a transzporter szabályozásának tanulmányozása. A rekonstrukció lépései: a) az organellum membránjának szolubilizálása detergensekkel b) a detergens eltávolítása c) a fehérjék beépítése unilamelláris foszfatidilkolin liposzómákba (ismételt fagyasztás-olvasztás ciklusokkal vagy hígítással) A fenti módszerrel a következő ligandokat transzportáló antiportereket sikerült rekonstruálni (a Golgiból): CMP-sziálsav, UDP-galaktóz, UDP-glukuronsav, ATP, UDP-N-acetil-galaktózamin, foszfoadenozin-foszfoszulfát. Az UDP-xilóz antiporter rekonstrukciójához a Golgi saját lipidjeit kellett felhasználni. Az ER transzportereinek rekonstruciója valamiért sokkal nehezebb, eddig csak az ATP/ADP antiportert rekonstruálták. Az antiporterek általános jellegzetességei: a) a transzport többé-kevésbé organellum-specifikus (ER vs. Golgi); b) a teljes molekula transzportálódik, nics metabolizmus a transzport során, a transzporter nem enzim; c) a transzport hőmérsékletfüggő és telíthető, a tipikus Km 1-10 µm; d) a transzport kompetitíven gátolható a ligand analógjaival; e) a ligand akkumulálódik a lumenben. Az ehhez szükséges energia egy intravezikuláris molekula koncentrációgrádiens irányába történő, kifelé irányuló transzportjából származik (antiport mechanizmus). Ez a molekula általában a ligand intravezikuláris metabolizmusából származó nukleozid monofoszfát (v. difoszfát). Az ER két legjobban ismert, transzporterek működéséhez kötött enzimrendszere a glukóz-6-foszfatáz és az UDP-glukuronoziltranszferáz rendszer. Az alábbiakban ezek működését ismertetem kissé részletesebben. Mindkét esetben a katalítikus aktivitású alegység (maga az enzim) aktív centruma a lumen felé irányul. Jellegzetes tulajdonságuk a latencia: az enzimaktivitás alacsony az intakt 4
 A fenti módszerrel a következő ligandokat transzportáló antiportereket sikerült rekonstruálni (a Golgiból): CMP-sziálsav, UDP-galaktóz,")
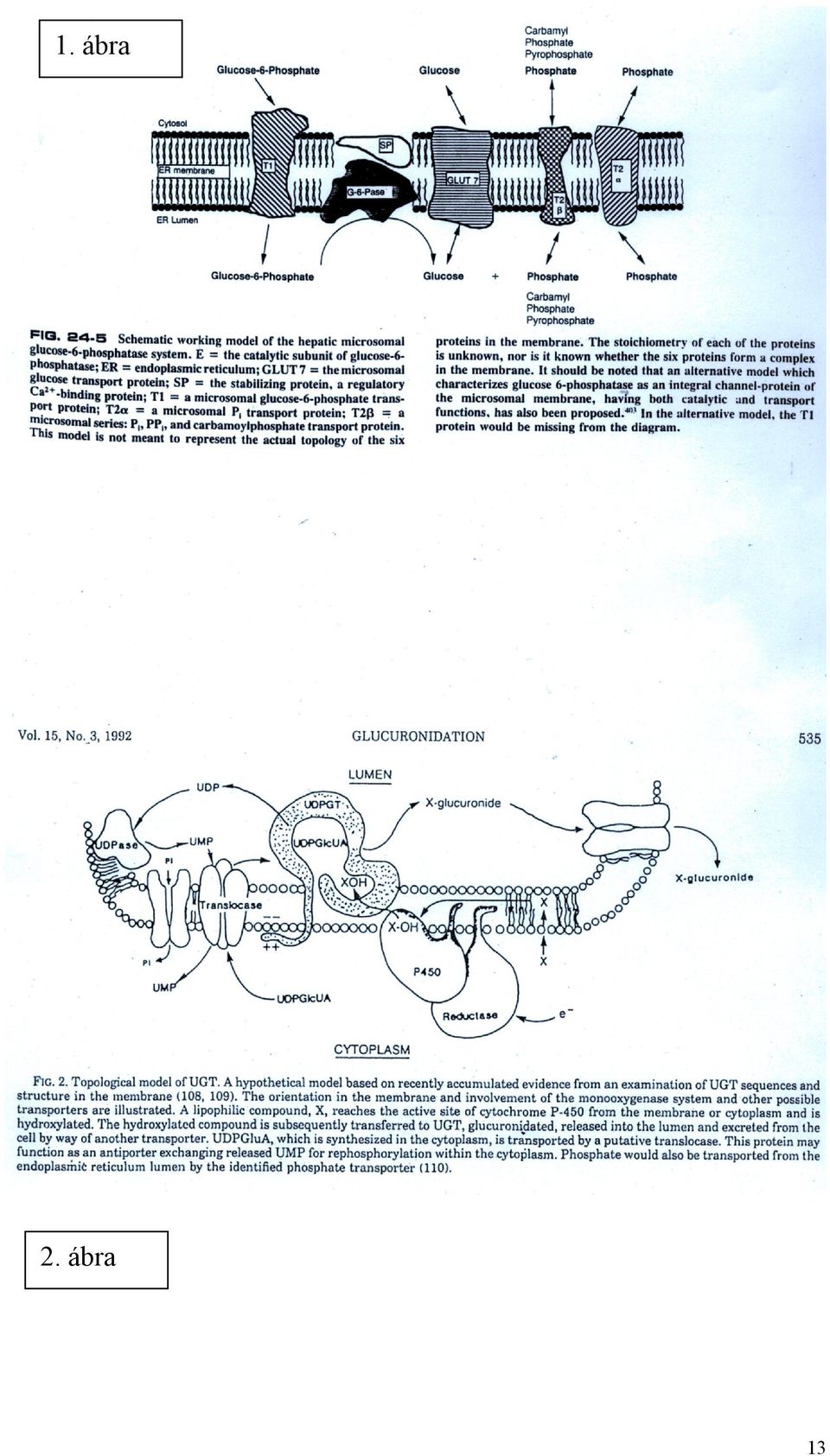
5 mikroszómális vezikulákban, és a membrán integritását megszüntető kezelés (detergens, pórusképzők, ultrahang, ismételt fagyasztás-olvasztás stb.) hatására növekszik. latencia (%) = x (enzimaktivitás intakt vezikulákban / teljes enzimaktivitás) A latencia jelensége jelzi, hogy a sebességmeghatározó lépés a szubsztrát transzportja, egyben lehetőséget ad a transzport sebességének indirekt meghatározására is (a transzport sebessége enzimaktivitás). A glukóz-6-foszfatáz rendszert legalább (?) három transzporter egészíti ki (l. 1. ábra). Mindegyik működési zavara ismert, a glikogéntárolási betegségek 1. típusának (GSD 1 v. Gierke-kór) különböző altípusait hozza létre. Az altípusok különböző manifesztációs formája valószínűvé teszi, hogy valóban különböző transzporterek hiány felelős az egyes kórképekért. Jelenleg molekuláris szinten csak a glukóz-6-foszfát transzporter (G6PT) ismert, a többi transzportert csak funkcionálisan jellemezték. A G6PT specifikus G6P-ra, míg a G6Páz bármilyen hexóz-foszfátot hidrolizálni tud. Tehát a transzporter biztosítja a G6Páz rendszer specificitását. A G6PT-t bakteriális analógiák alapján emberi hólyagrák cdns könyvtárból azonosították (Gerin et al, 1997). A transzporter elsősorban a glukoneogenezisre képes szervekben (máj, vese) expresszálódik, de jelen van szinte minden sejttípusban. A transzportert 429 aminosav alkotja, 10 transzmembrán hélixet képezve (3. ábra). Az agyból egy 22 aminosavval hosszabb variánsát lehetett kimutatni. A GSD 1b altípusban a gén különféle mutációit észlelték, melyek elsősorban a transzmembrán szakaszokat érintik. Ismertek a transzport gátlószerei is (klorogénsav és származékai), melyek esetleg a diabetes terápiájában is helyet kaphatnak majd. A G6PT mutációi esetén a mikroszómális G6P transzport defektusa kimutatható (fokozott v. teljes latencia), de a transzporter rekonstrukciója proteoliposzómában még nem volt sikeres. A G6PT gyakorlatilag minden sejtben expresszálódik, ennek ellenére a GSD 1b csak a glukoneogenetikus szerveket és a granulocitákat érinti. A kórképet a Gierke-kór általános jellemzőin (hipoglikémia, hiperlipémia, laktacidózis, máj- és vesemegnagyobbodás, máj adenómák) kívül granulocitopénia, csökkent granulocita funkciók és fertőzésekre való hajlam jellemzi. A granulociták nem tartalmaznak G6Pázt, glukóz termelésére nem képesek, így a transzporter funkciója misztikus, de fontos. Lehetséges hipotézisek: a G6P akkumuláció az ER lumenben a kálcium szekvesztrációhoz szükséges, vagy a transzporter G6P szenzorként működve befolyásolhatja a sejtciklust és apoptózist. A foszfát (és pirofoszfát) transzportjáért felelős fehérjék szerkezete nem ismert. A transzport zavaráért nem a G6PT gén mutációi felelősek (mint azt egy ideig hitték). A glukóz transzportját feltehetőleg egy GLUT típusú transzporter mediálja (mely más hexózokat és dehidroaszkorbátot is szállít). A transzporter viszonylag kis kapacitású, így a G6P hidrolízise során intraluminálisan glukóz akkumulálódik. Ez felveti, hogy a glukóz részben exocitózis útján hagyja el a májat, amit GLUT2 knockout állatokban is igazoltak. 5
 különböző altípusait hozza létre.")
