Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Nem vezikuláris release. Kapcsoltság
|
|
|
- Piroska Takácsné
- 8 évvel ezelőtt
- Látták:
Átírás
1 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia I. Kapcsoltság, Ca++ Kapcsoltság Glia szincícium Gap junctions Hemichannels Asztro network térbeli/időbeli szabályozása Gliális Ca++, Ca++ hullámok Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Transzporterek, egyéb glia eredetű faktorok Gliális neurotranszmitter transzporterek Glutamát transzporterek GABA transzporterek Glycin transzporterek Egyéb transzporterek Ozmolitikumok Glia eredetű neuropeptidek Glia eredetű növekedési faktorok

2 II. Agyi homeosztázis szabályozása Ion-homeosztázis az extracelluláris térben K+ spatial buffering Retina, K+ siphoning Extracellular space Cl - homeosztázis Ca++ homeosztázis ph szabályozás [Glu] ec szabályozása Ammónia [GABA] ec szabályozása Víz - homeosztázis szabályozása Swelling
![Cl - homeosztázis Ca++ homeosztázis ph szabályozás [Glu] ec](/docs-images/53/11818029/images/page_2.jpg "szabályozása Ammónia [GABA] ec szabályozása Víz - homeosztázis")
3 Érett makroglia: negatív nyugalmi membránpotenciál, ~ mv (negatívabb a neuronoknál), a K+ konduktancia túlsúlya miatt (ez a membránpotenciált a K+ egyensúlyi potenciáljához közel tartja) (Gliában nyugalmi állapotban csak K+-ra permeábilis a membrán, neuronban Na+, Cl- és K+ ionokra egyaránt). DE: glia e tekintetben nagyon heterogén társaság! Glia depolarizálható, de nem lesz regeneratív akciós potenciál Ioneloszlás gliasejtekben hasonló más sejtekhez, kivéve Cl -, mely magasabb asztroban és ODC-ben is más sejtekhez képest K + ic: ~ mm Na + ic: <10 mm Ca ++ ic: <0,0001 mm Cl - ic: ~30-40 mm

4 Glia fiziológia Verkhratsky és Butt, 2007



5 Ioncsatornák K+ csatornák (4 család, glián mindegyik jelen van) 1. Inward rectifier K+ csatornák - ha a membrán depolarizált, zárva vannak. - ha a membrán hiperpolarizált, (jobban mint E K ), nyitva vannak - inkább a befelé történő K+ influxot engedik - ezek felelősek elsősorban a glia a negatív nyugalmi membránpotenciáljáért - és fontosak az extracell K+ gyors eltávolításában!! See later! - 20 féle K IR - K IR 4.1 -/- egér: elpusztulnak 8-20 napon belül: fehérállomány nem fejlett eléggé Kir4.1.: pl. ODC sejteken, lásd 03. előadás



6 Ioncsatornák Kondicionális K IR 4.1 -/- csak GFAP expresszáló sejtekben: Ezek is elpusztulnak naposan. Szintén myelin károsodik!! Tehát van egy glia progenitor pool, mely GFAP-t is expresszál de tud oligodendrogliát is képezni (O2A de GFAP+ csak késői stádiumában): itt ez károsodik!! Fehérállomány vakuolizált...

7 Ioncsatornák 1. K IR csatornák - más típusok is jelen vannak klb. gliasejtekben, pl. - K IR 5.1 család coassembly with Kir4.1 - K IR 3.0 család G-protein kapcsolt neurotranszmitter receptorokhoz asszociáltan - K IR 6.1, 6.2 ATP függőek, akkor aktívak ha. ic. ATP szint nagyon lecsökken segítenek a negatív nyugalmi membránpotenciál fenntartásában metabolikus challenge idején 2. Delayed rectifier K+ csatornák, K D (lassan inaktválódók) 3. Rapidly inactivating A-type K+ channels, K A (gyorsan inaktválódók) 4. Calcium-activated K+ channels, K Ca - minden gliatípusban jelen vannak - sokféle K D csatorna de - K A -ból csak egyfélét, Kv1.4-et expresszálják gliasejtek - K Ca ból 3 féle van, gliában mindhárom (BK, IK, SK) előfordul - K D, K A, K Ca zárva vannak nyugalmi membránpotenciálon, - depolarizációkor nyitnak (-40 mv felett), ha ec K+ szint magas lesz - funkcióik: még bizonytalanok - Schwann sejtben Raniver-nél: K D -Kv1.5 és BK
 4. Calcium-activated K+ channels, K Ca - minden gliatípusban jelen vannak - sokféle K D csatorna de - K A -ból csak egyfélét, Kv1.")
8 Ioncsatornák Feszültségfüggő Na+ csatornák, Na V - sok gliatípusban - hasonlóak neuron/izom Na+ csatornáihoz - amiben különböznek: denzitásuk: 1 Na v /10 um 2 gliában (neuronban ez 10000/um 2 ) - szerepük??? de éretlen glia-típusokban és gliatumorokban több van Feszültségfüggő Ca++ csatornák, Ca v - általában glia prekurzorokon és éretlen gliasejteken: növekedés, migráció, prolif. - glia-fejlődés alatt downregulálódnak - reaktív gliában up-regulálódnak - éretlen oligodendrocita nyúlványokban jelen van myelinációban szerep? - érett asztro/myelináló oligo sejtekben mikrodoménekben: funkció itt? - glia-nyúlványok patch clampja problémás... Irodalom kevéske... egyelőre Ca v jelen vannak, működnek..

9 AnIoncsatornák Klorid és más anion csatornák - fontos és jellemző: asztrociták aktívan tudnak Cl t akkumulálni, magas az ic. Cl - koncentrációjuk (kb 35 mm) - ez főleg a Na + /K + /Cl - (NKCC) kotranszporter működésének az eredménye - csatorna nyitás: Cl - efflux - funkció: talán asztro swelling és ec. Cl - konc. szabályozása Jayakumar, Norenberg 2010

10 AnIoncsatornák - csak mostanában kezdjük megérteni jelentőségüket a glia-biológiában - még sok technikai limitáció: pl. csatorna-spec antagonisták, ellenanyagok hiánya - Cl- a legnagyobb mennyiségben jelenlevő anion - ezeken a csatornákon át általában más anionok is vándorolhatnak: pl aminosavak - Cl- csatorna-családok: - CFTR channels - Ca++-activated Cl- channels - voltage-dependent anion-selective channels (VDACs) - ClC channels Fontosak asztroban: volume regulated anion channels (VRACs) - Cl- csatornák asztroban való expressziója még csak néhány esetben igazolt, főleg in vivo adat kevés - Pl. ClC2 van HC asztrocitákban, VRAC gyakori swelling, ozmoreguláció
 - Cl- csatornák asztroban való expressziója még csak néhány esetben igazolt, főleg in vivo adat kevés - Pl.")
11 Klorid csatornák Összegyűjtötte: Jády Attila

12 Ioncsatornák Aquaporinok AQP - homotetramer - mindegyik monomer bidirekcionális H 2 O transzportot enged meg az adott ozmotikus gradiens mentén - központi pórus ionokra/gázokra áteresztő Badaut 2011

13 Ioncsatornák Aquaporinok I. Aquaporinok: - elsősorban vízcsatornák - AQP0, 1, 2, 4, 5, 6, 8 II. Aquaglyceroporins: - víz, urea, glicerol, monokarboxilát (piruvát, laktát) transzport - AQP3, 7, 9, 10 III. Super-Aquaporinok: - citoplazmatikusak, ic. víz transzport, - organellum és vezikulaméret és homeosztázis - AQP11, 12-7 féle AQP van rágcsáló agyban - AQP1,4,9 jelenleg legismertebbek/fontosabbak AQP1: - choroid plexus epitélben - cerebrospinal fluid formation AQP9: - májban a legnagyobb az expressziója - a hepatocitákban az AQP9 expresszióját a vér inzulin-tartalma szabályozza agyban is? - víz, glycerol, monokarboxilát diffúzió - agyban: tanyciták (ezek csak AQP9-et expresszálnak), endotél, egyes neuron populációk - glicerol, monokarboxilátok: energia szubsztrátok agyi energia metabolizmus!

14 Aquaporin4 Agyban leggyakoribb forma. Asztro végtalpon! M1: teljes hosszúságú AQP4 izoforma (splice variáns), ~34 kda, inkább egyedi csatornák M23: rövid izoforma -31 kda, nagy OAP, >100 partikulum TIRF images A: piros-quantum-dot jelölt AQP4 molekulák motilitása a membránban B: AQP4 szekvencia Crane 2008


15 Neurotranszmitter/neuromodulátor receptorok - szinte minden receptort expresszálnak, amit a neuronok: lokálisan azt, amit az adott környezet adott idegsejtjei - így képesek neuronális működést érzékelni Sőt: térbeli szegregáció: pl. gátló szinapszisok közelében Bergmann gliában inkább GABA receptorok koncentrálódnak - nagyon gyakran amire receptoruk van azt saját maguk is képesek szekretálni Verkhratsky és Butt, 2007

16 Neurotranszmitter/neuromodulátor receptorok Neurotransmitter/neuromodulator receptors in glial cells scheme showing the multiplicity of neurotransmitter receptors expressed in different types of glial cells. IICR InsP3-induced Ca2+release; CICR Ca2+-induced Ca2+ release. Ionotropic receptors: NChr Nicotinic Cholinoreceptors; GABAAR GABA receptors; GLY glycine receptors; GluR glutamate receptors (AMPA, NMDA and KA receptors); P2X purinoreceptors. Metabotropic receptors: VIP vasoactive intestinal polypeptide receptors; MChR muscarinic cholinoreceptors; NPY neuropeptide Y receptors; mglur metabotropic glutamate receptors; BK bradykinin receptors; V2 vasopressin receptors; H1R histamine receptors; OX oxytocin receptors; P2Y metabotropic purinoreceptors; 1AR adrenergic receptors; SbP substance P receptors; PAF platelet activating factor receptors; ETB endothelin receptors; 5-HT serotonin receptors Verkhratsky és Butt, 2007

