Excitáció-kontrakció csatolása szívizomsejtek. sejtekben. Dr. Tóth András
|
|
|
- Rezső Barna
- 8 évvel ezelőtt
- Látták:
Átírás
1 Excitáció-kontrakció csatolása szívizomsejtek sejtekben Dr. Tóth András

2 2+ i Intracelluláris szabad kálcium koncentráció

3 Témák Az ECc-ben szerepet játszó sejtstruktúrák Myofilamentumok az ECc végsı effektorai Az aktivátor Ca kinetikája Akciós potenciálok és ioncsatornák* A Ca beáramlás csatornái A szarkolemma Ca pumpája és az NCX A szarkoplazmás retikulum mőködése Excitációs kontrakciós csatolás SR Ca felszabadulás és Ca fluxusok Inotrópia Ca mismanagement

4 1 Hasonlóságok a szív- és a vázizom EC csatolásában! Mindkét izomtípus harántcsíkolt (fejlett T-tubulus, illetve intracelluláris SR hálózat) Az akciós potenciál biztosítja az excitációs stimulust, mely aktiválja a plazmamembrán Ca csatornáit Az aktivált Ca csatornák triggerelik a szarkoplazmás retikulum Ca release csatornáit Ennek eredményeként megemelkedik az intracelluláris szabad Ca és aktiválja a kontraktilis apparátust

5 2 Különbségek a szív- és a vázizom EC csatolásában! A szívizomban a T-tubulus, illetve szarkoplazmás retikulum rendszer kevésbé fejlett A szívben specializálódott ingerképzı (SA) szövet és vezetı (Purkinje) rostok találhatók A szívizom sejtek szincitiumot alkotnak azaz gap junction-ok segítségével elektromosan összekapcsoltak A kamrai szívizomsejtek akciós potenciál kb. 100x hosszabb (250 ms) mint a vázizom akciós potenciálja

6 3 Az ECc fontosabb lépései szívizomban! Az AP szomszédos myocitákról gap junction csatoláson keresztül átterjed a szívizomsejtre és végigterjed azon A depolarizáció aktiválja a membrán Ca csatornáit, s az elnyújtott AP alatt jelentıs mértékő Ca influx jön létre Az intracelluláris Ca lokális növekedése nagymértékő Ca felszabadulást triggerel az SR-bıl (CICR) Az intracelluláris Ca globális növekedése aktiválja a myofilamentumokat s így kontrakciót hoz létre β 1 -adrenerg stimuláció a Ca áram, release és reuptake növelésén keresztül növeli a szív kontraktilitását
 Az intracelluláris Ca globális növekedése aktiválja a myofilamentumokat s így kontrakciót hoz létre")
7 Az EC csatolásban szerepet játszó sejtstuktúrák

8 4 ~ 100 x 25 µm Hısünk, a kamrai szívizomsejt

9 5 Vázizom Szívizom Vázizomban az SR sőrő hálózatot alkot, jelentısen kiszélesedik a terminális ciszternáknál, a T-tubulusok átmérıje jóval kisebb, mint szívizomban. Szívizomban az SR hálózat ritkább, junkcionális csatolásokat tartalmaz a külsı szarkolemmával, illetve a T-tubulusokkal, melyek átmérıje sokkal nagyobb, mint vázizomban. A mitochondrium-denzitás is jóval nagyobb. Vázizomrost és szívizomsejt szerkezete!

10 6! A junkcionális SR/ER és a szarkolemma közötti restricted space külön intracelluláris kompartmentnek tekinthetı és kitüntetett szereppel rendelkezik az EC csatolás és a Ca-homeosztázis vonatkozásában. Ebben a térben jóval nagyobbak a Na +, K + és Ca 2+ koncentrációváltozások, mint a többi citoszol kompartmentben. A teret határoló szarkolemma L-típusú Ca csatorna és NCX fehérje denzitása sokkal nagyobb, mint a sejt más régióiban. A restricted space

11 7! Az SR belsejében számos calsequestrin molekula található A nem junkcionális SR felszínen nagyszámú Capumpa egység található A RyR lábak két párhuzamos sort alkotnak A DHP receptorok hasonló, de alternáló szimmetriájú struktúrát alkotnak a T-tubulus membránban A kulcsfehérjék relatív helyzete a vázizom triádban

12 8! Szívizomban a valószínőleg random módon elhelyezkedı DHP receptorok száma sokkal kisebb, mint vázizomban, ahol szabályos struktúrát alkotnak. T-tubulus junkciók szervezıdése váz-, illetve szívizomban

13 Myofilamentumok: az E-C csatolás végsı effektorai

14 9! Szív Troponin-C 1 db Ca kötı hely (reguláció) K d = 500 nm) 2 db Ca-Mg kötı hely (stabilitás) A szívizomsejtek kontraktilis fehérjéi

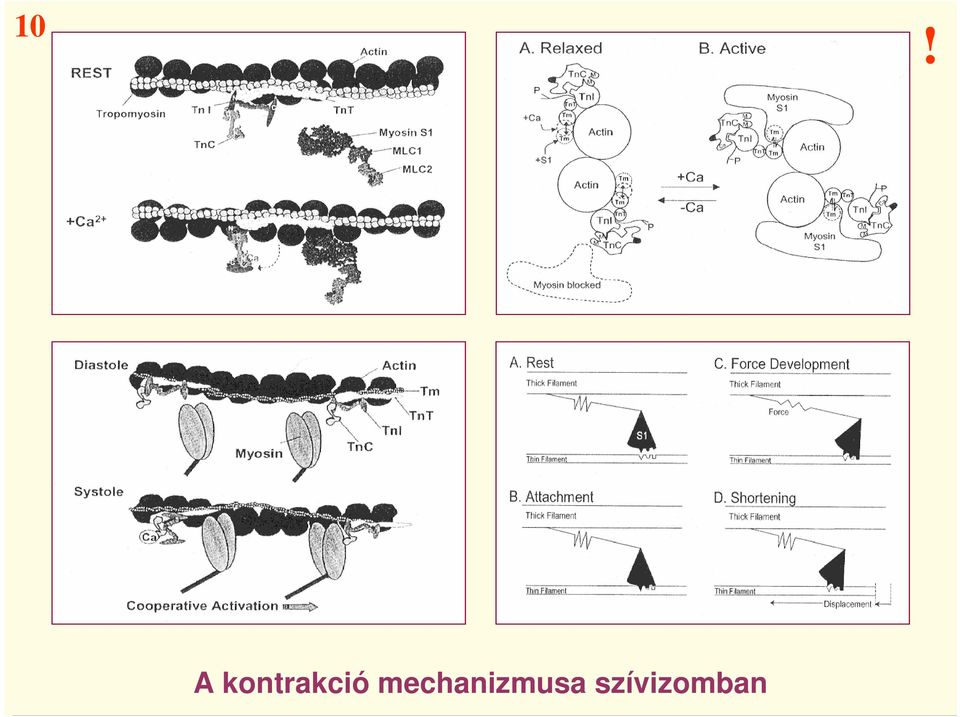
15 10! A kontrakció mechanizmusa szívizomban
16 11 Resting muscle Ca 2+! ADP+P i A + M ADP P i High actin affinity * ADP+P i A-M ADP P i * ATP A-M ATP Low actin affinity P i ATP A-M * Rigor Complex ADP A kereszthíd ciklus fontosabb lépései szívizomban

17 12 A vázizom kontrakciós erejét meghatározó tényezık A) Kontrakció szummáció (tetanusz) B) További rostok aktiválása (recruitment) C) Szarkomer hossz (myofilamentum overlap )! A szívizom kontrakciós erejét meghatározó tényezık A) Intracelluláris Ca koncentráció (analóg) (intrinsic reguláció) B) Szarkomer hossz (myofilamentum overlap ) (extrinsic reguláció) A kontrakciós erı szabályozása váz- és szívizomban
 B) Szarkomer hossz (myofilamentum overlap ) (extrinsic reguláció) A kontrakciós")
18 13! A kontrakció hossz-feszülés görbéje váz- és szívizomban

19 14 Ca szenzitizáló szerek Pozitív inotróp szerek Hipoxia iszkémia A Ca-érzékenységet befolyásoló fontosabb tényezık