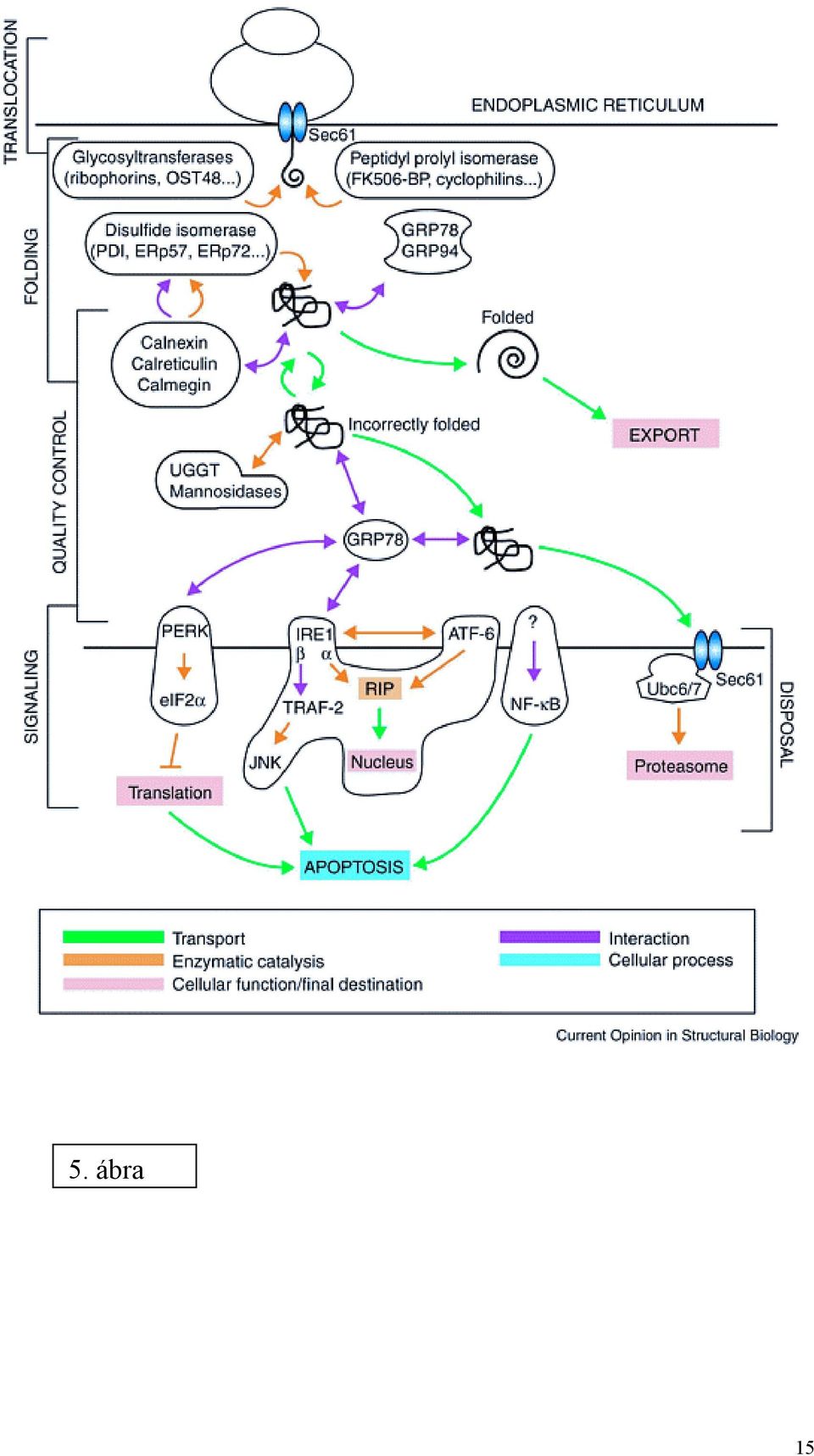
6 Az UDP-glukuronoziltranszferáz rendszer transzporterei csak funkcionálisan ismertek. Szemben a G6P rendszerrel, ahol a facilitált diffúzióért uniporterek felelősek, itt elsősorban antiporterek működnek (2. és 4. ábra). Az antiport általában elektroneutrális (azonos töltésű molekulák cserélődnek ki). Az UDP-glukuronsav befelé irányuló transzportját UMP távozása egészíti ki. (Lehetséges, hogy a folyamat kétlépcsős: az UDPGA UDP-N-acetilglukózaminnal cserélődik ki, s az utóbbi UMP-vel.) A képződött nagyobb molekulasúlyú glukuronidok távozása pedig szintén UDPGA belépésével járhat együtt. Az aniporterek tehát kevéssé specifikusnak tűnnek. MOLEKULÁRIS MECHANIZMUSOK AZ ENDOPLAZMÁS RETIKULUMBAN FOLYÓ FEHÉRJESZINTÉZISBEN (5. ábra) Fehérje folding (peptidil prolil cisz/transz izomeráz, oligoszaharil-transzfer komplex, glukóz-regulált chaperonok (GRP78, GRP94 stb), lektinszerű chaperonok (calnexin, calreticulin, calmegin), tiol-diszulfid oxidoreduktázok (protein diszulfid izomeráz, ERp72, Erp59 stb) Minőségellenőrzés (a fenti chaperonok, UDP-glukóz:glikoprotein glikoziltranszferáz, glukozidáz II, mannozidázok) ER-sejtmag jelpályák (UPR, EOR) Proteolízis (ERAD) A fenti folyamatok közül a konzultációs anyag az utóbbi kettőt tárgyalja. JELÁTVITEL AZ ENDOPLAZMÁS RETIKULUM ÉS A SEJTMAG KÖZÖTT Az ER egyik legfontosabb funkciója az exportra kerülő makromolekulák (fehérjék, lipoproteinek) szintézise, foldingja és poszttranszlációs módosítása, valamint a makromolekuláris protein komplexek összeállítása. Mielőtt a fehérje elhagyja az ER lumenét (a vezikuláris transzport során), gondos minőségellenőrzésen esik át. A vizsgálaton megbukott fehérjék a lumenben maradnak, további sorsuk kétféle lehet. Vagy kapnak még egy lehetőséget, további foldingon esnek át a lumenben, fokozva a zsúfoltságot (ER túltöltés, ER overload, ERO), vagy visszakerülnek a citoszólba, ahol 6

7 proteaszóma által végrehajtott proteolízisen esnek át (ER-asszociált degradáció, ERAD). Az ER folding funkcióját rontó legfontosabb hatások: a) A fehérjeglikoziláció gátlószerei: 2-deoxiglukóz, brefeldin A, tunikamicin, éhezés. b) Intraluminális kálciumot csökkentő szerek: tapsigargin, ciklopiazonsav (Ca 2+ - ATPáz gátlók), A23187, ionomicin és más ionofórok. Hatásuk közvetett: a legtöbb intraluminális enzim (chaperonok, glikoziláció enzimei, oxidoreduktázok) optimális működéséhez szükséges a lumen normális kálciumkoncentrációja (>400 µm). c) Intraluminális redoxpotenciált befolyásoló redukálószerek: ditiotreitol, merkaptoetanol. A protein diszulfid izomeráz és más tiol-diszulfid oxidoreduktázok működéséhez szükséges optimális redoxpotenciált (-180 mv) változtatják meg, jelenlétükben a diszulfid kötések képződése zavart szenved. d) Integráns membránfehérjék és szekréciós fehérjék túltermelése (pl. vírusinfekció), mutáns fehérjék termelése. A fenti eseményekről sajátos jelpályák szállítanak információt a sejtmagba. (Mivel a magot körülvevő membrán analóg az ER-mal, a jelpályának nem kell szükségszerűen érintenie a citoszólt.) A legismertebbek közülük: a selejtfehérje-válasz (unfolded protein response, UPR), az ER túltöltés válasz (EOR) és a szterol-válasz. UPR A jelpálya részleteit élesztőben írták le először, de legtöbb elemét már humán sejtekben is megtalálták (6. ábra). Az ER lumenében a minőségi hibás fehérjéket egy chaperon fehérje, a Kar2p köti. Ez a chaperon a hősokkfehérjék Hsp70 családjába tartozó BiP analógja. A hibás fehérjék tömeges megjelenése kivonja a Kar2p-t egy ER transzmembrán fehérjével, az Ire1p-vel alkotott kötéséből. Az Ire1p egy kináz aktivitású enzim, melynek citoszól felé néző doménje egy ribonukleázzal (RNázL) 7
 optimális működéséhez szükséges a lumen normális kálciumkoncentrációja (>400 µm).")