17 Neurotranszmitter/neuromodulátor receptorok Astrocyte NT receptors Ionotropic receptors A. Glutamate receptors: AMPA/Kainate, NMDA receptors B. GABAAreceptors C. P2X (ATP) Purinoreceptors D. Glycine receptors E. Nicotinic cholinoreceptors NChR Metabotropic receptors A. Glutamate receptors, mglurs B. GABAB receptors C. Adenosine receptors: A1, A2, A3 D. P2Y (ATP) Purinoreceptors E. Adrenergic receptors: 1AR, 2AR F. Muscarinic cholinoreceptors: mchr M1 M5 G. Oxytocin and vasopressin Receptors H. Vasoactive Intestinal: VIPR 1,2,3 I. Serotonin receptors: 5-HT1A, 5-HT2A, 5-HT5A J. Angiotensin receptors: AT1, AT2 K. Bradykinin receptors: B1, B2 L. Thyrotropic-releasing hormone receptors: TRH1 M. Opioid receptors N. Histamine receptors: H1, H2 O. Dopamine receptors: D1, D2 Asztro: - rengeteg-féle NT receptor, neuropeptid, kemokin, citokin receptor Verkhratsky és Butt, 2007

18 Neurotranszmitter/neuromodulátor receptorok Oligodendrocyte NT receptors Ionotropic receptors A. Glutamate receptors: AMPA/Kainate, NMDA B. GABAA receptors C. Glycine receptors Metabotropic receptors A. Muscarinic cholinoreceptors: mchr M1, M2 B. P2Y (ATP) Purinoreceptors ODC: kevesebb NT receptor, mint asztron, P2Y purinoreceptor a leggyakoribb, de van AMPA és NMDA is az érett, myelináló sejteken OPC : A1 adenozin rec., mgur, GABAB, gylcin rec. developmentally regulated and regulate OPC differentiation and myelination Verkhratsky és Butt, 2007
 Purinoreceptors ODC: kevesebb NT receptor, mint asztron, P2Y purinoreceptor a leggyakoribb, de van AMPA és NMDA is az érett,")
19 Neurotranszmitter/neuromodulátor receptorok Schwann cell NT receptors Ionotropic receptors A. P2X (ATP) Purinoreceptors Metabotropic receptors A. P2X (ATP) Purinoreceptors B. Endothelin receptors, ETB C. Tachykinin receptors, NK1 Schwann: kevesebb NT receptor, mint asztron, P2X és P2Y purinoreceptorok Endotelin receptorok chronic inflammatory pain Tachykinin (vazodilatátor..) rec.s Verkhratsky és Butt, 2007

20 Neurotranszmitter/neuromodulátor receptorok Microglia NT receptors Ionotropic receptors A. P2X (ATP) Purinoreceptors B. Glutamate receptors: AMPA/Kainate Metabotropic receptors A. P2Y (ATP) Purinoreceptors B. GABAB receptors C. Muscarinic cholinergic receptors D. Cytokine/complement receptors E. Chemokine receptors (CCR1 5, CXCR4..) F. Endothelin receptors: ETB Mikro: - sokféle NT receptor, kemokin, citokin receptor Verkhratsky és Butt, 2007
 F.")
21 Glutamát receptorok - minden típus: 4-5 subunit - AMPAR, KAR: főleg Na+/K+ - NMDAR: nagy Ca++ permeabilitás - AMPAR: leggyorsabb deszenzitizáció, - NMDAR: leglassabb deszenzitizáció InsP3/DAG kaszkád camp kaszkád - asztrocitákon mglur1,3,5 - éretlen OPC-n és mikroglián is vannak
22 Ionotróp Glutamát receptorok AMPA-R - GluR1-4 - Na+/K+, gyors - ha GluR2 hiányzik: Ca++ is - asztrocitákban : gyakori, pl. cortex, hippocampus, cerebellum, retina, spinal cord, corpus callosum minden agyterületen területén - mikroglia szubpopulációban (?) KA-R - GluR5-7 és KA1-2 subunits, Na+/K+ - asztrocitákon, oligodendroglián elektrofiziológia hiányzik!! NMDA-R - NR1-NR2A-D-NR3A-3B subunits, Ca++ - lassú válaszok - kérgi, gerincvelői asztrocitákon, Müller glián sőt oligodendrocitákon sőt myelinhüvelyen De szerintük NEM direkt a Glu/GABA hatás mikroglián! NMDARs might relay information about the volume of electrical traffic of the underlying axon to the sheath and perhaps even to the parent oligodendrocyte.
23 GABA receptorok GABA A -asztrocitákban minden agyterületen -ligand-vezérelt Cl - csatorna, olyan mint a neuronális DE - asztroban Cl - ic ~35mM, neuronban 3-5 mm!! - asztroban Cl - ekvilibrium potenciál: - 40 mv, - neuronban Cl - ekvilibrium potenciál: - 70 mv, GABA A akitváció ra neuronba Cl- influx és hiperpolarizáció - gliában GABA A akitváció: Cl - efflux és depolarizáció következik be, SŐT a GABA A akitváció a gliális K+ csatornákat is gátolja és így facilitálja a depolarizációt GABA B néhány asztrocita és OPC szubpopulációban
24 Citokin és kemokin receptorok Minden gliatípusban, általában osztódás, Type I citokin receptorok növekedés, metabolizmus kontroll IL2 (beta), IL3, IL4, IL5, IL6, IL7, IL9, IL11, IL12, GM-CSF, G-CSF, Epo, LIF, CNTF, Thrombopoietin (TPO), Prolactin, Growth hormone Type II citokin receptorok IFN-alpha, IFN-beta, IFN-gamma, IL10, IL22, and tissue factor Chemokine receptorok CC chemokine receptors, CXC chemokine receptors, CX3C chemokine receptors, XC chemokine receptor Tumor necrosis factor receptor (TNFR) TGF-beta receptors TGFBR1, TGFBR2, and TGFBR3 Immunoglobulin szupercsaládba tartozó citokin receptorok IL1R (type I and II), IL6R, PDGFR, SCFR, CSF-1R, etc. JAK/STAT általában html
 -ET B -t")
 - ET A és ET B receptor van asztrocitákon: aktivációjukra csökken a GJ kapcsoltság!")
25 Endotelin receptorok - vazoaktív peptidek: simaizom-kontrakció, vérnyomásnövekedés - főleg endotélen - ET A, ET B1 and ET B2 receptorok - G protein coupled, ic Ca++ - ligand: ET1, ET2, ET3 (asztro szekretálja) -ET B -t találtak mikroglián is (Ca++ release ic. raktárakból) - ET A és ET B receptor van asztrocitákon: aktivációjukra csökken a GJ kapcsoltság!!! - swelling szabályozása gliában pl.
26
27 Astrocytes may play an important role in the genesis of glaucoma... zöldhályog Intraocular pressure (IOP) Ischemia/ hypoxia Tumor necrosis factor-α (TNF-α)
28 - C3, C4, C5 komplement Komplement rendszer fehérjék nagy glikoproteinek - ezek kis darabjai, a C3a, C4a, C5a az anaphylotoxin-ok (vérszérumban inflammatorikus reakciót indukálnak és ér áteresztő-képességet növelik; kemotaxis-reguláció, ROS termelés..) - asztro és mikroglián ezek receptorai expresszálódnak, és több komplement-komponenst saját maguk is szekretálhatnak
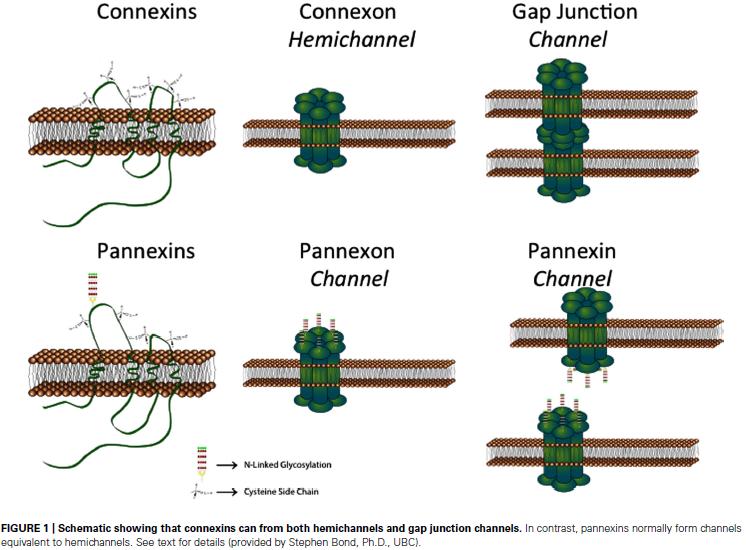
29 Purinoreceptorok Purinerg nukleotidok: ATP, adenozin és metabolitjaik Purinoreceptorok: P1-Adenozin és P2-ATP receptorok minden gliatípus expresszál valamennyit ezekből ATP: widespread gliotransmitter! 1972, Burnstock az ATP neurotranszmitter : nem adrenerg, nem kolinerg gátló neuronokban az autonóm idegrendszerben 1976, első purinreceptorok leírása 1992 után purinerg rendszer iránti érdeklődés nagyon megnő
30 Sejtből KI: exocitózis vagy transzportereken át AMP-nek jelenleg nincs ismert receptora NTPDases:ecto-nucleoside triphosphate diphosphohydrolases (ekto-atpázok) NTPDase 1-8 (humán), ATP ADP AMP 5-nucleotidase (5'-NT): AMP adenosine


31 Glia fiziológia Purinoreceptorok Adenozin P1 receptorok Koffein: AR antagonista A1, A2A, A2B, A3 receptorok - G protein coupled metabotrop - asztrocitákban mindhárom lehet - extracell. adenozin általában ATP bontás - ektonukleotidázok révén keletkezik (de lehetséges neuronális vagy gliális Adenozin release is) - AR stimulálás glutamát transzporter expressziót növeli asztrocitán - OPCn axon/glia kommunikációt közvetítenek