20 15! A kontrakciós erı-rövidülési sebesség görbe szívizomban

21 Az aktivátor kalcium forrásai, eltávolítása és kinetikája
22 16!! (From Bers, 2002) Ca transzportfolyamatok kamrai myocitákban
23 17 A) A myofilamentumok Ca érzékenysége (F = F max /(1 + (K m /[Ca] i ) n ) B) A kontrakció aktiválásához szükséges hozzáadott Ca mennyisége A görbék a skinned szívizomrostokra (Hill koefficiens n = 2), illetve intakt szívizomsejtekre (n = 4) vonatkoznak A myofilamentumok aktiválásához szükséges ( aktivátor ) Ca
24 18 Nyúl kamrai myocita modell A [Ca] i pontos értékének kísérletes meghatározása szinte lehetetlen!!!!!!! A) Változások a szabad és az összes intracelluláris Ca koncentrációkban B) Változások a citoplazmatikus Ca-kötı ligandokhoz kötött Ca mennyiségében C) Ca áram és transzporter fluxusok Dinamikus Ca változások (Ca tranziens) a kontrakció során
25 Extracelluláris tér (ECS 30% teljes testtérfogat) [Ca]: Influx: Efflux: Belsı szarkolemma felszín [Ca]: Szarkoplazmás retikulum [Ca]: Influx: Efflux: Mitochondriumok [Ca]: 2 mmol/l ECS x 0.55 L ECS/L citoszol = 1000 µmol/l citoszol VD Ca csatornák, Na/Ca kicserélı, leakage csatornák Na/Ca kicserélı, szarkolemma Ca-ATPáz 60 µmol/l citoszol (nincs szerepe az EC csatolásban!) (ui. gyors extracelluláris Ca eltávolítást követıen a depolarizáció nem hoz létre mérhetı [Ca] növekedést) µmol/l citoszol SR Ca-release csatorna (citoszolba!) Sr Ca ATP-áz (SERCA 2 ) (SR-be!) µmol/l citoszol (in vitro) (PO 4 3- matrix loading ) [Ca]: 100 µmol/l citoszol (in vivo) Influx: Na/Ca antiport (citoszolba!) Efflux: Ca uniport (mitochondriumba!) A citoszol Ca tartalma, Ca influx és efflux mechanizmusok
26 19 Coffein: felszabadítja az SR Ca tartalmát és gátolja visszavételét 0Na, 0Ca: gátolja az NCX-et FCCP: szétkapcsolja a mitochondriumokat Az egyes Ca transzporter mechanizmusok szerepe a relaxációban
27 20 Kritikus [Ca] i 500 nm A mitochondriális Ca ciklus a [Ca 2+ ] m változások tükrözıdnek a dehidrogenázok aktivitásában A mitochondrium szabad Ca tartalma [Ca] m a citoszol [Ca] i függvényében A mitochondrium szerepe az intracelluláris Ca regulációban
28 Akciós potenciálok és ioncsatornák a szívben*
29 21! Ioncsatornák szívizom sejtekben
30 Ca beáramlás a szarkolemma Ca csatornáin keresztül
31 22! (From Bers, 2002) Ca transzportfolyamatok kamrai myocitákban
32 23! A myokardium L - és T-típusú Ca csatornáinak legfontosabb tulajdonságai
33 24 Ca L csatorna: 3-5/µm 2 DHP receptor: 20/µm 2 Az L-típusú Ca csatornák tulajdonságai
34 25 A Ca-dependens I Ca -facilitáció (vagy gátlás) az un. lépcsı (starcase) jelenség oka a nyugalmi és folyamatos aktivitás alatti Ca szintek közötti eltérés. Egyes speciesekben (pl. kutya, nyúl, humán) az összefüggés pozitív, más speciesekben (pl. patkány, egér) negatív starcase jelenség mutatható ki PR: post rest (a nyugalom utáni elsı pulzus) SS: steady state pulzus Ca-dependens Ca-áram facilitáció I Ca positive starcase
35 26 Az AP alakja (idıtartama QT) meghatározza a transzportált Ca mennyiségét Hosszú AP: nagy Ca influx Rövid AP: kevés Ca influx Az I Ca mértéke négyszögimpulzus és AP-clamp alatt
36 27! Antagonisták: Dihidropiridin (DHP)-származékok (nifedipine, nitrendipine, nimodipine, nisoldipine, (+) Bay K 8644, azidopine, iodipine) Fenil-alkilamin (ΦAA)-származékok (Verapamil) Benzothiazepin (BTX)-származékok (Diltiazem) Agonisták: (-) Bay K8644 (+) S , stb. Agonisták: mode 2 ( tartósan nyitott állapot) (pl. Bay K 8644 esetén 0.6ms 20 ms) Antagonisták: mode 0 ( tartósan zárt állapot) A Ca csatorna mőködését moduláló fontosabb ágensek
 Az I Ca kétutas")
37 G s Adenil-cikláz camp PKA(REG) 2. G s direkt hatás (AKAP: PKA rögzítı protein, PLB: foszfolamban) Az I Ca kétutas β-adrenerg aktivációja szívizomsejtekben
38 Összefoglalás! A) Az L-típusú Ca csatornán bejutó áram (I Ca ) a Ca influx legfontosabb mechanizmusa. B) Az I Ca centrális szereppel rendelkezik a szív EC-csatolásában, a Ca regulációban és a kontrakcióban. C) Az I Ca AP alatti kinetikája és amplitúdója kritikus faktorok az SRbıl felszabaduló Ca mennyiségének meghatározásában (is). D) Az I Ca -ban belépı Ca közvetlenül hozzájárul a myofilamentumok aktiválásához, az SR Ca-raktárainak feltöltéséhez/fenntartásához. E) A tartós Ca steady state feltétele, hogy a belépı teljes Ca mennyiség ugyanazon szívciklus alatt (fıleg az NCX mőködése következtében) hagyja el a szívizomsejtet. Ha nem így történik, a sejt fokozatosan telítıdik kalciummal. F) Az L csatornák nagy Ca konduktanciájának következménye, hogy ha (viszonylag kevés) L-csatorna nem inaktiválódik (vagy reaktiválódik), jelentıs intracelluláris [Ca]-szint emelkedés jön létre, (különösen depolarizált sejtben). Ennek következtében a relaxáció gátlódik, illetve arritmia is kialakulhat
39 A Na/Ca kicserélı és a szarkolemma Ca pumpája
40 29! (From Bers, 2002) Szarkolemmális Ca-transzport folyamatok kamrai myocitákban
41 30 A) A PMCA lineáris reprezentációja: 10 TM régió, foszfolipid (PL) szenzitív régió, calmodulin-kötı régió (CaM-BD), stb. B) A PMCA kétdimenziós modellje: autoinhibíciós állapotban (bal), Ca 2+ -CaM stimulációt követıen (jobb) A plazmamembrán Ca 2+ -ATP-áz szerkezeti és mőködési modellje
42 31! A fiziológiás aktivitás alapfeltétele a calmodulin-kötés kialakulása!!! A plazmamembrán Ca 2+ -ATP-áz kinetikus tulajdonságai
 szerkezeti")
43 32 szabályozás XIP: exchange inhibitory protein A Na/Ca kicserélı (NCX) szerkezeti modellje
44 33 A. 2D topológiai modell B. TM 2, 3, 7, 8, illetve α-1 és α-2 loop-ok alkotják a transzport centrumot Az NCX transzport ciklusának lépései szívizomban. A kicserélési aktivitást az intracelluláris Na i & Ca i koncentrációk szabályozzák (E 1 befelé nyitott, E 2 kifelé nyitott) Az elektrogén Na/Ca kicserélı mőködési modellje
45 34! Spike fázis: Plato fázis Repolarizáció (E m > 0) a Ca influx dominál ion-eloszlások függvényében influx, vagy efflux egyaránt lehet az Ca efflux dominál (különösen magas [Ca 2+ ] i esetén) A Na/Ca kicserélı áram (I Na/Ca ) membránpotenciál (E m ) függése
46 35 Az NCX mőködését serkentı és gátló tényezık
47 36! Reverse mode : Ca 2+ influx, Na + efflux AP kezdeti fázisa Forward mode: Ca 2+ efflux, Na + influx AP késıi fázisai Az NCX AP alatti forward, illetve reverse irányú transzportját az ionkoncentrációk mellett a membránpotenciál értéke befolyásolja