8 homológ, míg luminális doménje kötődik a Kar2p-hez. Kar2p nélkül két Ire1p molekula homodimerizálódik, transz-autofoszforilálódik és így aktiválódik. Az aktív Ire1p egy trns ligázzal együttműködve kihasít egy darabot a HAC1 elsődleges mrns-ből. Az átalakított mrns kijut a citoszólba, átíródik fehérjévé, s az így képződött Hac1p transzkripciós faktorként működve fokozza az UPR cél-gének átíródását. Az ide tartozó gének promotere tartalmaz egy 22 bázispárból álló UPR elemet (UPRE), melyet minden UPR során indukálódó fehérje génjében megtaláltak. Az indukálódó fehérjék egyrészt szükségesek a foszfolipid bioszintézishez, másrészt a foldinghoz (luminális chaperonok, glikoziltranszferázok stb.). Élesztőben majdnem négyszáz gén tartozik az UPR által mediált csoportba, körülbelül 200-nak ismert a funkciója. Az UPR-t kiegészíti a fehérjék transzlációját gátló mechanizmus, mely az eukarióta iniciációs faktor eif2α foszforilációján keresztül érvényesül. Az ER membránjában leírtak egy másik, az Ire1p-hez hasonló transzmembrán kinázt. A PERK nevű fehérje intraluminális doménje, mely teljesen analóg az Ire1p hasonló doménjével, érzékeli a hibás fehérjék megjelenését. A citoszólban lévő domén protein kináz aktivitású, az eif2α 51-es szerinjét foszforilálja, ami gátolja a transzlációt (Harding et al., 1999). Egy harmadik lehetséges útvonal: az Ire1p humán homológja nem vesz részt az UPR cél-gének aktiválásában, hanem a fehérjeszintézist gátolja a 28S rrns specifikus hasításával (Iwawaki et al., 2001). EOR A fehérjék torlódása az ER lumenben beindítja az ER túltöltés válasznak nevezett jelpályát (7. ábra). Az EOR-t és az UPR-t kiváltó ágensek részben átfedőek, néhány hatás azonban (pl. vírusfertőzés utáni vírusfehérje túltermelés) csak az EOR jelpályát aktiválja. Az ER túltöltése kálcium felszabadulást vált ki, eddig ismeretlen mechanizmussal. A kálcium permeabilitást megváltozását okozhatja a membránban megjelenő idegen fehérjék Ca 2+ -ATPázt gátló hatása, vagy a magasabb fehérje/foszfolipid arány a membránban, vagy a membrán mechanikai feszülésére érzékeny kálcium csatornák megnyílása. Az emelkedett kálcium koncentráció a citoszólban reaktív oxigén intermediereket termelő enzimeket aktivál (pl. ciklooxigenáz, lipoxigenáz). A továbbiakban a jelpálya már közös számos más szignál 8
, melyet minden UPR során indukálódó fehérje génjében megtaláltak.")
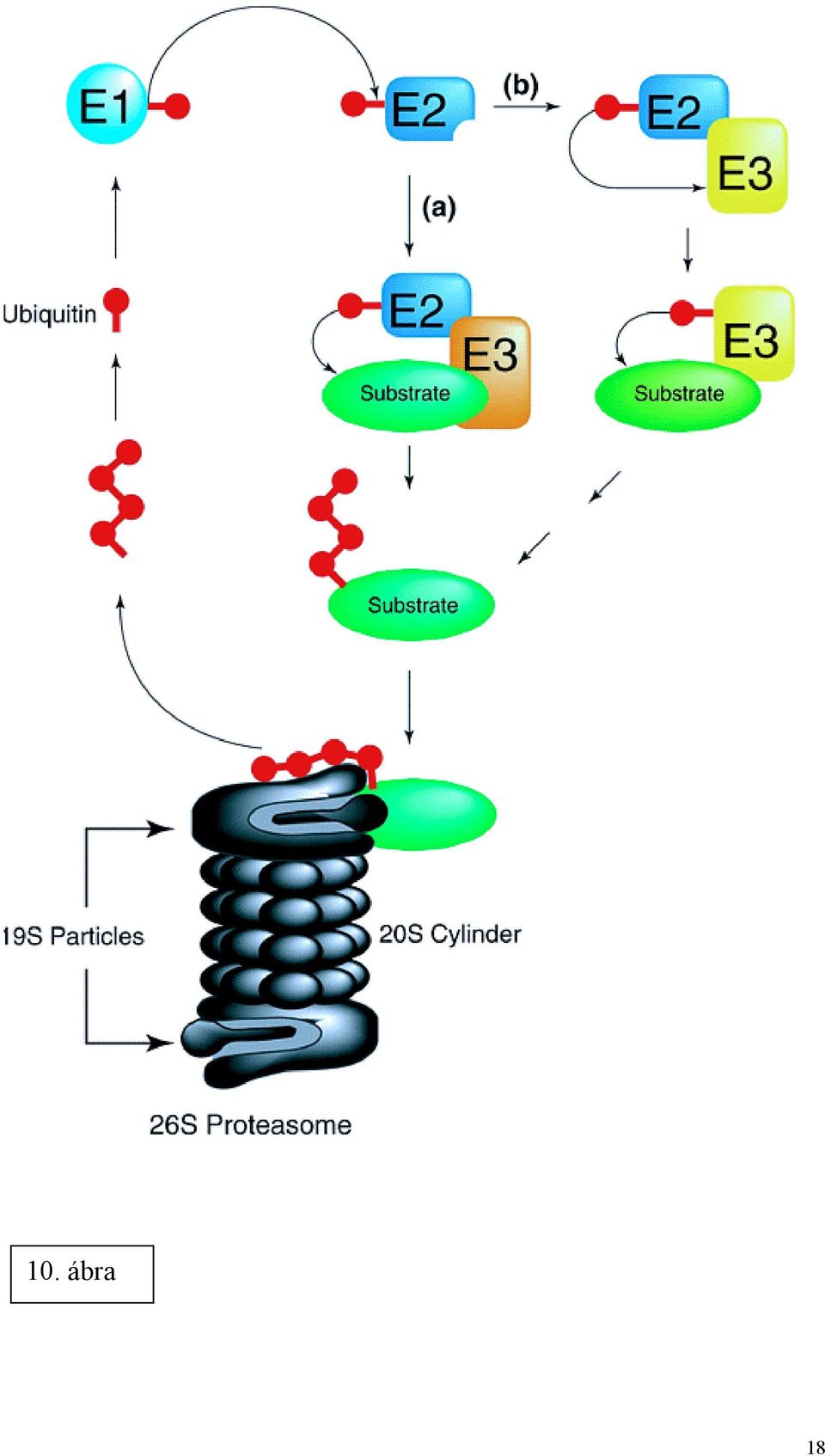
9 transzdukciós útvonallal: a reaktív oxigénszármazékok aktiválják az NF-κB nevű transzkripciós faktort, mely beindítja (egyebek közt) az immunválaszhoz szükséges interferon és citokin szintézist. A sémának megfelelően a hatás modellezhető kálcium felszabadulást kiváltó Ca 2+ -ATPáz gátlószerekkel (tapsigargin, ciklopiazonsav), illetve kivédhető Ca 2+ -kelátorokkal és antioxidánsokkal. Az EOR számos emberi betegségben megfigyelhető, így vírusfertőzésekben és a fehérje folding zavaraival járó kórképekben (Alzheimer-kór, cisztikus fibrózis stb). Szterol válasz A koleszterol az ER-ban szintetizálódik, és épül be a membránokba. Az ER-ban található az az apparátus is, mely a sejt koleszterol ellátottságát érzékeli (8. ábra). Koleszterol hiányában (melyet valószínűleg az ER membránban található SCAP fehérje érzékel), a membránban található SREBP (sterol regulatory element binding protein) két lépéses proteolízisen megy át. A proteolízishez a SCAP hozzájárulása szükséges. Az elhasított, aktív SREBP homodimerizálódik, és aktiválja a koleszterol szintézishez szükséges enzimek és más fehérjék génjeinek transzkripcióját (LDL receptor, HMG-KoA szintáz, HMG-KoA reduktáz, szkvalén szintáz stb). ERAD (ER-asszociált degradáció) Az ER minőségellenőrzési rendszere által visszatartott fehérjék eliminálására az ER speciális módszert alkalmaz. Mivel egy proteolítikus rendszer működtetése az ER lumenében veszélyes lehetne a részlegesen vagy egyáltalán nem natív konformációjú fehérjék jelenléte miatt, melyeket egy ilyen rendszer nagy valószínűséggel megtámadna, a degradációra ítélt fehérjéket el kell távolítani a lumenből. A folyamat egyes lépéseit élesztőben írták le. Az élesztő ER citoszól felé néző felszínén enzimek (Ubc6p, Ubc7p) találhatók, melyek az ubikvitin-konjugációt végző enzimek népes családjába tartoznak (l. tankönyv ábra). Ezek az enzimek egy kis polipeptidet, az ubikvitint (76 aminosav) kötik a lebontásra kerülő fehérjéhez, melyet így fel tud ismerni a 9
.")