32 PT Porcine testis sejt Asztrocita + virus Viral Epidemics in a Cell Culture Gönci 2010
33 Primer asztrocita tenyészet + virus saját eredmény unpublished
34
35
36 Purinoreceptorok -ligand vezérelt ioncsatornák -trimerek: homo vagy heteromerek - ATP kötésre gyors konformációváltás; Na+, K+, Ca subunit, külön géneken kódolva -P2X7 aktivációra extra nagy pórust képez és hosszantartó Ca++ influxot enged meg aktivációjához nagy ATP konc. kell : ez neuronsérüléskor jellemző -asztro, ODC, Müller glia, mikro gyors deszenzitizációs idők P2X7, P2Y12 Mikroglia!! - klasszikus 7 TM metabotrop receptorok - CNS-ben: P2Y1, P2Y6, P2Y11, P2Y12, P2Y13, P2Y14 - ic Ca++ transiens (secmin) növekedése Coddou 2011 Verkhratsky és Butt, 2007
37 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia I. Kapcsoltság, Ca++ Kapcsoltság Glia szincícium Gap junctions Hemichannels Asztro network térbeli/időbeli szabályozása Gliális Ca++, Ca++ hullámok Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Transzporterek, egyéb glia eredetű faktorok Gliális neurotranszmitter transzporterek Glutamát transzporterek GABA transzporterek Glycin transzporterek Egyéb transzporterek Ozmolitikumok Glia eredetű neuropeptidek Glia eredetű növekedési faktorok
38 Glia szincícium (syncytium) Kapcsoltság Gerschenfeld H.M. elsők közt írta le, hogy a gliasejtek karakterisztikus sajátsága, hogy rés-kapcsolatokon át kapcsoltak - a neurotranszmitterek ezt a kapcsoltságot modulálják, neuron-asztro kommunikáció izom egy, többmagvú sejt szívizom több sejt, gap junctions pan-glial syncytium asztro-odc-ependyma.. recently Christian Giaume Ken McCarthy Cx43 asztro tenyészet..glial cells are not organized as a syncytium, as initially proposed, but rather as networks of communicating cells with defined spatial organization and plasticity, as their modalities of intercellular communication are controlled by endogenous signals in normal and in pathological situations..
39 active contribution of glia to information processing Kapcsoltság glial cells have developed mechanisms alternative to synaptic transmission that is characteristic of neurons gap junction channels (GJ) gap junctional plaques gap junction kifejezés: gap : 2-2,5 nm rés a sejtek között Connexins Cx Pannexins Panx hemichannels (HCs) Giaume, Liu
40
 connexon - ~20 altípus")
 - minden connexinben 4 TM domén - homomer vagy heteromer connexonok - homocelluláris vagy")
41 Gap junctions Paul Sorgen - két sejt közti rés csak 2-2,5 nm - connexinek (6 db) connexon - ~20 altípus (26-62 kda; pl. Cx43 43 kda) - minden connexinben 4 TM domén - homomer vagy heteromer connexonok - homocelluláris vagy heterocelluláris kapcsolat - 1,5 nm pórus: relatív nagy, akár 1 kda molekulák is átjuthatnak, Ip3, ATP, vitaminok és ionok elektromos kapcsoltság is - open - closed állapotok - hemichannels
42 GJ csatorna nyitását szabályozza: - nagy Ca ++ ic : permeabilitás - ph : permeabilitás - connexin foszforiláltság, diszulfid hidak.. - feszültségfüggés Gap junctions Asztroban is van Hiánya: congenital deafness Plug gating model for transjunctional voltagedependent gating of the Cx26 gap junction channel. Maeda 2011 A: ha nincs feszültségklbség a két sejt között: N-terminális hélixek (NTH) az 1 TM régióhoz simulnak hidrofób módon B: pozitív elektromos mező kihúzza az Asp2-t (ez negatív töltésű-), NTH a pórusba kerül, dugót képez.
43 Connexinek - Pannexinek PanX - gerinctelen innexinekkel homológok így azonosítják őket - tulképp nagy transzmembrán csatornák: csak hemichannel-t képeznek Kapcsoltság Oroszok Asztro: - PanX: egyelőre nincs expressziójukra meggyőző bizonyíték - főleg: Cx43, Cx30, kevesebb: Cx26, Cx40, Cx45 - Cx43 embrionális kortól, Cx30-2. posztnat héttől csak szürkeállományban Mikroglia: - aktivációra Cx43, illetve Panx1, Cx32 Neuronok: - Cx26, Cx32, Cx36, Cx37, Cx40, Cx43, Cx45, Cx47 illetve Cx43 és Cx32 is jelen lehet - Cx36 csak neuronokban! Schwann sejt - myelináló sejt: Cx32 - éretlen Schwann: Cx46 NG2 glia: - nem kapcsolt (nincs erre jelenleg adat) - Panx1 is jelen van több neuronális populációban és PSD95-tel kolokalizál: szinaptikus funkcióban is szerepe lehet, szintje már születés környékén magas - Panx2 is jelen van neuronokban, főleg felnőttben Giaume, Liu 2011
44 Kapcsoltság Astrocyte astrocyte (A/A) coupling pan-glial syncytium - Cx30 és Cx43 kolokalizál asztro GJ plakkokban - variációk: Cx30/Cx30, Cx43/Cx30, and Cx43/Cx43 - eddigi elfiz adatok: kolokalizáció ellenére inkább homotipikus GJ valószínű (nagyobb festékterjedés, ha csak Cx30-at vagy Cx43-at expresszáltatnak pl. HeLa sejtekkel) Astrocyte oligodendrocyte (A/O) coupling Lásd még: 3. előadás! - GJ-t sokkal inkább a szomszédos asztrokkal képeznek, mint más ODC-kal - A/O kapcsolat heterotipikus kell legyen, mert ODC-k más Cx-ket expresszálnak, mint asztrok: Cx29, Cx32, Cx45, Cx47 Oligodendrocyte oligodendrocyte (O/O) coupling - újabban van csak néhány adat O/O GKJ kapcsolatra (corpus callosumban) - Cx32 és Cx47 vesz részt ezekben az O/O kacsolatokban Neuron-glia coupling - kevés bizonyíték van neuron-glia GJ kapcsoltságra - neuron-neuron inkább? Neuronális kapcsoltság nő, ha neuron sérül! Giaume, Liu 2011 Orthmann-Murphy 2008
45 Kapcsoltság - átlagosan egy asztro-pár a szürkeállományban 230 GJ-vel kapcsolt: magas fokú kapcsoltság!! 300 um slice - Lucifer yellow, Alexa dyes (~450 kda), biocitin: 1 asztroba injektálva szomszédos asztro jelölődik - kapcsoltság mértéke agyterületenként nagyon eltér: cortexben szinte teljes kapcsoltság (~100%); látóidegben 80%, HC-ban csak 50%, corpus callosumban szinte semmi 50 um slice Astrocytic coupling in the neocortex and in the corpus callosum. Biocytin. GFAP-GFP mice
46 Kapcsoltság P11 egér Asztro biocitin jelölése cortexben Asztro azonosítás morfológia/elfiz alapján Houade, Giaume 2006
 kapcsoltak, mint CA3-ban - ennek")
47 P11 egér, Asztro biocytin jelölése HC-ban, CA1 Glia fiziológia Kapcsoltság A1,A2: stratum radiatum közepén - koncentrikus B1 C2: stratum radiatum közel a piramissejt-réteghez : ekkor 2 féle jelölés B1,B2: biocytin jelölés megáll a pirmissejt-rétegben C1,C2: biocytin megjelöl asztrokat a stratum oriens-ben is - asztrociták a CA1-ben jobban (dye- coupled) kapcsoltak, mint CA3-ban - ennek funkcionális jelentősége még?? - de: CA1 kevés kapilláris CA3-hoz képest: metabolikus szignálok jobban terjedhetnek a kevésbé vaszkularizált területeken így?? Houade, Giaume 2006
48 Glia szincícium (syncytium) Kapcsoltság vizsgálata Glia fiziológia Kapcsoltság - elektrofiziológia, ionáramok, inhibitorok: carbenoxolone, GRA,.. - dye coupling - intracellular calcium imaging - flash photolysis of caged compounds - recovery after photobleaching (FRAP) slide - local activation (LAMP) of fluorescent probes
49 Kapcsoltság barrel cortex: szomatoszenzoros kéreg IV. réteg
50 Kapcsoltság Such anatomofunctional organization offers an ideal model to study how astrocytic networks are organized in reference to neuronal compartments A: akut szelet, infravörös megvilágítás B: morfológia és méret alapján kiválasztott asztro egy patch pipetta végén C, D: biocytin immunfestés, P5 és P10 E: carbenoxolone (100M) F: biocytin+ sejtek számolása: P5- P10 közt nagy váltás
51 Kapcsoltság A C: Barrel cortex, már P6-nál több a Cx43 a barrelekben. Diffúz, stellate pattern. D-E: Cx43 koronális és tangenciális metszeten, P10 F, G, Cx43 koronális metszeteken a barreleken belüli és azokon kívüli területeken, P20 H: Cx30 tangenciális metszeten, P20 I K: Cx43 és Cx30 fenőttben megmarad. Houades 2008


52 Kapcsoltság A1-A2: festékterjedés 20 perccel sulphorodamin beadása után, barrelen belüli asztrociták közt B1-B2, festékterjedés 20 perccel sulphorodamin beadása után, barrelek közti septumban levő asztrociták közt Houades 2008 P10, biocytin és sulforhodamine B-vel kapott kapcsoltság nagyon más!!! : kapcsoltsági csoportok mérete: Biocytin: és SB:15+-1 astrocytic networks parallel the columnar compartmentalization of neurons in the somatosensory cortex Houades 2008