48 37 Ilyen egyszerő az NCX áram közelítı kiszámítása
49 38 Szívelégtelenség: I NCX I SERCA Ca transzport-mechanizmusok versengése a szívizom relaxációja alatt
50 39 A nyugalmi csökkenés mértékét elsısorban az SL & SR szivárgási áramainak aránya, illetve a Na/Ca kicserélı aktivitása határozza meg A nyugalmi állapot nem fiziológiás állapota a szívizomsejtnek!!! A csökkenés mértéke erısen species-függı: patkányban igen kicsi, nyúlban viszont jelentıs Az SR Ca tartalmának nyugalmi csökkenése
51 Összefoglalás! A) A Na/Ca kicserélı mechanizmus alapvetı jelentıségő a miokardium intracelluláris Ca-regulációjában B) Ez az elsıdleges mechanizmus, mellyel az L-típusú Ca-csatornákon keresztül bekerülı Ca a relaxáció/diasztolé során kitranszportálódik a sejtekbıl. A szarkolemma Ca-pumpája (SLCP) szívizomban alárendelt jelentıségő. C) A Na/Ca kicserélı rendszer teljesítménye igen nagy, összemérhetı az SR Ca-ATPáz (SERCA 2 ) teljesítményével (~ 1 : 2), s ezáltal számottevıen hozzájárul a relaxációhoz D) Ugyanakkor az NCX elvben képes a kontrakcióhoz szükséges és elégséges Ca-influx létrehozására is. Erre fiziológiás körülmények között nincsen szükség. E) Az NCX által átlagosan kipumpált Ca mennyisége meg kell, hogy egyezzen az L-típusú Ca-csatornán bejutó Ca mennyiségével. F) Mivel az NCX a legfontosabb mechanizmus a Ca efflux kialakításában, jelentıs mértékő gátlása esetén várható az intracelluláris Ca szint progresszív emelkedése
52 Szarkoplazmás retikulum - Ca felvétel, tárolás és felszabadítás
53 40! (From Bers, 2002) Az SR Ca-transzport folyamatai kamrai myocitákban
54 41 Struktúra: 10 transzmembrán hélix. A protein 70%-a a membrán citoszol felıli oldalán van (β-lánc, foszforilációs és nukleotid kötı hely, membránon kívüli domainek, billenı tag) M 4 -M 6 & M 6 : Ca 2+ kötésben és transzportban fontos hélixek. A: Ca 2+ felvétel a citoszolból B: Ca 2+ leadás az SR lumenbe A B A transzport lépései: E 1 : 2 Ca 2+ magas affinitású kötése, ATP kötés, foszforiláció + konf. vált. E 2 -be E 2 : 2 Ca 2+ release az SR-be, két proton transzportjával, konf. vált. E 1 -be Az SR Ca-pumpa (SERCA 2 ) szerkezete és a transzport lépései
55 42 Arány: 2(-3) PLB monomer/serca 2 (nem szaturált) PLB-SERCA2 interakció: heterodimer PLB- SERCA gátolja a Ca transzportot foszforiláció vagy Ca kötés csökkenti a gátlást PLB A foszfolamban szerkezete és hatása az SR Ca transzportjára
56 43 A SR Ca-ATPáz (SERCA 2 ) farmakológiai inhibitorai Thapsigargin (TG) (K d < 2 pm) Ciklopiazonsav (CPA) 2,5-di(tert-butyl)-1,4-benzo-hidrokinon (TBQ)! A SERCA fontosabb (pato)fiziológiás szabályozó faktorai Ca: ph: ATP: Mg: az elérhetı Ca mennyisége ([Ca] i ) alapvetıen meghatározza a pumpa mindenkori aktivitását a pumpa mőködése szempontjából optimális ph 8, a ph jelentıs csökkenése (különösen ph < 7,4, azaz acidózis) esetén csökken a pumpa aktivitása (is), ezáltal a szívizom relaxációja (is) a pumpa 2 ATP-kötı hellyel rendelkezik (szubsztrát + reguláció) általában az ATP mennyisége nem limitáló tényezı, de súlyosiszkémia alatt, az ATP szintézis lassulása miatt csökkenhet a SERCA aktivitása (is), s ezáltal a szívizom relaxációja (is) a SERCA aktuális szubsztrátja valószínőleg Mg-ATP, így a Mg koncentráció jelentıs csökkenése gátolhatja mőködését (is) A SERCA 2 inhibitorai, illetve fontosabb szabályozó faktorai
57 44! SOC: store operated channels triadin, junctin: SR struktúrfehérjék A SR Ca tartalmát befolyásoló tényezık összegzése
58 45 Egyes feltételezések szerint a Ca csatorna kimenete a molekula oldalán lehet, s ezáltal a Ca ionok az SR-bıl közvetlenül a restricted space -be kerülhetnek. MW = Da FKBP: PKA kötı hely CaM: calmodulin kötı hely Az SR Ca release csatornájának (ryanodin receptor, RyR) modellje
59 46 A) 2 db Ca spark (2D konfokális) fluoreszcencia) B) 1 db Ca spark (1D line-scan ) C) Elıbbibıl nyert Ca tranziens D) A fenti sparkot 3D-ben mutató felületi Ca plot Az SR-bıl történı Ca felszabadulás elemi eseménye a lokális spark, mely gyakran spontán, sztohasztikus módon is létrejön RyR vesz benne részt, a T-tubulus közelében indul, 10 ms alatt nmol csúcs [Ca 2+ ] i -t hoz létre. A csökkenés oka Ca diffúzió, illetve Ca reuptake. Ca sparkok izolált kamrai szívizomsejtben Számos lokális spark fúziója Ca-tranziens kialakulásához, ezáltal kontrakcióhoz vezet!!!
60 47 A RyR makromolekuláris signaling komplex számos regulációs mechanizmus végsı integrátora!!! As SR-bıl történı Ca felszabadulást serkentı és gátló tényezık
61 48 Kiriazis 2000 A SR Ca-transzport genetikai modulációinak következményei
62 Összefoglalás! Az SR képes az szívizom-kontrakció aktiválásához elegendı Ca-t tárolni és megfelelıen gyorsan felszabadítani Néhány jellemzı számérték Tipikus kamrai szívizomsejtben ~ 2.5*10 5 DHPR, ~ *10 6 RyR, illetve ~ *10 9 SR Ca-ATPáz molekula található Tipikus nyugalmi Ca spark aktivitás: ~ 50/s, ehhez mindössze ~ 1000/s RyR aktivációja szükséges (az összes RyR ~ 0.02%-a) Csúcs SR Ca release-hez (~ 3 mm/s) ~ /s RyR szükséges (~ 4%) Teljes SR Ca release-hez (~ 50 µmol/l citoszol) ~ 7500 spark szükséges (azaz a RyR-ok ~ 5%-a) Hasonlóan, a mért Ca influx áram (max. 1 na) kialakulásához az L-típusú Ca csatornák (DHP receptorok) ~ 2-3%-a (~ 5000 db) szükséges
63 Excitációs-kontrakciós csatolás (ECc)
64 49! A) Izolált patkány kamrai szívizomsejt B) Béka vázizom rost A vázizommal ellentétben a szívizomban a kontrakció aktiválásához szükséges a külsı Ca beáramlása
65 50 Az SR-bıl történı Ca felszabadulás lehetséges aktivátor mechanizmusai
66 51 Aktiválás Delay idıtartam Gátlás A. Eltérı [Ca]-jú oldatok gyors cseréjét követıen mért tenziók B. Az SR Ca-release-ét indukáló (trigger) [Ca] az adott [Ca] eléréséhez szükséges (delay) idıtartam és a max. kontrakció kapcsolata (pca = - log[ca]) A tenzió (azaz Ca release) függ mind a trigger [Ca]-tól, mind annak változási sebességétıl (rate of change), azaz a mért delay mértékétıl Egy adott Ca-szint alatt pozitív, felette negatív feed-back van, azaz alacsony [Ca] sm aktiválja, magas deaktiválja a CICR mechanizmust. Ezt támasztja alá, hogy a delay idıtartam növekedésével a tenzió csökken! A CICR mechanizmus igazolása, illetve lokális (Ca) kontroll szívizomban (skinned Purkinje rostokon)
67 52! A RyR két Ca-kötıhelye eltérı kinetikával köti a Ca iont, ezáltal Ca-dependens aktiváció és inaktiváció is létre tud jönni: (1: gyors, alacsony affinitású kötıhely gyors aktiváció, 2: lassú, magasabb affinitású kötıhely lassú inaktiváció) Az SR-bıl indukált Ca-release (CICR) javasolt lépései szívizomban
68 53 TT SR TT SR + Vm + + Ca 2+ Ca 2+ Channel Release Channel Excitációs-kontrakciós csatolás szívizomban (Ca 2+ -Induced-Ca 2+ -Release)
69 54 A szívizom EC-csatolásának lokális kontrol elmélete!! Megfigyelések: Hipotézis: Tulajdonságok: Érvényesség: A [Ca] változás mértéke (sebessége) a RyR környezetében aktiválni, illetve inaktiválni tudja a RyR-t A DHPR és a RyR junkcionális kolokalizációja I Na ([Na] sm [Ca] sm ) SR Ca/release Lokalizált SR Ca-release események (Ca sparkok) kimutatása Közös Ca-pool modellek nem magyarázzák a graded CICR-t A RyR-t a fuzzy space azaz a [Ca] sm változásai modulálják Ca-szinapszis hipotézis: 1 DHPR 1 RyR-t triggerel Cluster bomb hipotézis: 1 DHPR cluster RyR-t aktivál Mindkét modell magyarázza a CICR analóg jellegét és nagy erısítését, de utóbbi valószerőbb, mivel nem igényel extra nagy single RyR Ca-fluxust A RyR-ok egy clusterjében (un. couplon) a Ca felszabadulás gyakorlatilag all or none jellegő és regeneratív. Aktív cluster nem képes a többit aktiválni (Ca-spark csak igen ritkán hoz létre újabbat) a graded jelleg az aktív clusterek számának változásából ered. A lokális kontrol elmélet az I Ca indukált SR Ca-release-re vonatkozik. Érvényessége nem igazolt más módokon (NCX, caged Ca) létrehozott SR Carelease vonatkozásában, melyek térbeli eloszlása kevésbé lokalizált a junkcionális régióhoz.