10 proteaszóma komplex, mely a peptidkötések hidrolízisét végzi (10. ábra). A folyamatnak ez a része nem specifikus az ER fehérjékre. Kérdés, hogyan jut ki a fehérje a membránból vagy a lumenből a citoszólba. Membránfehérjék esetében a proteaszóma komplex borotvaként működve levághatja a citoszól felé néző fragmentumokat. A luminális fehérjék esetében retrográd transzportot kell feltételezni, melyet alátámaszt az, hogy az ubikvitinált fehérjék egy része glikozilált is, tehát már megjárta az ER lumenét. A proteolítikus mechanizmust ezek szerint kiegészíti egy felismerési rendszer a lumenben és egy membrántranszport szisztéma. A következő elemeit ismerjük a teljes rendszernek (9. ábra): Kar2p (BiP): a hibás fehérjék felismerése és kötése a lumenben Sec61p: transzmembrán csatornát képező fehérje, kétirányú transzportot mediál Sec63p: más fehérjékkel együtt a transzport egyirányúsításáért felelős A fehérjék mutációja megszünteti az ERAD működését, de nem jár az élesztősejt életképtelenségével. Humán kórképek az ERAD egyes komponenseinek feltételezett közreműködésével: Számos bakteriális és növényi toxin (pl. Shiga toxin, ricin) endocitózis révén kerül a célsejtbe, ahol a vezikuláris transzporttal eljutnak egészen az ER lumenig. Mivel támadáspontjuk a citoszólban van (28S rrns), át kell transzportálódniuk az ER membránon. Valószínűleg az ERAD szubsztrátjait utánozza a szerkezetük, s így jutnak ki a citoszólba, de az ubikvitinálódást valahogy elkerülik. A cisztikus fibrózis az egyik leggyakoribb, légúti tünetekkel és hasnyálmirigyelégtelenséggel járó genetikai betegség. A kórképben a CFTR klorid csatorna génjének mutációja miatt nem képződik funkcióképes csatorna a plazma membránban. Leggyakrabban a F508 jelű mutáció fordul elő. Érdekes módon ez a mutáció nem eredményezne funkcióképtelen fehérjét, de az ER minőségellenőrzési rendszere felismeri a mutációt és halálra ítéli a fehérjét, mely így nem jut el rendeltetési helyére, a plazma membránba. Az ER minőségellenőrzési rendszerének elégtelen működése szerepet játszhat a prion betegségekben (bovin spongioform encephalitis, scrapie, Creutzfeld-Jakob kór). Ezekben a kórképekben a prion fehéje (PrP), egy 35 kda tömegű agy glikoprotein a patogenezis kiindulópontja. A PrP normál funkciója ismeretlen, a betegség a PrP egy rendellenes izoformájának a megjelenésével és felhalmozódásával függ össze. A PrP különböző topológiai formájú lehet, integrálódhat vagy beékelődhet az ER membránba. Új megfigyelések szerint az egyik transzmembrán forma potenciálisan patogén lehet; normál körülmények között ezt a formát az ER hibajavító rendszere kiküszöböli. Eddig ismeretlen okból ez a kóros forma néha elkerülheti az ERAD mediált degradációt, eljut más poszt-er kompartimentumokba és kiválthatja a betegség tüneteit. 10

11 Irodalom Cooper GM (1997) The Cell: A Molecular Approach. ASM Press, Washington, USA Subcellular Biochemistry. Vol. 21. Endoplasmic Reticulum. Eds: Borgese N and Harris JR. Plenum Press, New York, 1993 Hirschberg CB, Robbins PW and Abeijon C (1998) Transporters of nucleotide sugars, ATP, and nucleotide sulfate in the endoplasmic reticulum and Golgi apparatus. Annu. Rev. Biochem. 67, Abeijon C, Mandon EC and Hirschberg CB (1997) Transporters of nucleotide sugars, nucleotide sulfate and ATP in the Golgi apparatus. TIBS 22, Chen Y-T and Burchell A (1995) Glycogen storage diseases. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The molecular bases of inherited diseases, 7 th edition. McGraw-Hill, pp Lin B, Pan C-J and Chou JY (2000) Human variant glucose-6-phosphate transporter is active in microsomal transport. Hum. Genet. 107, Marcolongo P., Bánhegyi G., Benedetti A., Hinds C.J., Burchell A.: Liver microsomal transport of glucose-6-phosphate, glucose and phosphate in type 1 glycogen storage disease. J. Clin. Endocrinol. Metab. 83, , 1998 Bánhegyi G., Csala M., Braun L., Marcolongo P., Fulceri R., Mandl J., Benedetti A.: Evidence for a UDP-glucuronic acid - phenol glucuronide antiport in rat liver microsomal vesicles. Biochem. J. 315, , 1996 Corbett EF and Michalak M (2000) Calcium, a signaling molecule in the endoplasmic reticulum? Trends Biochem. Sci. 25, Pahl HL (1999) Signal transduction from the endoplasmic reticulum to the cell nucleus. Physiol. Rev. 79, Hampton RY (2000) Getting the UPR hand on misfolded proteins. Curr. Biol. 10, R518-R521 McCracken AA and Brodsky JL (2000) A molecular portrait of the response to unfolded proteins. Genome Biology 1, ( 11
 Glycogen storage diseases. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The molecular bases of inherited diseases, 7 th edition. McGraw-Hill, pp.")
12 Harding HP, Zhang Y and Ron D (1999) Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 397, Sidrauski C, Chapman R and Walter P (1998) The unfolded protein response: an intracellular signalling pathway with many surprising features. Trends Cell Biol. 8, Iwawaki T. et al. (2001) Translational control by the ER transmembrane kinase/ribonuclease IRE' under ER stress. Nature Cell Biol. 3, Bays NW et al. (2001) Hrd1p/Der3p is a membrane-anchored ubiquitin ligase required for the ERassociated degradation. Nature Cell Biol. 3, Chevet E et al. (2001) The endoplasmic reticulum: integration of protein folding, quality control, signaling and degradation. Curr. Opin. Struct. Biol. 11, Plemper RK and Wolf DH (1999) Retrograde protein translocation: ERADication of secretory proteins in health and disease. Trends Biochem. Sci. 24,
 Hrd1p/Der3p is a membrane-anchored ubiquitin ligase required for the ERassociated degradation. Nature Cell Biol. 3, 24-29 Chevet E et al.")