53 - asztrok kapcsoltak glomerulusokon belül, de glomerulusok között nem - KO egerek: Cx30 végzi itt a kapcsolást bár Cx43 is van Kapcsoltság
54 Sejtből KI: exocitózis vagy transzportereken át PURINERG RENDSZER AMP-nek jelenleg nincs ismert receptora NTPDases:ecto-nucleoside triphosphate diphosphohydrolases (ekto-atpázok) NTPDase 1-8 (humán), ATP ADP AMP 5-nucleotidase (5'-NT): AMP adenosine
55
56 Asztro network térbeli/időbeli szabályozása milyen connexint expresszálnak asztrok Cx kipakolódás/visszavétel szabályozása GJ nyitottság szabályozása neuronális aktivitás, más agyi sejtek által termelt faktorok, glia-aktiváció : neurotranszmitterek, endotelinek (gátlás!), citokinek Cx kompatibilitás (homo/heteromerek) csatorna szelektivitása (tradicionálisan poorly selective channels de a dye spreading nem azonos biomolekulák átjutásával!! ) csatorna feszültség-függése szintén szűrheti, milyen molekulák merre jutnak át Giaume, Liu 2011! Cx43 átengedi: Kapcsoltság - camp, InsP3 - Ca2+ - AS: glutamate, aspartate and taurine - nucleotidok: ADP, ATP, CTP, NAD - energia-metabolitok: glucose, glucose-6-phosphate and lactate - kis peptidek pl. glutathione - RNS Cx30 átengedi: - kevesebb info, eddig: -ATP, InsP3, aspartate, glutamate, glucose and lactate - selective for cations over anions Cx30/Cx30 és Cx43/Cx43 és Cx30/Cx43 csatornák is feszültségfüggőek (konduktanciájuk a transzmembrán feszültségtől függ) - sejtek izolálhatják magukat pl. sérült sejtektől
57 Ca++ szignalizáció nagyon általános, rengeteg minden szabályozza ic Ca++ szintet: térben/időben nagyfokú szabályozottság kell! Ca++ Glia fiziológia szabad Ca++ kevés a sejtben! NMDA rec! LTP...
58 Ca++ homeosztázis fő útvonalai/komponensei Glia fiziológia Ca++ Store operated Ca++ channel (SOOC) vagy capacitative Ca++ entry ic Ca++ raktár kiürülése Ca++ influxot indukál Verkhratsky Butt 2007 NCX Na+/Ca2+ exchanger; PMCA Plasmalemmal Calcium ATP-ase; Ca2+ BP Ca2+ binding proteins; InsP3R Inositol-1,4,5-trisphosphate Receptor/Inositol-1,4,5-trisphosphate-gated Ca2+ channel; RyR Ryanodine Receptors/Ca2+-gated Ca2+ channel; SERCA Sarco(Endo)plasmic Reticulum Calcium ATPase. Intra-ER Ca2+ binding proteins also act as Ca2+ dependent chaperones, which are enzymes controlling protein folding into the tertiary structure. PTP permeability transition pore
59 * Calcium-induced Ca++ release koffein! ic Ca++ koncentráció : RyR2, RyR3 receptorokat (csak ez a szignál!) aktiválja:er-ból is Ca++ release citoszolba * Depolarization-induced Ca++ release Depolarizáció RyR1 receptor-aktiváció * InsP3-induced Ca++ release ha ic Ca++ : InsP3-Rec érzékenysége nő InsP3-ra * Ca++ spark, puff egyetlen RyR vagy Insp3R nyitása miatt bekövetkező citoszolikus Ca++ és ez továbbadódhat: * propagating intracelllular Ca++ wave * Ca++ excitotoxicity ha túl sokáig magas az ic Ca++ gliában ez a fő útvonal gliában RyR szerepe Ca++ signalingban kicsi vagy semmi Glia fiziológia Ca++ RyR és InsP3 Ca++ érzékenysége miatt alakulhat ki ez a tovaterjedő Ca++ hullám nem maga a Ca++ ion terjed tova, hiszen ez azonnal kikötődik Ca++kötő fehérjékhez hanem újabb és újabb Ca++ efflux történik!
60 Ca++ - glia neuron közti különbség: gliasejtekben ritka a feszültségfüggő Ca++ csatorna: legtöbb érett gliában (asztro, ODG, Schwann) nincs, progenitorokban van éréskor eltűnik - másik különbség: glia ER-ben a Ca++ konc.: um (neuronban: uM) - Ca++ influx érett gliába: főleg ligand-gated csatornákon: ionotróp glutamát receptor, P2X purinoreceptor - metabotrop receptorok Ca++ hullám indításában fontosak Ca++ wave starfish egg Carroll 1997 metabotrop receptorok InsP3 InsP3Rec aktiváció ER magas Ca++ szint tovább fennmarad, mint maga a stimulus ER kimerül Ca++ influx (SOOC) ER-ből Ca++ release citoszolba - ez lehet egyszeri (peak), több száz mp-ig tartó Ca++ plateau - vagy ismétlődő oszcilláció!!


61 Asztroglia monolayer in vitro
62 propagating intercelllular Ca++ wave Glia fiziológia Ca++ calcium hullám átugrik a sejtmentes sávon Ca ++ hullám a stimulált sejt körül ATP Ca ++ hullám amit a perfúzió eltérít Hassinger 1996
63 propagating intercelllular Ca++ wave fenntartása: Glia fiziológia Ca++ waves A: intracell InsP3 diffúzió B: regeneratív Ca2+- függő gliotransmitter release és extracell diffúzió C: fokális gliotranszmitter release, mely nagy távolságra diffundál (több száz um-re is, ~15-20 um/sec) Asztrocitahálózatok az agy klb területein máshogy szabályozódnak: Pl. Cx43 KO esetén kéregben nincs hullám de HC-ban megmarad Neuronális akciós potenciál: ms Asztro Ca++ hullám: sec-min
64 Ca++ waves Spontán calcium oszcillációk asztrocitákban. Hippocampus, CA1. A: Calcium Green AM B: GFAP/S100 immunostaining. C: overlay D: average fluorescence intensities from boxes 1 5 normalized to baseline level, and plotted over time. Four of the 5 cells exhibited spontaneous calcium oscillations and all cells responded to tacpd (mglur agonist) application with a calcium increase. s.r., stratum radiatum; s.p., stratum pyramidale
65 Ca++ waves Ca++ oszcillációk HC asztro Nyúlványaiban Oregon Green BAPTA-1 töltés A: regions of interest (ROI) B: a 4 nyúlvány elég különböző Ca++ jeleket ad C. II. csoport kinagyítva: Ca++ ROIs 9-10 területén kezdődött, innen ic. calcium hullám halad végig a nyúlványon Nett 2002
66 Ca++ waves Nucleus accumbens, Patkány agyszelet Fluo-4AM töltött sejtek, ATP hatás Molnár, Kardos 2011
67 Na de mire jók ezek a Ca++ hullámok?
68 Intercelluláris Ca++ hullámok : - normál felnőtt agyban, in vivo :??? - in vitro - in vivo a fejlődő agyban - specializált szövetekben (pl. Retina) - patológiás esetekben (pl. Alzheimer kór, epilepszia ) ha szinaptikus aktivitást gátolják és fesz.függő Ca++ csatornákat blokkolják neuronokon akkor is kilakulhat lassú neuronális depolarizáció: photolysis of caged Ca2+ elegendő gliális Ca++ szint növekedés kiváltásához gliális Glutamát release epileptikus aktivitás
69 ..We monitored calcium activity simultaneously from hundreds of mouse hippocampal astrocytes in vivo and found that almost all astrocytes participated en masse in regenerative waves that propagated from cell to cell (referred to here as glissandi ). Glissandi emerged depending on the neuronal activity and accompanied a reduction in infraslow fluctuations of local field potentials and a decrease in the flow of red blood cells. This novel phenomenon was heretofore overlooked, probably because of the high vulnerability of astrocytes to light damage; glissandi occurred only when observed at much lower laser intensities than previously used......by using 2-photon microscopy in rodent cerebellar cortex labeled with fluorescent indicator dyes or the calcium-sensor protein G-CaMP2, we discovered spontaneous calcium waves that filled approximately ellipsoidal domains of Bergmann glia processes. Waves spread in 3 dimensions at a speed of 4-11 microm/s to a diameter of approximately 50 microm, slowed during expansion, and were reversibly blocked by P2 receptor antagonists. Consistent with the hypothesis that ATP acts as a diffusible trigger of calcium release waves, local ejection of ATP triggered P2 receptor-mediated waves that were refractory to repeated activation. Transglial waves represent a means for purinergic signals to act with local specificity to modulate activity or energetics in local neural circuits... genetically encoded calcium indicator
 of the mouse cerebellar cortex in vivo.")
70 Glial calcium waves constitute a means to spread signals between glial cells and to neighboring neurons and blood vessels. These waves occur spontaneously in Bergmann glia (BG) of the mouse cerebellar cortex in vivo. Here, we tested three hypotheses: (1) aging and reduced blood oxygen saturation alters wave activity; (2) glial Ca(2+) waves change cerebral oxygen metabolism; and (3) neuronal and glial wave activity is correlated. We used two-photon microscopy in the cerebellar cortexes of adult (8- to 15-week-old) and aging (48- to 80-week-old) ketamine-anesthetized mice after bolus loading with OGB-1/AM and SR101. We report that the occurrence of spontaneous waves is 20 times more frequent in the cerebellar cortex of aging as compared with adult mice, which correlated with a reduction in resting brain oxygen tension. In adult mice, spontaneous glial wave activity increased on reducing resting brain oxygen tension, and ATP-evoked glial waves reduced the tissue O(2) tension. Finally, although spontaneous Purkinje cell (PC) activity was not associated with increased glia wave activity, spontaneous glial waves did affect intracellular Ca(2+) activity in PCs. The increased wave activity during aging, as well as low resting brain oxygen tension, suggests a relationship between glial waves, brain energy homeostasis, and pathology.
71 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia I. Kapcsoltság, Ca++ Kapcsoltság Glia szincícium Gap junctions Hemichannels Asztro network térbeli/időbeli szabályozása Gliális Ca++, Ca++ hullámok Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Transzporterek, egyéb glia eredetű faktorok Gliális neurotranszmitter transzporterek Glutamát transzporterek GABA transzporterek Glycin transzporterek Egyéb transzporterek Ozmolitikumok Glia eredetű neuropeptidek Glia eredetű növekedési faktorok
72 Volterra, Meldolesi 2005 Glia fiziológia Gliotranszmitterek Asztrocitákban!
73 Gliotranszmitterek Nem vezikuláris release Transzporterek megfordulása pl. Glu release asztroból csak patológiás esetekben Hemichannels Glu, Asp, ATP release P2X7 purinoreceptorok nagy pórus ATP release patológiás esetekben, ha extracell ATP szint magas Volume-activated anion channels glutamát és taurin ürülése; hipozmotikus körülmények közötti asztro swelling hatására nyílnak Pl.: pituicitákból taurin release, ez VP/OT neuronok glicinreceptoraival kerül kapcsolatba: VP/OT release: a test ozmotikus homeosztázisának szabályozása
74 Gliotranszmitterek Vezikuláris release (exocitózis) - lokális!! Ca++ indukálja a plazmamembrán/vezikula fúziót - vezikuláris Ca++ szenzor: synaptotagmin I - vezikuláris még: synaptobrevin II - plazmamembrán: syntaxin, SNAP25 - ezek alkotják együtt a SNARE komplexet - asztroban minden komponens jelen van - VLUT is van asztroban - szinaptikus-szerű mikrovezikulák... stb, persze sokkal komplexebb...
75 ELMI: szinaptikus-szerű mikrovezikulák (SLMVs) asztrocitában (hippocampus) Gliotranszmitterek Vezikuláris release (exocitózis) méretre és formára is hasonlítanak a szinaptikus vezikulákhoz Volterra, Meldolesi 2005
76 Gliotranszmitterek Vezikuláris release (exocitózis) Asztro glutamát release-t stimulálhatják: - P2YR, mglur, bradykinin, BDNF receptor aktiváció - Ca++ kelátorok (pl. BAPTA-AM) teljesen gátolják - neuron: feszültség-függő Ca++ influx: gyors -glia: Ca++ belső raktárakból: lassú, de hosszan tartó Asztro szekretálhat D-serine-t is, ez speciális gliotranszmitter, glia gyártja L- serin-ből racemase enzim révén: NMDAR glycin kötő-helyeit stimulálja Asztro/neuron kommunikáció! - HC asztrocitákban speciális exocitózis: kiss and run exocitózis: vezikula rövid ideig (~2 ms) nyit, aztán zár nem ürül ki teljes tartalma!