70 55! Vázizomban A DHPR és RyR közötti fizikai kapcsolat alapvetı jelentıségő Nincsen szükség külsı Ca beáramlásra Szívizomban A DHPR és RyR közötti fizikai kapcsolat nem közvetíti a VDCR-t A külsı Ca beáramlása (I Ca ) alapvetı jelentıségő EC-csatolás vázizomban (VDCR), illetve szívizomban (CICR)
71 Összefoglalás Ca A) Lényegét tekintve a 3-féle alapvetı izomtípus modellként szolgálhat az SR-bıl történı Ca-release 3-féle alapvetı mechanizmusára. (VDCR: vázizom; CICR: szívizom; IP 3 ICR: simaizom). Ez szubsztanciális egyszerősítés. Mindhárom izomtípusban mindhárom mechanizmus jelenléte, és mőködése igazolható B) Vázizomban a release indítása szempontjából a VDCR alapvetı, de a CICR szerepe is vélhetıen nagyon fontos a DHPR-ekkel fizikailag nem kapcsolódó (kb. 50%), VDCR-rel nem aktivált RyR-ok aktiválásában (recruitment), illetve IP 3 is létre tud hozni (perinukleáris) felszabadulást (IP3ICR). C) Szívizomban a CICR az alapvetı jelentıségő EC-csatolási mechanizmus. Ugyanakkor az IP 3 ICR release-moduláló szereppel rendelkezhet. Néhány megfigyelés felveti a szarkolemma és az SR közötti funkcionális direkt kapcsolat (így a VDCR) létét. E kapcsolat tényleges súlya nem ismert. D) Simaizomban mind az IP 3 ICR, mind a CICR szerepe bizonyított, sıt az is bizonyított, hogy az IP 3 ICR kölcsönhat egy más típusú, a csatolásban is résztvevı plazmamembrán Ca csatornával (TRP).!
72 Az SR és SL Ca fluxusok szerepe a szív kontrakció szabályozásában
73 56 Koffein, illetve ryanodin elıkezelés hatása a kontrakciós erı nagyságára különbözı szívizom preparátumokban. A kontrakciós erı helyreállása szívizom preparátumokban 30 s nyugalmi fázis után, ryanodin kezelést követıen (alsó vonal), illetve kontroll-ban (felsı vonal). Species különbségek a steady state kontrakcióban, illetve a kontrakciós erı post-rest recovery-jében
74 57 A) A Ca transzport [Ca] i -függése szívsejtekben B) Relatív Ca-fluxusok kamrai preparátumokban C) Integrált Ca-fluxusok relaxáció alatt D) I Ca & SR eredető aktivátor Ca frakciók A Ca fluxusok analízise különbözı species-ekben
75 58 Frekvencia-függı változások a kontrakciós erıben nyúl kamrai szívizomban Erı-frekvencia összefüggés patkány, nyúl, tengeri malac és humán kamrai izomban Erı-frekvencia kapcsolat szívizomban
76 Összefoglalás! A) A [Ca] i szabályozás részleteiben, különbözı preparátumokban és kísérleti feltételek között jelentıs variabilitás áll fenn. E komplexitás lényegében érthetıvé válik kis számú közös kölcsönható rendszer, és néhány, a különbözı szívpreparátumokban eltérı lényeges funkcionális tulajdonság figyelembevételével. B) A Ca-influx speciális körülmények között képes kontrakció kiváltására, de normális esetben felnıtt emlıs szívizomban az aktivátor Ca legfıbb forrása az SR. C) Az SR-bıl felszabaduló Ca egyrészt újra az SR-ben akkumulálódhat, másrészt az NCX eltávolíthatja a sejtbıl. Steady state-ben a szívciklus során létrejövı Ca-influx és Ca-efflux meg kell, hogy egyezzen. D) Az SR Ca tartalma során fokozatosan kiürülhet (NCX), de az I Ca segítségével post-rest aktivitás során gyorsan helyreállhat (5-10 kontrakció alatt). A transz-szarkolemmális Na-gradiens értékétıl függıen nyugalomban az SR kiürülhet, de fel is töltıdhet. E) A szív [Ca] i szabályozására dinamikus, de érzékeny egyensúly jellemzı, és ennek megváltozása inotróp, illetve luzitróp hatással jár.
77 Pozitív inotrópia
78 59! A myokardium kontraktilis állapotát szabályozó mechanizmusok: 1. Szimpatikus idegrendszer, 2. Frank-Starling mechanizmus 3. Erı-frekvencia összefüggés 4. Adrenerg reguláció 5. Vaszkuláris funkció A szív inotróp állapotának fiziológiás szabályozása
79 Hormonreceptorok & ion transzporterek szívizomsejtekben 1.
80 Hormonreceptorok & ion transzporterek szívizomsejtekben 2.
81 60 Fenn: Lenn: β 1 -receptor aktiválása, deszenzitizációja és down-regulációja molekuláris szinten Különbségek a háromféle β-receptor G-protein csatolásában β-adrenerg mechanizmusok (pl. β 2,3 -receptorokon keresztül) gátló (kardioprotektív) hatást is közvetíthetnek (pl. NO-szintézisen keresztül kontraktilitás csökkentés)!!! β-adrenerg receptor szignál-mechanizmusok szívizomban
82 61! Az α-adrenerg reguláció G-protein PLC (& PLD) IP 3 +DAG lépéseken keresztül pozitív inotróp hatású, továbbá hipertrófia kialakulását is serkenti α-adrenerg traszdukciós mechanizmusok kamrai sejtekben
83 62 α-adrenerg aktiváció hatására a Ca-tranziens sokkal kevésbé nı mint az erı, β-adrenerg hatás jelentısen növeli a tranzienst β-adrenerg hatás csökkenti, α- adrenerg hatás viszont növeli a myofilamentumok Caérzékenységét! α- és β-adrenerg inotrópia összehasonlítása
84 63! Emelkedett diasztolés [Ca] i pozitív inotróp hatású
85 64 A Na-pumpa gátlása szívglikoziddal (strofantin) növeli a nyugalmi Na i szintet (A), ezáltal emeli az IC Ca szintjét és az SR Ca szintjét (B). Szívglikozid (acetilstrofantin) koffein vagy ryanodin jelenlétében is növeli a kontrakció erejét Acetil-strofantidin hatásai a Ca-homeosztázisra
86 65 Ca-blokkoló (Nifedipine) hatására szinte teljesen megszőnik a kontrakció, még koffein jelenlétében is (fenn), továbbá az APD is számottevıen rövidül. A Na-pumpa gátlását követıen (acetil-strofantin) a Nifedipine már nem képes az elızı mértékben csökkenteni a kontrakciós erıt, bár az APD csökken (lenn). Ca csatorna gátlás következményei
87 Összefoglalás! A) A szív inotrópiájának növelése több mechanizmussal is lehetséges - hypotermia (kísérleti célok) - β-adrenerg aktiváció (fiziológiás) - α-adrenerg aktiváció (fiziológiás) - CaMKII (Ca-CaM-függı protein kináz) aktiváció (fiziológiás) - kardioaktív szteroidok (szív glikozidok) (terápiás) B) Fiziológiás körülmények között a β 1 -adrenerg aktiváció (ANS) szerepe különlegesen jelentıs (inotróp, chronotróp, luzitróp, stb. hatások), de a β 2,3 -receptorok gátló, kardioprotektív hatásokat is közvetíthetnek (NO). C) Az α-adrenerg aktiváció szerepe az inotrópia növelésében humán szívben kevésbé jelentıs, ugyanakkor fontos szerepe van a szív hipertrófiájának indukciójában (PKC). α-adrenerg aktiváció növeli a myofilamentumok Ca érzékenységét, de nem fokozza a relaxáció sebességét. Az APD-t növeli. D) A CaMKII szerepe viszonylag kevésbé ismert. Mivel autofoszforilációra képes, szerepe lehet [Ca] i szignálok integrációjában. E) A digitalis a szív patológiás körülmények között csökkent inotrópiájának terápiás növelésére legrégebben (1785) használt anyag. A Na-K pumpa gátlásán keresztül növeli a Ca influxot és csökkenti az effluxot, ezáltal az [Ca] i is emelkedik, ami intrinsic módon inotrópia növelı hatású. Túlzott használata azonban negatív inotrópiához és arritmogenezishez vezet.
88 Ca mismanagement és negatív inotrópia
89 66! Az EAD-ok az I Ca újraaktiválódása következtében alakulnak ki, különösen az APD jelentıs megnyúlása esetén (pl. LQT szindróma) A DAD-ok oka vélhetıen az SR túltöltıdése következtében spontán kialakuló sparkok aktiváló hatása a Ca-függı csatornákra (I NA/Ca, I Cl(Ca), I NS(Ca) ) Spontán Ca felszabadulás és utópotenciálok szívizomsejtekben