13 1. ábra 2. ábra 13
14 3. ábra 4. ábra 14

15 5. ábra 15
16 6. ábra 7. ábra 16

17 8. ábra 9. ábra 17

18 10. ábra 18
1b. Fehérje transzport
 1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
1b. Fehérje transzport Fehérje transzport CITOSZÓL Nem-szekretoros útvonal sejtmag mitokondrium plasztid peroxiszóma endoplazmás retikulum Szekretoros útvonal lizoszóma endoszóma Golgi sejtfelszín szekretoros
A FAD transzportjának szerepe az oxidatív fehérje foldingban patkány máj mikroszómákban
 A FAD transzportjának szerepe az oxidatív fehérje foldingban patkány máj mikroszómákban PhD értekezés tézisek Varsányi Marianne 2005 Témavezető: Dr. Bánhegyi Gábor Semmelweis Egyetem Orvosi Vegytani, Molekuláris
A FAD transzportjának szerepe az oxidatív fehérje foldingban patkány máj mikroszómákban PhD értekezés tézisek Varsányi Marianne 2005 Témavezető: Dr. Bánhegyi Gábor Semmelweis Egyetem Orvosi Vegytani, Molekuláris
Antioxidánsok szerepe a fehérje diszulfid kötések kialakulásában SZARKA ANDRÁS
 Semmelweis Egyetem, Molekuláris Orvostudományok Doktori Iskola PATHOBIOKÉMIA DOKTORI PROGRAM Antioxidánsok szerepe a fehérje diszulfid kötések kialakulásában Doktori (Ph.D.) értekezés tézisei SZARKA ANDRÁS
Semmelweis Egyetem, Molekuláris Orvostudományok Doktori Iskola PATHOBIOKÉMIA DOKTORI PROGRAM Antioxidánsok szerepe a fehérje diszulfid kötések kialakulásában Doktori (Ph.D.) értekezés tézisei SZARKA ANDRÁS
A pályázat keretében a következő kérdéseket kívántuk részleteiben vizsgálni:
 A glikogenolízis és glukoneogenezis utolsó, közös lépését katalizáló glukóz-6-foszfatáz egy enzimrendszer, melyben a katalitikus alegység kevéssé specifikus, különböző foszfátésztereket tud hasítani és
A glikogenolízis és glukoneogenezis utolsó, közös lépését katalizáló glukóz-6-foszfatáz egy enzimrendszer, melyben a katalitikus alegység kevéssé specifikus, különböző foszfátésztereket tud hasítani és
transzláció DNS RNS Fehérje A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti fehérjék, transzportfehérjék
 Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció A molekuláris biológia centrális dogmája transzkripció transzláció DNS RNS Fehérje replikáció Reverz transzkriptáz A fehérjék jelenléte nélkülözhetetlen minden sejt számára: enzimek, szerkezeti
Transzláció. Szintetikus folyamatok Energiájának 90%-a
 Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Transzláció Transzláció Fehérje bioszintézis a genetikai információ kifejeződése Szükséges: mrns: trns: ~40 Riboszóma: 4 rrns + ~ 70 protein 20 Aminosav aktiváló enzim ~12 egyéb enzim Szintetikus folyamatok
Pro- és antioxidáns hatások szerepe az endoplazmás retikulum eredetű stresszben és apoptózisban
 Pro- és antioxidáns hatások szerepe az endoplazmás retikulum eredetű stresszben és apoptózisban Az endoplazmás retikulum (ER) számos környezeti és metabolikus hatás szenzora. Mindazon tényezők, melyek
Pro- és antioxidáns hatások szerepe az endoplazmás retikulum eredetű stresszben és apoptózisban Az endoplazmás retikulum (ER) számos környezeti és metabolikus hatás szenzora. Mindazon tényezők, melyek
A téma címe: Mikroszómális glukóz-6-foszfát szerepe granulocita apoptózisában
 Témavezető neve: Dr. Kardon Tamás Zoltán A téma címe: Mikroszómális glukóz-6-foszfát szerepe granulocita apoptózisában A kutatás időtartama: 2004-2007 Tudományos háttér A glukóz-6-foszfatáz multienzim-komplex
Témavezető neve: Dr. Kardon Tamás Zoltán A téma címe: Mikroszómális glukóz-6-foszfát szerepe granulocita apoptózisában A kutatás időtartama: 2004-2007 Tudományos háttér A glukóz-6-foszfatáz multienzim-komplex
A téma címe: Antioxidáns anyagcsere és transzportfolyamatok az endo/szarkoplazmás retikulumban A kutatás időtartama: 4 év
 Témavezető neve: Dr. Csala Miklós A téma címe: Antioxidáns anyagcsere és transzportfolyamatok az endo/szarkoplazmás retikulumban A kutatás időtartama: 4 év Háttér A glutation (GSH) és a glutation diszulfid
Témavezető neve: Dr. Csala Miklós A téma címe: Antioxidáns anyagcsere és transzportfolyamatok az endo/szarkoplazmás retikulumban A kutatás időtartama: 4 év Háttér A glutation (GSH) és a glutation diszulfid
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből.
 Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
Vércukorszint szabályozása: Szénhidrátok monoszacharidok formájában szívódnak fel a vékonybélből. Szövetekben monoszacharid átalakítás enzimjei: Szénhidrát anyagcserében máj központi szerepű. Szénhidrát
1. Előadás Membránok felépítése, mebrán raftok
 1. Előadás Membránok felépítése, mebrán raftok Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis biztosítása Klasszikus folyadékmozaik
1. Előadás Membránok felépítése, mebrán raftok Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis biztosítása Klasszikus folyadékmozaik
Biológiai membránok és membrántranszport
 Biológiai membránok és membrántranszport Biológiai membránok A citoplazma membrán funkciói: térrészek elválasztása (egész sejt, organellumok) transzport jelátvitel Milyen a membrán szerkezete? lipidek
Biológiai membránok és membrántranszport Biológiai membránok A citoplazma membrán funkciói: térrészek elválasztása (egész sejt, organellumok) transzport jelátvitel Milyen a membrán szerkezete? lipidek
Diabéteszes redox változások hatása a stresszfehérjékre
 Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola Pathobiokémia Program Doktori (Ph.D.) értekezés Diabéteszes redox változások hatása a stresszfehérjékre dr. Nardai Gábor Témavezeto:
Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola Pathobiokémia Program Doktori (Ph.D.) értekezés Diabéteszes redox változások hatása a stresszfehérjékre dr. Nardai Gábor Témavezeto:
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét. Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet
 Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
Fehérje szintézis 2. TRANSZLÁCIÓ Molekuláris biológia kurzus 7. hét Kun Lídia Genetikai, Sejt- és immunbiológiai Intézet Gén mrns Fehérje Transzkripció Transzláció A transzkriptum : mrns Hogyan mutatható
TRANSZPORTEREK Szakács Gergely
 TRANSZPORTEREK Szakács Gergely Összefoglalás A biológiai membránokon keresztüli anyagáramlást számos membránfehérje szabályozza. E fehérjék változatos funkciója és megjelenésük mintázata biztosítja a sejtek
TRANSZPORTEREK Szakács Gergely Összefoglalás A biológiai membránokon keresztüli anyagáramlást számos membránfehérje szabályozza. E fehérjék változatos funkciója és megjelenésük mintázata biztosítja a sejtek
Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont
 Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont Doktori tézisek Dr. Konta Laura Éva Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola
Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont Doktori tézisek Dr. Konta Laura Éva Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola
A glükóz reszintézise.
 A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
A glükóz reszintézise. A glükóz reszintézise. A reszintézis nem egyszerű megfordítása a glikolízisnek. A glikolízis 3 irrevezibilis lépése más úton játszódik le. Ennek oka egyrészt energetikai, másrészt
Membrántranszport. Gyógyszerész előadás Dr. Barkó Szilvia
 Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
Vezikuláris transzport
 Molekuláris Sejtbiológia Vezikuláris transzport Dr. habil KŐHIDAI László Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet 2005. november 3. Intracelluláris vezikul uláris transzport Kommunikáció
Molekuláris Sejtbiológia Vezikuláris transzport Dr. habil KŐHIDAI László Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet 2005. november 3. Intracelluláris vezikul uláris transzport Kommunikáció
TRANSZPORTFOLYAMATOK 1b. Fehérjék. 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS
 1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
1b. FEHÉRJÉK TRANSZPORTJA A MEMBRÁNONOKBA ÉS A SEJTSZERVECSKÉK BELSEJÉBE ÁLTALÁNOS DIA 1 Fő fehérje transzport útvonalak Egy tipikus emlős sejt közel 10,000 féle fehérjét tartalmaz (a test pedig összesen
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói
 1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
1. előadás Membránok felépítése, mebrán raftok, caveolák jellemzője, funkciói Plazmamembrán Membrán funkciói: sejt integritásának fenntartása állandó hő, energia, és információcsere biztosítása homeosztázis
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Dr. Csala Miklós OTKA NN 75275
 Az endoplazmás retikulum piridin-nukleotid rendszerének redox változásai: összefüggés az elhízással, a 2-es típusú diabetes-szel és a metabolikus szindrómával Bevezetés A prohormonnak tekinthető kortizon
Az endoplazmás retikulum piridin-nukleotid rendszerének redox változásai: összefüggés az elhízással, a 2-es típusú diabetes-szel és a metabolikus szindrómával Bevezetés A prohormonnak tekinthető kortizon
A koleszterin-anyagcsere szabályozása (Csala Miklós)
") A koleszterin-anyagcsere szabályozása (Csala Miklós) A koleszterin fontos építőeleme az emberi sejteknek, fontos szerepe van a biológiai membránok fluiditásának szabályozásában. E mellett hormonok és epesavak
A koleszterin-anyagcsere szabályozása (Csala Miklós) A koleszterin fontos építőeleme az emberi sejteknek, fontos szerepe van a biológiai membránok fluiditásának szabályozásában. E mellett hormonok és epesavak
térrészek elválasztása transzport jelátvitel Milyen a membrán szerkezete? Milyen a membrán szerkezete? lipid kettısréteg, hidrofil/hidrofób részek
 Biológiai membránok A citoplazma membrán funkciói: Biológiai membránok és membrántranszport térrészek elválasztása (egész sejt, organellumok) transzport jelátvitel Milyen a membrán szerkezete? lipidek
Biológiai membránok A citoplazma membrán funkciói: Biológiai membránok és membrántranszport térrészek elválasztása (egész sejt, organellumok) transzport jelátvitel Milyen a membrán szerkezete? lipidek
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Glikolízis. emberi szervezet napi glukózigénye: kb. 160 g
 Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
Glikolízis Minden emberi sejt képes glikolízisre. A glukóz a metabolizmus központi tápanyaga, minden sejt képes hasznosítani. glykys = édes, lysis = hasítás emberi szervezet napi glukózigénye: kb. 160
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül.