![[Ca ++ ] ic hatására FM-64 kiürül a vezikulumokból](/docs-images/27/11818029/images/77-0.png "VGLUT EGFP TIRF FM-64 festék együtt Volterra,")
77 Gliotranszmitterek Glutamát exocitózis asztrocitákból TIRF mikroszkópia 2004 Bezzi, Paola [Ca ++ ] ic hatására FM-64 kiürül a vezikulumokból VGLUT EGFP TIRF FM-64 festék együtt Volterra, Meldolesi 2005


78 astrocytes expressing the fluorescent synaptobrevin 2 derivative, synapto-phluorin
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Nem vezikuláris release. Kapcsoltság
 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák. Nem vezikuláris release
 I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz mitter/neuro
I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz mitter/neuro
Glia fiziológia I. Glia szincícium azaz network! Gap junctions. Hemichannels. Asztro network térbeli/időbeli szabályozása
 Ioncsatornák Aquaporinok Neurotranszmitter/neuro -modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia
Ioncsatornák Aquaporinok Neurotranszmitter/neuro -modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia
Glia fiziológia I. Glia szincícium azaz network! Gap junctions. Hemichannels. Asztro network térbeli/időbeli szabályozása
 Ioncsatornák Aquaporinok Neurotranszmitter/neuro -modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia
Ioncsatornák Aquaporinok Neurotranszmitter/neuro -modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok Glia fiziológia
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Nem vezikuláris release. Kapcsoltság
 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Glia fiziológia I. Kapcsoltság. Glia szincícium/network. Gap junctions. Hemichannels. Asztro network térbeli/időbeli szabályozása
 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Purinoreceptorok Endotelin receptorok Citokin és kemokin receptorok Komplement
Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Purinoreceptorok Endotelin receptorok Citokin és kemokin receptorok Komplement
CSF1*-R antagonizálás = szelektív mikroglia irtás (depléció) ugyanakkor: asztroglia depléció: letális
 ugyanakkor: asztroglia depléció: letális") 22222222222222,----------------------------------------------*/444444 *colony stimulating factor 1 (CSF1) = macrophage colony-stimulating factor (M-CSF) CSF1*-R antagonizálás = szelektív mikroglia irtás
22222222222222,----------------------------------------------*/444444 *colony stimulating factor 1 (CSF1) = macrophage colony-stimulating factor (M-CSF) CSF1*-R antagonizálás = szelektív mikroglia irtás
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus
 Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Ca 2+ Transients in Astrocyte Fine Processes Occur Via Ca 2+ Influx in the Adult Mouse Hippocampus Ravi L. Rungta, Louis-Philippe Bernier, Lasse Dissing-Olesen, Christopher J. Groten,Jeffrey M. LeDue,
Asztrociták: a központi idegrendszer sokoldalú sejtjei. 2009.11.04. Dr Környei Zsuzsanna
 Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
Asztrociták: a központi idegrendszer sokoldalú sejtjei 2009.11.04. Dr Környei Zsuzsanna Caenorhabditis elegans 1090 testi sejt 302 idegsejt 56 gliasejt Idegi sejttípusok Neural cell types Idegsejtek Gliasejtek
AsztroGlia - neuron interakció
 2011.04. 06. AsztroGlia - neuron interakció protoplazmás asztroglia (szürkeállomány); rostos asztroglia (fehérállomány); oligodendroglia (CNS); Schwann sejt (PNS); radiális glia (cortex); Bergmann glia
2011.04. 06. AsztroGlia - neuron interakció protoplazmás asztroglia (szürkeállomány); rostos asztroglia (fehérállomány); oligodendroglia (CNS); Schwann sejt (PNS); radiális glia (cortex); Bergmann glia
9. előadás Sejtek közötti kommunikáció
 9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
9. előadás Sejtek közötti kommunikáció Intracelluláris kommunikáció: Elmozdulás aktin szálak mentén miozin segítségével: A mikrofilamentum rögzített, A miozin mozgékony, vándorol az aktinmikrofilamentum
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika
 Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Az ioncsatorna fehérjék szerkezete, működése és szabályozása. A patch-clamp technika Panyi György 2014. November 12. Mesterséges membránok ionok számára átjárhatatlanok Iontranszport a membránon keresztül:
Humán asztrociták. Nagyobb és komplexebb. idegrendszeri fejlődésben jelentős szerepű
 Humán asztrociták Nagyobb és komplexebb idegrendszeri fejlődésben jelentős szerepű Forrás: Human vs Rodent astrocytes. (Courtesy Alexi Verkhratsky (Chapter 3), Neuroglia by Kettenmann) Glial Progenitor
Humán asztrociták Nagyobb és komplexebb idegrendszeri fejlődésben jelentős szerepű Forrás: Human vs Rodent astrocytes. (Courtesy Alexi Verkhratsky (Chapter 3), Neuroglia by Kettenmann) Glial Progenitor
Egy idegsejt működése. a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza
A kémiai szinapszis (alapok)
") A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
A preszinapszis A kémiai szinapszis (alapok) preszinaptikus neuron 1 akciós potenciál 2 Ca 2+ axon végbunkó (preszinapszis) Ca 2+ szinaptikus vezikula feszültség-függő Ca 2+ csatorna citoplazma szinaptikus
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet
 Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
Debreceni Egyetem Orvos- és Egészségtudományi Centrum Biofizikai és Sejtbiológiai Intézet Az ioncsatorna fehérjék szerkezete, működése és szabályozása Panyi György www.biophys.dote.hu Mesterséges membránok
a. Nyugalmi potenciál b. Transzport proteinek c. Akciós potenciál. Nyugalmi potenciál. 3 tényező határozza meg:
 Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Egy idegsejt működése a. Nyugalmi potenciál b. Transzport proteinek c. Nyugalmi potenciál Az ionok vándorlása 5. Alacsonyabb koncentráció ioncsatorna membrán Passzív Aktív 3 tényező határozza meg: 1. Koncentráció
Gyógyszerészeti neurobiológia. Idegélettan
 Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az idegrendszert felépítő sejtek szerepe Gyógyszerészeti neurobiológia. Idegélettan Neuronok, gliasejtek és a kémiai szinapszisok működési sajátságai Neuronok Információkezelés Felvétel Továbbítás Feldolgozás
Az adenozin Adenozin receptorok:
 Az adenozin Nukleinsavak és energiaraktározó vegyületek építőeleme Jelenléte ATP hidrolízisére utal -> extracelluláris szintje utal a korábbi neuronális és gliális aktivitásra Adenozin receptorok: 1-es
Az adenozin Nukleinsavak és energiaraktározó vegyületek építőeleme Jelenléte ATP hidrolízisére utal -> extracelluláris szintje utal a korábbi neuronális és gliális aktivitásra Adenozin receptorok: 1-es
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA NOVEMBER
 Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
Asztroglia Ca 2+ szignál szerepe az Alzheimer kórban FAZEKAS CSILLA LEA 2017. NOVEMBER Az Alzheimer kór Neurodegeneratív betegség Gyógyíthatatlan 65 év felettiek Kezelés: vakcinákkal inhibitor molekulákkal
Computational Neuroscience
 Computational Neuroscience Zoltán Somogyvári senior research fellow KFKI Research Institute for Particle and Nuclear Physics Supporting materials: http://www.kfki.hu/~soma/bscs/ BSCS 2010 Lengyel Máté:
Computational Neuroscience Zoltán Somogyvári senior research fellow KFKI Research Institute for Particle and Nuclear Physics Supporting materials: http://www.kfki.hu/~soma/bscs/ BSCS 2010 Lengyel Máté:
A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA
 A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA 2017.11.14. AZ ASZTROGLIA SEJTEK FONTOSABB TULAJDONSÁGAI AZ EPILEPTIKUS AKTIVITÁS SZEMPONTJÁBÓL (Devinsky és mtsai.,
A GLIASEJTEK ÉS AZ EPILEPTIKUS AKTIVITÁS KAPCSOLATA GÁSPÁR ATTILA GLIA SEJTEK ÉLETTANA EA 2017.11.14. AZ ASZTROGLIA SEJTEK FONTOSABB TULAJDONSÁGAI AZ EPILEPTIKUS AKTIVITÁS SZEMPONTJÁBÓL (Devinsky és mtsai.,
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV
 KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
KÉSZÍTETTE: BALOGH VERONIKA ELTE IDEGTUDOMÁNY ÉS HUMÁNBIOLÓGIA SZAKIRÁNY MSC 2015/16 II. FÉLÉV TÉNYEK, CÉLOK, KÉRDÉSEK Kísérlet központja Neuronok és réskapcsolatokkal összekötött asztrocita hálózatok
Ioncsatorna szerkezetek
 Jellegzetes Ioncsatorna szerkezetek Ördög Balázs Farmakológiai és Farmakoterápiai Intézet Kapuzás Feszültség szabályozott Voltage-gated Fesz. szab. Na +, +, Ca 2+ 2+,, K + + csatornák channels Transiens
Jellegzetes Ioncsatorna szerkezetek Ördög Balázs Farmakológiai és Farmakoterápiai Intézet Kapuzás Feszültség szabályozott Voltage-gated Fesz. szab. Na +, +, Ca 2+ 2+,, K + + csatornák channels Transiens
Az idegsejtek kommunikációja. a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus PERIFÉRIÁS IDEGRENDSZER Receptor
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban
 A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
A sejtmembrán szabályozó szerepe fiziológiás körülmények között és kóros állapotokban 17. Központi idegrendszeri neuronok ingerületi folyamatai és szinaptikus összeköttetései 18. A kalciumháztartás zavaraira
Glia - neuron interakció