90 67 C Acidózis hatása a szív EC-csatolására D Papilláris izom preparátum-ban ph o változtatása után az erı csökkent, a Ca-tranziens nıtt (A+B). A ph-csökkenés hatására lényegesen csökken a maximális kontrakciós erı (C). Az intracelluláris ph i szabályozásának legfontosabb transzport mechanizmusai (D).
91 68 Akut iszkémia-reperfúzió alatt kialakuló fontosabb változások
92 69 Hipertrófiához vezetı szignál kaszkádok
93 70! Szívelégtelenségben módosult funkcióval rendelkezı struktúrák : NCX (reverz és forward módban is) Feszültség szenzor Foszfolamban SR-Ca-ATPáz (SERCA2a) A szarkolemma Ca-transzportjának (NCX) relatív súlya nı A szarkoplazmás retikulum Ca-transzportjának (SERCA) relatív jelentısége csökken. Szívelégtelenségben az EC-csatolásban kialakuló fontosabb elváltozások
94 70!!! + feedback A csökkent pumpa funkciót (fiziológiás) negatív feedback folyamatok próbálják kompenzálni. Ezek következtében az intracelluláris Ca-szint és a Ca tranziens emelkedik.!!!! - feedback Az intracelluláris Ca-szint emelkedése maladaptív gén-expressziót indukál, mely tovább rontja a pumpa funkciót, és kóros pozitív feed-back kör, circulus viciosus jön létre. A genetikai változások pozitív feed-back-je
95 72 A Ca-homeosztázisban szívelégtelenségben kialakuló változások
96 73 Kontraktilis diszfunkció és arritmogenezis szívelégtelenségben
97 74 Változások a Ca homeosztázissal kapcsolatos gének expressziójában súlyos fokú szívelégtelenségben
98 Összefoglalás! A) A szív (kamrai szívizomsejt) nagyon finoman hangolt rendszer, amely képes a kontrakció paramétereit rövid idı alatt a fiziológiás igények rendkívül széles tartományban történı változásaihoz igazítani az ion-áramok, Ca-homeosztázis és a myofilamentumok paramétereinek megváltoztatásával. Ez a rendszer sok ponton redundáns, de amennyiben a normális [Ca]-regulációs mechanizmusok jelentısen sérülnek, súlyos negatív inotrópia jöhet létre. B) Ca túlterhelés gyakran spontán SR Ca-release-hez, és Ca-hullámok kialakulásához vezet, melyek, ha nagyszámú sejtben, random módon generálódnak, a kontrakciós erı jelentıs csökkenéséhez vezetnek. C) A késıi utópotenciálok és utókontrakciók kialakulásában az SR Ca-release által indukált I ti (tranziens inward áram: I Na/Ca + I Cl(Ca) + I NS(Ca) ) vesz részt. D) Az acidózis, mely gyakran myokardiális iszkémia következménye, a Catranszport több komponensének (NCX, SERCA, stb.) gátlása révén jelentıs kardiodepresszív hatással rendelkezik (kontrakciós erı & Ca-érzékenység). E) A hipoxia, iszkémia/reperfúzió és szívelégtelenség rendkívül komplex, multifaktoriális patomechanizmusainak progressziójában mindig jelentıs szerepet játszanak az intracelluláris Ca-homeosztázis folyamatosan súlyosbodó zavarai, melyek egyik legfontosabb következménye a szív kontrakciós erejének sokszor a keringés összeomlásához vezetı mértékő csökkenése.
Vázizom elektrofiziológia alapjai. Tóth András, PhD
 Vázizom elektrofiziológia alapjai Tóth András, PhD Témák Struktúra Kontrakció és relaxáció Aktiváció Excitáció-kontrakció csatolás Akciós potenciál Ioncsatornák* Ca 2+ homeosztázis Struktúra Vázizom
Vázizom elektrofiziológia alapjai Tóth András, PhD Témák Struktúra Kontrakció és relaxáció Aktiváció Excitáció-kontrakció csatolás Akciós potenciál Ioncsatornák* Ca 2+ homeosztázis Struktúra Vázizom
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
Excitáció-kontrakció csatolása szívizomsejtekben
 Excitáció-kontrakció csatolása szívizomsejtekben A háztartás feladata ARTÉRIÁS VÉRNYOMÁS + + PERCTÉRFOGAT + + Teljes perifériás rezisztencia Szívfrekvencia VERŐTÉRFOGAT + KONTRAKCIÓ Intracelluláris ciklus
Excitáció-kontrakció csatolása szívizomsejtekben A háztartás feladata ARTÉRIÁS VÉRNYOMÁS + + PERCTÉRFOGAT + + Teljes perifériás rezisztencia Szívfrekvencia VERŐTÉRFOGAT + KONTRAKCIÓ Intracelluláris ciklus
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András
 Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Szívelektrofiziológiai alapjelenségek. Dr. Tóth András 2018
 Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
A szívizomsejt ioncsatornái és azok működése
 A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
IONCSATORNÁK. Osztályozás töltéshordozók szerint: pozitív töltésű ion: Na+, K+, Ca2+ negatív töltésű ion: Cl-, HCO3-
 Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
A harántcsíkolt izom struktúrája általános felépítés
 harántcsíkolt izom struktúrája általános felépítés LC-2 Izom LC1/3 Izom fasciculus LMM S-2 S-1 HMM rod Miozin molekula S-1 LMM HMM S-2 S-1 Izomrost H Band Z Disc csík I csík M Z-Szarkomér-Z Miofibrillum
harántcsíkolt izom struktúrája általános felépítés LC-2 Izom LC1/3 Izom fasciculus LMM S-2 S-1 HMM rod Miozin molekula S-1 LMM HMM S-2 S-1 Izomrost H Band Z Disc csík I csík M Z-Szarkomér-Z Miofibrillum
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
ÖSSZ-TARTALOM. 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3.
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés 2. A sejtkommunikáció
ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás
 Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ-TARTALOM 1. Az alapok - 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi- és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
A szív élettana. Aszív élettana I. A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG
 A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa
A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa
Jelutak ÖSSZ TARTALOM. Jelutak. 1. a sejtkommunikáció alapjai
 Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
Jelutak ÖSSZ TARTALOM 1. Az alapok 1. előadás 2. A jelutak komponensei 1. előadás 3. Főbb jelutak 2. előadás 4. Idegi és hormonális kommunikáció 3. előadás Jelutak 1. a sejtkommunikáció alapjai 1. Bevezetés
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák
 A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
A miokardium intracelluláris kalcium homeosztázisa: iszkémiás és kardiomiopátiás változások
 Doktori értekezés A miokardium intracelluláris kalcium homeosztázisa: iszkémiás és kardiomiopátiás változások Dr. Szenczi Orsolya Témavezető: Dr. Ligeti László Klinikai Kísérleti Kutató- és Humán Élettani
Doktori értekezés A miokardium intracelluláris kalcium homeosztázisa: iszkémiás és kardiomiopátiás változások Dr. Szenczi Orsolya Témavezető: Dr. Ligeti László Klinikai Kísérleti Kutató- és Humán Élettani
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az akciós potenciál (AP) 2.rész. Szentandrássy Norbert
 2.rész. Szentandrássy Norbert") Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Új terápiás lehetőségek (receptorok) a kardiológiában
 a kardiológiában") Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
Új terápiás lehetőségek (receptorok) a kardiológiában Édes István Kardiológiai Intézet, Debreceni Egyetem Kardiomiociták Ca 2+ anyagcseréje és új terápiás receptorok 2. 1. 3. 6. 6. 7. 4. 5. 8. 9. Ca
Érzékszervi receptorok
 Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Kollár Veronika
 A harántcsíkolt izom szerkezete, az izommőködés és szabályozás molekuláris alapjai Kollár Veronika 2010. 11. 11. Az izom citoszkeletális filamentumok és motorfehérjék rendezett összeszervezıdésébıl álló
A harántcsíkolt izom szerkezete, az izommőködés és szabályozás molekuláris alapjai Kollár Veronika 2010. 11. 11. Az izom citoszkeletális filamentumok és motorfehérjék rendezett összeszervezıdésébıl álló