 7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Folyadékkristályok; biológiai és mesterséges membránok
 Folyadékkristályok; biológiai és mesterséges membránok Dr. Voszka István Folyadékkristályok: Átmenet a folyadékok és a kristályos szilárdtestek között (anizotróp folyadékok) Fonal, pálcika, korong alakú
Folyadékkristályok; biológiai és mesterséges membránok Dr. Voszka István Folyadékkristályok: Átmenet a folyadékok és a kristályos szilárdtestek között (anizotróp folyadékok) Fonal, pálcika, korong alakú
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
Szerkezet és funkció kapcsolata a membránműködésben. Folyadékkristályok típusai (1) Dr. Voszka István
 Dr. Voszka István") MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2012/2013 II. félév II. 7. Szerkezet és funkció kapcsolata a membránműködésben Dr. Voszka István II. 21. Liposzómák előállítási módjai Dr.
MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2012/2013 II. félév II. 7. Szerkezet és funkció kapcsolata a membránműködésben Dr. Voszka István II. 21. Liposzómák előállítási módjai Dr.
CzB 2010. Élettan: a sejt
 CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
A transzlokon szerepe kismolekulák transzportjában az endoplazmás retikulumban
 A transzlokon szerepe kismolekulák transzportjában az endoplazmás retikulumban Doktori tézisek Dr. Lizák Beáta Semmelweis Egyetem, Molekuláris Orvostudományok Doktori Iskola, Pathobiokémia program Témavezető:Dr.
A transzlokon szerepe kismolekulák transzportjában az endoplazmás retikulumban Doktori tézisek Dr. Lizák Beáta Semmelweis Egyetem, Molekuláris Orvostudományok Doktori Iskola, Pathobiokémia program Témavezető:Dr.
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
A KOLESZTERIN SZERKEZETE. (koleszterin v. koleszterol)
") 19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
19 11 12 13 C 21 22 20 18 D 17 16 23 24 25 26 27 HO 2 3 1 A 4 5 10 9 B 6 8 7 14 15 A KOLESZTERIN SZERKEZETE (koleszterin v. koleszterol) - a koleszterin vízben rosszul oldódik - szabad formában vagy koleszterin-észterként
Immunológia alapjai. 10. előadás. Komplement rendszer
 Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
Immunológia alapjai 10. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Miért fontos a komplement rendszer? A veleszületett (nem-specifikus) immunválasz része Azonnali válaszreakció A veleszületett
A T sejt receptor (TCR) heterodimer
 heterodimer") Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
Immunbiológia - II A T sejt receptor (TCR) heterodimer 1 kötőhely lánc lánc 14. kromoszóma 7. kromoszóma V V C C EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL lánc: VJ régió lánc: VDJ régió Nincs szomatikus
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Szerkezet és funkció kapcsolata a membránműködésben. Folyadékkristályok típusai (1) Dr. Voszka István
 Dr. Voszka István") MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2015/2016 II. félév Időpont: szerda 17 30-19 00 Helyszín Elméleti Orvostudományi Központ Szent-Györgyi Albert előadóterme II. 3. Szerkezet
MODELLMEMBRÁNOK (LIPOSZÓMÁK) ORVOSI, GYÓGYSZERÉSZI ALKALMAZÁSA 2015/2016 II. félév Időpont: szerda 17 30-19 00 Helyszín Elméleti Orvostudományi Központ Szent-Györgyi Albert előadóterme II. 3. Szerkezet
Immunológia alapjai előadás MHC. szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Antigén felismerés Az ellenanyagok és a B sejt receptorok natív formában
Endocitózis - Exocitózis
 Molekuláris sejtbiológia Endocitózis - Exocitózis Dr. habil.. Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immnubiológiai Intézet Budapest Endocitózis Fagocitózis szilárd fázishoz közel álló
Molekuláris sejtbiológia Endocitózis - Exocitózis Dr. habil.. Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immnubiológiai Intézet Budapest Endocitózis Fagocitózis szilárd fázishoz közel álló
Biokémiai és Molekuláris Biológiai Intézet. Sejtbiológiai alapok. Sarang Zsolt
 Biokémiai és Molekuláris Biológiai Intézet Sejtbiológiai alapok Sarang Zsolt Víz (felnőtt emberi test 57-60%-a víz) Élő szervezetek inorganikus felépítő elemei Anionok (foszfát, klorid, karbonát ion, stb.)
Biokémiai és Molekuláris Biológiai Intézet Sejtbiológiai alapok Sarang Zsolt Víz (felnőtt emberi test 57-60%-a víz) Élő szervezetek inorganikus felépítő elemei Anionok (foszfát, klorid, karbonát ion, stb.)
MITOCHONDRIUM. Molekuláris sejtbiológia: Dr. habil. Kőhidai László egytemi docens Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet
 Molekuláris sejtbiológia: MITOCHONDRIUM külső membrán belső membrán lemezek / crista matrix Dr. habil. Kőhidai László egytemi docens Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Tudomány-történet
Molekuláris sejtbiológia: MITOCHONDRIUM külső membrán belső membrán lemezek / crista matrix Dr. habil. Kőhidai László egytemi docens Semmelweis Egyetem, Genetikai, Sejt- és Immunbiológiai Intézet Tudomány-történet
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE
 ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE") Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
Glükoproteinek (GP) ELŐADÁSVÁZLAT ORVOSTANHALLGATÓK RÉSZÉRE SZTE ÁOK Biokémia Intézet összeállította: dr Keresztes Margit Jellemzők - relative rövid oligoszacharid láncok ( 30) (sok elágazás) (1-85% GP
(1) A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.