 Glia - neuron interakció 2017.05.05. kornyei@koki.hu Gliális sejttípusok az idegrendszerben neuroektodermális eredet (kivéve mikroglia) NEUROGLIA glia glia (görög): ragadós 1858, Rudolf Virchow agyi kötőszövet
Glia - neuron interakció 2017.05.05. kornyei@koki.hu Gliális sejttípusok az idegrendszerben neuroektodermális eredet (kivéve mikroglia) NEUROGLIA glia glia (görög): ragadós 1858, Rudolf Virchow agyi kötőszövet
Egy idegsejt működése
 2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
2a. Nyugalmi potenciál Egy idegsejt működése A nyugalmi potenciál (feszültség) egy nem stimulált ingerelhető sejt (neuron, izom, vagy szívizom sejt) membrán potenciálját jelenti. A membránpotenciál a plazmamembrán
A membránpotenciál. A membránpotenciál mérése
 A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A membránpotenciál Elektromos potenciál különbség a membrán két oldala közt, E m Cink Galvani (1791) Réz ideg izom A membránpotenciál mérése Mérési elv: feszültségmérő áramkör Erősítő (feszültségmérő műszer)
A sejtek közöti kommunikáció formái. BsC II. Sejtélettani alapok Dr. Fodor János
 A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
A sejtek közöti kommunikáció formái BsC II. Sejtélettani alapok Dr. Fodor János 2010. 03.19. I. Kommunikáció, avagy a sejtek informálják egymást Kémiai jelátvitel formái Az üzenetek kémiai úton történő
a. Szinaptikus jelátvitel b. Receptorok c. Szignál transzdukció neuronokban d. Neuromoduláció. Szinaptikus jelátvitel.
 Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
Az idegsejtek kommunikációja a. Szinaptikus jelátvitel b. eceptorok c. Szignál transzdukció neuronokban d. Neuromoduláció Szinaptikus jelátvitel Terjedő szignál 35. Stimulus eceptor végződések Érző neuron
A sejtek közötti közvetlen (direkt) kapcsolat
 kapcsolat") A sejtek közötti közvetlen (direkt) kapcsolat rés-kapcsolat vagy gap junction ingerlékeny sejteknél elektromos szinapszis - kétirányú jeladás - gyors jelátadás (nincs "szünet") - egyszerű szabályozás,
A sejtek közötti közvetlen (direkt) kapcsolat rés-kapcsolat vagy gap junction ingerlékeny sejteknél elektromos szinapszis - kétirányú jeladás - gyors jelátadás (nincs "szünet") - egyszerű szabályozás,
A jel-molekulák útja változó hosszúságú lehet. A jelátvitel. hírvivő molekula (messenger) elektromos formában kódolt információ
 elektromos formában kódolt információ") A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
A jelátvitel hírvivő molekula (messenger) elektromos formában kódolt információ A jel-molekulák útja változó hosszúságú lehet 1. Endokrin szignalizáció: belső elválasztású mirigy véráram célsejt A jelátvitel:
1. Mi jellemző a connexin fehérjékre?
 Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
Sejtbiológia ea (zh2) / (Áttekintés) (1. csoport) : Start 2019-02-25 20:35:53 : Felhasznált idő 00:01:02 Név: Minta Diák Eredmény: 0/121 azaz 0% Kijelentkezés 1. Mi jellemző a connexin fehérjékre? (1.1)
S-2. Jelátviteli mechanizmusok
 S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
S-2. Jelátviteli mechanizmusok A sejtmembrán elválaszt és összeköt. Ez az információ-áramlásra különösen igaz! 2.1. A szignál-transzdukció elemi lépései Hírvivô (transzmitter, hormon felismerése = kötôdés
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Nem vezikuláris release. Kapcsoltság
 Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
Csatornák, receptorok Ioncsatornák Aquaporinok Neurotransz mitter/neuro modulátor receptorok Glutamát receptorok GABA receptorok Citokin és kemokin receptorok Endotelin receptorok Komplement rendszer Purinoreceptorok
-Két fő korlát: - asztrogliák rendkívüli morfológiája -Ca szignálok értelmezési nehézségei
 Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Nature reviewes 2015 - ellentmondás: az asztrociták relatív lassú és térben elkent Ca 2+ hullámokkal kommunikálnak a gyors és pontos neuronális körökkel - minőségi ugrás kell a kísérleti és analitikai
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: Az orvosi biotechnológiai mesterképzés
Intracelluláris ion homeosztázis I.-II. Február 15, 2011
 Intracelluláris ion homeosztázis I.II. Február 15, 2011 Ca 2 csatorna 1 Ca 2 1 Ca 2 EC ~2 mm PLAZMA Na /Ca 2 cserélő Ca 2 ATPáz MEMBRÁN Ca 2 3 Na ATP ADP 2 H IC ~100 nm citoszol kötött Ca 2 CR CSQ SERCA
Intracelluláris ion homeosztázis I.II. Február 15, 2011 Ca 2 csatorna 1 Ca 2 1 Ca 2 EC ~2 mm PLAZMA Na /Ca 2 cserélő Ca 2 ATPáz MEMBRÁN Ca 2 3 Na ATP ADP 2 H IC ~100 nm citoszol kötött Ca 2 CR CSQ SERCA
IONCSATORNÁK. I. Szelektivitás és kapuzás. III. Szabályozás enzimek és alegységek által. IV. Akciós potenciál és szinaptikus átvitel
 IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
IONCSATORNÁK I. Szelektivitás és kapuzás II. Struktúra és funkció III. Szabályozás enzimek és alegységek által IV. Akciós potenciál és szinaptikus átvitel V. Ioncsatornák és betegségek VI. Ioncsatornák
Az agyi értónust befolyásoló tényezők
 2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
2016. október 13. Az agyi értónust befolyásoló tényezők Vazoaktív metabolitok EC neurotranszmitterek SIMAIZOM ENDOTHELIUM LUMEN Kereszthíd aktiváció a simaizomban Ca 2+ -által stimulált myosin foszforiláció
In vitro elektrofiziológiai technikák Mike Árpád
 In vitro elektrofiziológiai technikák Mike Árpád 2011-05-20 1. A sejt szintű elektrofiziológia alapjai: Története Technikák Ionáramok szelektivitása, iránya, nagysága, hatása a membránpotenciálra 2. FAQ
In vitro elektrofiziológiai technikák Mike Árpád 2011-05-20 1. A sejt szintű elektrofiziológia alapjai: Története Technikák Ionáramok szelektivitása, iránya, nagysága, hatása a membránpotenciálra 2. FAQ
Érzékszervi receptorok
 Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
Érzékszervi receptorok működése Akciós potenciál Érzékszervi receptorok Az akciós potenciál fázisai Az egyes fázisokat kísérő ionáram változások 214.11.12. Érzékszervi receptorok Speciális sejtek a környezetből
2012.11.27. Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai. Neuronok izolálása I
 Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai Sejtszintű elektrofiziológia 1.: csatornák funkcionális Sejtszintű elektrofiziológia 2.: izolált/sejtkultúrában
Neuronok előkészítése funkcionális vizsgálatokra. Az alkalmazható technikák előnyei és hátrányai Sejtszintű elektrofiziológia 1.: csatornák funkcionális Sejtszintű elektrofiziológia 2.: izolált/sejtkultúrában
Glia - neuron interakció
 Glia - neuron interakció 2016.04.27. kornyei@koki.hu Neuronális heterogenitás: Cajal rajzai alapján Gliasejtek morfológiai diverzitása Gustaf Retzius hgfap-gfp Emsley 2006 Gliális sejttípusok az idegrendszerben
Glia - neuron interakció 2016.04.27. kornyei@koki.hu Neuronális heterogenitás: Cajal rajzai alapján Gliasejtek morfológiai diverzitása Gustaf Retzius hgfap-gfp Emsley 2006 Gliális sejttípusok az idegrendszerben
IONCSATORNÁK. Osztályozás töltéshordozók szerint: pozitív töltésű ion: Na+, K+, Ca2+ negatív töltésű ion: Cl-, HCO3-
 Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
Ionáromok IONCSATORNÁK 1. Osztályozás töltéshordozók szerint: 1. pozitív töltésű ion: Na+, K+, Ca2+ 2. negatív töltésű ion: Cl-, HCO3-3. Non-specifikus kationcsatornák: h áram 4. Non-specifikus anioncsatornák
2006 1. Nemszinaptikus receptorok és szubmikronos Ca2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra.
 2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
2006 1. Nemszinaptikus receptorok és szubmikronos Ca 2+ válaszok: A két-foton lézermikroszkópia felhasználása a farmakológiai vizsgálatokra. A kutatócsoportunkban Közép Európában elsőként bevezetett két-foton
Szinapszis, szinaptogenezis
 Szinapszis, szinaptogenezis en passant és terminális szinapszisok parakrin szinapszis Kémiai szinapszis Jena, B.J., J. Cell. Mol. Med. Vol 8, No 1, 2004 SNARE proteins ("SNAP and NSF a>achment receptors")
Szinapszis, szinaptogenezis en passant és terminális szinapszisok parakrin szinapszis Kémiai szinapszis Jena, B.J., J. Cell. Mol. Med. Vol 8, No 1, 2004 SNARE proteins ("SNAP and NSF a>achment receptors")
IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója
 IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója A Golgi-impregnáció kulcsfontosságú módszer a struktúra megismerésében rer: tigroid vs Nissl rögök Tigroid: Lenhossék
IDEGSZÖVET 1. neuronok felépítése, típusai, végszervei 2. gliasejtek típusai és funkciója A Golgi-impregnáció kulcsfontosságú módszer a struktúra megismerésében rer: tigroid vs Nissl rögök Tigroid: Lenhossék
Sejt - kölcsönhatások az idegrendszerben
 Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Sejt - kölcsönhatások az idegrendszerben dendrit Sejttest Axon sejtmag Axon domb Schwann sejt Ranvier mielinhüvely csomó (befűződés) terminális Sejt - kölcsönhatások az idegrendszerben Szinapszis típusok
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Interneurális kommunikáció
 Interneurális kommunikáció 2010/2011 Sejtélettan II. Szinapszisok osztályozása Na channel Transmitter vesicle Local circuit current Na 2+ Ca channel PRE- SYNAPTIC Ca++ PRE- SYNAPTIC Ca-induced exocytosis