Elektrofiziológiai alapjelenségek. Dr. Tóth András
 Elektrofiziológiai alapjelenségek Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Lokális és akciós potenciálok Az ingerület terjedése
Elektrofiziológiai alapjelenségek Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Lokális és akciós potenciálok Az ingerület terjedése
Membránpotenciál, akciós potenciál
 A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
-Két fő korlát: - asztrogliák rendkívüli morfológiája -Ca szignálok értelmezési nehézségei
 Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika
 Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
4. Egy szarkomer sematikus rajza látható az alanti ábrán. Aktív kontrakció esetén mely távolságok csökkenése lesz észlelhető? (3)
") Budapesti Műszaki és Gazdaságtudományi Egyetem, Budapest, 2009. jan. 6. Villamosmérnöki és Informatikai Kar Semmelweis Egyetem Budapest Egészségügyi Mérnök Mesterképzés Felvételi kérdések orvosi élettanból
Budapesti Műszaki és Gazdaságtudományi Egyetem, Budapest, 2009. jan. 6. Villamosmérnöki és Informatikai Kar Semmelweis Egyetem Budapest Egészségügyi Mérnök Mesterképzés Felvételi kérdések orvosi élettanból
Transzportfolyamatok a biológiai rendszerekben
 A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
Receptorok, szignáltranszdukció jelátviteli mechanizmusok
 Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV
 KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
A szív ingerképző és vezető rendszere
 A szív ingerképző és vezető rendszere A ritmikus működés miogén eredetű Az elektromos aktivitás alakja az elvezetés helyétől függ: 1. Nodális szövetről (SA és AV csomó) Pacemaker potenciál 2. Munkaizomzatról,
A szív ingerképző és vezető rendszere A ritmikus működés miogén eredetű Az elektromos aktivitás alakja az elvezetés helyétől függ: 1. Nodális szövetről (SA és AV csomó) Pacemaker potenciál 2. Munkaizomzatról,
Vázizom Simaizom. Szentesi Péter
 Vázizom Simaizom Szentesi Péter A harántcsíkolt izom struktúrája általános felépítés LC-2 Izom LC1/3 Izom fasciculus LMM S-2 S-1 HMM rod Miozin molekula S-1 LMM HMM S-2 S-1 Izomrost H Band Z Disc A csík
Vázizom Simaizom Szentesi Péter A harántcsíkolt izom struktúrája általános felépítés LC-2 Izom LC1/3 Izom fasciculus LMM S-2 S-1 HMM rod Miozin molekula S-1 LMM HMM S-2 S-1 Izomrost H Band Z Disc A csík
Intracelluláris ion homeosztázis I.-II. Február 15, 2011
 Intracelluláris ion homeosztázis I.II. Február 15, 2011 Ca 2 csatorna 1 Ca 2 1 Ca 2 EC ~2 mm PLAZMA Na /Ca 2 cserélő Ca 2 ATPáz MEMBRÁN Ca 2 3 Na ATP ADP 2 H IC ~100 nm citoszol kötött Ca 2 CR CSQ SERCA
Intracelluláris ion homeosztázis I.II. Február 15, 2011 Ca 2 csatorna 1 Ca 2 1 Ca 2 EC ~2 mm PLAZMA Na /Ca 2 cserélő Ca 2 ATPáz MEMBRÁN Ca 2 3 Na ATP ADP 2 H IC ~100 nm citoszol kötött Ca 2 CR CSQ SERCA
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Membránpotenciál. Nyugalmi membránpotenciál. Akciós potenciál
 Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Energia források a vázizomban
 Energia források a vázizomban útvonal sebesség mennyiség ATP/glükóz 1. direkt foszforiláció igen gyors igen limitált - 2. glikolízis gyors limitált 2-3 3. oxidatív foszforiláció lassú nem limitált 36 Izomtípusok
Energia források a vázizomban útvonal sebesség mennyiség ATP/glükóz 1. direkt foszforiláció igen gyors igen limitált - 2. glikolízis gyors limitált 2-3 3. oxidatív foszforiláció lassú nem limitált 36 Izomtípusok
Biofizika I 2013-2014 2014.12.02.
 ÁTTEKINTÉS AZ IZOM TÍPUSAI: SZERKEZET és FUNKCIÓ A HARÁNTCSÍKOLT IZOM SZERKEZETE MŰKÖDÉSÉNEK MOLEKULÁRIS MECHANIZMUSA IZOM MECHANIKA Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet
ÁTTEKINTÉS AZ IZOM TÍPUSAI: SZERKEZET és FUNKCIÓ A HARÁNTCSÍKOLT IZOM SZERKEZETE MŰKÖDÉSÉNEK MOLEKULÁRIS MECHANIZMUSA IZOM MECHANIKA Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus
 Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Izomműködés. Az izommozgás. az állati élet legszembetűnőbb külső jele a mozgás amőboid, ostoros ill. csillós és izomösszehúzódással
 Izomműködés Az izommozgás az állati élet legszembetűnőbb külső jele a mozgás amőboid, ostoros ill. csillós és izomösszehúzódással történő mozgás van Galenus id. II.szd. - az idegekből animal spirit folyik
Izomműködés Az izommozgás az állati élet legszembetűnőbb külső jele a mozgás amőboid, ostoros ill. csillós és izomösszehúzódással történő mozgás van Galenus id. II.szd. - az idegekből animal spirit folyik
Zárójelentés. A) A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.
 A cervix nyújthatóságának (rezisztencia) állatkísérletes meghatározása terhes és nem terhes patkányban.") Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
Zárójelentés A kutatás fő célkitűzése a β 2 agonisták és altípus szelektív α 1 antagonisták hatásának vizsgálata a terhesség során a patkány cervix érésére összehasonlítva a corpusra gyakorolt hatásokkal.
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ IZOMMŰKÖDÉS 1. kulcsszó cím: A SZERVEZETBEN ELŐFORDULÓ IZOM- SZÖVETEK TÍPUSAI 1. képernyő cím: Sima izomszövet
 Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ IZOMMŰKÖDÉS 1. kulcsszó cím: A SZERVEZETBEN ELŐFORDULÓ IZOM- SZÖVETEK TÍPUSAI 1. képernyő cím: Sima izomszövet G001 akaratunktól függetlenül működik; lassú,
Modul cím: MEDICINÁLIS ALAPISMERETEK BIOKÉMIA AZ IZOMMŰKÖDÉS 1. kulcsszó cím: A SZERVEZETBEN ELŐFORDULÓ IZOM- SZÖVETEK TÍPUSAI 1. képernyő cím: Sima izomszövet G001 akaratunktól függetlenül működik; lassú,
Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén keresztül
 Dr. Miklós Zsuzsanna Semmelweis Egyetem, ÁOK Klinikai Kísérleti Kutató- és Humán Élettani Intézet Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén
Dr. Miklós Zsuzsanna Semmelweis Egyetem, ÁOK Klinikai Kísérleti Kutató- és Humán Élettani Intézet Szívbetegségek hátterében álló folyamatok megismerése a ciklusosan változó szívélettani paraméterek elemzésén
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.
. Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.") Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
1. Mi jellemző a connexin fehérjékre?
 Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon
 Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon Válasz Tóth Attila, PhD, MTA doktora opponensi véleményére Először szeretnék
Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon Válasz Tóth Attila, PhD, MTA doktora opponensi véleményére Először szeretnék
Az izomszövet biokémiája. Izombetegségek. Szerkesztette: Fekete Veronika
 Az izomszövet biokémiája. Izombetegségek Szerkesztette: Fekete Veronika Az izomtípusok jellemzői Simaizom lassú nincs rajzolat Szívizom gyors harántcsíkolt Harántcsíkolt izom belső szervek akaratlan nem
Az izomszövet biokémiája. Izombetegségek Szerkesztette: Fekete Veronika Az izomtípusok jellemzői Simaizom lassú nincs rajzolat Szívizom gyors harántcsíkolt Harántcsíkolt izom belső szervek akaratlan nem
Jelutak. 2. A jelutak komponensei Egy tipikus jelösvény sémája. 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája Receptor fehérje Jel molekula (ligand; elsődleges
Membránszerkezet Nyugalmi membránpotenciál
 Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Biofizika I 2013-2014 2014.12.03.
 Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet A KERESZTHÍD CIKLUSHOZ KAPCSOLÓDÓ ERŐKIEJTÉS egy kereszthíd ciklus során a miozin II fej elmozdulása: í ~10 nm 10 10 egy kereszthíd
Biofizika I. -2014. 12. 02. 03. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet A KERESZTHÍD CIKLUSHOZ KAPCSOLÓDÓ ERŐKIEJTÉS egy kereszthíd ciklus során a miozin II fej elmozdulása: í ~10 nm 10 10 egy kereszthíd
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
ZÁRÓJELENTÉS SZAKMAI BESZÁMOLÓ
 ZÁRÓJELENTÉS SZAKMAI BESZÁMOLÓ Pályázat címe: A szívritmuszavarok és a myocardiális repolarizáció mechanizmusainak vizsgálata; antiaritmiás és proaritmiás gyógyszerhatások elemzése (NI 61902) Vezetı kutató:
ZÁRÓJELENTÉS SZAKMAI BESZÁMOLÓ Pályázat címe: A szívritmuszavarok és a myocardiális repolarizáció mechanizmusainak vizsgálata; antiaritmiás és proaritmiás gyógyszerhatások elemzése (NI 61902) Vezetı kutató:
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
A szelektív NCX gátlás kardioprotektív hatásának vizsgálata szívizomsejteken
 A szelektív NCX gátlás kardioprotektív hatásának vizsgálata szívizomsejteken PhD Tézis Kormos Anita, MSc Szegedi Tudományegyetem Általános Orvostudományi Kar Farmakológiai és Farmakoterápiai Intézet Szeged
A szelektív NCX gátlás kardioprotektív hatásának vizsgálata szívizomsejteken PhD Tézis Kormos Anita, MSc Szegedi Tudományegyetem Általános Orvostudományi Kar Farmakológiai és Farmakoterápiai Intézet Szeged
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
Sejtek membránpotenciálja
 Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Izomélettan. Vázizom
 Izomélettan VÁZIZOM, SIMAIZOM ÉS SZÍVIZOM 2 0 1 6. 0 9. 2 7. Ö S S Z E V O N T S Z E M I N Á R I U M Vázizom Harántcsíkolt Funkcionális egysége az izomrost: sok magvú, hosszú hengeres sejtek Izomrostok
Izomélettan VÁZIZOM, SIMAIZOM ÉS SZÍVIZOM 2 0 1 6. 0 9. 2 7. Ö S S Z E V O N T S Z E M I N Á R I U M Vázizom Harántcsíkolt Funkcionális egysége az izomrost: sok magvú, hosszú hengeres sejtek Izomrostok
Vérkeringés. A szív munkája
 Vérkeringés. A szív munkája 2014.11.04. Keringési Rendszer Szív + erek (artériák, kapillárisok, vénák) alkotta zárt rendszer. Funkció: vér pumpálása vér áramlása az erekben oxigén és tápanyag szállítása
Vérkeringés. A szív munkája 2014.11.04. Keringési Rendszer Szív + erek (artériák, kapillárisok, vénák) alkotta zárt rendszer. Funkció: vér pumpálása vér áramlása az erekben oxigén és tápanyag szállítása
Az izommőködéssel járó élettani jelenségek
 Az izommőködéssel járó élettani jelenségek Az izomszövet az egyetlen olyan szövet, amely hosszúságát változtatni tudja. Egy nem elhízott (non-obese) hölgyben az izomtömeg a testsúly 25-35 %-a, férfiben
Az izommőködéssel járó élettani jelenségek Az izomszövet az egyetlen olyan szövet, amely hosszúságát változtatni tudja. Egy nem elhízott (non-obese) hölgyben az izomtömeg a testsúly 25-35 %-a, férfiben
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt
 az információ kémiailag kódolt") Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
Keringési Rendszer. Vérkeringés. A szív munkája. Számok a szívről. A szívizom. Kis- és nagyvérkör. Nyomás terület sebesség
 Keringési Rendszer Vérkeringés. A szív munkája 2010.11.03. Szív + erek (artériák, kapillárisok, vénák) alkotta zárt rendszer. Funkció: Oxigén és tápanyag szállítása a szöveteknek. Metabolikus termékek
Keringési Rendszer Vérkeringés. A szív munkája 2010.11.03. Szív + erek (artériák, kapillárisok, vénák) alkotta zárt rendszer. Funkció: Oxigén és tápanyag szállítása a szöveteknek. Metabolikus termékek
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A szívizomsejtek ionáramai
 A szívizomsejtek ionáramai Dr. Szentesi Péter DE OEC Élettani Intézet 2009 A szivet alkotó szívizomsejtek A sejtmembrán szerkezete Csatornák Pórus Szőrı Kapu A Patch Clamp módszer Egyedi csatorna izolálása
A szívizomsejtek ionáramai Dr. Szentesi Péter DE OEC Élettani Intézet 2009 A szivet alkotó szívizomsejtek A sejtmembrán szerkezete Csatornák Pórus Szőrı Kapu A Patch Clamp módszer Egyedi csatorna izolálása
BIOMECHANIKA 3 Erőhatások eredete és következményei biológiai rendszerekben
 BIOMECHANIKA 3 Erőhatások eredete és következményei biológiai rendszerekben A MOZGÁS MOLEKULÁRIS MECHANIZMUSAI SZERVEZET SZINTŰ MOZGÁS AZ IZOMMŰKÖDÉS MOLEKULÁRIS MECHANIZMUSAI DR. BUGYI BEÁTA- BIOFIZIKA
BIOMECHANIKA 3 Erőhatások eredete és következményei biológiai rendszerekben A MOZGÁS MOLEKULÁRIS MECHANIZMUSAI SZERVEZET SZINTŰ MOZGÁS AZ IZOMMŰKÖDÉS MOLEKULÁRIS MECHANIZMUSAI DR. BUGYI BEÁTA- BIOFIZIKA
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYEK KÁLIUM TÁPLÁLKOZÁSÁNAK GENETIKAI ALAPJAI előadás áttekintése A kálium szerepe a növényi szervek felépítésében
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYEK KÁLIUM TÁPLÁLKOZÁSÁNAK GENETIKAI ALAPJAI előadás áttekintése A kálium szerepe a növényi szervek felépítésében
FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL
 Eke András, Kollai Márk FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL Szerkesztette: Ivanics Tamás Semmelweis Kiadó www.semmelweiskiado.hu B u d a p e s t, 2 0 0 7 Szerkesztette: Ivanics Tamás egyetemi docens, Semmelweis
Eke András, Kollai Márk FEJEZETEK AZ ÉLETTAN TANTÁRGYBÓL Szerkesztette: Ivanics Tamás Semmelweis Kiadó www.semmelweiskiado.hu B u d a p e s t, 2 0 0 7 Szerkesztette: Ivanics Tamás egyetemi docens, Semmelweis
A veseműködés élettana, a kiválasztás funkciója, az emberi test víztereinek élettana (5)
") A veseműködés élettana, a kiválasztás funkciója, az emberi test víztereinek élettana (5) Dr. Attila Nagy 2016 Kalcium és foszfátháztartás (Tanulási támpont: 63) A szabályozásban a pajzsmirigy, mellékpajzsmirigy
A veseműködés élettana, a kiválasztás funkciója, az emberi test víztereinek élettana (5) Dr. Attila Nagy 2016 Kalcium és foszfátháztartás (Tanulási támpont: 63) A szabályozásban a pajzsmirigy, mellékpajzsmirigy
ZSÍRSAVAK OXIDÁCIÓJA. FRANZ KNOOP német biokémikus írta le először a mechanizmusát. R C ~S KoA. a, R-COOH + ATP + KoA R C ~S KoA + AMP + PP i
 máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
máj, vese, szív, vázizom ZSÍRSAVAK XIDÁCIÓJA FRANZ KNP német biokémikus írta le először a mechanizmusát 1 lépés: a zsírsavak aktivációja ( a sejt citoplazmájában, rövid zsírsavak < C12 nem aktiválódnak)
A légzési lánc és az oxidatív foszforiláció
 A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
A légzési lánc és az oxidatív foszforiláció Csala Miklós Semmelweis Egyetem Orvosi Vegytani, Molekuláris Biológiai és Patobiokémiai Intézet intermembrán tér Fe-S FMN NADH mátrix I. komplex: NADH-KoQ reduktáz
Hemoglobin - myoglobin. Konzultációs e-tananyag Szikla Károly
 Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Hemoglobin - myoglobin Konzultációs e-tananyag Szikla Károly Myoglobin A váz- és szívizom oxigén tároló fehérjéje Mt.: 17.800 153 aminosavból épül fel A lánc kb 75 % a hélix 8 db hélix, köztük nem helikális
Adatelemzési eljárások az idegrendszer kutatásban Somogyvári Zoltán
 Adatelemzési eljárások az idegrendszer kutatásban Somogyvári Zoltán MTA KFKI Részecske és Magfizikai Intézet, Biofizikai osztály Az egy adatsorra (idősorra) is alkalmazható módszerek Példa: Az epileptikus
Adatelemzési eljárások az idegrendszer kutatásban Somogyvári Zoltán MTA KFKI Részecske és Magfizikai Intézet, Biofizikai osztály Az egy adatsorra (idősorra) is alkalmazható módszerek Példa: Az epileptikus
Bevezetés I. Diabéteszes kardiomiopátia (10, 16)