 A T sejtek aktiválása (2) Az ön reaktív T sejtek toleranciája. α lánc. β lánc. V α. V β. C β. C α.") Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
Immunbiológia II A T sejt receptor () heterodimer α lánc kötőhely β lánc 14. kromoszóma 7. kromoszóma 1 V α V β C α C β EXTRACELLULÁRIS TÉR SEJTMEMBRÁN CITOSZÓL αlánc: VJ régió β lánc: VDJ régió Nincs
sejt működés jovo.notebook March 13, 2018
 1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
1 A R É F Z S O I B T S Z E S R V E Z D É S I S E Z I N E T E K M O I B T O V N H C J W W R X S M R F Z Ö R E W T L D L K T E I A D Z W I O S W W E T H Á E J P S E I Z Z T L Y G O A R B Z M L A H E K J
DNS replikáció. DNS RNS Polipeptid Amino terminus. Karboxi terminus. Templát szál
 DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
DNS replikáció DNS RNS Polipeptid Amino terminus Templát szál Karboxi terminus Szuper-csavarodott prokarióta cirkuláris DNS Hisztonok komplexe DNS hisztonokra történő felcsvarodása Hiszton-kötött negatív
A citoszol szolubilis fehérjéi. A citoplazma matrix (citoszol) Caspase /Kaszpáz/ 1. Enzimek. - Organellumok nélküli citoplazma
 Caspase /Kaszpáz/ 1. Enzimek. - Organellumok nélküli citoplazma") A citoplazma matrix (citoszol) A citoszol szolubilis fehérjéi 1. Enzimek - Organellumok nélküli citoplazma -A sejt fejlődéstani szempontból legősibb része (a sejthártyával együtt) Glikolízis teljes enzimrendszere
A citoplazma matrix (citoszol) A citoszol szolubilis fehérjéi 1. Enzimek - Organellumok nélküli citoplazma -A sejt fejlődéstani szempontból legősibb része (a sejthártyával együtt) Glikolízis teljes enzimrendszere
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Stressz, gyulladás és a központi idegrendszer
 Stressz, gyulladás és a központi idegrendszer Továbbképzés 2012 november Mandl József Az alábbi ábrák a szintentartó tanfolyamon elhangzott előadás szemléltetői. Ebből következően szöveg nélkül önmagukban
Stressz, gyulladás és a központi idegrendszer Továbbképzés 2012 november Mandl József Az alábbi ábrák a szintentartó tanfolyamon elhangzott előadás szemléltetői. Ebből következően szöveg nélkül önmagukban
TRANSZPORTFOLYAMATOK A SEJTEKBEN
 16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
16 A sejtek felépítése és mûködése TRANSZPORTFOLYAMATOK A SEJTEKBEN 1. Sejtmembrán elektronmikroszkópos felvétele mitokondrium (energiatermelõ és lebontó folyamatok) citoplazma (fehérjeszintézis, anyag
Gyógyszerrezisztenciát okozó fehérjék vizsgálata
 Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
Gyógyszerrezisztenciát okozó fehérjék vizsgálata AKI kíváncsi kémikus kutatótábor 2017.06.25-07.01. Témavezetők : Telbisz Ágnes, Horváth Tamás Kutatók : Dobolyi Zsófia, Bereczki Kristóf, Horváth Ákos Gyógyszerrezisztencia
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Transzporterek vizsgálata lipidmembránokban Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest
 Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
Transzporterek vizsgálata lipidmembránokban 2016. Sarkadi Balázs MTA-SE Molekuláris Biofizikai Kutatócsoport, MTA-TTK Budapest Membrántranszport fehérjék típusok, lipid-kapcsolatok A membránok szerkezete
BIOFIZIKA. Membránpotenciál és transzport. Liliom Károly. MTA TTK Enzimológiai Intézet
 BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
Darvas Zsuzsa László Valéria. Sejtbiológia. Negyedik, átdolgozott kiadás
 Darvas Zsuzsa László Valéria Sejtbiológia Negyedik, átdolgozott kiadás Írták: DR. DARVAS ZSUZSA egyetemi docens Semmelweis Egyetem Genetikai, Sejtés Immunbiológiai Intézet DR. LÁSZLÓ VALÉRIA egyetemi docens
Darvas Zsuzsa László Valéria Sejtbiológia Negyedik, átdolgozott kiadás Írták: DR. DARVAS ZSUZSA egyetemi docens Semmelweis Egyetem Genetikai, Sejtés Immunbiológiai Intézet DR. LÁSZLÓ VALÉRIA egyetemi docens
Energiatermelés a sejtekben, katabolizmus. Az energiaközvetítő molekula: ATP
 Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Energiatermelés a sejtekben, katabolizmus Az energiaközvetítő molekula: ATP Elektrontranszfer, a fontosabb elektronszállító molekulák NAD: nikotinamid adenin-dinukleotid FAD: flavin adenin-dinukleotid
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Immunológia alapjai. 16. előadás. Komplement rendszer
 Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
Immunológia alapjai 16. előadás Komplement rendszer A gyulladás molekuláris mediátorai: Plazma enzim mediátorok: - Kinin rendszer - Véralvadási rendszer Lipid mediátorok Kemoattraktánsok: - Chemokinek:
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK
 POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
POSZTTRANSZLÁCIÓS MÓDOSÍTÁSOK: GLIKOZILÁLÁSOK Dr. Pécs Miklós Budapesti Műszaki és Gazdaságtudományi Egyetem, Alkalmazott Biotechnológia és Élelmiszertudomány Tanszék 1 Glikozilálás A rekombináns fehérjék
A koleszterin és az epesavak bioszintézise
 A koleszterin és az epesavak bioszintézise Koleszterin A koleszterin a biológia legkitüntetettebb kis molekulája. Tizenhárom Nobel-díjat ítéltek oda azon tudósoknak, aki karrierjük legnagyobb részét a
A koleszterin és az epesavak bioszintézise Koleszterin A koleszterin a biológia legkitüntetettebb kis molekulája. Tizenhárom Nobel-díjat ítéltek oda azon tudósoknak, aki karrierjük legnagyobb részét a
Kevéssé fejlett, sejthártya betüremkedésekből. Citoplazmában, cirkuláris DNS, hisztonok nincsenek
 1 A sejtek felépítése Szerkesztette: Vizkievicz András A sejt az élővilág legkisebb, önálló életre képes, minden életjelenséget mutató szerveződési egysége. Minden élőlény sejtes szerveződésű, amelyek
1 A sejtek felépítése Szerkesztette: Vizkievicz András A sejt az élővilág legkisebb, önálló életre képes, minden életjelenséget mutató szerveződési egysége. Minden élőlény sejtes szerveződésű, amelyek
Natív antigének felismerése. B sejt receptorok, immunglobulinok
 Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
Natív antigének felismerése B sejt receptorok, immunglobulinok B és T sejt receptorok A B és T sejt receptorok is az immunglobulin fehérje család tagjai A TCR nem ismeri fel az antigéneket, kizárólag az
3. Főbb Jelutak. 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3.
 Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Jelutak 3. Főbb Jelutak 1. G protein-kapcsolt receptor által közvetített jelutak 2. Enzim-kapcsolt receptorok által közvetített jelutak 3. Egyéb jelutak I. G-protein-kapcsolt receptorok 1. által közvetített
Integráció. Csala Miklós. Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet
 Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Integráció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet Anyagcsere jóllakott állapotban Táplálékkal felvett anyagok sorsa szénhidrátok fehérjék lipidek
Intelligens molekulákkal a rák ellen
 Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
Intelligens molekulákkal a rák ellen Kotschy András Servier Kutatóintézet Rákkutatási kémiai osztály A rákos sejt Miben más Hogyan él túl Áttekintés Rákos sejtek célzott támadása sejtmérgekkel Fehérjék
A nukleinsavak polimer vegyületek. Mint polimerek, monomerekből épülnek fel, melyeket nukleotidoknak nevezünk.
 Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Nukleinsavak Szerkesztette: Vizkievicz András A nukleinsavakat először a sejtek magjából sikerült tiszta állapotban kivonni. Innen a név: nucleus = mag (lat.), a sav a kémhatásukra utal. Azonban nukleinsavak
Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont
 Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont Doktori értekezés Dr. Konta Laura Éva Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola
Fehérjeglikoziláció az endoplazmás retikulumban mint lehetséges daganatellenes támadáspont Doktori értekezés Dr. Konta Laura Éva Semmelweis Egyetem Molekuláris Orvostudományok Tudományági Doktori Iskola
Citrátkör, terminális oxidáció, oxidatív foszforiláció
 Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
Citrátkör, terminális oxidáció, oxidatív foszforiláció A citrátkör jelentősége tápanyagok oxidációjának közös szakasza anyag- és energiaforgalom központja sejtek anyagcseréjében elosztórendszerként működik:
TDK lehetőségek az MTA TTK Enzimológiai Intézetben
 TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
TDK lehetőségek az MTA TTK Enzimológiai Intézetben Vértessy G. Beáta egyetemi tanár TDK mind 1-3 helyezettek OTDK Pro Scientia különdíj 1 második díj Diákjaink Eredményei Zsűri különdíj 2 első díj OTDK
Glukuronidtranszport az endoplazmás retikulumban
 Glukuronidtranszport az endoplazmás retikulumban Doktori értekezés Dr. Révész Katalin Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Dr. Csala Miklós egyetemi docens, Ph.D. Hivatalos
Glukuronidtranszport az endoplazmás retikulumban Doktori értekezés Dr. Révész Katalin Semmelweis Egyetem Molekuláris Orvostudományok Doktori Iskola Témavezető: Dr. Csala Miklós egyetemi docens, Ph.D. Hivatalos
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás.
 Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Immunológia alapjai 5-6. előadás MHC szerkezete és genetikája, és az immunológiai felismerésben játszott szerepe. Antigén bemutatás. Az immunrendszer felépítése Veleszületett immunitás (komplement, antibakteriális
Elválasztástechnikai és bioinformatikai kutatások. Dr. Harangi János DE, TTK, Biokémiai Tanszék
 Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Elválasztástechnikai és bioinformatikai kutatások Dr. Harangi János DE, TTK, Biokémiai Tanszék Fő kutatási területek Enzimek vizsgálata mannozidáz amiláz OGT Analitikai kutatások Élelmiszer analitika Magas
Zsírsav szintézis. Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P. 2 i
 Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
Zsírsav szintézis Az acetil-coa aktivációja: Acetil-CoA + CO + ATP = Malonil-CoA + ADP + P 2 i A zsírsav szintáz reakciói Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H = Palmitát + 8 CoA-SH + 7 CO 2 + 7
A bioenergetika a biokémiai folyamatok során lezajló energiaváltozásokkal foglalkozik.