Interneurális kommunikáció 2010/2011 Sejtélettan II. Szinapszisok osztályozása Na channel Transmitter vesicle Local circuit current Na 2+ Ca channel PRE- SYNAPTIC Ca++ PRE- SYNAPTIC Ca-induced exocytosis
A sejtek közötti kommunikáció módjai és mechanizmusa. kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok
 A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
A sejtek közötti kommunikáció módjai és mechanizmusa kommunikáció a szomszédos vagy a távoli sejtek között intracellulári jelátviteli folyamatok A kommunikáció módjai szomszédos sejtek esetén autokrin
A somatomotoros rendszer
 A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
A somatomotoros rendszer Motoneuron 1 Neuromuscularis junctio (NMJ) Vázizom A somatomotoros rendszer 1 Neurotranszmitter: Acetil-kolin Mire hat: Nikotinos kolinerg-receptor (nachr) Izom altípus A parasympathicus
AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN
 AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN Kormann Eszter Idegi sejtdifferenciáció 2. 2012.12.10. AZ EPILEPSZIÁRÓL RÖVIDEN Definíció: az agyban kialakuló betegség, melyet legalább két alkalommal
AZ ASZTROCITA DISZFUNKCIÓ SZEREPE AZ EPILEPSZIÁBAN Kormann Eszter Idegi sejtdifferenciáció 2. 2012.12.10. AZ EPILEPSZIÁRÓL RÖVIDEN Definíció: az agyban kialakuló betegség, melyet legalább két alkalommal
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András
 Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Elektrofiziológiai alapjelenségek 1. Dr. Tóth András Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál Ioncsatornák alaptulajdonságai Nehézségi fok Belépı szint (6 év alatt is) Hallgató
Membránpotenciál, akciós potenciál
 A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
A nyugalmi membránpotenciál Membránpotenciál, akciós potenciál Fizika-Biofizika 2015.november 3. Nyugalomban valamennyi sejt belseje negatív a külső felszínhez képest: negatív nyugalmi potenciál (Em: -30
A nemi különbségek vizsgálatáról lévén szó, elsődleges volt a nemi hormonok, mint belső környezetbeli különbségeket létrehozó tényezők szerepének
 Kutatási beszámoló Pályázatunk célja annak kiderítése volt, hogy az agyi asztrociták mutatnak-e nemi különbségeket, akár struktura, akár területi megoszlás, akár reaktivitás tekintetében. Alkalmazott megközelítésünk
Kutatási beszámoló Pályázatunk célja annak kiderítése volt, hogy az agyi asztrociták mutatnak-e nemi különbségeket, akár struktura, akár területi megoszlás, akár reaktivitás tekintetében. Alkalmazott megközelítésünk
Az idegrendszer határfelszínei és a neurovaszkuláris egység
 Az idegrendszer határfelszínei és a neurovaszkuláris egység Határfelszínek az idegrendszerben vér-agy gát [blood-brain barrier (BBB)] vér-liquor gát [bloodcerebrospinal fluid barrier (BCSFB)] arachnoid
Az idegrendszer határfelszínei és a neurovaszkuláris egység Határfelszínek az idegrendszerben vér-agy gát [blood-brain barrier (BBB)] vér-liquor gát [bloodcerebrospinal fluid barrier (BCSFB)] arachnoid
Receptorok, szignáltranszdukció jelátviteli mechanizmusok
 Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Receptorok, szignáltranszdukció jelátviteli mechanizmusok Sántha Péter 2016.09.16. A sejtfunkciók szabályozása - bevezetés A sejtek közötti kommunikáció fő típusai: Endokrin Parakrin - Autokrin Szinaptikus
Sejtek közötti kommunikáció
 Sejtek közötti kommunikáció Szerv/szövet homeosztázisa szempontjából fontos: A sejt érzékeli a változásokat környezetében és arra megfelelő választ ad. Többsejtűekben a szignál molekulák koordinálják a
Sejtek közötti kommunikáció Szerv/szövet homeosztázisa szempontjából fontos: A sejt érzékeli a változásokat környezetében és arra megfelelő választ ad. Többsejtűekben a szignál molekulák koordinálják a
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen
 Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
Az orvosi biotechnológiai mesterképzés megfeleltetése az Európai Unió új társadalmi kihívásainak a Pécsi Tudományegyetemen és a Debreceni Egyetemen Azonosító szám: TÁMOP-4.1.2-08/1/A-2009-0011 Az orvosi
- Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál. ellenállás. kondenzátor
 - Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál ellenállás kondenzátor Alap struktúra: 4x6TM S5-S6: vizes pórus, szelektivitás S1-S4: feszültség-függés gating: általában
- Csatornák pumpák - Ellenállás kondenzátor komponens - Fordulási-, membrán potenciál ellenállás kondenzátor Alap struktúra: 4x6TM S5-S6: vizes pórus, szelektivitás S1-S4: feszültség-függés gating: általában
Celluláris és Molekuláris Neurobiológia 2016
 Az asztro- és mikrogliasejtek biológiája Celluláris és Molekuláris Neurobiológia 2016 kornyei@koki.hu glia glia (görög): ragadós A GLIA felfedezése 1858, Rudolf Ludwig Karl Virchow alkotta meg a passzív
Az asztro- és mikrogliasejtek biológiája Celluláris és Molekuláris Neurobiológia 2016 kornyei@koki.hu glia glia (görög): ragadós A GLIA felfedezése 1858, Rudolf Ludwig Karl Virchow alkotta meg a passzív
A plazmamembrán felépítése
 A plazmamembrán felépítése Folyékony mozaik membrán Singer-Nicholson (1972) Lipid kettősréteg Elektronmikroszkópia Membrán kettősréteg Intracelluláris Extracelluláris 1 Lipid kettősréteg foszfolipidek
A plazmamembrán felépítése Folyékony mozaik membrán Singer-Nicholson (1972) Lipid kettősréteg Elektronmikroszkópia Membrán kettősréteg Intracelluláris Extracelluláris 1 Lipid kettősréteg foszfolipidek
CzB 2010. Élettan: a sejt
 CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
CzB 2010. Élettan: a sejt Sejt - az élet alapvető egysége Prokaryota -egysejtű -nincs sejtmag -nincsenek sejtszervecskék -DNS = egy gyűrű - pl., bactériumok Eukaryota -egy-/többsejtű -sejmag membránnal
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák
 A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
A szívizom akciós potenciálja, és az azt meghatározó ioncsatornák Dr. Jost Norbert SZTE, ÁOK Farmakológiai és Farmakoterápiai Intézet Az ingerület vezetése a szívben Conduction velocity in m/s Time to
A szívizomsejt ioncsatornái és azok működése
 A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
A szívizomsejt ioncsatornái és azok működése Dr. Bárándi László Viktor Passzív transzport Egyszerű diffúzió: H 2 O, O 2, CO 2, lipid oldékony anyagok, ionok Csatornán át történő diffúzió: Permeabilitás:
Idegrendszer egyedfejlődése. Az idegszövet jellemzése
 Idegrendszer egyedfejlődése. Az idegszövet jellemzése Központi idegrendszer egyedfejlődése: Ektoderma dorsális részéből velőcső Velőcső középső és hátsó részéből: gerincvelő Velőcső elülső részéből 3 agyhólyag:
Idegrendszer egyedfejlődése. Az idegszövet jellemzése Központi idegrendszer egyedfejlődése: Ektoderma dorsális részéből velőcső Velőcső középső és hátsó részéből: gerincvelő Velőcső elülső részéből 3 agyhólyag:
Sejtek közötti kommunikáció
 Sejtek közötti kommunikáció Szerv/szövet homeosztázisa szempontjából fontos: A sejt érzékeli a változásokat környezetében és arra megfelelő választ ad. Többsejtűekben a szignál molekulák koordinálják a
Sejtek közötti kommunikáció Szerv/szövet homeosztázisa szempontjából fontos: A sejt érzékeli a változásokat környezetében és arra megfelelő választ ad. Többsejtűekben a szignál molekulák koordinálják a
Membrántranszport. Gyógyszerész előadás Dr. Barkó Szilvia
 Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
Membrántranszport Gyógyszerész előadás 2017.04.10 Dr. Barkó Szilvia Sejt membránok A sejtmembrán funkciói Védelem Kommunikáció Molekulák importja és exportja Sejtmozgás Általános szerkezet Lipid kettősréteg
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák. Nem vezikuláris release
 Glia fiziológia I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz
Glia fiziológia I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz
Az akciós potenciál (AP) 2.rész. Szentandrássy Norbert
 2.rész. Szentandrássy Norbert") Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Az akciós potenciál (AP) 2.rész Szentandrássy Norbert Ismétlés Az akciós potenciált küszöböt meghaladó nagyságú depolarizáció váltja ki Mert a feszültségvezérelt Na + -csatornákat a depolarizáció aktiválja,
Membránszerkezet Nyugalmi membránpotenciál
 Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Membránszerkezet Nyugalmi membránpotenciál 2011.11.15. A biológiai membránok fő komponense. Foszfolipidek foszfolipid = diglicerid + foszfát csoport + szerves molekula (pl. kolin). Poláros fej (hidrofil)
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.
. Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet.") Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
Termodinamikai egyensúlyi potenciál (Nernst, Donnan). Diffúziós potenciál, Goldman-Hodgkin-Katz egyenlet. Biológiai membránok passzív elektromos tulajdonságai. A sejtmembrán kondenzátorként viselkedik
NÖVÉNYGENETIKA. Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP /1/A
 NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
NÖVÉNYGENETIKA Az Agrármérnöki MSc szak tananyagfejlesztése TÁMOP-4.1.2-08/1/A-2009-0010 A NÖVÉNYI TÁPANYAG TRANSZPORTEREK az előadás áttekintése A tápionok útja a növényben Növényi tápionok passzív és
Szívelektrofiziológiai alapjelenségek. Dr. Tóth András 2018
 Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
Szívelektrofiziológiai alapjelenségek 1. Dr. Tóth András 2018 Témák Membrántranszport folyamatok Donnan egyensúly Nyugalmi potenciál 1 Transzmembrán transzport A membrántranszport-folyamatok típusai J:
Intracelluláris és intercelluláris kommunikáció
 Intracelluláris és intercelluláris kommunikáció Transzportfolyamatok a sejten belül Ciklózis: Az endoplazma sejten belüli (sejtmag körüli) áramlása A ciklózis teszi lehetővé, hogy a sejten belül az egyik
Intracelluláris és intercelluláris kommunikáció Transzportfolyamatok a sejten belül Ciklózis: Az endoplazma sejten belüli (sejtmag körüli) áramlása A ciklózis teszi lehetővé, hogy a sejten belül az egyik
Membránpotenciál. Nyugalmi membránpotenciál. Akciós potenciál
 Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Membránpotenciál Vig Andrea 2014.10.29. Nyugalmi membránpotenciál http://quizlet.com/8062024/ap-11-nervous-system-part-5-electrical-flash-cards/ Akciós potenciál http://cognitiveconsonance.info/2013/03/21/neuroscience-the-action-potential/
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt
 az információ kémiailag kódolt") Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
Szignáltranszdukció Mediátorok (elsődleges hírvivők) az információ kémiailag kódolt apoláros szerkezet (szabad membrán átjárhatóság) szteroid hormonok, PM hormonok, retinoidok hatásmech.: sejten belül
Receptorok és szignalizációs mechanizmusok
 Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Molekuláris sejtbiológia: Receptorok és szignalizációs mechanizmusok Dr. habil Kőhidai László Semmelweis Egyetem Genetikai, Sejt- és Immunbiológiai Intézet Sejtek szignalizációs kapcsolatai Sejtek szignalizációs
Neurovaszkuláris csatolás
 Neurovaszkuláris csatolás Farkas Eszter 2016. október 20. Az első bizonyíték a neurovaszkuláris csatolásra Kognitív feladat végzése az agytérfogat változásával jár (Mosso, 1881) A Roy-Sherrington elv Neurovaszkuláris
Neurovaszkuláris csatolás Farkas Eszter 2016. október 20. Az első bizonyíték a neurovaszkuláris csatolásra Kognitív feladat végzése az agytérfogat változásával jár (Mosso, 1881) A Roy-Sherrington elv Neurovaszkuláris
BIOFIZIKA. Membránpotenciál és transzport. Liliom Károly. MTA TTK Enzimológiai Intézet
 BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
BIOFIZIKA 2012 10 15 Membránpotenciál és transzport Liliom Károly MTA TTK Enzimológiai Intézet liliom@enzim.hu A biofizika előadások temamkája 1. 09-03 Biofizika: fizikai szemlélet, modellalkotás, biometria
Sejt - kölcsönhatások. az idegrendszerben és az immunrendszerben
 Sejt - kölcsönhatások az idegrendszerben és az immunrendszerben A sejttől a szervezetig A sejtek között, ill. a sejtek és környezetük közötti jelátviteli folyamatok összessége az a struktúrált kölcsönhatásrendszer,
Sejt - kölcsönhatások az idegrendszerben és az immunrendszerben A sejttől a szervezetig A sejtek között, ill. a sejtek és környezetük közötti jelátviteli folyamatok összessége az a struktúrált kölcsönhatásrendszer,
Szignalizáció - jelátvitel
 Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Jelátvitel autokrin Szignalizáció - jelátvitel Összegezve: - a sejt a,,külvilággal"- távolabbi szövetekkel ill. önmagával állandó anyag-, információ-, energia áramlásban áll, mely autokrin, parakrin,
Glutamát transzporterek asztrocitákban. EAA- Excitatory Amino Acid Transporter (EAAT) család. főleg asztroglia, de bizonyos neuronokon is
 család. főleg asztroglia, de bizonyos neuronokon is") Glia fiziológia Gliális neurotranszmitter transzporterek Glutamát transzporterek asztrocitákban EAA- Excitatory Amino Acid Transporter (EAAT) család GLAST (EAAT1) GLT1 (EAAT2) EAAC1 (EAAT3) EAAT4 EAAT5
Glia fiziológia Gliális neurotranszmitter transzporterek Glutamát transzporterek asztrocitákban EAA- Excitatory Amino Acid Transporter (EAAT) család GLAST (EAAT1) GLT1 (EAAT2) EAAC1 (EAAT3) EAAT4 EAAT5
Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György
 Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György Nyugalmi membránpotenciál: TK. 284-285. Akciós potenciál: TK. 294-301. Elektromos ingerelhetőség:
Nyugalmi potenciál, akciós potenciál és elektromos ingerelhetőség. A membránpotenciál mérése. Panyi György Nyugalmi membránpotenciál: TK. 284-285. Akciós potenciál: TK. 294-301. Elektromos ingerelhetőség:
Apoptózis. 1. Bevezetés 2. Külső jelút 3. Belső jelút
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag 1. Kondenzálódó sejtmag apoptózis autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita
Az fmri alapjai BOLD fiziológia. Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika
 Az fmri alapjai BOLD fiziológia Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika T2* Az obszervált transzverzális relaxáció (T2*) több különböző komponens összege Many physical effects result
Az fmri alapjai BOLD fiziológia Dr. Kincses Tamás Szegedi Tudományegyetem Neurológiai Klinika T2* Az obszervált transzverzális relaxáció (T2*) több különböző komponens összege Many physical effects result
2. A jelutak komponensei. 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék
 Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak 2. A jelutak komponensei 1. Egy tipikus jelösvény sémája 2. Ligandok 3. Receptorok 4. Intracelluláris jelfehérjék Egy tipikus jelösvény sémája 1. Receptor fehérje Jel molekula (ligand; elsődleges
Jelutak. Apoptózis. Apoptózis Bevezetés 2. Külső jelút 3. Belső jelút. apoptózis autofágia nekrózis. Sejtmag. Kondenzálódó sejtmag
 Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Jelutak Apoptózis 1. Bevezetés 2. Külső jelút 3. Belső jelút Apoptózis Sejtmag Kondenzálódó sejtmag 1. autofágia nekrózis Lefűződések Összezsugorodás Fragmentálódó sejtmag Apoptotikus test Fagocita bekebelezi
Szinaptikus folyamatok
 Szinaptikus folyamatok Jelátvitel az idegrendszerben Elektromos szinapszisok Kémiai szinapszisok Neurotranszmitterek és receptoraik Szinaptikus integráció Szinaptikus plaszticitás Kettős információátvitel
Szinaptikus folyamatok Jelátvitel az idegrendszerben Elektromos szinapszisok Kémiai szinapszisok Neurotranszmitterek és receptoraik Szinaptikus integráció Szinaptikus plaszticitás Kettős információátvitel
A citokin egyensúly. Gyulladásgátló cytokinek. Gyulladáskeltő citokinek. Védelem és sejttúlélés. Gyulladás, sejtpusztulás NA DA.
 Apoptózis; Asztrocita: fagocitózis; PGD4; Növekedési faktorok: TGFβ1; neurosteroidok Gyulladásgátló cytokinek A citokin egyensúly NEUROTRANSZMITTEREK Reaktív asztroglia, aktivált mikroglia; Perifériás
Apoptózis; Asztrocita: fagocitózis; PGD4; Növekedési faktorok: TGFβ1; neurosteroidok Gyulladásgátló cytokinek A citokin egyensúly NEUROTRANSZMITTEREK Reaktív asztroglia, aktivált mikroglia; Perifériás
Glia fiziológia I. Gliotranszmisszió. Gliotranszmitterek. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák. Nem vezikuláris release
 Glia fiziológia I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz
Glia fiziológia I. Csatornák, receptorok Kapcsoltság, Ca ++ Ioncsatornák Kapcsoltság Aquaporinok Glia szincícium Gliotranszmisszió Gliotranszmitterek Nem vezikuláris release Vezikuláris release Neurotransz
Sejtek membránpotenciálja
 Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Sejtek membránpotenciálja Termodinamikai egyensúlyi potenciál (Nernst, Donnan) Diffúziós potenciál, (Goldman-Hodgkin-Katz egyenlet) A nyugalmi membránpotenciál: TK. 284-285. A nyugalmi membránpotenciál
Transzportfolyamatok a biológiai rendszerekben
 A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
A nyugalmi potenciál jelentősége Transzportfolyamatok a biológiai rendszerekben Transzportfolyamatok a sejt nyugalmi állapotában a sejt homeosztázisának (sejttérfogat, ph) fenntartása ingerlékenység érzékelés
Degeneráció és regeneráció az idegrendszerben
 Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
Degeneráció és regeneráció az idegrendszerben Axonális sérülés és regeneráció August Waller, 1850: Waller-féle degeneráció (Wallerian degeneration) disztális axonális csonk: degeneráció 24-36 órán belül
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül.
 7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
7. előadás: A plazma mebrán szerkezete és funkciója. Anyagtranszport a plazma membránon keresztül. A plazma membrán határolja el az élő sejteket a környezetüktől Szelektív permeabilitást mutat, így lehetővé
Glia-neuron interakció aspektusai. Neuron-glia metabolikus együttműködés
 Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
Glia-neuron interakció aspektusai Agyi mikrokörnyezet szabályozása Neuron-glia metabolikus együttműködés Neurovaszkuláris kapcsoltság ion, víz homeosztázis szabályozása neurotranszmitter homeosztázis szabályozása
A szív élettana. Aszív élettana I. A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG
 A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa
A szív élettana A szív pumpafunkciója A szívciklus A szívizom sajátosságai A szív elektrofiziológiája Az EKG prof. Sáry Gyula 1 Aszív élettana I. A szívizom sajátosságai A szívciklus A szív mint pumpa