") Bevezetés A szívizom elégtelenség epidémiás jelleget öltött, a daganatos megbetegedések mellett a halálozások fő kórokának tekinthető. Az intracelluláris kalcium (Ca 2+ i) homeosztázis alapvetően szabja
Bevezetés A szívizom elégtelenség epidémiás jelleget öltött, a daganatos megbetegedések mellett a halálozások fő kórokának tekinthető. Az intracelluláris kalcium (Ca 2+ i) homeosztázis alapvetően szabja
Szívmőködés. Dr. Cseri Julianna
 Szívmőködés Dr. Cseri Julianna A keringési szervrendszer funkcionális szervezıdése Szív Vérerek Nagyvérkör Kisvérkör Nyirokerek A szív feladata: a vérkeringés fenntartása A szív szívó-nyomó pumpa Automáciával
Szívmőködés Dr. Cseri Julianna A keringési szervrendszer funkcionális szervezıdése Szív Vérerek Nagyvérkör Kisvérkör Nyirokerek A szív feladata: a vérkeringés fenntartása A szív szívó-nyomó pumpa Automáciával
EGYETEMI DOKTORI (Ph.D.) ÉRTEKEZÉS TÉZISEI
 ÉRTEKEZÉS TÉZISEI") EGYETEMI DOKTORI (Ph.D.) ÉRTEKEZÉS TÉZISEI A SEA0400 hatása az emlıs kamrai szívizom kalcium homeosztázisára Birinyi Péter Témavezetık: Dr. Nánási Péter és Dr. Magyar János DEBRECENI EGYETEM ORVOS- ÉS
EGYETEMI DOKTORI (Ph.D.) ÉRTEKEZÉS TÉZISEI A SEA0400 hatása az emlıs kamrai szívizom kalcium homeosztázisára Birinyi Péter Témavezetık: Dr. Nánási Péter és Dr. Magyar János DEBRECENI EGYETEM ORVOS- ÉS
Az ingerületi folyamat sejtélettani alapjai
 Az ingerületi folyamat sejtélettani alapjai Dr. Oláh Attila DEOEC Élettani Intézet 2011.09.15. Alapvetések I. Mi az a membránpotenciál? Az intakt sejtmembrán elektromosan szigetel -> a rajta keresztül
Az ingerületi folyamat sejtélettani alapjai Dr. Oláh Attila DEOEC Élettani Intézet 2011.09.15. Alapvetések I. Mi az a membránpotenciál? Az intakt sejtmembrán elektromosan szigetel -> a rajta keresztül
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Az emlıs keringési rendszer felépítése
 Az emlıs keringési rendszer felépítése tüdı artériák kis vérkör tüdı vénák zárt keringés: magas nyomás, gyors áramlás, gyors szabályozás (diffúzió nem lenne elég) szív nyirokkeringés nyirokcsomó aorta
Az emlıs keringési rendszer felépítése tüdı artériák kis vérkör tüdı vénák zárt keringés: magas nyomás, gyors áramlás, gyors szabályozás (diffúzió nem lenne elég) szív nyirokkeringés nyirokcsomó aorta
SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár
 SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár TDK felelős: Dr. Pataricza János egyetemi docens tel.: (62) 545-674 fax: (62) 545-680 E-mail: Pataricza@phcol.szote.u
SZTE Farmakológiai és Farmakoterápiai Intézet Igazgató: Dr. Varró András egyetemi tanár TDK felelős: Dr. Pataricza János egyetemi docens tel.: (62) 545-674 fax: (62) 545-680 E-mail: Pataricza@phcol.szote.u
INTRACELLULÁRIS Ca 2+ HOMEOSZTÁZIS-
 MTA-Doktori Értekezés Tézisei INTRACELLULÁRIS Ca 2+ HOMEOSZTÁZIS- VÁLTOZÁSOK HATÁSAINAK ELEMZÉSE IZOLÁLT SZÍVPREPARÁTUMOKON Tóth András Szegedi Tudományegyetem ÁOK Farmakológiai és Farmakoterápiai Intézet
MTA-Doktori Értekezés Tézisei INTRACELLULÁRIS Ca 2+ HOMEOSZTÁZIS- VÁLTOZÁSOK HATÁSAINAK ELEMZÉSE IZOLÁLT SZÍVPREPARÁTUMOKON Tóth András Szegedi Tudományegyetem ÁOK Farmakológiai és Farmakoterápiai Intézet
Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon
 Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon Válasz Szokodi István MTA doktor opponensi véleményére Először is szeretnék
Tóth András MTA doktori értekezés Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon Válasz Szokodi István MTA doktor opponensi véleményére Először is szeretnék
Bírálat Dr. Tóth András Intracelluláris Ca2+ homeosztázis-változások hatásainak elemzése izolált szívpreparátumokon című MTA Doktori értekezéséről.
 DEBRECENI EGYETEM Általános Orvostudományi Kar Kardiológiai Intézet Igazgató: Prof. Dr. Édes István egyetemi tanár Tel. / Fax: 52-255-928 Klinikai Fiziológiai Tanszék Tanszékvezető: Prof. Dr. Papp Zoltán
DEBRECENI EGYETEM Általános Orvostudományi Kar Kardiológiai Intézet Igazgató: Prof. Dr. Édes István egyetemi tanár Tel. / Fax: 52-255-928 Klinikai Fiziológiai Tanszék Tanszékvezető: Prof. Dr. Papp Zoltán
Biofizika I. DIFFÚZIÓ OZMÓZIS
 1. KÍSÉRLET 1. kísérlet: cseppentsünk tintát egy üveg vízbe Biofizika I. OZMÓZIS 2012. szeptember 5. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet 1. megfigyelés: a folt lassan szétterjed és megfesti az egész
1. KÍSÉRLET 1. kísérlet: cseppentsünk tintát egy üveg vízbe Biofizika I. OZMÓZIS 2012. szeptember 5. Dr. Bugyi Beáta PTE ÁOK Biofizikai Intézet 1. megfigyelés: a folt lassan szétterjed és megfesti az egész
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
A jel-molekulák útja változó hosszúságú lehet. A jelátvitel. hírvivő molekula (messenger) elektromos formában kódolt információ
 elektromos formában kódolt információ") A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
Az intracelluláris kalcium túltelítődés kivédése az NCX gátlásával
 Az intracelluláris kalcium túltelítődés kivédése az NCX gátlásával PhD Értekezés Szepesi Judit, Msc Farmakológiai és Farmakoterápiaiai Intézet, Általános Orvostudományi Kar Szegedi Tudományegyetem Szeged
Az intracelluláris kalcium túltelítődés kivédése az NCX gátlásával PhD Értekezés Szepesi Judit, Msc Farmakológiai és Farmakoterápiaiai Intézet, Általános Orvostudományi Kar Szegedi Tudományegyetem Szeged
Az agyi értónust befolyásoló tényezők
 2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
repolarizációs tartalék
 A projekt négy munkaévében, a kutatási tervben kitűzött céloknak megfelelően, az antiaritmiás és proaritmiás hatás mechanizmusában szereplő tényezők vizsgálatára került sor, amely az alábbi fontosabb új
A projekt négy munkaévében, a kutatási tervben kitűzött céloknak megfelelően, az antiaritmiás és proaritmiás hatás mechanizmusában szereplő tényezők vizsgálatára került sor, amely az alábbi fontosabb új
A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására
 OTKA 62707 A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására Zárójelentés A gesztációs diabetes mellitus (GDM) egyike a leggyakoribb terhességi komplikációknak, megfelelő
OTKA 62707 A diabetes hatása a terhes patkány uterus működésére és farmakológiai reaktivitására Zárójelentés A gesztációs diabetes mellitus (GDM) egyike a leggyakoribb terhességi komplikációknak, megfelelő
A MASP-1 dózis-függő módon vazorelaxációt. okoz egér aortában
 Analog input Analog input 157.34272 167.83224 178.32175 188.81127 Relaxáció (prekontrakció %) Channel 8 Channel 8 Analog input Volts Volts Channel 12 A dózis-függő módon vazorelaxációt Vehikulum 15.80
Analog input Analog input 157.34272 167.83224 178.32175 188.81127 Relaxáció (prekontrakció %) Channel 8 Channel 8 Analog input Volts Volts Channel 12 A dózis-függő módon vazorelaxációt Vehikulum 15.80
A membránpotenciál. A membránpotenciál mérése
 A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A sejtek közötti kommunikáció módjai és mechanizmusa. kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok
 A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
Membránszerkezet, Membránpotenciál, Akciós potenciál. Biofizika szeminárium
 Membránszerkezet, Membránpotenciál, Akciós potenciál Biofizika szeminárium 2013. 09. 09. Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens
Membránszerkezet, Membránpotenciál, Akciós potenciál Biofizika szeminárium 2013. 09. 09. Membránszerkezet Biológiai membránok (citoplazma, sejten belüli membránféleségek) közös jellemzője: Nem kovalens