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA BIOENERGETIKA I. 1. kulcsszó cím: Energia A termodinamika első főtétele kimondja, hogy a különböző energiafajták átalakulhatnak egymásba ez az energia megmaradásának
OZMÓZIS, MEMBRÁNTRANSZPORT
 OZMÓZIS, MEMBRÁNTRANSZPORT Vig Andrea PTE ÁOK Biofizikai Intézet 2014.10.28. ÁTTEKINTÉS DIFFÚZIÓ BROWN-MOZGÁS a részecskék rendezetlen hőmozgása DIFFÚZIÓ a részecskék egyenletlen (inhomogén) eloszlásának
OZMÓZIS, MEMBRÁNTRANSZPORT Vig Andrea PTE ÁOK Biofizikai Intézet 2014.10.28. ÁTTEKINTÉS DIFFÚZIÓ BROWN-MOZGÁS a részecskék rendezetlen hőmozgása DIFFÚZIÓ a részecskék egyenletlen (inhomogén) eloszlásának
LIPID ANYAGCSERE (2011)
") LIPID ANYAGCSERE LIPID ANYAGCSERE (2011) 5 ELİADÁS: 1, ZSÍRK EMÉSZTÉSE, FELSZÍVÓDÁSA + LIPPRTEINEK 2, ZSÍRSAVAK XIDÁCIÓJA 3, ZSÍRSAVAK SZINTÉZISE 4, KETNTESTEK BIKÉMIÁJA, KLESZTERIN ANYAGCSERE 5, MEMBRÁN
LIPID ANYAGCSERE LIPID ANYAGCSERE (2011) 5 ELİADÁS: 1, ZSÍRK EMÉSZTÉSE, FELSZÍVÓDÁSA + LIPPRTEINEK 2, ZSÍRSAVAK XIDÁCIÓJA 3, ZSÍRSAVAK SZINTÉZISE 4, KETNTESTEK BIKÉMIÁJA, KLESZTERIN ANYAGCSERE 5, MEMBRÁN
15. Fehérjeszintézis: transzláció. Fehérje lebontás (proteolízis)
") 15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
15. Fehérjeszintézis: transzláció Fehérje lebontás (proteolízis) 1 Transzláció fordítás A C G T/U A C D E F G H I K L M N P Q R S T V W Y 4 betűs írás (nukleinsavak) 20 betűs írás (fehérjék) 2 Amit már
DER (Felületén riboszómák találhatók) Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.
 Feladata a biológiai fehérjeszintézis Riboszómák. Az endoplazmatikus membránrendszer. A kódszótár.") Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
Az endoplazmatikus membránrendszer Részei: DER /durva (szemcsés) endoplazmatikus retikulum/ SER /sima felszínű endoplazmatikus retikulum/ Golgi készülék Lizoszómák Peroxiszómák Szekréciós granulumok (váladékszemcsék)
Tantárgyi kód BIB 1211 Meghirdetés féléve 2 Kreditpont 3 Összóraszám (elm.+gyak) 3+0. Előfeltétel (tantárgyi kód):
 3+0. Előfeltétel (tantárgyi kód):") Tantárgy neve Sejtbiológia Tantárgyi kód BIB 1211 Meghirdetés féléve 2 Kreditpont 3 Összóraszám (elm.+gyak) 3+0 Számonkérés módja kollokvium Előfeltétel (tantárgyi kód): A tantárgyfelelős neve Kalucza
Tantárgy neve Sejtbiológia Tantárgyi kód BIB 1211 Meghirdetés féléve 2 Kreditpont 3 Összóraszám (elm.+gyak) 3+0 Számonkérés módja kollokvium Előfeltétel (tantárgyi kód): A tantárgyfelelős neve Kalucza
A citoszolikus NADH mitokondriumba jutása
 A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
A citoszolikus NADH mitokondriumba jutása Energiaforrásaink Fototróf: fotoszintetizáló élőlények, szerves vegyületeket állítanak elő napenergia segítségével (a fényenergiát kémiai energiává alakítják át)
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
6. Zárványtestek feldolgozása
 6. Zárványtestek feldolgozása... 1 6.1. A zárványtestek... 1 6.1.1. A zárványtestek kialakulása... 2 6.1.2. A feldolgozási technológia... 3 6.1.2.1. Sejtfeltárás... 3 6.1.2.2. Centrifugálás, tisztítás...
6. Zárványtestek feldolgozása... 1 6.1. A zárványtestek... 1 6.1.1. A zárványtestek kialakulása... 2 6.1.2. A feldolgozási technológia... 3 6.1.2.1. Sejtfeltárás... 3 6.1.2.2. Centrifugálás, tisztítás...
A sejtfelszíni receptorok három fő kategóriája
 A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
A sejtfelszíni receptorok három fő kategóriája 1. Saját enzimaktivitás nélküli receptorok 1a. G proteinhez kapcsolt pl. adrenalin, szerotonin, glukagon, bradikinin receptorok 1b. Tirozin kinázhoz kapcsolt
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Glikolízis. Csala Miklós
 Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
Glikolízis Csala Miklós Szubsztrát szintű (SZF) és oxidatív foszforiláció (OF) katabolizmus Redukált tápanyag-molekulák Szállító ADP + P i ATP ADP + P i ATP SZF SZF Szállító-H 2 Szállító ATP Szállító-H
A fotoszintézis molekuláris biofizikája (Vass Imre, 2000) 39
 39") A fotoszintézis molekuláris biofizikája (Vass Imre, 2000) 39 6. A citokróm b 6 f komplex A két fotokémiai rendszer közötti elektrontranszportot a citokróm b 6 f komplex közvetíti. Funkciója a kétszeresen
A fotoszintézis molekuláris biofizikája (Vass Imre, 2000) 39 6. A citokróm b 6 f komplex A két fotokémiai rendszer közötti elektrontranszportot a citokróm b 6 f komplex közvetíti. Funkciója a kétszeresen
A zsírszövet mellett az agyvelő lipidekben leggazdagabb szervünk. Pontosabban az agy igen gazdag hosszú szénláncú politelítetlen zsírsavakban
 BEVEZETÉS ÉS A KUTATÁS CÉLJA A zsírszövet mellett az agyvelő lipidekben leggazdagabb szervünk. Pontosabban az agy igen gazdag hosszú szénláncú politelítetlen zsírsavakban (LCPUFA), mint az arachidonsav
BEVEZETÉS ÉS A KUTATÁS CÉLJA A zsírszövet mellett az agyvelő lipidekben leggazdagabb szervünk. Pontosabban az agy igen gazdag hosszú szénláncú politelítetlen zsírsavakban (LCPUFA), mint az arachidonsav
13. RNS szintézis és splicing
 13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
13. RNS szintézis és splicing 1 Visszatekintés: Az RNS típusai és szerkezete Hírvivő RNS = mrns (messenger RNA = mrna) : fehérjeszintézis pre-mrns érett mrns (intronok kivágódnak = splicing) Transzfer
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András
 Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Maléth József. Az endoplazmás retikulum - plazma membrán mikrodomének szerepe az intracelluláris Ca 2+ szignalizáció szabályzásában
 Az endoplazmás retikulum - plazma membrán mikrodomének szerepe az intracelluláris Ca 2+ szignalizáció szabályzásában Maléth József Tudományos munkatárs MTA Orvosi Tudományok Osztálya 2017. 04. 19. Az endoplazmás
Az endoplazmás retikulum - plazma membrán mikrodomének szerepe az intracelluláris Ca 2+ szignalizáció szabályzásában Maléth József Tudományos munkatárs MTA Orvosi Tudományok Osztálya 2017. 04. 19. Az endoplazmás
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
Bevezetés a biokémiába fogorvostan hallgatóknak
 Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Bevezetés a biokémiába fogorvostan hallgatóknak Munkafüzet 14. hét METABOLIZMUS III. LIPIDEK, ZSÍRSAVAK β-oxidációja Szerkesztette: Jakus Péter Név: Csoport: Dátum: Labor dolgozat kérdések 1.) ATP mennyiségének
Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására
 Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
Szalma Katalin Biológiai módszerek alkalmazása környezeti hatások okozta terhelések kimutatására Témavezető: Dr. Turai István, OSSKI Budapest, 2010. október 4. Az ionizáló sugárzás sejt kölcsönhatása Antone
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak
 BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
BIOGÉN ELEMEK Azok a kémiai elemek, amelyek az élőlények számára létfontosságúak A több mint száz ismert kémiai elem nagyobbik hányada megtalálható az élőlények testében is, de sokuknak nincsen kimutatható
Biokémia. Alkalmazott Biotechnológia és Élelmiszer-tudományi Tanszék: Ch épület III.
 Biokémia Szarka András szarka@mail.bme.hu 463-3858 Wunderlich Lívius livius@mail.bme.hu 463-1407 Alkalmazott Biotechnológia és Élelmiszer-tudományi Tanszék: Ch épület III. Tantárgyi követelmények A biokémia
Biokémia Szarka András szarka@mail.bme.hu 463-3858 Wunderlich Lívius livius@mail.bme.hu 463-1407 Alkalmazott Biotechnológia és Élelmiszer-tudományi Tanszék: Ch épület III. Tantárgyi követelmények A biokémia